NOD-like Receptors in the Eye: Uncovering Its Role in Diabetic Retinopathy


 ,
, 
Abstract
1. Introduction
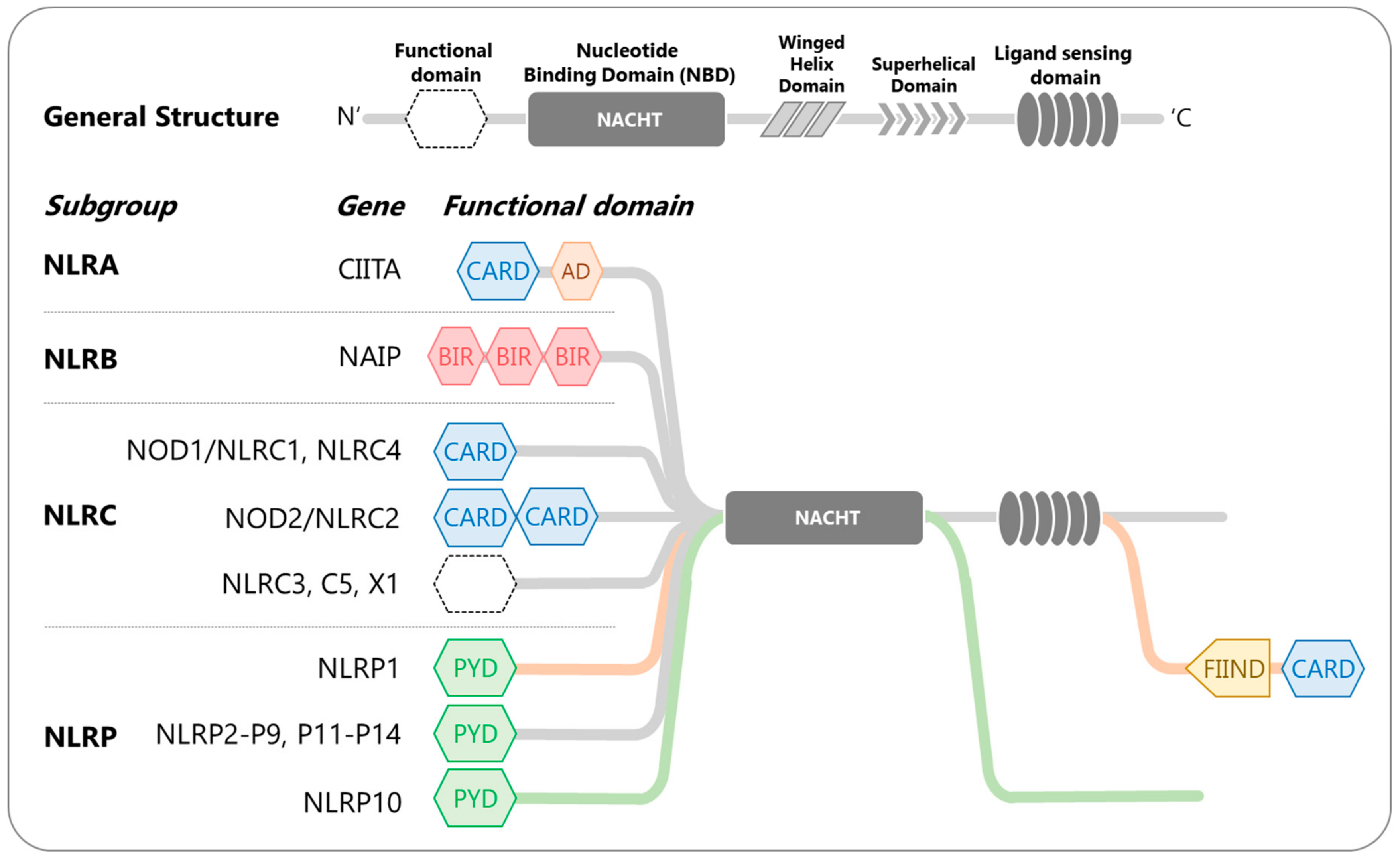
2. Nucleotide-Binding and Oligomerization Domain (NOD)-like Receptors (NLRs)
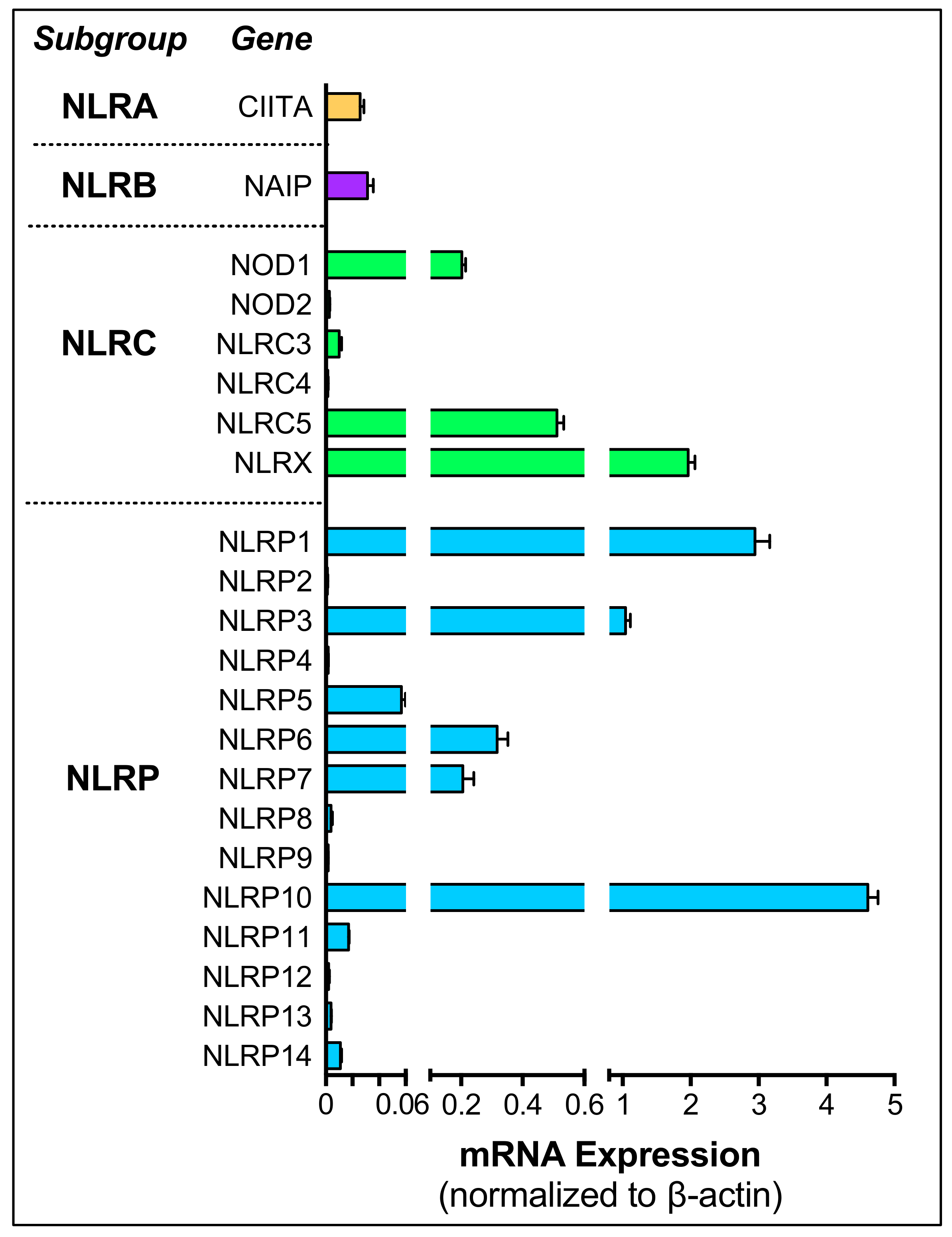
3. NOD-like Receptors in the Ocular Tissues
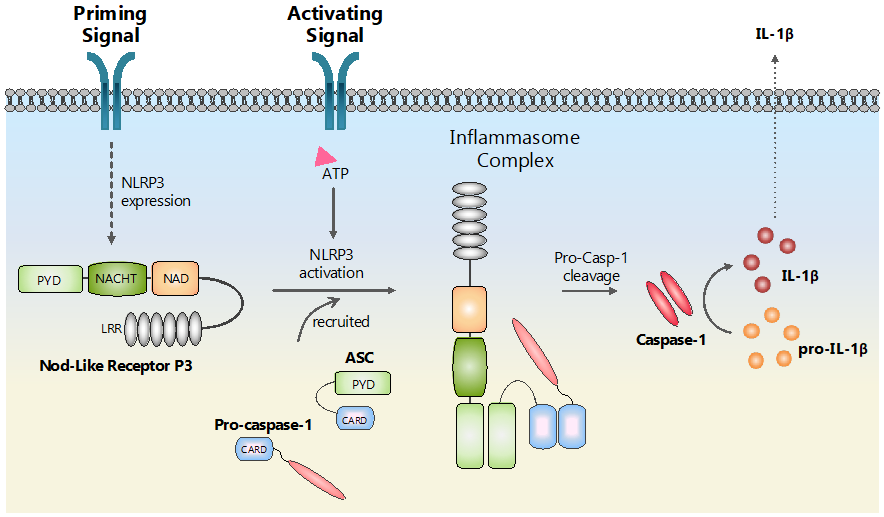
4. NLRs in Diabetic Retinopathy
5. Future Directions
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMD | Age-related macular degeneration |
| AGEs | Advanced glycation endproducts |
| ARPE-19 | Human retinal pigment epithelial cell line |
| ASC | Apoptosis speck-like protein containing a CARD |
| BRB | Blood-retinal barrier |
| CAPs | Cryopyrin-associated periodic syndrome |
| CNV | Choroidal neovascularization |
| DAMPS | Damage-associated molecular patterns |
| DM | Diabetes mellitus |
| DR | Diabetic retinopathy |
| GA | Geographic atrophy |
| HMC3 | Human brain microglia cell line |
| HRMEC | Human retinal microvascular endothelial cells |
| I/R | Ischemic/Reperfusion |
| IOP | Intraocular pressure |
| NLRs | NOD-like receptors |
| NOD | Nucleotide-Binding and Oligomerization Domain |
| NPDR | Non-Proliferative DR |
| ONC | Optic nerve crush |
| PAMPS | Pathogen-associated molecular patterns |
| PDR | Proliferative DR |
| RGCs | Retinal ganglion cells |
| ROS | Reactive oxygen species |
| TGS | Transcriptional gene silencing |
| TXNIP | Thioredoxin-interacting protein |
References
- Tang, J.; Kern, T. Inflammation in Diabetic Retinopathy. Prog. Retin. Eye Res. 2011, 30, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Rübsam, A.; Parikh, S.; Fort, P.E. Role of Inflammation in diabetic retinopathy. Int. J. Mol. Sci. 2018, 19, 942. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.Y.; Nuñez, G. Sterile inflammation: Sensing and reacting to damage. Nat. Rev. Immunol. 2010, 10, 826–837. [Google Scholar] [CrossRef] [PubMed]
- Kuemmerle-Deschner, J.B. Caps—Pathogenesis, presentation and treatment of an autoinflammatory disease. Semin. Immunopathol. 2015, 37, 377–385. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.; Al-Shabrawey, M.; Caldwell, R.W.; Caldwell, R.B. Inflammation and diabetic retinal microvascular complications. J. Cardiovasc. Dis. Res. 2011, 2, 96–103. [Google Scholar] [CrossRef]
- Wooff, Y.; Man, S.M.; Aggio-Bruce, R.; Natoli, R.; Fernando, N. IL-1 Family Members Mediate Cell Death, Inflammation and Angiogenesis in Retinal Degenerative Diseases. Front. Immunol. 2019, 10, 1618. [Google Scholar] [CrossRef]
- Xu, H.; Chen, M. Diabetic retinopathy and dysregulated innate immunity. Vision Res. 2017, 139, 39–46. [Google Scholar] [CrossRef]
- Ferguson, T.A.; Griffith, T.S. A vision of cell death: Insights into immune privilege. Immunol. Rev. 1997, 156, 167–184. [Google Scholar] [CrossRef]
- Forrester, J.V.; Xu, H. Good news-bad news: The Yin and Yang of immune privilege in the eye. Front. Immunol. 2012, 3, 338. [Google Scholar] [CrossRef]
- Nakao, S.; Hafezi-Moghadam, A.; Ishibashi, T. Lymphatics and lymphangiogenesis in the eye. J. Ophthalmol. 2012, 2012, 783163. [Google Scholar] [CrossRef]
- Wenkel, H.; Streilein, J.W. Evidence that retinal pigment epithelium functions as an immune-privileged tissue. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3467–3473. [Google Scholar]
- Abcouwer, S.F. Neural inflammation and the microglial response in diabetic retinopathy. J. Ocul. Biol. Dis. Infor. 2011, 4, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.R.; Vaidya, T.; Gadde, S.G.; Yadav, N.K.; Sethu, S.; Hainsworth, D.P.; Mohan, R.R.; Ghosh, A.; Chaurasia, S.S. Correlation between systemic S100A8 and S100A9 levels and severity of diabetic retinopathy in patients with type 2 diabetes mellitus. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Yerramothu, P.; Vijay, A.K.; Willcox, M.D.P. Inflammasomes, the eye and anti-inflammasome therapy. Eye 2018, 32, 491–505. [Google Scholar] [CrossRef]
- Huang, H.; Gandhi, J.K.; Zhong, X.; Wei, Y.; Gong, J.; Duh, E.J.; Vinores, S.A. TNFα Is Required for Late BRB Breakdown in Diabetic Retinopathy, and Its Inhibition Prevents Leukostasis and Protects Vessels and Neurons from Apoptosis. Investig. Opthalmology Vis. Sci. 2011, 52, 1336–1343. [Google Scholar] [CrossRef]
- Funatsu, H.; Noma, H.; Mimura, T.; Eguchi, S.; Hori, S. Association of Vitreous Inflammatory Factors with Diabetic Macular Edema. Ophthalmology 2009, 116, 73–79. [Google Scholar] [CrossRef]
- Vujosevic, S.; Simó, R. Local and systemic inflammatory biomarkers of diabetic retinopathy: An integrative approach. Investig. Ophthalmol. Vis. Sci. 2017, 58, BIO68–BIO75. [Google Scholar] [CrossRef]
- Yau, J.W.Y.; Rogers, S.L.; Kawasaki, R.; Lamoureux, E.L.; Kowalski, J.W.; Bek, T.; Chen, S.J.; Dekker, J.M.; Fletcher, A.; Grauslund, J.; et al. Global prevalence and major risk factors of diabetic retinopathy. Diabetes Care 2012, 35, 556–564. [Google Scholar] [CrossRef]
- Ding, J.; Wong, T.Y. Current Epidemiology of Diabetic Retinopathy and Diabetic Macular Edema. Curr. Diab. Rep. 2012, 12, 346–354. [Google Scholar] [CrossRef]
- International Diabetes Federation. IDF Diabetes Atlas, 8th ed.; International Diabetes Federation: Brussels, Belgium, 2017. [Google Scholar]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report, 2017; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2017. [Google Scholar]
- Saaddine, J.B.; Honeycutt, A.A.; Narayan, K.M.V.; Zhang, X.; Klein, R.; Boyle, J.P. Projection of diabetic retinopathy and other major eye diseases among people with diabetes mellitus: United States, 2005–2050. Arch. Ophthalmol. 2008, 126, 1740–1747. [Google Scholar] [CrossRef]
- Jones, C.D.; Greenwood, R.H.; Misra, A.; Bachmann, M.O. Incidence and Progression of Diabetic Retinopathy During 17 Years of a Population-Based Screening Program in England. Diabetes Care 2012, 35, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Kern, T.S. Contributions of inflammatory processes to the development of the early stages of diabetic retinopathy. Exp. Diabetes Res. iabetes Res. 2007, 2007, 95103. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.; Mitchell, P.; Wong, T.Y. Diabetic Retinopathy. Lancet 2010, 376, 124–136. [Google Scholar] [CrossRef]
- Wilkinson, C.P.; Ferris, F.L.; Klein, R.E.; Lee, P.P.; Agardh, C.D.; Davis, M.; Dills, D.; Kampik, A.; Pararajasegaram, R.; Verdaguer, J.T.; et al. Proposed international clinical diabetic retinopathy and diabetic macular edema disease severity scales. Ophthalmology 2003, 110, 1677–1682. [Google Scholar] [CrossRef]
- Kern, T.S.; Engerman, R.L. Vascular lesions in diabetes are distributed non-uniformly within the retina. Exp. Eye Res. 1995, 60, 545–549. [Google Scholar] [CrossRef]
- Curtis, T.M.; Gardiner, T.A.; Stitt, A.W. Microvascular lesions of diabetic retinopathy: Clues towards understanding pathogenesis? Eye 2009, 23, 1496–1508. [Google Scholar] [CrossRef]
- Durham, J.T.; Herman, I.M. Microvascular modifications in diabetic retinopathy. Curr. Diab. Rep. 2011, 11, 253–264. [Google Scholar] [CrossRef]
- Cai, J.; Boulton, M. The pathogenesis of diabetic retinopathy: Old concepts and new questions. Eye 2002, 16, 242–260. [Google Scholar] [CrossRef]
- Stewart, M.W. Treatment of diabetic retinopathy: Recent advances and unresolved challenges. World J. Diabetes 2016, 7, 333–341. [Google Scholar] [CrossRef]
- Gross, J.G.; Glassman, A.R.; Jampol, L.M.; Inusah, S.; Aiello, L.P.; Antoszyk, A.N.; Baker, C.W.; Berger, B.B.; Bressler, N.M.; Browning, D.; et al. Panretinal photocoagulation vs. intravitreous ranibizumab for proliferative diabetic retinopathy: A randomized clinical trial. JAMA 2015, 314, 2137–2146. [Google Scholar]
- Oellers, P.; Mahmoud, T.H. Surgery for proliferative diabetic retinopathy: New tips and tricks. J. Ophthalmic Vis. Res. 2016, 11, 93–99. [Google Scholar] [PubMed]
- Bolinger, M.T.; Antonetti, D.A. Moving past anti-VEGF: Novel therapies for treating diabetic retinopathy. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Zorena, K. Anti-Inflammatory Therapy in Diabetic Retinopathy. Mediators Inflamm. 2014, 2014, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Motta, V.; Soares, F.; Sun, T.; Philpott, D.J. NOD-like receptors: Versatile cytosolic sentinels. Physiol. Rev. 2015, 95, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Fritz, J.H.; Ferrero, R.L.; Philpott, D.J.; Girardin, S.E. Nod-like proteins in immunity, inflammation and disease. Nat. Immunol. 2006, 7, 1250–1257. [Google Scholar] [CrossRef]
- Martinon, F.; Mayor, A.; Tschopp, J. The Inflammasomes: Guardians of the Body. Annu. Rev. Immunol. 2009, 27, 229–265. [Google Scholar] [CrossRef]
- Duéñez-Guzmán, E.A.; Haig, D. The evolution of reproduction-related NLRP genes. J. Mol. Evol. 2014, 78, 194–201. [Google Scholar] [CrossRef]
- McDaniel, P.; Wu, X. Identification of oocyte-selective NLRP genes in rhesus macaque monkeys (Macaca mulatta). Mol. Reprod. Dev. 2009, 76, 151–159. [Google Scholar] [CrossRef]
- Peng, H.; Chang, B.; Lu, C.; Su, J.; Wu, Y.; Lv, P.; Wang, Y.; Liu, J.; Zhang, B.; Quan, F.; et al. Nlrp2, a maternal effect gene required for early embryonic development in the mouse. PLoS ONE 2012, 7, e30344. [Google Scholar] [CrossRef]
- Wu, X. Maternal depletion of NLRP5 blocks early embryogenesis in rhesus macaque monkeys (Macaca mulatta). Hum. Reprod. 2009, 24, 415–424. [Google Scholar] [CrossRef]
- Zhang, P.; Dixon, M.; Zucchelli, M.; Hambiliki, F.; Levkov, L.; Hovatta, O.; Kere, J. Expression analysis of the NLRP gene family suggests a role in human preimplantation development. PLoS ONE 2008, 3, e2755. [Google Scholar] [CrossRef] [PubMed]
- Ogura, Y.; Bonen, D.K.; Inohara, N.; Nicolae, D.L.; Chen, F.F.; Ramos, R.; Britton, H.; Moran, T.; Karaliuskas, R.; Duerr, R.H.; et al. A frameshift mutation in NOD2 associated with susceptibility to Crohn’s disease. Nature 2001, 411, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Guerder, S.; Hong, S.C.; Van Ewijk, W.; Flavell, R.A. Mice lacking the MHC class II transactivator (CIITA) show tissue-specific impairment of MHC class II expression. Immunity 1996, 4, 167–178. [Google Scholar] [CrossRef]
- Staehli, F.; Ludigs, K.; Heinz, L.X.; Seguín-Estévez, Q.; Ferrero, I.; Braun, M.; Schroder, K.; Rebsamen, M.; Tardivel, A.; Mattmann, C.; et al. NLRC5 Deficiency Selectively Impairs MHC Class I- Dependent Lymphocyte Killing by Cytotoxic T Cells. J. Immunol. 2012, 188, 3820–3828. [Google Scholar] [CrossRef] [PubMed]
- Kofoed, E.M.; Vance, R.E. Innate immune recognition of bacterial ligands by NAIPs determines inflammasome specificity. Nature 2011, 477, 592–595. [Google Scholar] [CrossRef] [PubMed]
- Lightfield, K.L.; Persson, J.; Brubaker, S.W.; Witte, C.E.; von Moltke, J.; Dunipace, E.A.; Henry, T.; Sun, Y.H.; Cado, D.; Dietrich, W.F.; et al. Critical function for Naip5 in inflammasome activation by a conserved carboxy-terminal domain of flagellin. Nat. Immunol. 2008, 9, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Caruso, R.; Warner, N.; Inohara, N.; Núñez, G. NOD1 and NOD2: Signaling, host defense, and inflammatory disease. Immunity 2014, 41, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Inohara, N.; Hernandez, L.D.; Galán, J.E.; Núñez, G.; Janeway, C.A.; Medzhitov, R.; Flavell, R.A. RICK/Rip2/CARDIAK mediates signalling for receptors of the innate and adaptive immune systems. Nature 2002, 416, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Zambetti, L.P.; Laudisi, F.; Licandro, G.; Ricciardi-Castagnoli, P.; Mortellaro, A. The rhapsody of NLRPs: Master players of inflammation… and a lot more. Immunol. Res. 2012, 53, 78–90. [Google Scholar] [CrossRef]
- Martinon, F.; Pétrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef]
- Pazár, B.; Ea, H.-K.; Narayan, S.; Kolly, L.; Bagnoud, N.; Chobaz, V.; Roger, T.; Lioté, F.; So, A.; Busso, N. Basic calcium phosphate crystals induce monocyte/macrophage IL-1β secretion through the NLRP3 inflammasome in vitro. J. Immunol. 2011, 186, 2495–2502. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Ojala, J.; Suuronen, T.; Kaarniranta, K.; Kauppinen, A. Amyloid-beta oligomers set fire to inflammasomes and induce Alzheimer’s pathology. J. Cell. Mol. Med. 2008, 12, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Gris, D.; Lei, Y.; Jha, S.; Zhang, L.; Huang, M.T.H.; Brickey, W.J.; Ting, J.P.Y. Fatty acid-induced NLRP3-ASC inflammasome activation interferes with insulin signaling. Nat. Immunol. 2011, 12, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The Inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Elinav, E.; Strowig, T.; Kau, A.L.; Henao-Mejia, J.; Thaiss, C.A.; Booth, C.J.; Peaper, D.R.; Bertin, J.; Eisenbarth, S.C.; Gordon, J.I.; et al. NLRP6 inflammasome regulates colonic microbial ecology and risk for colitis. Cell 2011, 145, 745–757. [Google Scholar] [CrossRef]
- Khare, S.; Dorfleutner, A.; Bryan, N.B.; Yun, C.; Radian, A.D.; de Almeida, L.; Rojanasakul, Y.; Stehlik, C. An NLRP7-containing inflammasome mediates recognition of microbial lipopeptides in human macrophages. Immunity 2012, 36, 464–476. [Google Scholar] [CrossRef]
- Vladimer, G.I.; Weng, D.; Paquette, S.W.M.; Vanaja, S.K.; Rathinam, V.A.K.; Aune, M.H.; Conlon, J.E.; Burbage, J.J.; Proulx, M.K.; Liu, Q.; et al. The NLRP12 inflammasome recognizes Yersinia pestis. Immunity 2012, 37, 96–107. [Google Scholar] [CrossRef]
- Kufer, T.A.; Sansonetti, P.J. NLR functions beyond pathogen recognition. Nat. Immunol. 2011, 12, 121–128. [Google Scholar] [CrossRef]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nũez, G.; Schnurr, M.; et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef]
- Rajamäki, K.; Lappalainen, J.; Oörni, K.; Välimäki, E.; Matikainen, S.; Kovanen, P.T.; Eklund, K.K. Cholesterol crystals activate the NLRP3 inflammasome in human macrophages: A novel link between cholesterol metabolism and inflammation. PLoS ONE 2010, 5, e11765. [Google Scholar] [CrossRef]
- Hu, Y.; Lin, H.; Dib, B.; Atik, A.; Bouzika, P.; Lin, C.; Yan, Y.; Tang, S.; Miller, J.W.; Vavvas, D.G. Cholesterol crystals induce inflammatory cytokines expression in a human retinal pigment epithelium cell line by activating the NF-κB pathway. Discov. Med. 2014, 18, 7–14. [Google Scholar] [PubMed]
- He, Q.; You, H.; Li, X.M.; Liu, T.H.; Wang, P.; Wang, B.E. HMGB1 promotes the synthesis of pro-il-1β and pro-il-18 by activation of p38 MAPK and NF-kB through receptors for advanced glycation end-products in macrophages. Asian Pacific J. Cancer Prev. 2012, 13, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.D.; Frank, M.G.; Tracey, K.J.; Watkins, L.R.; Maier, S.F. Stress induces the danger-associated molecular pattern HMGB-1 in the hippocampus of male Sprague Dawley rats: A priming stimulus of microglia and the NLRP3 inflammasome. J. Neurosci. 2015, 35, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-L.; Zhang, X.-D.; Li, Y.-Y.; Chen, X.-M.; Tang, D.-R.; Ran, R.-J. Involvement of HMGB1 mediated signalling pathway in diabetic retinopathy: Evidence from type 2 diabetic rats and ARPE-19 cells under diabetic condition. Br. J. Ophthalmol. 2013, 97, 1598–1603. [Google Scholar] [CrossRef] [PubMed]
- Benko, S.; Tozser, J.; Miklossy, G.; Varga, A.; Kadas, J.; Csutak, A.; Berta, A.; Rajnavolgyi, E. Constitutive and UV-B modulated transcription of NOD-like receptors and their functional partners in human corneal epithelial cells. Mol. Vis. 2008, 14, 1575–1583. [Google Scholar] [PubMed]
- Oh, J.Y.; Ko, J.H.; Ryu, J.S.; Lee, H.J.; Kim, M.K.; Wee, W.R. Transcription Profiling of NOD-like receptors in the Human Cornea with Disease. Ocul. Immunol. Inflamm. 2017, 25, 364–369. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, J.; Xin, Z.; Wu, X. Aspergillus fumigatus triggers innate immune response via NOD1 signaling in human corneal epithelial cells. Exp. Eye Res. 2014, 127, 170–178. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Xin, Z.; Wu, X. The crosstalk between TLR2 and NOD2 in Aspergillus fumigatus keratitis. Mol. Immunol. 2015, 64, 235–243. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, G.; Yan, J.; Xu, R.; Che, C.; Zheng, H.; Zhu, G.; Zhang, J. Pannexin 1 Channels Contribute to IL-1β Expression via NLRP3/Caspase-1 Inflammasome in Aspergillus Fumigatus Keratitis. Curr. Eye Res. 2019, 44, 716–725. [Google Scholar] [CrossRef]
- McClellan, S.A.; Jerome, A.; Suvas, S.; Hazlett, L.D. NLRC4 regulates caspase-1 and IL-1beta production in a CD11blowLy6Glow population of cells required for resistance to Pseudomonas aeruginosa keratitis. PLoS ONE 2017, 12, e0185718. [Google Scholar] [CrossRef]
- Karmakar, M.; Katsnelson, M.; Malak, H.A.; Greene, N.G.; Howell, S.J.; Hise, A.G.; Camilli, A.; Kadioglu, A.; Dubyak, G.R.; Pearlman, E. Neutrophil IL-1β Processing Induced by Pneumolysin Is Mediated by the NLRP3/ASC Inflammasome and Caspase-1 Activation and Is Dependent on K + Efflux. J. Immunol. 2015, 194, 1763–1775. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Sakimoto, T.; Yamagami, S. Pro-inflammatory role of NLRP3 inflammasome in experimental sterile corneal inflammation. Sci. Rep. 2019, 9, 9596. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Abbondante, S.; Karmakar, M.; de Jesus Carrion, S.; Che, C.; Hise, A.G.; Pearlman, E. Neutrophil Caspase-11 Is Required for Cleavage of Caspase-1 and Secretion of IL-1β in Aspergillus fumigatus Infection. J. Immunol. 2018, 201, 2767–2775. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Ren, Y.; Reinach, P.S.; Xiao, B.; Lu, H.; Zhu, Y.; Qu, J.; Chen, W. Reactive oxygen species activated NLRP3 inflammasomes initiate inflammation in hyperosmolarity stressed human corneal epithelial cells and environment-induced dry eye patients. Exp. Eye Res. 2015, 134, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Hua, X.; Chen, X.; Bian, F.; Yuan, X.; Zhang, L.; Wang, X.; Chen, D.; Deng, R.; Li, Z.; et al. Mitochondrial DNA oxidation induces imbalanced activity of NLRP3/NLRP6 inflammasomes by activation of caspase-8 and BRCC36 in dry eye. J. Autoimmun. 2017, 80, 65–76. [Google Scholar] [CrossRef]
- Zheng, Q.; Ren, Y.; Reinach, P.S.; She, Y.; Xiao, B.; Hua, S.; Qu, J.; Chen, W. Reactive oxygen species activated NLRP3 inflammasomes prime environment-induced murine dry eye. Exp. Eye Res. 2014, 125, 1–8. [Google Scholar] [CrossRef]
- Bian, F.; Xiao, Y.; Zaheer, M.; Volpe, E.A.; Pflugfelder, S.C.; Li, D.Q.; De Paiva, C.S. Inhibition of NLRP3 inflammasome pathway by butyrate improves corneal wound healing in corneal alkali burn. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef]
- Zheng, Q.; Tan, Q.; Ren, Y.; Reinach, P.S.; Li, L.; Ge, C.; Qu, J.; Chen, W. Hyperosmotic stress–induced TRPM2 channel activation stimulates NLRP3 inflammasome activity in primary human corneal epithelial cells. Investig. Ophthalmol. Vis. Sci. 2018, 59, 3259–3268. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, J.; Xiang, J.; Li, Y.; Wu, D.; Xu, J. Calcitriol inhibits ROS-NLRP3-IL-1β signaling axis via activation of Nrf2-antioxidant signaling in hyperosmotic stress stimulated human corneal epithelial cells. Redox Biol. 2019, 21, 101093. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, S.; Cancino-Díaz, M.E.; Jiménez-Zamudio, L.; García-Latorre, E.; Cancino-Díaz, J.C. TLRs and NODs mRNA expression pattern in healthy mouse eye. Br. J. Ophthalmol. 2005, 89, 904–910. [Google Scholar] [CrossRef]
- Del Mar Cendra, M.; Christodoulides, M.; Hossain, P. Signaling mediated by Toll-Like receptor 5 sensing of Pseudomonas aeruginosa flagellin influences IL-1β and IL-18 production by primary fibroblasts derived from the human cornea. Front. Cell. Infect. Microbiol. 2017, 7, 130. [Google Scholar]
- Chaurasia, S.; Lim, R.; Lakshminarayanan, R.; Mohan, R. Nanomedicine Approaches for Corneal Diseases. J. Funct. Biomater. 2015, 6, 277–298. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Ishida, W.; Fukushima, A.; Nishida, T. Corneal fibroblasts as sentinel cells and local immune modulators in infectious keratitis. Int. J. Mol. Sci. 2017, 18, 1831. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wei, C.; Wang, S.; Liu, T.; Zhai, H.; Shi, W. Upregulation of NLRP3 inflammasome components in Mooren’s ulcer. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 607–612. [Google Scholar] [CrossRef]
- McGilligan, V.E.; Gregory-Ksander, M.S.; Li, D.; Moore, J.E.; Hodges, R.R.; Gilmore, M.S.; Moore, T.C.B.; Dartt, D.A. Staphylococcus aureus Activates the NLRP3 Inflammasome in Human and Rat Conjunctival Goblet Cells. PLoS ONE 2013, 8, e74010. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, H. Pyroptosis in pterygium pathogenesis. Biosci. Rep. 2018, 38, 1–15. [Google Scholar] [CrossRef]
- Li, D.; Hodges, R.R.; Bispo, P.; Gilmore, M.S.; Gregory-Ksander, M.; Dartt, D.A. Neither non-toxigenic Staphylococcus aureus nor commensal S. epidermidi activates NLRP3 inflammasomes in human conjunctival goblet cells. BMJ Open Ophthalmol. 2017, 2. [Google Scholar] [CrossRef]
- Scurrell, E.; Stanley, R.; Schöniger, S. Immunohistochemical detection of NOD1 and NOD2 in the healthy murine and canine eye. Vet. Ophthalmol. 2009, 12, 269–275. [Google Scholar] [CrossRef]
- Chi, W.; Li, F.; Chen, H.; Wang, Y.; Zhu, Y.; Yang, X.; Zhu, J.; Wu, F.; Ouyang, H.; Ge, J.; et al. Caspase-8 promotes NLRP1/NLRP3 inflammasome activation and IL-1β production in acute glaucoma. Proc. Natl. Acad. Sci. USA 2014, 111, 11181–11186. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.; Qi, Y.; Feng, J.; Zhu, L.; Bai, Y.; Wu, H. Protective effects of Erigeron breviscapus Hand.-Mazz. (EBHM) extract in retinal neurodegeneration models. Mol. Vis. 2018, 24, 315–325. [Google Scholar]
- Albalawi, F.; Lu, W.; Beckel, J.M.; Lim, J.C.; McCaughey, S.A.; Mitchell, C.H. The P2 × 7 receptor primes IL-1β and the NLRP3 inflammasome in astrocytes exposed to mechanical strain. Front. Cell. Neurosci. 2017, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Pronin, A.; Pham, D.; An, W.; Dvoriantchikova, G.; Reshetnikova, G.; Qiao, J.; Kozhekbaeva, Z.; Reiser, A.E.; Slepak, V.Z.; Shestopalov, V.I. Inflammasome activation induces pyroptosis in the retina exposed to ocular hypertension injury. Front. Mol. Neurosci. 2019, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhao, M.; Bai, Y.; Huang, L.; Yu, W.; Bian, Z.; Zhao, M.; Li, X. Retinal ischemia/reperfusion injury is mediated by tolllike receptor 4 activation of NLRP3 inflammasomes. Investig. Ophthalmol. Vis. Sci. 2014, 55, 5466–5475. [Google Scholar] [CrossRef] [PubMed]
- Qijun, Z.; Huan, Z.; Ling, G.; Kaijian, C.; Wei, L.; Shuxing, J.; Xiang, C.; Rongdi, Y.; Jian, Y. The levels and significance of inflammasomes in the mouse retina following optic nerve crush. Int. Immunopharmacol. 2019, 71, 313–320. [Google Scholar] [CrossRef]
- Puyang, Z.; Feng, L.; Chen, H.; Liang, P.; Troy, J.B.; Liu, X. Retinal ganglion cell loss is delayed following optic nerve crush in nlrp3 knockout mice. Sci. Rep. 2016, 6, 6–13. [Google Scholar] [CrossRef]
- Harun-Or-Rashid, M.; Inman, D.M. Reduced AMPK activation and increased HCAR activation drive anti-inflammatory response and neuroprotection in glaucoma. J. Neuroinflammation 2018, 15, 313. [Google Scholar] [CrossRef]
- Xu, Z.; Fouda, A.Y.; Lemtalsi, T.; Shosha, E.; Rojas, M.; Liu, F.; Patel, C.; Caldwell, R.W.; Narayanan, S.P.; Caldwell, R.B. Retinal Neuroprotection from Optic Nerve Trauma by Deletion of Arginase 2. Front. Neurosci. 2018, 12, 1–13. [Google Scholar] [CrossRef]
- Chi, W.; Chen, H.; Li, F.; Zhu, Y.; Yin, W.; Zhuo, Y. HMGB1 promotes the activation of NLRP3 and caspase-8 inflammasomes via NF-κB pathway in acute glaucoma. J. Neuroinflammation 2015, 12, 1–12. [Google Scholar] [CrossRef]
- Lin, C.; Wu, F.; Zheng, T.; Wang, X.; Chen, Y.; Wu, X. Kaempferol attenuates retinal ganglion cell death by suppressing NLRP1/NLRP3 inflammasomes and caspase-8 via JNK and NF-κB pathways in acute glaucoma. Eye 2019, 33, 777–784. [Google Scholar] [CrossRef]
- Lei, X.; Zhao, Y. Neovascular glaucoma regulation by arylsulfonyl indoline-benzamide (ASIB) through targeting NF-kB signalling pathway. 3 Biotech. 2019, 9, 211. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, Y.; Wang, J.; Mao, P.; Lv, X.; Yuan, S.; Huang, Z.; Ding, Y.; Xie, P.; Liu, Q. Knockout of Ccr2 alleviates photoreceptor cell death in rodent retina exposed to chronic blue light. Cell Death Dis. 2016, 7, e2468. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Ryals, R.C.; Huang, S.J.; Weller, K.K.; Titus, H.E.; Robb, B.M.; Saad, F.W.; Salam, R.A.; Hammad, H.; Yang, P.; et al. Monomethyl fumarate protects the retina from light- induced retinopathy. Investig. Ophthalmol. Vis. Sci. 2019, 60, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Tsoka, P.; Barbisan, P.R.; Kataoka, K.; Chen, X.N.; Tian, B.; Bouzika, P.; Miller, J.W.; Paschalis, E.I.; Vavvas, D.G. NLRP3 inflammasome in NMDA-induced retinal excitotoxicity. Exp. Eye Res. 2019, 181, 136–144. [Google Scholar] [CrossRef] [PubMed]
- El-Azab, M.F.; Baldowski, B.R.B.; Mysona, B.A.; Shanab, A.Y.; Mohamed, I.N.; Abdelsaid, M.A.; Matragoon, S.; Bollinger, K.E.; Saul, A.; El-Remessy, A.B. Deletion of thioredoxin-interacting protein preserves retinal neuronal function by preventing inflammation and vascular injury. Br. J. Pharmacol. 2014, 171, 1299–1313. [Google Scholar] [CrossRef] [PubMed]
- Viringipurampeer, I.A.; Metcalfe, A.L.; Bashar, A.E.; Sivak, O.; Yanai, A.; Mohammadi, Z.; Moritz, O.L.; Gregory-Evans, C.Y.; Gregory-Evans, K. NLRP3 inflammasome activation drives bystander cone photoreceptor cell death in a P23H rhodopsin model of retinal degeneration. Hum. Mol. Genet. 2016, 25, 1501–1516. [Google Scholar] [CrossRef] [PubMed]
- Appelbaum, T.; Santana, E.; Aguirre, G.D. Strong upregulation of inflammatory genes accompanies photoreceptor demise in canine models of retinal degeneration. PLoS ONE 2017, 12, e0177224. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, C.; Wan, X.S.; Li, S.W. NLRP1 deficiency attenuates diabetic retinopathy (DR) in mice through suppressing inflammation response. Biochem. Biophys. Res. Commun. 2018, 501, 351–357. [Google Scholar] [CrossRef]
- Wang, S.; Ji, L.Y.; Li, L.; Li, J.M. Oxidative stress, autophagy and pyroptosis in the neovascularization of oxygen-induced retinopathy in mice. Mol. Med. Rep. 2019, 19, 927–934. [Google Scholar] [CrossRef]
- Rivera, J.C.; Sitaras, N.; Noueihed, B.; Hamel, D.; Madaan, A.; Zhou, T.; Honoré, J.-C.; Quiniou, C.; Joyal, J.-S.; Hardy, P.; et al. Microglia and interleukin-1β in ischemic retinopathy elicit microvascular degeneration through neuronal semaphorin-3A. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1881–1891. [Google Scholar] [CrossRef]
- Davey, M.P.; Martin, T.M.; Planck, S.R.; Lee, J.; Zamora, D.; Rosenbaum, J.T. Human endothelial cells express NOD2/CARD15 and increase IL-6 secretion in response to muramyl dipeptide. Microvasc. Res. 2006, 71, 103–107. [Google Scholar] [CrossRef]
- Mohamed, I.N.; Hafez, S.S.; Fairaq, A.; Ergul, A.; Imig, J.D.; El-Remessy, A.B. Thioredoxin-interacting protein is required for endothelial NLRP3 inflammasome activation and cell death in a rat model of high-fat diet. Diabetologia 2014, 57, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, S.S.; Lim, R.R.; Parikh, B.H.; Yeo, S.W.; Tun, B.B.; Wong, T.Y.; Luu, C.D.; Agrawal, R.; Ghosh, A.; Mortellaro, A.; et al. The NLRP3 Inflammasome May Contribute to Pathologic Neovascularization in the Advanced Stages of Diabetic Retinopathy. Sci. Rep. 2018, 8, 2847. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.T.; Gao, J.; Cao, S.; Sandhu, N.; Cui, J.Z.; Chou, C.L.; Fang, E.; Matsubara, J.A. Inflammatory mediators induced by amyloid-beta in the retina and RPE in vivo: Implications for inflammasome activation in age-related macular degeneration. Invest. Ophthalmol. Vis. Sci. 2013, 54, 2225–2237. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Lin, R.; Wang, J.; Tao, L.; Fu, X.; Qiu, Y.; Lei, B. Amelioration of amyloid β-induced retinal inflammatory responses by a LXR agonist TO901317 is associated with inhibition of the NF-κB signaling and NLRP3 inflammasome. Neuroscience 2017, 360, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hanus, J.W.; Abu-Asab, M.S.; Shen, D.; Ogilvy, A.; Ou, J.; Chu, X.K.; Shi, G.; Li, W.; Wang, S.; et al. NLRP3 upregulation in retinal pigment epithelium in age-related macular degeneration. Int. J. Mol. Sci. 2016, 17, 73. [Google Scholar] [CrossRef] [PubMed]
- Aredo, B.; Li, T.; Chen, X.; Zhang, K.; Wang, C.X.Z.; Gou, D.; Zhao, B.; He, Y.; Ufret-Vincenty, R.L. A chimeric Cfh transgene leads to increased retinal oxidative stress, inflammation, and accumulation of activated subretinal microglia in mice. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3427–3440. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Fowler, B.J.; Kerur, N.; Arnvig, K.B.; Rao, N.A. NLRP3 inflammasome activation by mycobacterial ESAT-6 and dsRNA in intraocular tuberculosis. Microb. Pathog. 2018, 114, 219–224. [Google Scholar] [CrossRef]
- Zhang, Y.; Lv, X.; Hu, Z.; Ye, X.; Zheng, X.; Ding, Y.; Xie, P.; Liu, Q. Protection of Mcc950 against high-glucose-induced human retinal endothelial cell dysfunction. Cell Death Dis. 2017, 8, e2941. [Google Scholar] [CrossRef]
- Li, S.; Yang, H.; Chen, X. Protective effects of sulforaphane on diabetic retinopathy: Activation of the Nrf2 pathway and inhibition of NLRP3 inflammasome formation. Exp. Anim. 2019, 68, 221–231. [Google Scholar] [CrossRef]
- Hao, J.; Zhang, H.; Yu, J.; Chen, X.; Yang, L. Methylene Blue Attenuates Diabetic Retinopathy by Inhibiting NLRP3 Inflammasome Activation in STZ-induced Diabetic Rats. Ocul. Immunol. Inflamm. 2018, 27, 1–8. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, F.; Zhang, X.; Cheng, R.; Ma, J.X.; Yi, J.; Li, J. Fenofibrate ameliorates diabetic retinopathy by modulating Nrf2 signaling and NLRP3 inflammasome activation. Mol. Cell. Biochem. 2018, 445, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Indaram, M.; Ma, W.; Zhao, L.; Fariss, R.N.; Rodriguez, I.R.; Wong, W.T. 7-Ketocholesterol Increases Retinal Microglial Migration, Activation, and Angiogenicity: A Potential Pathogenic Mechanism Underlying Age-related Macular Degeneration. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Devi, T.S.; Lee, I.; Hüttemann, M.; Kumar, A.; Nantwi, K.D.; Singh, L.P. TXNIP links innate host defense mechanisms to oxidative stress and inflammation in retinal muller glia under chronic hyperglycemia: Implications for diabetic retinopathy. Exp. Diabetes Res. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Coucha, M.; Mohamed, I.N.; Elshaer, S.L.; Mbata, O.; Bartasis, M.L.; El-Remessy, A.B. High fat diet dysregulates microRNA-17-5p and triggers retinal inflammation: Role of endoplasmic-reticulum-stress. World J. Diabetes 2017, 8, 56. [Google Scholar] [CrossRef]
- Hu, L.; Yang, H.; Ai, M.; Jiang, S. Inhibition of TLR4 alleviates the inflammation and apoptosis of retinal ganglion cells in high glucose. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 2199–2210. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, L.; Curtiss, E.; Steinle, J.J. Epac1 Blocks NLRP3 Inflammasome to Reduce IL-1 β in Retinal Endothelial Cells and Mouse Retinal Vasculature. Mediators Inflamm. 2017, 2017, 2860956. [Google Scholar] [CrossRef]
- Tseng, W.A.; Thein, T.; Kinnunen, K.; Lashkari, K.; Gregory, M.S.; D’Amore, P.A.; Ksander, B.R. NLRP3 inflammasome activation in retinal pigment epithelial cells by lysosomal destabilization: Implications for age-related macular degeneration. Invest. Ophthalmol. Vis. Sci. 2013, 54, 110–120. [Google Scholar] [CrossRef]
- Tarallo, V.; Hirano, Y.; Gelfand, B.D.; Dridi, S.; Kerur, N.; Kim, Y.; Cho, W.G.; Kaneko, H.; Fowler, B.J.; Bogdanovich, S.; et al. DICER1 loss and Alu RNA induce age-related macular degeneration via the NLRP3 inflammasome and MyD88. Cell 2012, 149, 847–859. [Google Scholar] [CrossRef]
- Liu, R.T.; Wang, A.; To, E.; Gao, J.; Cao, S.; Cui, J.Z.; Matsubara, J.A. Vinpocetine inhibits amyloid-beta induced activation of NF-κB, NLRP3 inflammasome and cytokine production in retinal pigment epithelial cells. Exp. Eye Res. 2014, 127, 49–58. [Google Scholar] [CrossRef]
- Gelfand, B.D.; Wright, C.B.; Kim, Y.; Yasuma, T.; Yasuma, R.; Li, S.; Fowler, B.J.; Bastos-Carvalho, A.; Kerur, N.; Uittenbogaard, A.; et al. Iron toxicity in the retina requires Alu RNA and the NLRP3 inflammasome. Cell Rep. 2015, 11, 1686–1693. [Google Scholar] [CrossRef]
- Marneros, A. NLRP3 inflammasome blockade inhibits VEGF-A-induced age-related macular degeneration. Cell Rep. 2013, 4, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Prager, P.; Hollborn, M.; Steffen, A.; Wiedemann, P.; Kohen, L.; Bringmann, A. P2Y1 receptor signaling contributes to high salt-induced priming of the NLRP3 inflammasome in retinal pigment epithelial cells. PLoS ONE 2016, 11, e0165653. [Google Scholar] [CrossRef] [PubMed]
- Hollborn, M.; Ackmann, C.; Kuhrt, H.; Doktor, F.; Kohen, L.; Wiedemann, P.; Bringmann, A. Osmotic and hypoxic induction of the complement factor C9 in cultured human retinal pigment epithelial cells: Regulation of VEGF and NLRP3 expression. Mol. Vis. 2018, 24, 518–535. [Google Scholar] [PubMed]
- Zhang, S.; Yu, N.; Zhang, R.; Zhang, S.; Wu, J. Interleukin-17A induces IL-1β secretion from RPE cells via the NLRP3 inflammasome. Investig. Ophthalmol. Vis. Sci. 2016, 57, 312–319. [Google Scholar] [CrossRef]
- Kerur, N.; Hirano, Y.; Tarallo, V.; Fowler, B.J.; Bastos-Carvalho, A.; Yasuma, T.; Yasuma, R.; Kim, Y.; Hinton, D.R.; Kirschning, C.J.; et al. TLR-independent and P2 × 7-dependent signaling mediate Alu RNA-induced NLRP3 inflammasome activation in geographic atrophy. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7395–7401. [Google Scholar] [CrossRef]
- Gnanaguru, G.; Choi, A.R.; Amarnani, D.; D’Amore, P.A. Oxidized lipoprotein uptake through the CD36 receptor activates the NLRP3 inflammasome in human retinal pigment epithelial cells. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4704–4712. [Google Scholar] [CrossRef]
- Jin, X.; Wang, C.; Wu, W.; Liu, T.; Ji, B.; Zhou, F. Cyanidin-3-glucoside alleviates 4-Hydroxyhexenal-induced NLRP3 inflammasome activation via JNK-c-Jun/AP-1 pathway in human retinal pigment epithelial cells. J. Immunol. Res. 2018, 2018. [Google Scholar] [CrossRef]
- Cao, S.; Wang, J.C.C.; Gao, J.; Wong, M.; To, E.; White, V.A.; Cui, J.Z.; Matsubara, J.A. CFH Y402H polymorphism and the complement activation product C5a: Effects on NF-κB activation and inflammasome gene regulation. Br. J. Ophthalmol. 2016, 100, 713–718. [Google Scholar] [CrossRef]
- Piippo, N.; Korkmaz, A.; Hytti, M.; Kinnunen, K.; Salminen, A.; Atalay, M.; Kaarniranta, K.; Kauppinen, A. Decline in cellular clearance systems induces inflammasome signaling in human ARPE-19 cells. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 3038–3046. [Google Scholar] [CrossRef]
- Mao, X.; Pan, T.; Shen, H.; Xi, H.; Yuan, S.; Liu, Q. The rescue effect of mesenchymal stem cell on sodium iodate-induced retinal pigment epithelial cell death through deactivation of NF-κB-mediated NLRP3 inflammasome. Biomed. Pharmacother. 2018, 103, 517–523. [Google Scholar] [CrossRef]
- Mugisho, O.O.; Green, C.R.; Kho, D.T.; Zhang, J.; Graham, E.S.; Acosta, M.L.; Rupenthal, I.D. The inflammasome pathway is amplified and perpetuated in an autocrine manner through connexin43 hemichannel mediated ATP release. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 385–393. [Google Scholar] [CrossRef]
- Wang, K.; Zhu, X.; Zhang, K.; Yao, Y.; Zhuang, M.; Tan, C.; Zhou, F.; Zhu, L. Puerarin inhibits amyloid β-induced NLRP3 inflammasome activation in retinal pigment epithelial cells via suppressing ROS-dependent oxidative and endoplasmic reticulum stresses. Exp. Cell Res. 2017, 357, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Mao, K. Salvianolic Acid A Protects Retinal Pigment Epithelium from O… Salvianolic Acid A Protects Retinal Pigment Epithelium from O. Discov. Med. 2019, 23, 129–147. [Google Scholar]
- Wang, K.; Yao, Y.; Zhu, X.; Zhang, K.; Zhou, F.; Zhu, L. Amyloid β induces NLRP3 inflammasome activation in retinal pigment epithelial cells via NADPH oxidase- and mitochondria-dependent ROS production. J. Biochem. Mol. Toxicol. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Zhang, Z.; Wang, X.; Li, R.; Hou, W.; Bi, W.; Zhang, X. Inhibition of autophagy induces IL-1β release from ARPE-19 cells via ROS mediated NLRP3 inflammasome activation under high glucose stress. Biochem. Biophys. Res. Commun. 2015, 463, 1071–1076. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, Y.; Zhang, Y.; Yang, J.; He, G.; Chen, S. Photo-Oxidative Blue-Light Stimulation in Retinal Pigment Epithelium Cells Promotes Exosome Secretion and Increases the Activity of the NLRP3 Inflammasome. Curr. Eye Res. 2019, 44, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Piippo, N.; Korhonen, E.; Hytti, M.; Skottman, H.; Kinnunen, K.; Josifovska, N.; Petrovski, G.; Kaarniranta, K.; Kauppinen, A. Hsp90 inhibition as a means to inhibit activation of the NLRP3 inflammasome. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Gao, J.; Liu, R.T.; Cao, S.; Cui, J.Z.; Wang, A.; To, E.; Matsubara, J.A. NLRP3 Inflammasome: Activation and Regulation in Age-Related Macular Degeneration. Mediators Inflamm. 2015, 2015, 690243. [Google Scholar] [CrossRef]
- Doyle, S.L.; Campbell, M.; Ozaki, E.; Salomon, R.G.; Mori, A.; Kenna, P.F.; Farrar, G.J.; Kiang, A.S.; Humphries, M.M.; Lavelle, E.C.; et al. NLRP3 has a protective role in age-related macular degeneration through the induction of IL-18 by drusen components. Nat. Med. 2012, 18, 791–798. [Google Scholar] [CrossRef]
- Anderson, O.A.; Finkelstein, A.; Shima, D.T. A2E Induces IL-1ß Production in Retinal Pigment Epithelial Cells via the NLRP3 Inflammasome. PLoS ONE 2013, 8, e67263. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Kowluru, A.; Mishra, M.; Kumar, B. Oxidative stress and epigenetic modifications in the pathogenesis of diabetic retinopathy. Prog. Retin. Eye Res. 2015, 48, 40–61. [Google Scholar] [CrossRef]
- Kanwar, M.; Chan, P.S.; Kern, T.S.; Kowluru, R.A. Oxidative damage in the retinal mitochondria of diabetic mice: Possible protection by superoxide dismutase. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3805–3811. [Google Scholar] [CrossRef] [PubMed]
- Goto, H.; Nishikawa, T.; Sonoda, K.; Kondo, T.; Kukidome, D.; Fujisawa, K.; Yamashiro, T.; Motoshima, H.; Matsumura, T.; Tsuruzoe, K.; et al. Endothelial MnSOD overexpression prevents retinal VEGF expression in diabetic mice. Biochem. Biophys. Res. Commun. 2008, 366, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Atasi, L.; Ho, Y.S. Role of mitochondrial superoxide dismutase in the development of diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1594–1599. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, M. The polyol pathway as a mechanism for diabetic retinopathy: Attractive, elusive, and resilient. Exp. Diabetes Res. 2007, 2007, 61038. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Odenbach, S. Role of interleukin-1β in the development of retinopathy in rats: Effect of antioxidants. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4161–4166. [Google Scholar] [CrossRef]
- Kirk, S.L.; Karlik, S.J. VEGF and vascular changes in chronic neuroinflammation. J. Autoimmun. 2003, 21, 353–363. [Google Scholar] [CrossRef]
- Rangasamy, S.; McGuire, P.G.; Franco Nitta, C.; Monickaraj, F.; Oruganti, S.R.; Das, A. Chemokine mediated monocyte trafficking into the retina: Role of inflammation in alteration of the blood-retinal barrier in diabetic retinopathy. PLoS ONE 2014, 9, e108508. [Google Scholar] [CrossRef]
- Joussen, A.M.; Poulaki, V.; Le, M.L.; Koizumi, K.; Esser, C.; Janicki, H.; Schraermeyer, U.; Kociok, N.; Fauser, S.; Kirchhof, B.; et al. A central role for inflammation in the pathogenesis of diabetic retinopathy. FASEB J. 2004, 18, 1450–1452. [Google Scholar] [CrossRef]
- Vincent, J.A.; Mohr, S. Inhibition of caspase-1/interleukin-1beta signaling prevents degeneration of retinal capillaries in diabetes and galactosemia. Diabetes 2007, 56, 224–230. [Google Scholar] [CrossRef]
- Xie, T.X.; Xia, Z.; Zhang, N.; Gong, W.; Huang, S. Constitutive NF-κB activity regulates the expression of VEGF and IL-8 and tumor angiogenesis of human glioblastoma. Oncol. Rep. 2010, 23, 725–732. [Google Scholar] [PubMed]
- Zheng, L.; Szabó, C.; Kern, T.S. Poly(ADP-ribose) polymerase is involved in the development of diabetic retinopathy via regulation of nuclear factor-κB. Diabetes 2004, 53, 2960–2967. [Google Scholar] [CrossRef] [PubMed]
- Drel, V.R.; Xu, W.; Zhang, J.; Kador, P.F.; Ali, T.K.; Shin, J.; Julius, U.; Slusher, B.; El-Remessy, A.B.; Obrosova, I.G. Poly(ADP-ribose)polymerase inhibition counteracts cataract formation and early retinal changes in streptozotocin-diabetic rats. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1778–1790. [Google Scholar] [CrossRef]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; Macdonald, K.; Speert, D.; Fernandes-Alnemri, T.; Monks, B.G.; Wu, J.; Fitzgerald, K.A.; et al. Cutting Edge: NF-κ B Activating Pattern Recognition and Cytokine Receptors License NLRP3 Inflammasome Activation by Regulating NLRP3 Expression. J. Immunol. 2009, 187, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Wang, P.; Qi, J.; Zhang, L.; Gao, C. TLR-induced NF-κB activation regulates NLRP3 expression in murine macrophages. FEBS Lett. 2012, 586, 1022–1026. [Google Scholar] [CrossRef] [PubMed]
- Tukhvatulin, A.I.; Logunov, D.Y.; Gitlin, I.I.; Shmarov, M.M.; Kudan, P.V.; Adzhieva, C.A.C.A.; Moroz, A.F.; Kostyukova, N.N.; Burdelya, L.G.; Naroditsky, B.S.; et al. A In Vitro and In Vivo Study of the Ability of NOD1 Ligands to Activate the Transcriptional Factor NF-kB. Acta Naturae 2011, 3, 77–84. [Google Scholar] [CrossRef]
- Ting, J.P.Y.; Duncan, J.A.; Lei, Y. How the noninflammasome NLRs function in the innate immune system. Science 2010, 327, 286–290. [Google Scholar] [CrossRef]
- Fontalba, A.; Gutierrez, O.; Fernandez-Luna, J.L. NLRP2, an inhibitor of the NF-kappaB pathway, is transcriptionally activated by NF-kappaB and exhibits a nonfunctional allelic variant. J. Immunol. 2007, 179, 8519–8524. [Google Scholar] [CrossRef]
- Anand, P.K.; Malireddi, R.K.S.; Lukens, J.R.; Vogel, P.; Bertin, J.; Lamkanfi, M.; Kanneganti, T.-D. NLRP6 negatively regulates innate immunity and host defence against bacterial pathogens. Nature 2012, 488, 389–393. [Google Scholar] [CrossRef]
- Chen, G.Y.; Liu, M.; Wang, F.; Bertin, J.; Núñez, G. A functional role for Nlrp6 in intestinal inflammation and tumorigenesis. J. Immunol. 2011, 186, 7187–7194. [Google Scholar] [CrossRef]
- Schneider, M.; Zimmermann, A.G.; Roberts, R.A.; Zhang, L.; Swanson, K.V.; Wen, H.; Davis, B.K.; Allen, I.C.; Holl, E.K.; Ye, Z.; et al. The innate immune sensor NLRC3 attenuates Toll-like receptor signaling via modification of the signaling adaptor TRAF6 and transcription factor NF-κB. Nat. Immunol. 2012, 13, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Allen, I.C.; Wilson, J.E.; Schneider, M.; Lich, J.D.; Roberts, R.A.; Arthur, J.C.; Woodford, R.M.T.; Davis, B.K.; Uronis, J.M.; Herfarth, H.H.; et al. NLRP12 Suppresses Colon Inflammation and Tumorigenesis through the Negative Regulation of Noncanonical NF-κB Signaling. Immunity 2012, 36, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.B.; Bergstralh, D.T.; Duncan, J.A.; Lei, Y.; Morrison, T.E.; Zimmermann, A.G.; Accavitti-Loper, M.A.; Madden, V.J.; Sun, L.; Ye, Z.; et al. NLRX1 is a regulator of mitochondrial antiviral immunity. Nature 2008, 451, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Lupfer, C.; Kanneganti, T.-D. Unsolved Mysteries in NLR Biology. Front. Immunol. 2013, 4, 285. [Google Scholar] [CrossRef]
- Kawai, M.; Yoshikawa, T.; Nishikomori, R.; Heike, T.; Takahashi, K. Obvious optic disc swelling in a patient with cryopyrin-associated periodic syndrome. Clin. Ophthalmol. 2013, 7, 1581–1585. [Google Scholar]
- Alejandre, N.; Ruiz-Palacios, A.; García-Aparicio, A.M.; Blanco-Kelly, F.; Bermúdez, S.; Fernández-Sanz, G.; Romero, F.I.; Aróstegui, J.I.; Ayuso, C.; Jiménez-Alfaro, I.; et al. Description of a new family with cryopyrin-associated periodic syndrome: Risk of visual loss in patients bearing the R260W mutation. Rheumatol. (United Kingdom) 2014, 53, 1095–1099. [Google Scholar] [CrossRef]
- Oberg, T.J.; Vitale, A.T.; Hoffman, R.O.; Bohnsack, J.F.; Warner, J.E. Cryopyrin-associated periodic syndromes and the eye. Ocul. Immunol. Inflamm. 2013, 21, 306–309. [Google Scholar] [CrossRef]
- Mohr, S.; Xi, A.; Tang, J.; Kern, T.S. Caspase activation in retinas of diabetic and galactosemic mice and diabetic patients. Diabetes 2002, 51, 1172–1179. [Google Scholar] [CrossRef]
- El Asrar, A.M.A.; Maimone, D.; Morse, P.H.; Gregory, S.; Reder, A.T. Cytokines in the vitreous of patients with proliferative diabetic retinopathy. Am. J. Ophthalmol. 1992, 114, 731–736. [Google Scholar] [CrossRef]
- Feenstra, D.J.; Yego, E.C.; Mohr, S. Modes of Retinal Cell Death in Diabetic Retinopathy. J. Clin. Exp. Ophthalmol. 2013, 4, 298. [Google Scholar]
- Kowluru, R.A. Mitochondrial stability in diabetic retinopathy: Lessons learned from epigenetics. Diabetes 2019, 68, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Crother, T.R.; Karlin, J.; Dagvadorj, J.; Chiba, N.; Chen, S.; Ramanujan, V.K.; Wolf, A.J.; Vergnes, L.; Ojcius, D.M.; et al. Oxidized Mitochondrial DNA Activates the NLRP3 Inflammasome during Apoptosis. Immunity 2012, 36, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, D.; Hu, D.; Zhou, X.; Zhou, Y. The role of mitochondria in NLRP3 inflammasome activation. Mol. Immunol. 2018, 103, 115–124. [Google Scholar] [CrossRef]
- Singh, L.P.; Yumnamcha, T.; Swornalata Devi, T. Mitophagic Flux Deregulation, Lysosomal Destabilization and NLRP3 Inflammasome Activation in Diabetic Retinopathy: Potentials of Gene Therapy Targeting TXNIP and The Redox System. Ophthalmol. Res. reports 2018, 3, ORRT-126. [Google Scholar]
- McGuire, K.A.; Barlan, A.U.; Griffin, T.M.; Wiethoff, C.M. Adenovirus Type 5 Rupture of Lysosomes Leads to Cathepsin B-Dependent Mitochondrial Stress and Production of Reactive Oxygen Species. J. Virol. 2011, 85, 10806–10813. [Google Scholar] [CrossRef]
- Klettner, A.; Kauppinen, A.; Blasiak, J.; Roider, J.; Salminen, A.; Kaarniranta, K. Cellular and molecular mechanisms of age-related macular degeneration: From impaired autophagy to neovascularization. Int. J. Biochem. Cell Biol. 2013, 45, 1457–1467. [Google Scholar] [CrossRef]
- Nakahira, K.; Haspel, J.A.; Rathinam, V.A.K.; Lee, S.J.; Dolinay, T.; Lam, H.C.; Englert, J.A.; Rabinovitch, M.; Cernadas, M.; Kim, H.P.; et al. Autophagy proteins regulate innate immune responses by inhibiting the release of mitochondrial DNA mediated by the NALP3 inflammasome. Nat. Immunol. 2011, 12, 222–230. [Google Scholar] [CrossRef]
- Perrone, L.; Devi, T.S.; Hosoya, K.; Terasaki, T.; Singh, L.P. Thioredoxin interacting protein (TXNIP) induces inflammation through chromatin modification in retinal capillary endothelial cells under diabetic conditions. J. Cell. Physiol. 2009, 221, 262–272. [Google Scholar] [CrossRef]
- Devi, T.S.; Hosoya, K.-I.; Terasaki, T.; Singh, L.P. Critical role of TXNIP in oxidative stress, DNA damage and retinal pericyte apoptosis under high glucose: Implications for diabetic retinopathy. Exp. Cell Res. 2013, 319, 1001–1012. [Google Scholar] [CrossRef]
- Parikh, H.; Carlsson, E.; Chutkow, W.A.; Johansson, L.E.; Storgaard, H.; Poulsen, P.; Saxena, R.; Ladd, C.; Schulze, P.C.; Mazzini, M.J.; et al. TXNIP regulates peripheral glucose metabolism in humans. PLoS Med. 2007, 4, e158. [Google Scholar] [CrossRef]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-interacting protein links oxidative stress to inflammasome activation. Nat. Immunol. 2010, 11, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Trueblood, K.E.; Mohr, S.; Dubyak, G.R. Purinergic regulation of high-glucose-induced caspase-1 activation in the rat retinal Müller cell line rMC-1. Am. J. Physiol. Cell Physiol. 2011, 301, C1213–C1223. [Google Scholar] [CrossRef] [PubMed]
- Devi, T.S.; Somayajulu, M.; Kowluru, R.A.; Singh, L.P. TXNIP regulates mitophagy in retinal Müller cells under high-glucose conditions: Implications for diabetic retinopathy. Cell Death Dis. 2017, 8, e2777. [Google Scholar]
- Schulze, P.C.; Yoshioka, J.; Takahashi, T.; He, Z.; King, G.L.; Lee, R.T. Hyperglycemia promotes oxidative stress through inhibition of thioredoxin function by thioredoxin-interacting protein. J. Biol. Chem. 2004, 279, 30369–30374. [Google Scholar] [CrossRef]
- Perrone, L.; Devi, T.S.; Hosoya, K.-I.; Terasaki, T.; Singh, L.P. Inhibition of TXNIP expression in vivo blocks early pathologies of diabetic retinopathy. Cell Death Dis. 2010, 1, e65. [Google Scholar] [CrossRef]
- Rashid, K.; Akhtar-Schaefer, I.; Langmann, T. Microglia in Retinal Degeneration. Front. Immunol. 2019, 10, 1–19. [Google Scholar] [CrossRef]
- Zeng, H.Y.; Green, W.R.; Tso, M.O.M. Microglial activation in human diabetic retinopathy. Arch. Ophthalmol. 2008, 126, 227–232. [Google Scholar] [CrossRef]
- Vujosevic, S.; Bini, S.; Midena, G.; Berton, M.; Pilotto, E.; Midena, E. Hyperreflective intraretinal spots in diabetics without and with nonproliferative diabetic retinopathy: An in vivo study using spectral domain OCT. J. Diabetes Res. 2013, 2013, 491835. [Google Scholar] [CrossRef]
- Zeng, X.X.; Ng, Y.K.; Ling, E.A. Neuronal and microglial response in the retina of streptozotocin-induced diabetic rats. Vis. Neurosci. 2000, 17, 463–471. [Google Scholar] [CrossRef]
- Lim, R.R.; Hainsworth, D.P.; Mohan, R.R.; Chaurasia, S.S. Characterization of a functionally active primary microglial cell culture from the pig retina. Exp. Eye Res. 2019, 185, 107670. [Google Scholar] [CrossRef]
- Krady, J.K.; Basu, A.; Allen, C.M.; Xu, Y.; LaNoue, K.F.; Gardner, T.W.; Levison, S.W. Minocycline reduces proinflammatory cytokine expression, microglial activation, and caspase-3 activation in a rodent model of diabetic retinopathy. Diabetes 2005, 54, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Mangan, M.S.J.; Olhava, E.J.; Roush, W.R.; Seidel, H.M.; Glick, G.D.; Latz, E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat. Rev. Drug Discov. 2018, 17, 588–606. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zhang, H.; He, D.; Wang, Y.; Cai, B.; Chen, J.; Ma, J.; Liu, Z.; Wu, Y. Retinal pigment epithelium cell death is associated with NLRP3 inflammasome activation by all-trans retinal. Investig. Ophthalmol. Vis. Sci. 2019, 60, 3034–3045. [Google Scholar] [CrossRef] [PubMed]
- Yumnamcha, T.; Devi, T.S.; Singh, L.P. Auranofin Mediates Mitochondrial Dysregulation and Inflammatory Cell Death in Human Retinal Pigment Epithelial Cells: Implications of Retinal Neurodegenerative Diseases. Front. Neurosci. 2019, 13, 1065. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.R.; Grant, D.G.; Olver, T.D.; Padilla, J.; Czajkowski, A.M.; Schnurbusch, T.R.; Mohan, R.R.; Hainsworth, D.P.; Walters, E.M.; Chaurasia, S.S. Young Ossabaw Pigs Fed a Western Diet Exhibit Early Signs of Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2325–2338. [Google Scholar] [CrossRef]
- Masland, R.H. Cell populations of the retina: The Proctor lecture. Invest. Ophthalmol. Vis. Sci. 2011, 52, 4581–4591. [Google Scholar] [CrossRef]
- Masland, R.H. The fundamental plan of the retina. Nat. Neurosci. 2001, 4, 877–886. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ocular Tissues/Cells | Human | Transcript/Protein | Ref | Rat | Transcript/Protein | Ref | Mouse | Transcript/Protein | Ref | Canine | Transcript/Protein | Ref |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cornea | NLRP1 | mRNA | [68] | NLRP1 | mRNA | - | NLRP1 | mRNA | - | NLRP1 | mRNA | - |
| NLRP3 | mRNA | [68] | NLRP3 | mRNA | - | NLRP3 | mRNA | [74,78,79] | NLRP3 | mRNA | - | |
| Protein | - | Protein | - | Protein | [71,72,78] | Protein | - | |||||
| NLRC4 | Protein | - | NLRC4 | Protein | - | NLRC4 | Protein | [72] | NLRC4 | Protein | - | |
| NLRB | mRNA | - | NLRB | mRNA | - | NLRB | mRNA | [74] | NLRB | mRNA | - | |
| NOD1 | mRNA | [68] | NOD1 | mRNA | - | NOD1 | mRNA | - | NOD1 | mRNA | - | |
| NOD2 | mRNA | [68] | NOD2 | mRNA | - | NOD2 | mRNA | [102] | NOD2 | mRNA | - | |
| Protein | - | Protein | - | Protein | [102] | Protein | - | |||||
| Corneal Epithelium | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | [77] | NLRP3 | mRNA | - |
| Protein | - | Protein | Protein | [77,78,79] | Protein | - | ||||||
| NOD1 | mRNA | [67] | NOD1 | mRNA | - | NOD1 | mRNA | - | NOD1 | mRNA | - | |
| Protein | - | Protein | - | Protein | [90] | Protein | [90] | |||||
| NOD2 | mRNA | [67] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - | |
| Protein | - | Protein | - | Protein | [90] | Protein | [90] | |||||
| NLRC5 | mRNA | [67] | NLRC5 | mRNA | - | NLRC5 | mRNA | - | NLRC5 | mRNA | - | |
| NLRX1 | mRNA | [67] | NLRX1 | mRNA | - | NLRX1 | mRNA | - | NLRX1 | mRNA | - | |
| Corneal Limbal Epithelial (cell culture) | NLRP1 | mRNA | [68] | NLRP1 | mRNA | - | NLRP1 | mRNA | - | NLRP1 | mRNA | - |
| NLRP3 | mRNA | [68] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - | |
| NOD1 | mRNA | [68] | NOD1 | mRNA | - | NOD1 | mRNA | - | NOD1 | mRNA | - | |
| NOD2 | mRNA | [68] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - | |
| Corneal Epithelial (cell culture) | NLRP1 | mRNA | [67] | NLRP1 | mRNA | - | NLRP1 | mRNA | - | NLRP1 | mRNA | - |
| Protein | [67] | Protein | - | Protein | - | Protein | - | |||||
| NLRP2 | mRNA | [67] | NLRP2 | mRNA | - | NLRP2 | mRNA | - | NLRP2 | mRNA | - | |
| NLRP3 | mRNA | [67,76,77,80,81] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - | |
| Protein | [76,77,80,81] | Protein | - | Protein | - | Protein | - | |||||
| NLRP6 | mRNA | [77] | NLRP6 | mRNA | - | NLRP6 | mRNA | - | NLRP6 | mRNA | - | |
| Protein | [77] | Protein | - | Protein | - | Protein | - | |||||
| NLRP7 | mRNA | [67] | NLRP7 | mRNA | - | NLRP7 | mRNA | - | NLRP7 | mRNA | - | |
| NLRP10 | mRNA | [67] | NLRP10 | mRNA | - | NLRP10 | mRNA | - | NLRP10 | mRNA | - | |
| NOD1 | mRNA | [67,69] | NOD1 | mRNA | - | NOD1 | mRNA | - | NOD1 | mRNA | - | |
| Protein | [69] | Protein | - | Protein | - | Protein | - | |||||
| NOD2 | mRNA | [67,70] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - | |
| Protein | [70] | Protein | - | Protein | - | Protein | - | |||||
| NLRC4 | mRNA | [67] | NLRC4 | mRNA | - | NLRC4 | mRNA | - | NLRC4 | mRNA | - | |
| NLRX1 | mRNA | [67] | NLRX1 | mRNA | - | NLRX1 | mRNA | - | NLRX1 | mRNA | - | |
| Corneal Fibroblast (cell culture) | NLRP1 | mRNA | [68] | NLRP1 | mRNA | - | NLRP1 | mRNA | - | NLRP1 | mRNA | - |
| NLRP3 | mRNA | [68] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - | |
| NLRC4 | Protein | [83] | NLRC4 | Protein | - | NLRC4 | Protein | - | NLRC4 | Protein | - | |
| NOD1 | mRNA | [68] | NOD1 | mRNA | - | NOD1 | mRNA | [82] | NOD1 | mRNA | - | |
| NOD2 | mRNA | [68] | NOD2 | mRNA | - | NOD2 | mRNA | [82] | NOD2 | mRNA | ||
| Corneal Endothelium | NOD1 | Protein | - | NOD1 | Protein | - | NOD1 | Protein | [90] | NOD1 | Protein | [90] |
| NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | [90] | NOD2 | Protein | [90] | |
| Corneal Endothelial (cell culture) | NLRP1 | mRNA | [68] | NLRP1 | mRNA | - | NLRP1 | mRNA | - | NLRP1 | mRNA | - |
| NLRP3 | mRNA | [68] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - | |
| NOD1 | mRNA | [68] | NOD1 | mRNA | - | NOD1 | mRNA | - | NOD1 | mRNA | - | |
| Protein | - | Protein | - | Protein | - | Protein | - | |||||
| NOD2 | mRNA | [68] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - | |
| Conjunctiva | NLRP3 | mRNA | [86] | NLRP3 | mRNA | - | NLRP3 | mRNA | [78] | NLRP3 | mRNA | - |
| Protein | [86,87] | Protein | [87] | Protein | [78] | Protein | - | |||||
| Conjunctival Epithelium | NLRP3 | mRNA | [76,88] | NLRP3 | mRNA | - | NLRP3 | mRNA | [77] | NLRP3 | mRNA | - |
| Protein | [88] | Protein | - | Protein | [77,78] | Protein | - | |||||
| NLRP6 | mRNA | - | NLRP6 | mRNA | - | NLRP6 | mRNA | [77] | NLRP6 | mRNA | - | |
| Protein | - | Protein | - | Protein | [77] | Protein | - | |||||
| NOD1 | Protein | - | NOD1 | Protein | - | NOD1 | Protein | [90] | NOD1 | Protein | [90] | |
| NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | [90] | NOD2 | Protein | [90] | |
| Conjunctival Goblet Cell (cell culture) | NLRP3 | mRNA | [89] | NLRP3 | mRNA | [87] | NLRP3 | mRNA | - | NLRP3 | mRNA | - |
| Protein | [87,89] | Protein | [87] | Protein | - | Protein | - | |||||
| Conjunctival Substantia Propria | NOD1 | Protein | - | NOD1 | Protein | - | NOD1 | Protein | [90] | NOD1 | Protein | [90] |
| NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | [90] | |
| Iris Vascular Endothelial Cells (primary culture) | NOD2 | mRNA | [112] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - |
| Nonpigmented Ciliary Body Epithelium | NOD1 | Protein | - | NOD1 | Protein | - | NOD1 | Protein | - | NOD1 | Protein | [90] |
| NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | [90] | |
| Retina | NLRP1 | mRNA | - | NLRP1 | mRNA | [91,95,113] | NLRP1 | mRNA | [109,114] | NLRP1 | mRNA | - |
| Protein | - | Protein | [91,95] | Protein | [101,109] | Protein | - | |||||
| NLRP1b | mRNA | - | NLRP1b | mRNA | - | NLRP1b | mRNA | [96] | NLRP1b | mRNA | - | |
| NLRP3 | mRNA | - | NLRP3 | mRNA | [91,93,95,111,113,115] | NLRP3 | mRNA | [93,96,98,100,104,105,114,116,117,118] | NLRP3 | mRNA | [108] | |
| Protein | [119,120] | Protein | [91,95,107,113,121,122] | Protein | [92,94,98,99,100,101,103,110,114,116,123] | Protein | [108] | |||||
| NLRC4 | mRNA | - | NLRC4 | mRNA | [113] | NLRC4 | mRNA | - | NLRC4 | mRNA | - | |
| Retinal Microglia | NLRP3 | Protein | - | NLRP3 | Protein | [107] | NLRP3 | Protein | [97,103,118] | NLRP3 | Protein | - |
| Retinal Microglia (cell culture) | NLRP3 | mRNA | - | NLRP3 | mRNA | [111] | NLRP3 | mRNA | - | NLRP3 | mRNA | - |
| Protein | - | Protein | - | Protein | [92,124] | Protein | - | |||||
| Retinal Müller (cell culture) | NLRP3 | Protein | - | NLRP3 | Protein | [121,125] | NLRP3 | Protein | [106,126] | NLRP3 | Protein | - |
| Retina Astrocytes | NLRP3 | Protein | - | NLRP3 | Protein | - | NLRP3 | Protein | [94] | NLRP3 | Protein | - |
| Retina Ganglion Cell | NLRP1 | Protein | - | NLRP1 | Protein | - | NLRP1 | Protein | [94] | NLRP1 | Protein | - |
| NLRP3 | Protein | - | NLRP3 | Protein | - | NLRP3 | Protein | [94] | NLRP3 | Protein | - | |
| Retina Ganglion Cell (cell culture) | NLRP1 | Protein | - | NLRP1 | Protein | - | NLRP1 | Protein | [109] | NLRP1 | Protein | - |
| NLRP3 | mRNA | - | NLRP3 | mRNA | [127] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | |
| Protein | - | Protein | [127] | Protein | - | Protein | - | |||||
| Retina Microvascular Endothelial Cell (cell culture) | NLRP3 | mRNA | [120] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - |
| Protein | [113,120,128] | Protein | - | Protein | - | Protein | - | |||||
| NOD2 | mRNA | [112] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - | |
| Retina Pigmented Epithelium | NLRP3 | mRNA | [117] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - |
| Protein | [119,129,130] | Protein | [115,131] | Protein | [132,133] | Protein | ||||||
| Retina Pigmented Epithelial Cell (primary culture) | NLRP2 | mRNA | [134] | NLRP2 | mRNA | - | NLRP2 | mRNA | - | NLRP2 | mRNA | - |
| NLRP3 | mRNA | [134,135,136,137,138] | NLRP3 | mRNA | - | NLRP3 | mRNA | [119,137] | NLRP3 | mRNA | - | |
| Protein | [134,136,137] | Protein | - | Protein | [119] | Protein | - | |||||
| Retina Pigmented Epithelial Cell (ARPE-19) | NLRP3 | mRNA | [117,138,139,140,141] | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | - |
| Protein | [129,141,142,143,144,145,146,147,148,149] | Protein | - | Protein | - | Protein | - | |||||
| Choroid | NLRP3 | mRNA | - | NLRP3 | mRNA | - | NLRP3 | mRNA | [116] | NLRP3 | mRNA | - |
| Protein | [119,129] | Protein | [115] | Protein | [116] | Protein | - | |||||
| Choroid Vascular Endothelium | NOD2 | Protein | [112] | NOD2 | Protein | - | NOD2 | Protein | - | NOD2 | Protein | - |
| Choroidal Vascular Endothelial Cell (primary culture) | NOD2 | mRNA | [112] | NOD2 | mRNA | - | NOD2 | mRNA | - | NOD2 | mRNA | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, R.R.; Wieser, M.E.; Ganga, R.R.; Barathi, V.A.; Lakshminarayanan, R.; Mohan, R.R.; Hainsworth, D.P.; Chaurasia, S.S. NOD-like Receptors in the Eye: Uncovering Its Role in Diabetic Retinopathy. Int. J. Mol. Sci. 2020, 21, 899. https://doi.org/10.3390/ijms21030899
Lim RR, Wieser ME, Ganga RR, Barathi VA, Lakshminarayanan R, Mohan RR, Hainsworth DP, Chaurasia SS. NOD-like Receptors in the Eye: Uncovering Its Role in Diabetic Retinopathy. International Journal of Molecular Sciences. 2020; 21(3):899. https://doi.org/10.3390/ijms21030899
Chicago/Turabian StyleLim, Rayne R., Margaret E. Wieser, Rama R. Ganga, Veluchamy A. Barathi, Rajamani Lakshminarayanan, Rajiv R. Mohan, Dean P. Hainsworth, and Shyam S. Chaurasia. 2020. "NOD-like Receptors in the Eye: Uncovering Its Role in Diabetic Retinopathy" International Journal of Molecular Sciences 21, no. 3: 899. https://doi.org/10.3390/ijms21030899
APA StyleLim, R. R., Wieser, M. E., Ganga, R. R., Barathi, V. A., Lakshminarayanan, R., Mohan, R. R., Hainsworth, D. P., & Chaurasia, S. S. (2020). NOD-like Receptors in the Eye: Uncovering Its Role in Diabetic Retinopathy. International Journal of Molecular Sciences, 21(3), 899. https://doi.org/10.3390/ijms21030899