
Lipidomic Profile of Human Sperm Membrane Identifies a Clustering of Lipids Associated with Semen Quality and Function


 ,
,  ,
,  ,
, 
 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
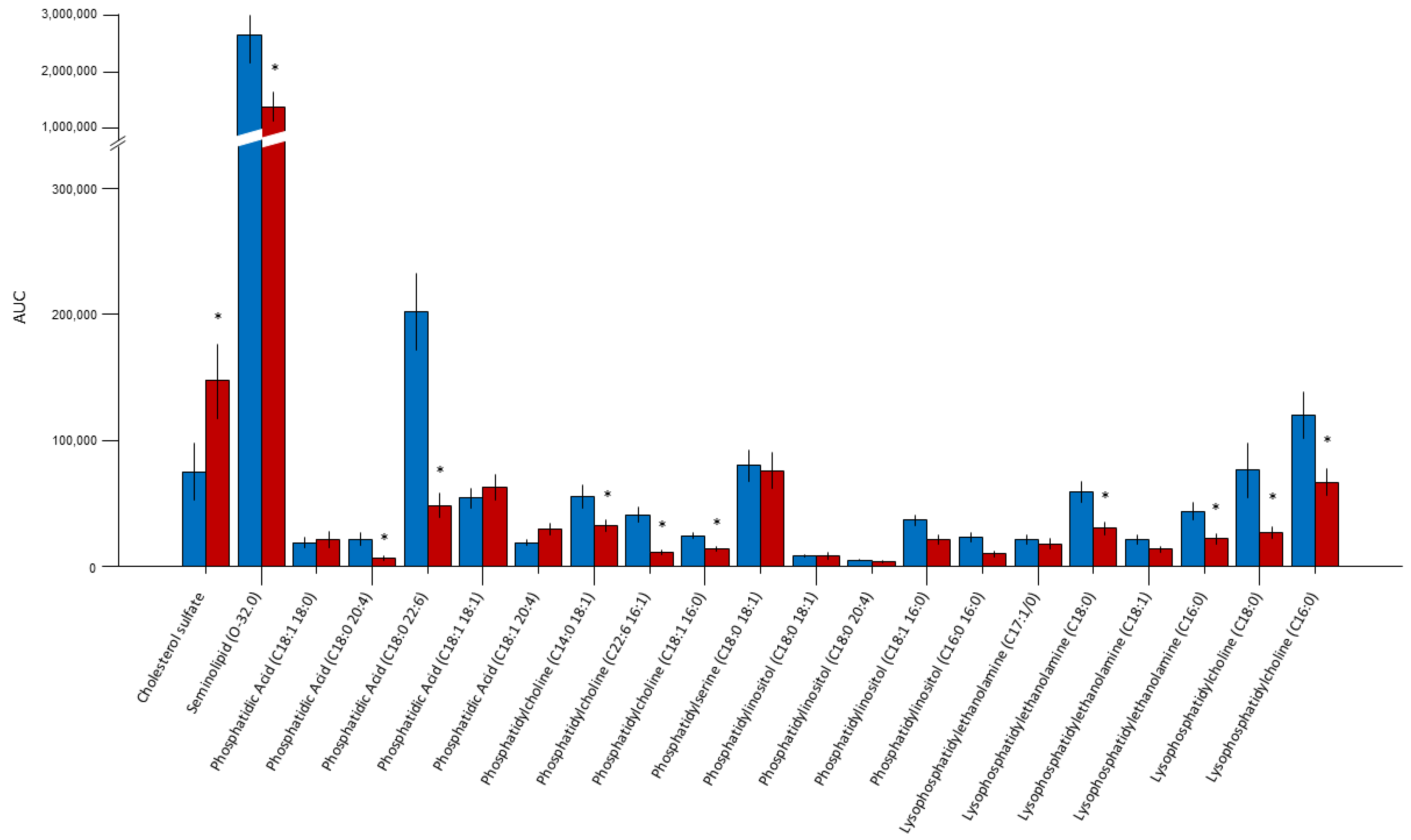
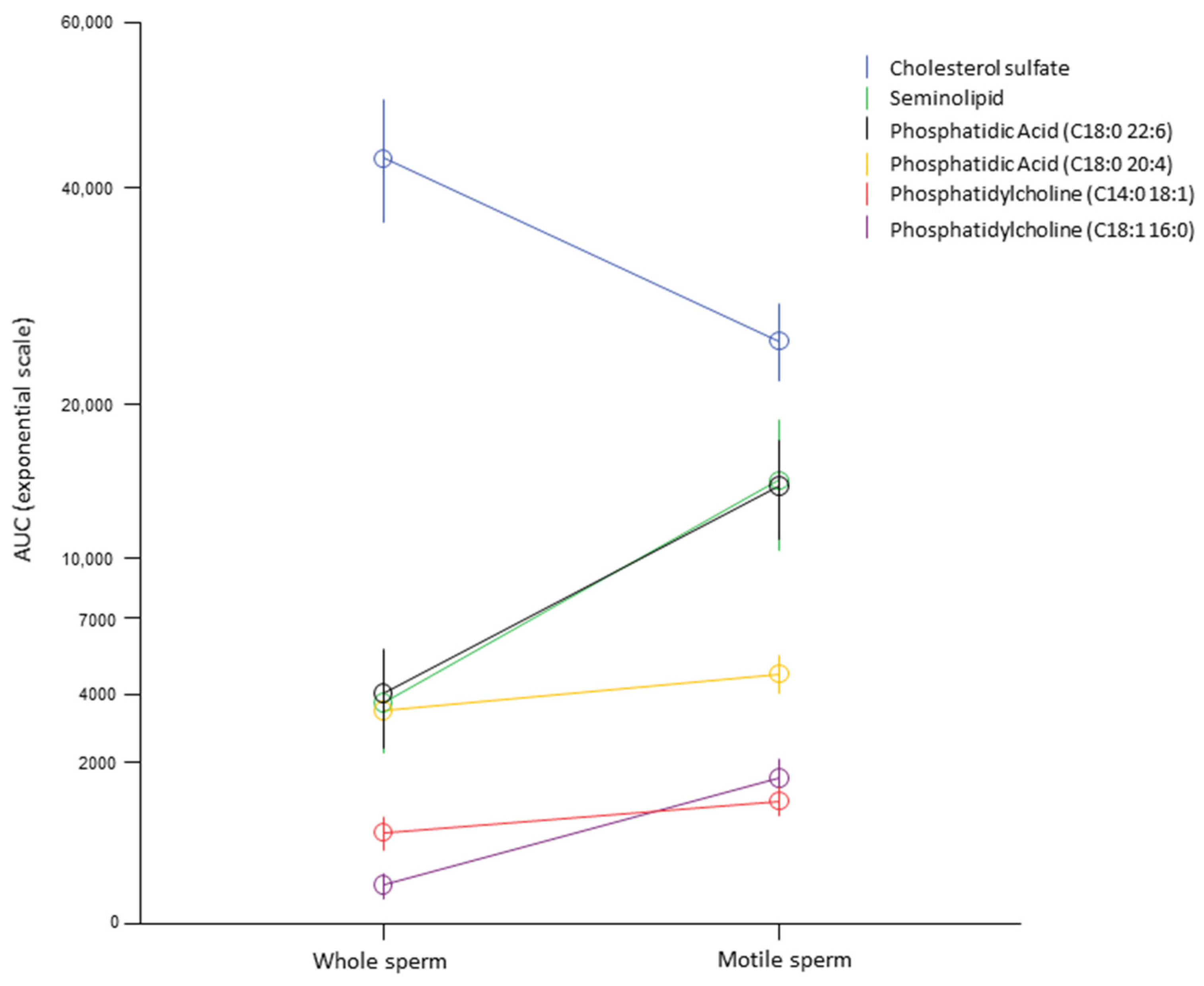
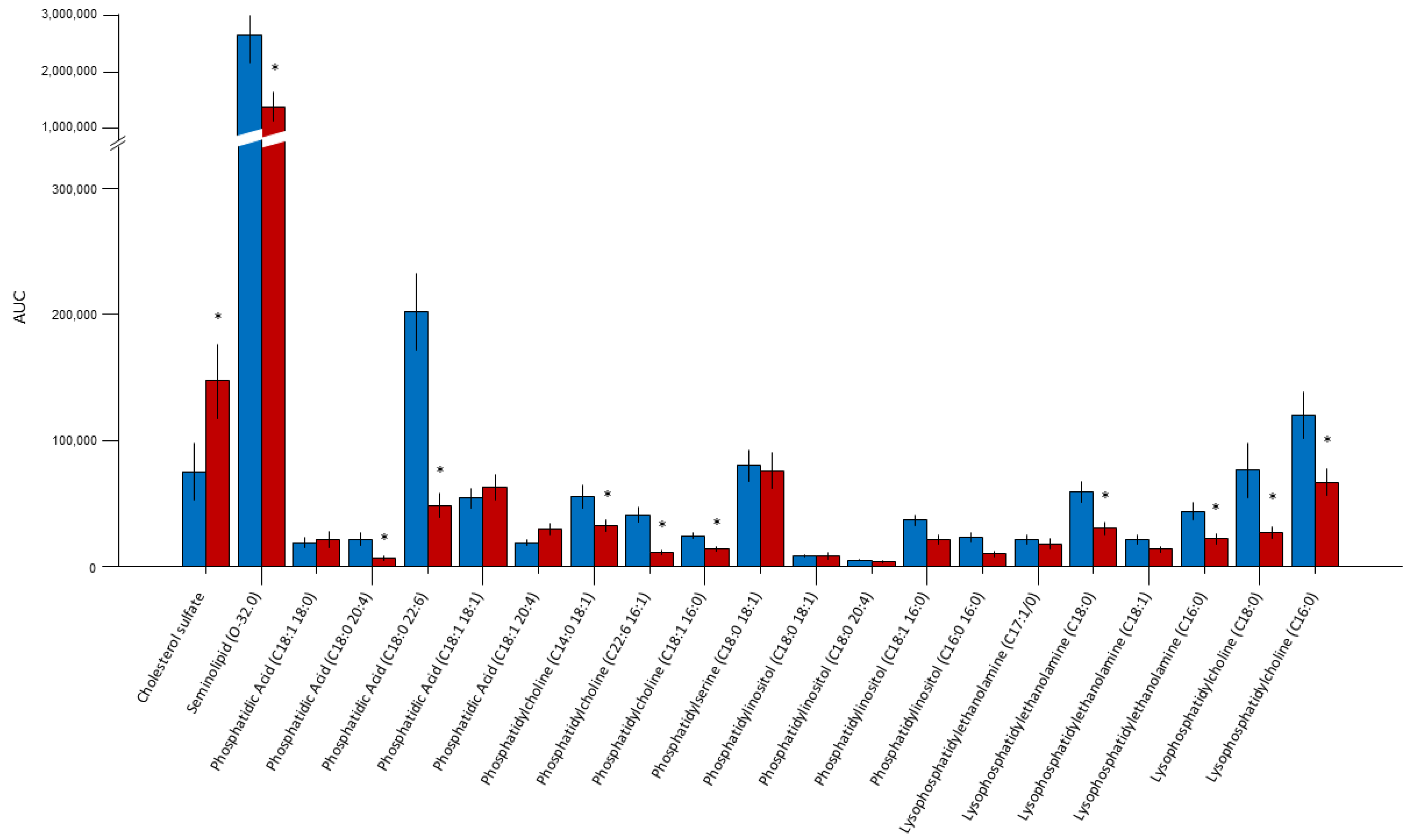
2.1. Lipidomic Analysis
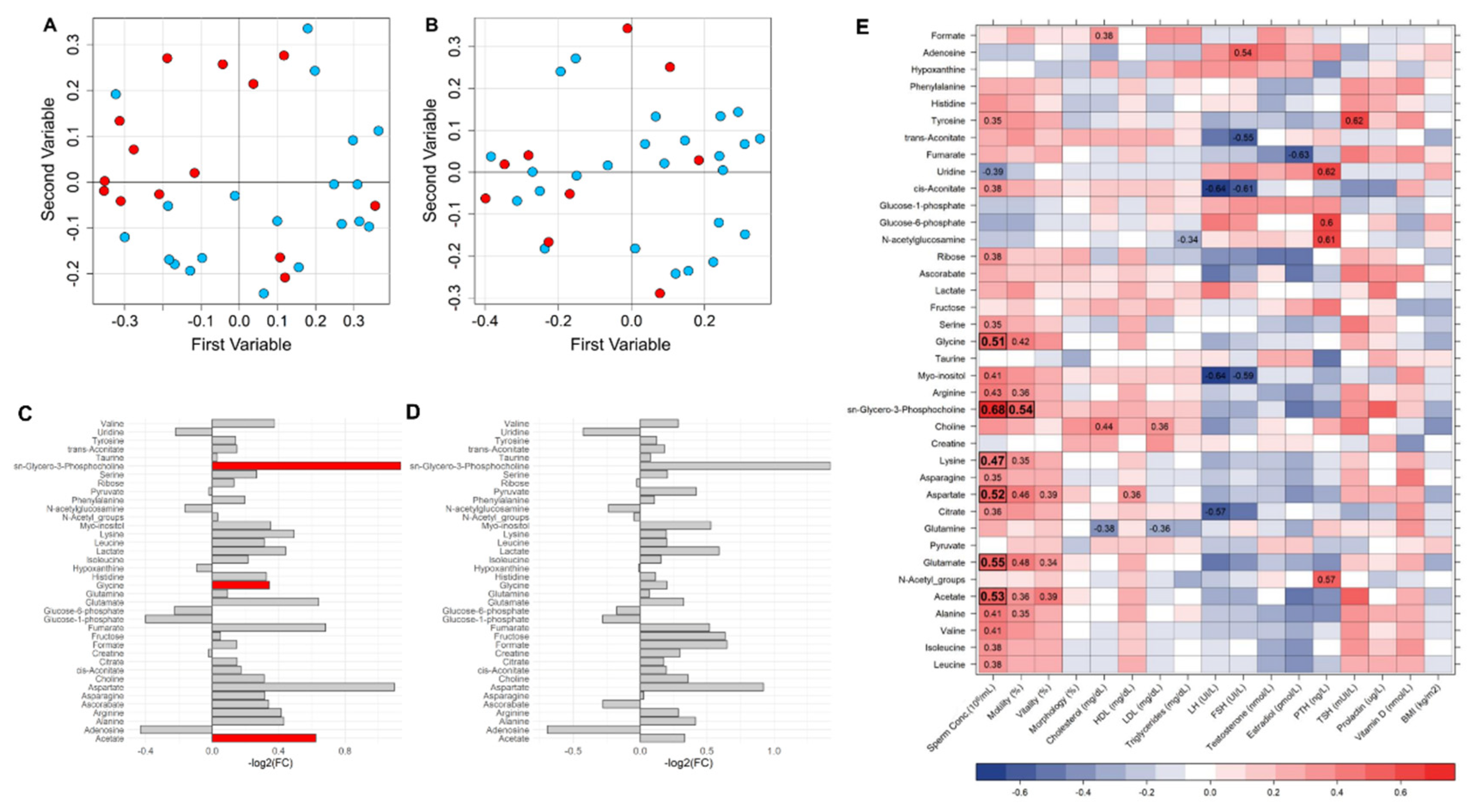
2.2. Metabolomic Analysis
3. Discussion
4. Materials and Methods
4.1. Subjects and Semen Analysis
4.2. Swim-Up
4.3. Sperm Cell Isolation by Density Gradient
4.4. Extraction and Quantitative Determination of Lipids by LC-MS
4.5. NMR Analysis
4.6. Sex Hormone Quantification
4.7. Biochemical Serum Markers Evaluation
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nikolopoulou, M.; Soucek, D.A.; Vary, J.C. Changes in the Lipid Content of Boar Sperm Plasma Membranes during Epididymal Maturation. Biochim. Biophys. Acta 1985, 815, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Zerbinati, C.; Caponecchia, L.; Rago, R.; Leoncini, E.; Bottaccioli, A.G.; Ciacciarelli, M.; Pacelli, A.; Salacone, P.; Sebastianelli, A.; Pastore, A.; et al. Fatty Acids Profiling Reveals Potential Candidate Markers of Semen Quality. Andrology 2016, 4, 1094–1101. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. The Amoroso Lecture. The Human Spermatozoon—A Cell in Crisis? J. Reprod. Fertil. 1999, 115, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lopalco, P.; Vitale, R.; Cho, Y.S.; Totaro, P.; Corcelli, A.; Lobasso, S. Alteration of Cholesterol Sulfate/Seminolipid Ratio in Semen Lipid Profile of Men With Oligoasthenozoospermia. Front. Physiol. 2019, 10, 1344. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Kassan, A.; Busija, A.R.; Rangamani, P.; Patel, H.H. The Plasma Membrane as a Capacitor for Energy and Metabolism. Am. J. Physiol. Cell Physiol. 2016, 310, C181–C192. [Google Scholar] [CrossRef] [PubMed]
- Potter, J.E.R.; James, M.J.; Kandutsch, A.A. Sequential Cycles of Cholesterol and Dolichol Synthesis in Mouse Spleens during Phenylhydrazine-Induced Erythropoiesis. J. Biol. Chem. 1981, 256, 2371–2376. [Google Scholar] [CrossRef] [PubMed]
- Yanagimachi, R. Fertility of Mammalian Spermatozoa: Its Development and Relativity. Zygote 1994, 2, 371–372. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Shiratsuchi, A. Phagocytic Removal of Apoptotic Spermatogenic Cells by Sertoli Cells: Mechanisms and Consequences. Biol. Pharm. Bull. 2004, 27, 13–16. [Google Scholar] [CrossRef]
- Visconti, P.E.; Ning, X.P.; Fornés, M.W.; Alvarez, J.G.; Stein, P.; Connors, S.A.; Kopf, G.S. Cholesterol Efflux-Mediated Signal Transduction in Mammalian Sperm: Cholesterol Release Signals an Increase in Protein Tyrosine Phosphorylation during Mouse Sperm Capacitation. Dev. Biol. 1999, 214, 429–443. [Google Scholar] [CrossRef]
- Needham, D.; Mcintosh, T.J.; Evans, E. Thermomechanical and Transition Properties of Dimyristoylphosphatidylcholine/Cholesterol Bilayers. Biochemistry 1988, 27, 4668–4673. [Google Scholar] [CrossRef]
- Pelletier, R.M. The Blood-Testis Barrier: The Junctional Permeability, the Proteins and the Lipids. Prog. Histochem. Cytochem. 2011, 46, 49–127. [Google Scholar] [CrossRef] [PubMed]
- Garolla, A.; Šabović, I.; Tescari, S.; De Toni, L.; Menegazzo, M.; Cosci, I.; De Filippis, V.; Giarola, M.; Foresta, C. Impaired Sperm Function in Infertile Men Relies on the Membrane Sterol Pattern. Andrology 2018, 6, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Wood, P.L.; Scoggin, K.; Ball, B.A.; Troedsson, M.H.; Squires, E.L. Lipidomics of Equine Sperm and Seminal Plasma: Identification of Amphiphilic (O-Acyl)-ω-Hydroxy-Fatty Acids. Theriogenology 2016, 86, 1212–1221. [Google Scholar] [CrossRef] [PubMed]
- Sandhoff, K.; Kolter, T. Biosynthesis and Degradation of Mammalian Glycosphingolipids. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 847–861. [Google Scholar] [CrossRef]
- Vos, J.P.; Lopes-Cardozo, M.; Gadella, B.M. Metabolic and Functional Aspects of Sulfogalactolipids. Biochim. Biophys. Acta 1994, 1211, 125–149. [Google Scholar] [CrossRef]
- Tanphaichitr, N.; Kongmanas, K.; Faull, K.F.; Whitelegge, J.; Compostella, F.; Goto-Inoue, N.; Linton, J.J.; Doyle, B.; Oko, R.; Xu, H.; et al. Properties, Metabolism and Roles of Sulfogalactosylglycerolipid in Male Reproduction. Prog. Lipid Res. 2018, 72, 18–41. [Google Scholar] [CrossRef]
- Macías García, B.; González Fernández, L.; Ortega Ferrusola, C.; Salazar-Sandoval, C.; Morillo Rodríguez, A.; Rodríguez Martinez, H.; Tapia, J.A.; Morcuende, D.; Peña, F.J. Membrane Lipids of the Stallion Spermatozoon in Relation to Sperm Quality and Susceptibility to Lipid Peroxidation. Reprod. Domest. Anim. 2011, 46, 141–148. [Google Scholar] [CrossRef]
- Frenette, G.; Girouard, J.; Sullivan, R. Comparison between Epididymosomes Collected in the Intraluminal Compartment of the Bovine Caput and Cauda Epididymidis. Biol. Reprod. 2006, 75, 885–890. [Google Scholar] [CrossRef]
- Kawano, N.; Yoshida, K.; Miyado, K.; Yoshida, M. Lipid Rafts: Keys to Sperm Maturation, Fertilization, and Early Embryogenesis. J. Lipids 2011, 2011, 264706. [Google Scholar] [CrossRef]
- Furse, S.; Kusinski, L.C.; Ray, A.; Glenn-Sansum, C.; Williams, H.E.L.; Koulman, A.; Meek, C.L. Relative Abundance of Lipid Metabolites in Spermatozoa across Three Compartments. Int. J. Mol. Sci. 2022, 23, 11655. [Google Scholar] [CrossRef]
- Lu, J.C.; Jing, J.; Yao, Q.; Fan, K.; Wang, G.H.; Feng, R.X.; Liang, Y.J.; Chen, L.; Ge, Y.F.; Yao, B. Relationship between Lipids Levels of Serum and Seminal Plasma and Semen Parameters in 631 Chinese Subfertile Men. PLoS ONE 2016, 11, e0146304. [Google Scholar] [CrossRef] [PubMed]
- Schisterman, E.F.; Mumford, S.L.; Chen, Z.; Browne, R.W.; Boyd Barr, D.; Kim, S.; Buck Louis, G.M. Lipid Concentrations and Semen Quality: The LIFE Study. Andrology 2014, 2, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Sion, B.; Grizard, G.; Boucher, D. Quantitative Analysis of Desmosterol, Cholesterol and Cholesterol Sulfate in Semen by High-Performance Liquid Chromatography. J. Chromatogr. A 2001, 935, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.A.P.; Ashworth, P.J.C.; Miller, N.G.A. Bicarbonate/CO2, an Effector of Capacitation, Induces a Rapid and Reversible Change in the Lipid Architecture of Boar Sperm Plasma Membranes. Mol. Reprod. Dev. 1996, 45, 378–391. [Google Scholar] [CrossRef]
- Osheroff, J.E.; Visconti, P.E.; Valenzuela, J.P.; Travis, A.J.; Alvarez, J.; Kopf, G.S. Regulation of Human Sperm Capacitation by a Cholesterol Efflux-Stimulated Signal Transduction Pathway Leading to Protein Kinase A-Mediated up-Regulation of Protein Tyrosine Phosphorylation. Mol. Hum. Reprod. 1999, 5, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Sèdes, L.; Thirouard, L.; Maqdasy, S.; Garcia, M.; Caira, F.; Lobaccaro, J.M.A.; Beaudoin, C.; Volle, D.H. Cholesterol: A Gatekeeper of Male Fertility? Front. Endocrinol. 2018, 9, 369. [Google Scholar] [CrossRef]
- Wiebe, J.P.; Tilbe, K.S. De Novo Synthesis of Steroids (from Acetate) by Isolated Rat Sertoli Cells. Biochem. Biophys. Res. Commun. 1979, 89, 1107–1113. [Google Scholar] [CrossRef]
- Maqdasy, S.; Baptissart, M.; Vega, A.; Baron, S.; Lobaccaro, J.M.A.; Volle, D.H. Cholesterol and Male Fertility: What about Orphans and Adopted? Mol. Cell. Endocrinol. 2013, 368, 30–46. [Google Scholar] [CrossRef]
- Eisenberg, M.L.; Kim, S.; Chen, Z.; Sundaram, R.; Schisterman, E.F.; Buck Louis, G.M. The Relationship between Male BMI and Waist Circumference on Semen Quality: Data from the LIFE Study. Hum. Reprod. 2015, 30, 493–494. [Google Scholar] [CrossRef]
- Ladha, S. Lipid Heterogeneity and Membrane Fluidity in a Highly Polarized Cell, the Mammalian Spermatozoon. J. Membr. Biol. 1998, 165, 1–10. [Google Scholar] [CrossRef]
- Collodel, G.; Castellini, C.; Lee, J.C.Y.; Signorini, C. Relevance of Fatty Acids to Sperm Maturation and Quality. Oxid. Med. Cell. Longev. 2020, 2020, 7038124. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Abayasekara, D.R.E.; Aitken, R.J. Polyunsaturated Fatty Acids in Male and Female Reproduction. Biol. Reprod. 2007, 77, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Picardo, M.; Gandini, L.; Dondero, F. Lipids of the Sperm Plasma Membrane: From Polyunsaturated Fatty Acids Considered as Markers of Sperm Function to Possible Scavenger Therapy. Hum. Reprod. Update 1996, 2, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Xu, F.; Hirschfeld, M.; Brenig, B. Sperm Lipid Markers of Male Fertility in Mammals. Int. J. Mol. Sci. 2021, 22, 8767. [Google Scholar] [CrossRef] [PubMed]
- Safarinejad, M.R.; Hosseini, S.Y.; Dadkhah, F.; Asgari, M.A. Relationship of Omega-3 and Omega-6 Fatty Acids with Semen Characteristics, and Anti-Oxidant Status of Seminal Plasma: A Comparison between Fertile and Infertile Men. Clin. Nutr. 2010, 29, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Diao, R.; Zhu, X.; Li, Z.; Cai, Z. Metabolic Characterization of Asthenozoospermia Using Nontargeted Seminal Plasma Metabolomics. Clin. Chim. Acta. 2015, 450, 254–261. [Google Scholar] [CrossRef]
- Tang, B.; Shang, X.; Qi, H.; Li, J.; Ma, B.; An, G.; Zhang, Q. Metabonomic Analysis of Fatty Acids in Seminal Plasma between Healthy and Asthenozoospermic Men Based on Gas Chromatography Mass Spectrometry. Andrologia 2017, 49, e12744. [Google Scholar] [CrossRef]
- Barbonetti, A.; Castellini, C.; Di Giammarco, N.; Santilli, G.; Francavilla, S.; Francavilla, F. In Vitro Exposure of Human Spermatozoa to Bisphenol A Induces Pro-Oxidative/Apoptotic Mitochondrial Dysfunction. Reprod. Toxicol. 2016, 66, 61–67. [Google Scholar] [CrossRef]
- Lenzi, A.; Gandini, L.; Maresca, V.; Rago, R.; Sgrò, P.; Dondero, F.; Picardo, M. Fatty Acid Composition of Spermatozoa and Immature Germ Cells. Mol. Hum. Reprod. 2000, 6, 226–231. [Google Scholar] [CrossRef]
- Vignoli, A.; Tenori, L.; Giusti, B.; Takis, P.G.; Valente, S.; Carrabba, N.; Balzi, D.; Barchielli, A.; Marchionni, N.; Gensini, G.F.; et al. NMR-Based Metabolomics Identifies Patients at High Risk of Death within Two Years after Acute Myocardial Infarction in the AMI-Florence II Cohort. BMC Med. 2019, 17, 3. [Google Scholar] [CrossRef]
- Juyena, N.S.; Stelletta, C. Seminal Plasma: An Essential Attribute to Spermatozoa. J. Androl. 2012, 33, 536–551. [Google Scholar] [CrossRef] [PubMed]
- Rovira, J.; Mari, M.; Nadal, M.; Schuhmacher, M.; Domingo, J.L. Levels of Metals and PCDD/Fs in the Vicinity of a Cement Plant: Assessment of Human Health Risks. J. Environ. Sci. Health-Part A Toxic/Hazard. Subst. Environ. Eng. 2011, 46, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic Quotient Normalization as Robust Method to Account for Dilution of Complex Biological Mixtures. Application in 1H NMR Metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Guo, A.C.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]


{kind=link}
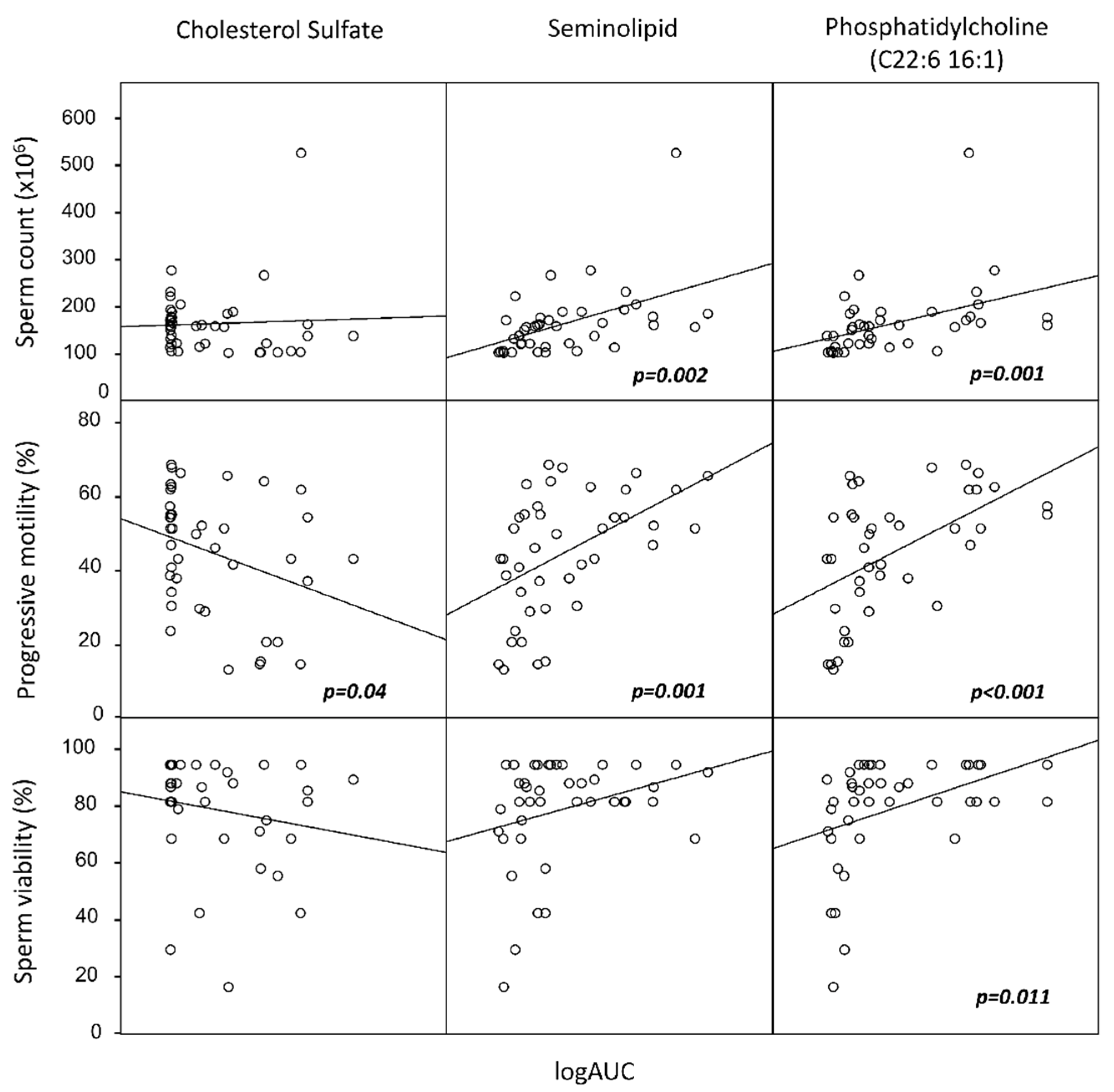{kind=link}
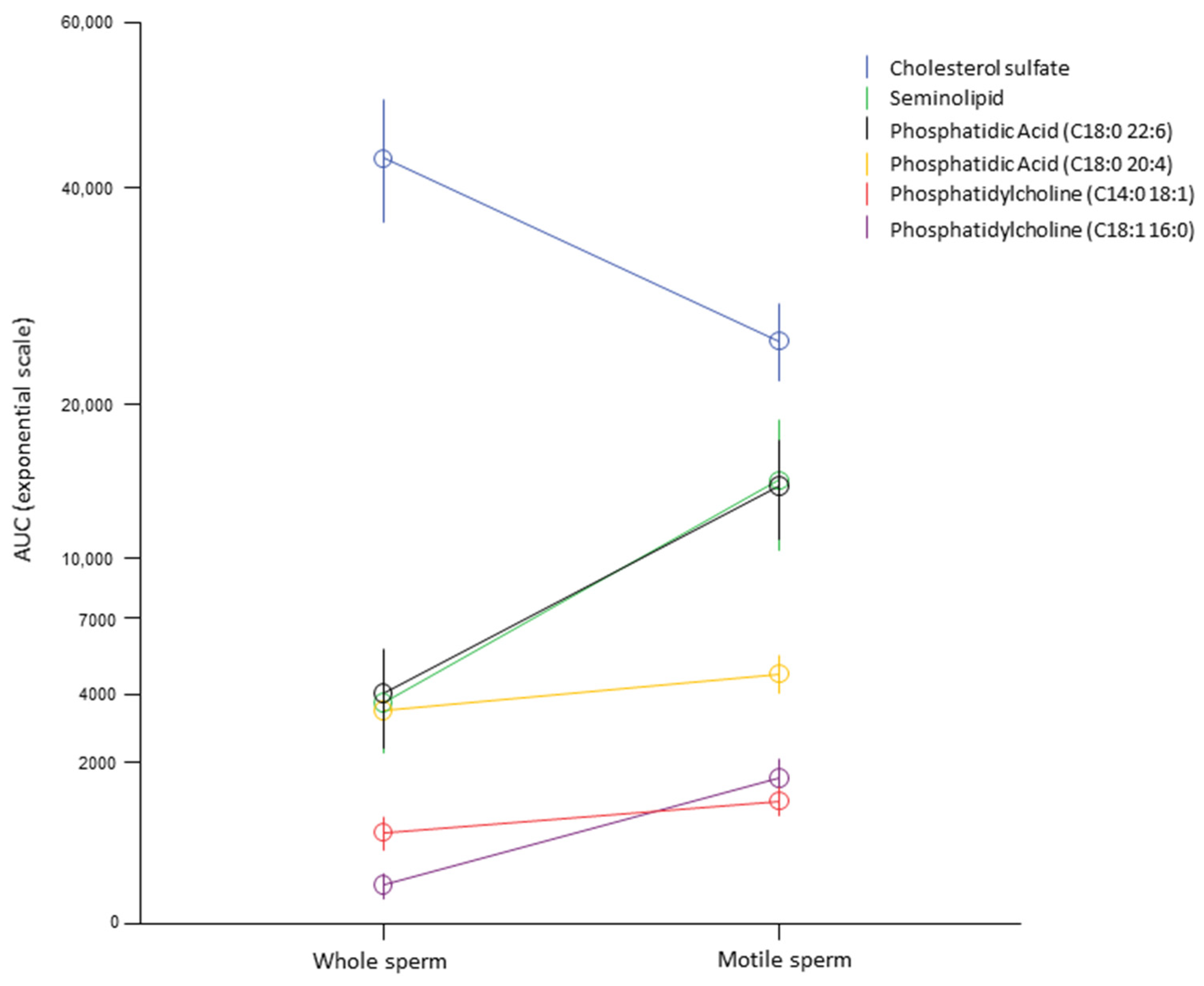{kind=link}
{kind=link}
| Parameters | Fertile Group (N = 33) | Infertile Group (N = 29) | p Value |
|---|---|---|---|
| Age (years) | 31.3 ± 8.3 | 32.8 ± 6.1 | 0.623 |
| BMI (Kg/m2) | 23.6 ± 2.4 | 24.4 ± 3.1 | 0.422 |
| Luteinizing hormone (IU/L) | 4.63 ± 1.65 | 4.52 ± 4.94 | 0.929 |
| Follicle-stimulating hormone (IU/L) | 4.04 ± 1.61 | 5.37 ± 5.41 | 0.320 |
| Estradiol (pmol/L) | 73.90 ± 24.10 | 80.20 ± 34.10 | 0.189 |
| Testosterone (nmol/L) | 19.79 ± 5.79 | 18.45 ± 7.23 | 0.603 |
| Semen volume (mL) | 3.14 ± 1.23 | 1.93 ± 1.03 | <0.001 |
| Total sperm count (106 cells) | 220.06 ± 89.42 | 26.88 ± 24.89 | <0.001 |
| Sperm concentration (106 cells/mL) | 81.89 ± 64.42 | 14.21 ± 10.69 | <0.001 |
| Progressive motility (%) | 51.80 ± 17.98 | 21.22 ± 15.32 | <0.001 |
| Normal morphology (%) | 6.44 ± 3.31 | 5.50 ± 3.83 | 0.376 |
| Viability (%) | 81.92 ± 11.94 | 57.08 ± 24.68 | <0.001 |
| Total cholesterol (mg/dL) | 183.48 ± 47.93 | 192.11 ± 55.09 | 0.529 |
| HDL (mg/dL) | 55.28 ± 10.05 | 48.64 ± 11.45 | 0.612 |
| LDL (mg/dL) | 108.39 ± 41.06 | 105.22 ± 45.84 | 0.817 |
| Triglycerides (mg/dL) | 102.60 ± 82.98 | 131.14 ± 78.60 | 0.178 |
| Lipid | Ion Molecular Mass (Da) | Main Observed Fragment | Retention Time (min) |
|---|---|---|---|
| Cholesterol sulfate | 465 | Fragments | 8.5 |
| Seminolipid (O-32.0) | 795 | 539 315 | 9.6 |
| Phosphatidic acid (C18:1 18:0) | 701 | 419 283 | 13.0 |
| Phosphatidic acid (C18:0 20:4) | 723 | 419 303 283 | 13.4 |
| Phosphatidic acid (C18:0 22:6) | 747 | 419 283 327 | 13.5 |
| Phosphatidic acid (C18:1 18:1) | 700.7 | 463 281 | 15.1 |
| Phosphatidic acid (C18:1 20:4) | 722.8 | 436 303 | 13.4 |
| Phosphatidylcholine (C14:0 18:1) | 716.7 | 452 281 | 13.8 |
| Phosphatidylcholine (C22:6 16:1) | 850 | −60 (790) 533 480 327 | 14.6 |
| Phosphatidylcholine (C18:1 16:0) | 804 | −60 (744) 481 281 | 14.8 |
| Phosphatidylserine (C18:0 18:1) | 788.8 | (−87) 701 419 283 | 15.0 |
| Phosphatidylinositol (C18:0 18:1) | 863 | (−180) 683 581 420 | 17.1 |
| Phosphatidylinositol (C18:0 20:4) | 885 | (−161) 724 600 581 419 | 15.4 |
| Phosphatidylinositol (C18:1 16:0) | 836 | 581 553 417 | 16.0 |
| Phosphatidylinositol (C16:0 16:0) | 810 | 553 417 | 15.1 |
| LysoPE (P-18:0/0:0) phospho-ether lipid | 464.9 | 403 267 196 | 18.6 |
| Lysophosphatidylethanolamine (C18:0) | 480 | 283 | 17.6 |
| Lysophosphatidylethanolamine (C18:1) | 478 | 281 | 17.6 |
| Lysophosphatidylethanolamine (C16:0) | 452 | 255 | 18.0 |
| Lysophosphatidylcholine (C18:0) | 508 | 283 | 20.5 |
| Lysophosphatidylcholine (C16:0) | 480 | 255 | 19.5 |
| Matrix | Lipids Included | % of Explained Variance (Cumulative Variance) |
|---|---|---|
| PC 1 | LysoPE(P-18:0/0:0) phospho-ether lipid Lysophosphatidylethanolamine (C18:0) Lysophosphatidylethanolamine (C18:1) Lysophosphatidylethanolamine (C16:0) Lysophosphatidylcholine (C16:0) | 29.3% (29.3%) |
| PC 2 | Seminolipid (O-32.0) Phosphatidic acid (C18:0 20:4) Phosphatidylcholine (C22:6 16:1) Phosphatidylcholine (C18:1 16:0) Phosphatidylinositol (C18:0 18:1) Phosphatidylinositol (C18:0 20:4) Phosphatidylinositol (C18:1 16:0) Phosphatidylinositol (C16:0 16:0) | 16.3% (45.6%) |
| PC 3 | Phosphatidic acid (C18:1 18:1) Phosphatidylcholine (C14:0 18:1) Phosphatidylcholine (C22:6 16:1) Phosphatidylcholine (C18:1 16:0) Phosphatidylserine (C18:0 18:1) Phosphatidylinositol (C18:1 16:0) | 11.0% (56.6%) |
| PC 4 | Phosphatidic acid (C18:0 20:4) Phosphatidic acid (C18:0 22:6) Lysophosphatidylcholine (C18:0) | 8.9% (65.5%) |
| PC 5 | Cholesterol sulfate Phosphatidic acid (C18:1 18:0) Phosphatidylserine (C18:0 18:1) | 8.5% (74.0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Nisio, A.; De Toni, L.; Sabovic, I.; Vignoli, A.; Tenori, L.; Dall’Acqua, S.; Sut, S.; La Vignera, S.; Condorelli, R.A.; Giacone, F.; et al. Lipidomic Profile of Human Sperm Membrane Identifies a Clustering of Lipids Associated with Semen Quality and Function. Int. J. Mol. Sci. 2024, 25, 297. https://doi.org/10.3390/ijms25010297
Di Nisio A, De Toni L, Sabovic I, Vignoli A, Tenori L, Dall’Acqua S, Sut S, La Vignera S, Condorelli RA, Giacone F, et al. Lipidomic Profile of Human Sperm Membrane Identifies a Clustering of Lipids Associated with Semen Quality and Function. International Journal of Molecular Sciences. 2024; 25(1):297. https://doi.org/10.3390/ijms25010297
Chicago/Turabian StyleDi Nisio, Andrea, Luca De Toni, Iva Sabovic, Alessia Vignoli, Leonardo Tenori, Stefano Dall’Acqua, Stefania Sut, Sandro La Vignera, Rosita Angela Condorelli, Filippo Giacone, and et al. 2024. "Lipidomic Profile of Human Sperm Membrane Identifies a Clustering of Lipids Associated with Semen Quality and Function" International Journal of Molecular Sciences 25, no. 1: 297. https://doi.org/10.3390/ijms25010297
APA StyleDi Nisio, A., De Toni, L., Sabovic, I., Vignoli, A., Tenori, L., Dall’Acqua, S., Sut, S., La Vignera, S., Condorelli, R. A., Giacone, F., Ferlin, A., Foresta, C., & Garolla, A. (2024). Lipidomic Profile of Human Sperm Membrane Identifies a Clustering of Lipids Associated with Semen Quality and Function. International Journal of Molecular Sciences, 25(1), 297. https://doi.org/10.3390/ijms25010297