Abstract
The World Health Organization (WHO) herald of the “End TB Strategy” has defined goals and targets for tuberculosis prevention, care, and control to end the global tuberculosis endemic. The emergence of drug resistance and the relative dreadful consequences in treatment outcome has led to increased awareness on immunization against Mycobacterium tuberculosis (Mtb). However, the proven limited efficacy of Bacillus Calmette-Guérin (BCG), the only licensed vaccine against Mtb, has highlighted the need for alternative vaccines. In this review, we seek to give an overview of Mtb infection and failure of BCG to control it. Afterward, we focus on the protein- and peptide-based subunit vaccine subtype, examining the advantages and drawbacks of using this design approach. Finally, we explore the features of subunit vaccine candidates currently in pre-clinical and clinical evaluation, including the antigen repertoire, the exploited adjuvanted delivery systems, as well as the spawned immune response.
1. Introduction
According to the WHO’s Report of 2020, around 10 million people fell ill with tuberculosis (TB) each year, and 1.5 million died only in 2018, making it the leading cause of death from a single infectious agent [1].
TB represents an urgent health priority, especially in high-burden countries, where 90% of cases are registered. Most of these countries are low income and developing states in which TB has severe economic, social, and public health consequences. The epidemiological scenario in these countries is worsened by health-related risk factors, like the correlation with HIV in TB-areas, likewise by broader determinants, such as undernutrition and poverty [1]. Additionally, deep concern is currently emerging about the COVID-19 outbreak impingement on TB handling due to the limited access to treatment and diagnosis, as well as the partial overlap of disease symptoms [2]. Despite drug treatments continue to improve, reports of multidrug resistance tuberculosis (MDR-TB) and extensively drug-resistance tuberculosis (XDR-TB) are increasing worldwide, reducing the treatment success rate at unacceptably low levels. The latest available data show an 85% success rate for drug-susceptible TB, 56% for MDR-TB, and 39% for XDR-TB [3]. Nevertheless, the estimated treating cost for MDR-TB ranges from eight-fold to 15-fold higher compared to sensitive-TB, while it can rise to 32-fold higher in XDR-TB cases [1,4,5].
2. BCG: A Cornerstone in TB Handling
BCG vaccine is an attenuated form of Mycobacterium bovis, the primary cause of bovine tuberculosis, which is genetically related to Mycobacterium tuberculosis (Mtb). Used for the first time in humans in 1921, it still represents the gold standard in TB vaccination, although several aspects of this vaccine remain incompletely characterized, including its overall efficacy, duration of protective immunity, and how age at vaccination affects the protection [6,7].
The role of BCG in preventing severe forms of extra-pulmonary TB in children, like TB meningitis and miliary TB, has been proved [8,9]. Conversely, a limited protective effect was observed against pulmonary TB, especially in adults, which is not only the most prevalent form of the disease but also the principal source of transmission [10]. Epidemiological evidence highlighted that the BCG degree of protection varies geographically [6,7,8,11,12,13,14,15], while several studies suggested a lower and short-lasting BCG protection in people previously sensitized with Mtb and non-tuberculous mycobacteria (NTB) [7,11]. Additionally, it has an unfavorable risk profile in immunocompromised conditions, for example, congenital immunodeficiencies and HIV infections, because of BCG bacteremia that follows vaccination [9,13]. Commonly, BCG has been shown to protect infants and young children up to approximately 10 years of age, although some degree of protection might last for up to 20 years following school-aged BCG vaccination and as long as 50–60 years following infant vaccination [8,9,15]. However, it cannot provide lifelong immunity, and there are no studies that currently demonstrate a reliable effect against TB of BCG revaccination in adolescents [8]. Nevertheless, a Phase II clinical trial is ongoing to demonstrate the efficacy of revaccination against sustained Mtb infection in previously BCG vaccinated people (NCT04152161).
Given the scale of the global TB epidemic and the slow pace of TB control efforts, it is clear that a novel and more effective TB vaccine is needed to achieve WHO End TB Strategy targets by reducing TB deaths by 95% and new cases of TB by 90% by 2035 [1].
3. Tuberculosis Immunopathology
Mtb infection is a complex, multi-stage process that generally arises after the inhalation of Mtb-containing aerosolized droplets released from people with active pulmonary TB (Figure 1). The respiratory tract and bronchoalveolar spaces represent a unique immunological compartment where the innate immune system and the respiratory epithelium act as the first barrier to providing early clearance of the pathogen [11,16,17,18,19]. Following initial exposure to Mtb, a small percentage of exposed humans can quickly clear the infection. Besides, a growing number of studies evidence that a subset of people does not show immunodiagnostic relevance of Mtb sensitization after exposure. Even though this phenomenon is subject to several potential clinical and immunological interpretations, some insights suggest the involvement of the innate immune response [8,19,20,21]. Generally, innate immunity fails to clear the infection in the majority of exposed humans, and Mtb dominates the initial stage of infection, taking advantage of phagocytes recruitment to infection sites for its expansion [22,23]. The bacteria can survive and replicate in infected cells via several immunosuppressive mechanisms, including inhibition of phagosome maturation, phagosome–lysosome fusion, production of reactive oxygen species, hamper major histocompatibility (MHC) class-II antigen presentation [21]. Additionally, Mtb manipulates death modalities of infected cells to improve cell-to-cell spreading [18,23]. These and other virulence factors contribute to the severe innate immune suppression in the lung and lead to the slowed-down recruitment and trafficking of dendritic cells (DC) to the drain lymph nodes. This peculiarity of Mtb infection results in a delayed onset of the adaptive immune response [24]. Moreover, DC migration from the primary site of infection to the bloodstream causes the colonization of previously uninfected lobes of the lung and the establishment of extra-pulmonary disease [18].
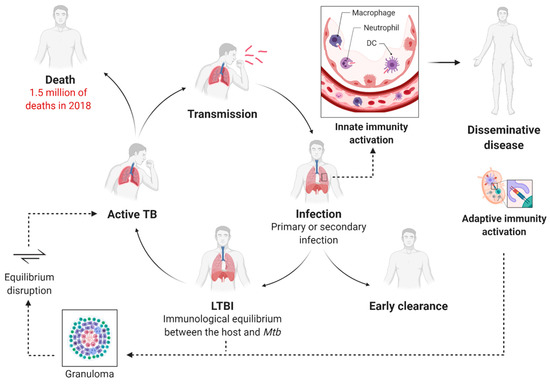
Figure 1.
The immunological lifecycle of Mycobacterium tuberculosis and the progression of the disease. Mtb is commonly aerosol transmitted, and the innate immune response acts as a first barrier in the alveolar space. At the early stage of infection, Mtb may be rapidly eradicated. The trafficking of the antigen-presenting cells to the lymph nodes leads to the dissemination of the disease as well as to the activation of the adaptive immune response. T-lymphocytes regulate the formation of the granuloma within Mtb is contained, and the individual develops latent TB infection (LTBI). When the immunological equilibrium between the host and the pathogen is disrupted, latent TB evolves to active disease. At this stage, the patient becomes symptomatic and can transmit the infection to healthy individuals.
T-lymphocytes stimulated in the draining lymph nodes mediate the formation of the granulomas, primarily composed of macrophages, DCs, and T- and B-lymphocytes. Mtb containment within these lesions results in the development of latent TB infection (LTBI) [10]. In this stage, the individual becomes asymptomatic and cannot transmit the disease to healthy people. In the LTBI stage, a dynamic immunological equilibrium is established between the host and the pathogen. Mtb is prevalently in the dormant state, and its activities are markedly downregulated, while a subpopulation of bacteria is metabolically active and continues to replicate [18]. Hypothetically, the expression of a modified antigen repertoire causes the physiological shift of the bacteria into a non-replicative phenotype, as confirmed by human studies showing that the tubercular antigens recognized by healthy individuals with latent TB are different from those by active TB patients [25].
Approximately in 10% of the cases, reactivation of latent TB can progress to active, symptomatic disease [10]. This phenomenon is widely attributed to ‘weakened’ immunity, and that is the main reason for the emerging syndemic interaction between HIV and TB epidemics [26]. HIV-associated TB contributes substantially to the number of deaths, and it is estimated that people living with HIV are 15–22 times more likely to develop active TB than people without HIV [27].
4. Strategic Goals for New TB Vaccine Development
4.1. Target Population
One of the main challenges in developing a new TB vaccine is understanding who should be vaccinated and when. Therefore, host infection status required for efficacy (pre- or post-infection) and the desired effect type (prevention of infection or disease) represent essential factors to take into consideration in the design of a new vaccine [28]. Considering that the risk of Mtb infection and progression to TB disease increases with age, the WHO has proposed two sets of preferred product characteristics to guide vaccine candidate’s development, one addressing adolescents and adults, the other targeting neonates. Despite the development of a more effective vaccine than BCG would be desirable, it is expected that infant-targeted vaccination would require many years to become a cost-effective measure unless lifelong protection and high efficacy would be observed [9,29]. Children show lower rates of TB notifications, lower proportions of smear-positive pulmonary TB, and, consequently, they exhibited a smaller contribution to TB transmission [29]. Therefore, adolescents and adults should be defined as a priority strategic target as they represent the leading source of Mtb dissemination [9,30,31].
Novel TB vaccine candidates that enter clinical trials (Figure 2) can be categorized as follows: (i) preventive vaccines that are administered before Mtb infection—typically to neonates; (ii) booster vaccines, administered after previous BCG vaccination—typically to adolescents and adults with LTBI; (iii) post-exposure therapeutic vaccines drugs—to adults with to treat TB in adjunct with drug therapy [32].
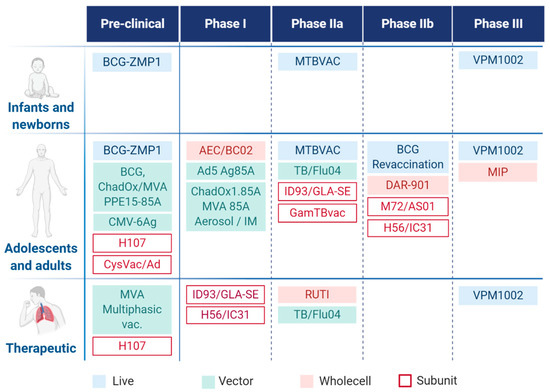
Figure 2.
Tuberculosis vaccine pipeline with target population.
4.2. Tuberculosis Vaccine Pipeline
Currently, 15 vaccine candidates are being investigated in active clinical trials (Figure 2), and additional candidates are tested in the pre-clinical phase. MVA85A (modified vaccine Ankara 85A) was the first booster vaccine candidate to enter vaccine efficacy evaluation since the introduction of BCG [33]. Unfortunately, the Phase IIb trial with previously BCG vaccinated healthy infants ended up with undesired results because the difference between the vaccinated and the placebo groups was not significant, and the interpretation was the absence of efficacy of MVA85. However, results were very informative to the scientific community, leading to a return to basic discovery with the primary goal to explore a greater diversity of approaches and develop better efficacy markers as well as improved in vitro and in vivo model systems [34,35].
In the recent vaccine pipeline, three live attenuated, four whole-cell or fragmented, three viral vectored, and five subunit-type vaccines are studied, of which six candidates enter the clinical IIb and III phases [36].
4.3. Subunit Vaccines against Tuberculosis
Traditionally, cell-based vaccines have been in the spotlight of vaccine development strategies because they generally produce long-lasting immunity. Nonetheless, they display several drawbacks, such as manufacturing difficulties, high feasibility costs, and critical stability. However, the main concern regards their safety as they may cause autoimmune or allergic reactions. Moreover, attenuation or inactivation of such vaccines might not be perfect, leading to the pathogen’s return to its virulent state, as it happened in the “Lübeck disaster” where 72 BCG-vaccinated children out of 252 died after developing pulmonary TB.
However, subunit vaccines, which consist primarily of proteins or peptides, can face limitations concerning immunogenicity. Therefore, they require immunostimulatory adjuvant or delivery systems to enhance the immunological response [37].
4.3.1. Protein Antigens and Immunological Response
The development of an efficient vaccine requires not only accurate antigens choice but also activating the right ratio of a protective and suppressive immune response against the selected pathogen.
Even if intensive efforts have been underway, the identification of well-defined protective antigens has proved to be a challenging goal in TB subunit vaccine development.
While Mtb is a complex microorganism, composed of approximately 4000 genes, the selection of a small number of antigens among them is complicated because of the limited knowledge about the function of several antigens and their differential expression at different stages of the pathogen life cycle [31,38]. Moreover, Mtb escape mechanisms, as the downregulation of antigen processing and presentation, have made the identification procedure more difficult, resulting in the current lack of stringent immunological markers as correlates of protection [38,39].
The protein adjuvanted subunit vaccines that are currently under clinical evaluation use 12 mycobacterial antigens in different combinations and formulations (see Table 1).

Table 1.
Mycobacterial antigens used in subunit-type vaccines being under clinical investigations.
Antigens involved in the active bacteria replication, such as Ag85, ESAT6, and CFP10, are extensively used in TB vaccination as they have proved to be highly immunogenic and showed protection in different animal models [41]. EsxV, EsxW, and TB10.4 are members of the Esx family and they are associated with the virulence of the bacteria as well. PPE18 and PPE42 belong to the so-called PPE family because of the presence of conserved Pro-Pro-Glu (PPE) motifs at the N-termini. As revealed by genome sequence analysis, genes encoding PPE proteins are immediately upstream of the Esx gene family, and they seem to play a role in driving localization or secretion of different proteins, such as ESAT6 [42,43]. On the other hand, Rv1813 and Rv2660 are stress-induced antigens associated with the dormant state of the pathogen. While the exact function of PepA, also called MTB32A antigen, is still unknown, although it seems to be a putative serine protease.
Several studies highlighted that these antigens are sharing a common characteristic, immunodominance. The immunogenicity of a protein antigen is heterogeneous; hence, some regions can elicit a more efficient expansion of T-cells than others can.
The current TB vaccine pipeline suffers the shortcoming of being focused on a narrow set of antigens characterized by suboptimal activity. Moreover, clinical trial data complicate to clarify the real effectiveness of these antigens’ subset and whether their intrinsic features represent an advantage for a TB vaccine candidate [7,38,41]. Therefore, the current strategy suggests to broad antigen repertoire, involving a more diversified array of antigens to improve the outcome.
The majority of vaccine candidates in the TB pipeline were selected based on their ability to induce interferon-gamma producing T-cells. However, while the protective role of CD4+ T-cells against Mtb is well-established; it seems evident that focusing on this classical T-cells subtype response is essential, but maybe not sufficient to obtain a fully protective response [8,38,39,44]. Notable attempts have focused upon identifying antigens capable of stimulating CD8+ T-cell mediated responses since Mtb-specific CD8+ T-cells have been detected in individuals with TB and LTBI as well [45].
Recently, the potential role of Donor Unrestricted T-cells (DURTs) is subject to increasing interest; it is a subset of T-cells that interact with human antigen-presenting cells (APCs) through non-MHC antigens presenting systems [8,46]. These non-conventional T-cells can recognize non-protein antigens, such as lipids and glycolipids that are part of the Mtb lipidome, which could be immunogenic to humans and may act as a booster of the immune response against the pathogen. Moreover, the multi-epitope approach is catching the eye of the scientific world as a promising strategy against different diseases, from oncology to infections, because it can expand the targeting spectra [47]. For example, it is possible to target several strains, different stages of the life cycle, or even distinct pathogens. The concept of targeting both the latent and the active state of the pathogen is not a novelty in TB vaccine design. Subunit vaccines ID93/GLA-SE and H56:IC31 are both candidates under clinical evaluation characterized by the presence of antigens associated with the virulence of the bacteria and the latency as well.
4.3.2. Adjuvants and Formulation Strategies for Subunit Vaccines
As previously mentioned, peptide- or protein-based vaccination is not free from challenges to overcome, such as low immunogenicity and bioavailability limitations. For this reason, a suitable formulation needs to be accurately designed. Nowadays, adjuvant and delivery systems are no longer mutually exclusive components in the field of vaccine development. Both of them can have the potential to adequately stimulate an immune response while simultaneously protecting the antigen degradation and transporting it to the desired tissue [48]. Indeed, most adjuvants effective in TB animal models, including the ones in clinical development, rely on vaccine adjuvant-delivery system (VADS) (Figure 3).
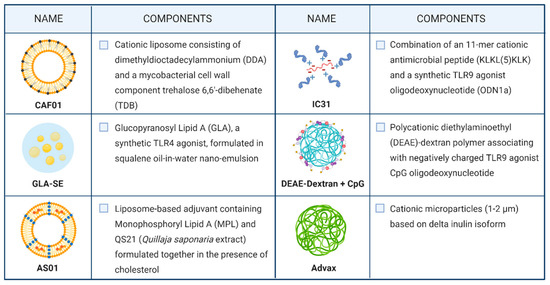
Figure 3.
Vaccine adjuvant delivery system used in candidates undergoing clinical trials.
For decades alum was the only adjuvant used in licensed human vaccines, and it still represents a benchmark in this field. However, it has been considered unsuitable for vaccines against intracellular pathogens, like Mtb, because of its low ability to induce Th1-cellular immunity as well as CD8+ cytotoxic responses [49]. On the other hand, recent studies suggest that the modification of alum physicochemical properties, including the reduction of the particle size to the nanometer scale, can induce cellular immune responses characterized by Th1-cytokine secreting CD4+ T-cells [50,51].
Currently, the use of liposomes and emulsions as VADS is a prominent strategy used in TB vaccine formulation; examples include AS01, CAF01, and GLA-SE.
CAF01 is the first member of the Cationic Adjuvant Formulation (CAF) series developed by the Statens Serum Institut. This two-component liposomal adjuvant system is composed of a cationic liposome vehicle (N,N-dimethyl-N,N-dioctadecylammonium (DDA)) stabilized with a glycolipid immunomodulator (trehalose 6,6′-dibehenate (TDB)). While DDA alone can promote antibody-mediated and cellular immunological responses, it has shown intrinsic physical instability. Therefore, TDB incorporation act as a stabilizer of the system by enabling hydrogen bonding between the liposomal membrane and the surrounding water. Accordingly, CAF01 has been demonstrated as a highly stable formulation with an attested shelf life at 4 °C for more than two years [52]. From an immunological point of view, TDB is characterized by intrinsic immunostimulatory properties, as it is a synthetic analog of the cord factor (trehalose 6,6′-dibehenate (TDB)) located in the mycobacterial cell wall. Thus, it enhances the cellular immune response induced by DDA liposomes [53]. CAF01 has proved to promote a long-lived immunity with a Th1 profile in animal models and to elicit a Th17 response due to TDB signaling through the C-type lectin receptor Mincle. A recently conducted study on subcutaneous immunization with H56:CAF01 followed by an intranasal boosting did not skew the Th1/17 profile established by parenteral vaccine administration and did not confer additional control of pulmonary TB after aerosol Mtb-challenge [54]. However, a phase I clinical trial aimed to evaluate the CAF01 safety profile highlighted no concerns associated with the administration of the CAF01-adjuvanted vaccine to healthy adults (NCT00922363) [55].
Glucopyranosyl Lipid Adjuvant (GLA), a synthetic derivative of Monophosphoryl Lipid A (MPL) with TLR4 agonistic action, is formulated in a stable oil-in-water squalene emulsion (SE) to conceive GLA-SE. GLA is a fully synthetic derivative of lipopolysaccharide (LPS) that maintains mostly the immunostimulatory activity of the original molecule while it displays considerably reduced toxicity [56]. The combination of GLA-SE adjuvant and the fusion protein ID93 is currently under ongoing phase IIa clinical trial as a TB vaccine candidate, wherein its safety, immunogenicity, and efficacy are evaluated in previously BCG-vaccinated healthy healthcare workers. Phase I clinical trials previously conducted on healthy adult individuals highlighted the good tolerability of the adjuvant [57,58]. Furthermore, the inclusion of GLA-SE led to higher T-cell response magnitudes and to improve the overall T-cell response quality by eliciting a significantly higher proportion of polyfunctional CD4+ T-cells, producing TNF+, IL-2+, IFN-γ cytokines. Additionally, an increased antibody production, prevalently of IgG1 and IgG3 subclasses, was observed after ID93:GLA-SE vaccination [58].
M72:AS01E, one of the most advanced subunit vaccines in the pipeline was recently evaluated in a phase IIb clinical trial (NCT01755598), showing 54% efficacy in HIV-negative individuals with latent TB when administered intramuscularly in emulsion form [59]. Furthermore, the study evidenced that M72:AS01E vaccination protects against progression to pulmonary tuberculosis disease for at least three years [60]. AS01 is a liposome-based adjuvant that contains two immunostimulants, MPL and QS-21, formulated together in the presence of cholesterol to abrogate the haemolytic activity of QS-21. As mentioned above, MPL is a well-established pro-inflammatory adjuvant that directly activates APCs expressing TLR4 and stimulating NF-ĸB transcriptional activity. QS-21 is a natural saponin composed of a purified fraction of two isomeric triterpene glycosides from the tree Quillaja saponaria. While the exact mechanism of action of QS-21 has not been fully elucidated, the latest hypothesis suggested its action on both T-cells and APCs to induce a Th1 immune response [61]. The mechanism of action studies on QS-21 suggested the aldehyde group as the principal mediator of T-cell interaction and further activation, whereas QS-21 forms complexes with cholesterol to intercalate on the cell membrane of APCs. Moreover, it allows the early endosomal escape of the antigens for further processing inside the cell that results in peptides presented on the DCs surface to naïve CD8+ T-cells to yield cytotoxic T-lymphocytes [61].
A different approach was used in the development of the two-component adjuvant IC31, currently under clinical evaluation in combination with the fusion protein H56. IC31 is composed of the synthetic antimicrobial positively charged peptide KLK (KLKL5KLK) and ODN1a, a phosphodiester-backboned DNA oligonucleotide consisting of repeats of the dinucleotides deoxyinosine and deoxycytosine (poly I:C). KLK and ODN1a are formulated in a stoichiometrically fixed molar ratio of 25:1, respectively. While KLK is a poor adjuvant itself, its poly-cationic structure is suggested to play a crucial role in the delivery of ODN1a and the antigens to target APCs. It also induces depot formation after subcutaneous immunization, essential to provide constant antigen-specific stimulation of immune responses over a long period. ODN1a is an immunostimulatory molecule, acting through the TLR9/MyD88 pathway to promote Th1 biased immune responses [62]. H56:IC31 is currently under clinical evaluation both as a vaccine for adults and adolescents and as a therapeutic vaccine. However, the vaccine showed acceptable safety and tolerability profile in Mtb-infected and Mtb-uninfected adults as well as durable antigen-specific CD4+ T-cell responses [63,64].
Recently, polysaccharides are gaining attention as adjuvants because of their biocompatibility, biodegradability. Dextran is one of the most investigated α-glucan in drug and antigen delivery and has a long history of medical use because generally considered safe.
GamTBVac is a subunit vaccine under clinical development wherein a fusion protein with a dextran-binding domain is combined with a novel dextran/CpG adjuvant. The mixture results in nanoparticles consisting of DEAE-dextran core covered with a mixture of TLR9 agonist CpG through relatively week electrostatic interaction. The safety and immunogenicity of GamTBvac have been evaluated in a recently completed phase IIa trial in healthy BCG-vaccinated adults (NCT03878004), following successful clinical evaluation in a phase I trial in Russia [65].
Advax is a novel plant-derived adjuvant based on inulin delta isoform with the ability to form cationic particles of approximately 1–2 µm in diameter. Advax-based adjuvants have previously been proved to induce protective immunity against several pathogens in a wide range of animal species. Furthermore, this adjuvant is well-tolerated and immunogenic in human subjects. Currently, the use of Advax-based adjuvants in TB vaccine design is under pre-clinical evaluation. Recently, the Advax-based formulation of CysVac2 fusion protein has shown to confer protection against aerosol challenge at a comparable level to other TB vaccine candidates that have entered clinical trials, such as ID93/GLA-SE and M72/AS02. Moreover, the addition of CpG oligonucleotide, a TLR9 agonist, has resulted in enhanced immune cell recruitment and the subsequent multifunctional effector T-cell response [66,67].
4.3.3. Peptide-Based Subunit Vaccines
Conversely, the peptide-based vaccine approach has become an attractive strategy in vaccine development because its manufacture is simple, fast, safe, reproducible, and cost-effective. Peptides are chemically well-defined compounds with good stability, and they do not rely on refrigerated storage. Overcoming the cold chain dependency is another advantage of the use of peptide-based vaccines. Besides, customizable design results in a reduced risk of harmful responses and side effects [48,68,69].
Numerous peptide-based vaccine candidates can be found in advanced clinical development targeting (i) infectious agents such as the highly polymorphic malaria, influenza virus, Hepatitis C virus, and HIV; (ii) Alzheimer’s disease where immunization with peptides could result in the clearance of neurotoxic forms of β-amyloid and Tau proteins and (iii) tumor cells [37]. The immuno-oncology approach to fighting against cancer and the use of peptides to provoke a sufficient T-cell response against tumor cells became one of the most promising measures of the decade [70,71,72].
While seven protein-based candidates are present in the TB vaccine pipeline [73], peptide-based vaccine candidates are being investigated only in pre-clinical studies yet (Table 2). For example, a synthetic long-peptide (SLP) derived from immunodominant proteins of Mtb showed protection in a murine model of tuberculosis [74]. Also, the immunization with self-adjuvating lipopeptide vaccine candidates is attracting more and more attention [75]. For example, Agrewala [76] focused on a self-adjuvating synthetic vaccine construct (L91). They recently proved that the conjugate is immunogenic in the murine and guinea pig models of TB and conferred better protection than BCG against Mtb [77]. They also showed that the combination of drug therapy and L91 (D-L91) significantly declined the bacterial load in Mtb infected animals [78]. Lately, self-assembled peptide nanofibers have received interest as vaccines and immunotherapies for several applications, including infectious diseases. A recently published study highlighted that peptide nanofiber vaccines in combination with BCG reduce bacterial load in the lungs of Mtb-infected mice and thus may be an effective strategy for boosting BCG-primed individuals [79]. Moreover, some studies have suggested that developing subunit vaccines that include classical T-cell epitopes derived from proteins that are expressed by the active and latent form of the causative agent may represent a new frontier in tuberculosis vaccine research [80,81]. In this context, several supportive tools are used to predict the most suitable epitopes to involve in the vaccine design, including immunoinformatic, genomic, and proteomic approaches [39,82,83,84].
Table 2.
A selection of peptide-based subunit vaccines under pre-clinical investigation.
5. Conclusions
Over the last decade, the TB vaccine pipeline has significantly progressed. Despite the advances made, several questions are still unsolved, such as the deeper understanding of Mtb pathogenesis and resistance strategies, as well as the immunological profile of BCG.
Despite BCG being developed over a century ago, there is still a lack in the full comprehension of its mechanism of action in preventing TB, as well as its off-target effect linked to protection against other infectious diseases. For example, increasing interest has been placed in its potential role in improving COVID-19 outcomes [90,91]. Though WHO officially disclaim the hypothesis of BCG protective effect against SARS-CoV-2, several surveys suggested a higher propensity of countries with no BCG vaccination coverage to be severely affected by the virus. On the other hand, these findings cannot provide definitive proof of causality because of some intrinsic bias, such as variances in the demographic and genetic structure of each country population, differences in non-pharmaceutical measures adopted (like quarantine and social distancing), and in diagnosis and reporting COVID-19 cases as well. Therefore, randomized controlled trials using BCG are under evaluation to endow the highest quality proof for the hypothesis that BCG may protect against COVID-19 [92,93,94,95]. Three clinical trials have been registered this year to evaluate the protective role of BCG vaccination against SARS-CoV-2 infection [96,97,98].
While primary TB has been extensively studied in humans and animals, post-primary TB is seldom recognized or studied. Nowadays, we have greater ability to manipulate vaccines to induce desired immune responses, including the adoption of the vaccine technology, the choice of the antigen and the adjuvant as well as the preferred route of administration.
The main concern is related to what type of immune response should be induced. Clinical trials are essential to answer this question as we are suffering from the lack of good biomarkers of protective immunity for adult pulmonary TB [99]. Moreover, we experienced disagreement in between animal TB models and between different human TB vaccine studies. This could reflect that a correlate of protection may be specific for each vaccine platform or may vary for different TB antigens. In this regard, standardization of vaccine study protocols and harmonization of animal and human studies could improve the discovery of the protection correlates [100].
On the other hand, the increasing number of subunit vaccines in the clinical phase has evidenced the proof of concept that protein-based antigens empowered by an adequate adjuvant can represent a promising immunization strategy against Mtb, but further studies are required to broaden the antigen repertoire, diversify the immune response, and find suitable adjuvant systems to develop a vaccine efficiently preventive TB infection, especially now that the outbreak of MDR-TB has made new struggles to control TB prevalence.
Author Contributions
Writing—Original Draft Preparation and Visualization, C.B.; Conceptualization, Writing—Review & Editing, K.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been funded by the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement no. 860325 (BactiVax), by the ELTE Thematic Excellence Programme (Szint+) supported by the Hungarian Ministry for Innovation and Technology, and by the National Research, Development and Innovation Fund of Hungary, financed under the 2018-1.2.1-NKP funding scheme (2018-1.2.1-NKP-2018-00005).
Acknowledgments
Kata Horváti thanks for the support of the Premium Post-Doctoral Fellowship of the Hungarian Academy of Sciences. The artwork was created with BioRender.
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization. Global Tuberculosis Report 2020; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Manyazewal, T.; Woldeamanuel, Y.; Blumberg, H.M.; Fekadu, A.; Marconi, V.C. The fight to end tuberculosis must not be forgotten in the COVID-19 outbreak. Nat. Med. 2020, 26, 811–812. [Google Scholar] [CrossRef] [PubMed]
- Dheda, K.; Gumbo, T.; Gandhi, N.R.; Murray, M.; Theron, G.; Udwadia, Z.; Migliori, G.B.; Warren, R. Global control of tuberculosis: From extensively drug-resistant to untreatable tuberculosis. Lancet Respir. Med. 2014, 2, 321–338. [Google Scholar] [CrossRef]
- Oh, P.; Pascopella, L.; Barry, P.M.; Flood, J.M. A systematic synthesis of direct costs to treat and manage tuberculosis disease applied to California. BMC Res. Notes 2017, 10, 434. [Google Scholar] [CrossRef] [PubMed]
- Diel, R.; Vandeputte, J.; De Vries, G.; Stillo, J.; Wanlin, M.; Nienhaus, A. Costs of tuberculosis disease in the European Union: A systematic analysis and cost calculation. Eur. Respir. J. 2014, 43, 554–565. [Google Scholar] [CrossRef]
- Angelidou, A.; Diray-Arce, J.; Conti, M.G.; Smolen, K.K.; van Haren, S.D.; Dowling, D.J.; Husson, R.N.; Levy, O. BCG as a Case Study for Precision Vaccine Development: Lessons from Vaccine Heterogeneity, Trained Immunity, and Immune Ontogeny. Front. Microbiol. 2020, 11, 332. [Google Scholar] [CrossRef]
- Li, J.; Zhao, A.; Tang, J.; Wang, G.; Shi, Y.; Zhan, L.; Qin, C. Tuberculosis vaccine development: From classic to clinical candidates. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1405–1425. [Google Scholar] [CrossRef]
- Schrager, L.K.; Vekemens, J.; Drager, N.; Lewinsohn, D.M.; Olesen, O.F. The status of tuberculosis vaccine development. Lancet Infect. Dis. 2020, 20, e28–e37. [Google Scholar] [CrossRef]
- Hatherill, M.; White, R.G.; Hawn, T.R. Clinical Development of New TB Vaccines: Recent Advances and Next Steps. Front. Microbiol. 2020, 10, 3154. [Google Scholar] [CrossRef]
- Kaufmann, S.H.E. Vaccination against Tuberculosis: Revamping BCG by Molecular Genetics Guided by Immunology. Front. Immunol. 2020, 11, 316. [Google Scholar] [CrossRef]
- Brazier, B.; McShane, H. Towards new TB vaccines. Semin. Immunopathol. 2020, 42, 315–331. [Google Scholar] [CrossRef]
- Mangtani, P.; Abubakar, I.; Ariti, C.; Beynon, R.; Pimpin, L.; Fine, P.E.; Rodrigues, L.C.; Smith, P.G.; Lipman, M.; Whiting, P.F.; et al. Protection by BCG vaccine against tuberculosis: A systematic review of randomized controlled trials. Clin. Infect. Dis. 2014, 58, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Scriba, T.J.; Mizrahi, V. Renewing the Fight against TB with an Old Vaccine. Cell 2020, 180, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Aronson, N.E.; Santosham, M.; Comstock, G.W.; Howard, R.S.; Moulton, L.H.; Rhoades, E.R.; Harrison, L.H. Long-term Efficacy of BCG Vaccine in American Indians and Alaska Natives: A 60-Year Follow-up Study. JAMA 2004, 291, 2086–2091. [Google Scholar] [CrossRef] [PubMed]
- Mangtani, P.; Nguipdop-Djomo, P.; Keogh, R.H.; Sterne, J.; Abubakar, I.; Smith, P.G.; Fine, P.; Vynnycky, E.; Watson, J.M.; Elliman, D.; et al. The duration of protection of school-aged BCG vaccination in England: A population-based case-control study. Int. J. Epidemiol. 2018, 47, 193–201. [Google Scholar] [CrossRef]
- Schwander, S.; Dheda, K. Human lung immunity against Mycobacterium tuberculosis: Insights into pathogenesis and protection. Am. J. Respir. Crit. Care Med. 2011, 183, 696–707. [Google Scholar] [CrossRef]
- Ferguson, J.S.; Schlesinger, L.S. Pulmonary surfactant in innate immunity and the pathogenesis of tuberculosis. Tuber. Lung Dis. 2000, 80, 173–184. [Google Scholar] [CrossRef]
- Delogu, G.; Provvedi, R.; Sali, M.; Manganelli, R. Mycobacterium tuberculosis virulence: Insights and impact on vaccine development. Future Microbiol. 2015, 10, 1177–1194. [Google Scholar] [CrossRef]
- Schorey, J.S.; Schlesinger, L.S. Innate Immune Responses to Tuberculosis. Microbiol. Spectr. 2016, 4, TBTB2-0010-2016. [Google Scholar] [CrossRef]
- Verrall, A.J.; Netea, M.G.; Alisjahbana, B.; Hill, P.C.; van Crevel, R. Early clearance of Mycobacterium tuberculosis: A new frontier in prevention. Immunology 2014, 141, 506–513. [Google Scholar] [CrossRef]
- Jeyanathan, M.; Yao, Y.; Afkhami, S.; Smaill, F.; Xing, Z. New Tuberculosis Vaccine Strategies: Taking Aim at Un-Natural Immunity. Trends Immunol. 2018, 39, 419–433. [Google Scholar] [CrossRef]
- de Martino, M.; Lodi, L.; Galli, L.; Chiappini, E. Immune Response to Mycobacterium tuberculosis: A Narrative Review. Front. Pediatr. 2019, 7, 350. [Google Scholar] [CrossRef] [PubMed]
- Ernst, J.D. The immunological life cycle of tuberculosis. Nat. Rev. Immunol. 2012, 12, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Jeyanathan, M.; McCormick, S.; Lai, R.; Afkhami, S.; Shaler, C.R.; Horvath, C.N.; Damjanovic, D.; Zganiacz, A.; Barra, N.; Ashkar, A.; et al. Pulmonary M. tuberculosis infection delays Th1 immunity via immunoadaptor DAP12-regulated IRAK-M and IL-10 expression in antigenpresenting cells. Mucosal Immunol. 2014, 7, 670–683. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Oe, T.; Kawakami, K.; Osada-Oka, M.; Ozeki, Y.; Terahara, K.; Yasuda, I.; Edwards, T.; Tanaka, T.; Tsunetsugu-Yokota, Y.; et al. CD4+ T Responses Other Than Th1 Type Are Preferentially Induced by Latency-Associated Antigens in the State of Latent Mycobacterium tuberculosis Infection. Front. Immunol. 2019, 10, 2807. [Google Scholar] [CrossRef]
- Kwan, C.; Ernst, J.D. HIV and tuberculosis: A deadly human syndemic. Clin. Microbiol. Rev. 2011, 24, 351–376. [Google Scholar] [CrossRef]
- Bares, S.H.; Swindells, S. Latent Tuberculosis and HIV Infection. Curr. Infect. Dis. Rep. 2020, 22, 17. [Google Scholar] [CrossRef]
- Harris, R.C.; Sumner, T.; Knight, G.M.; White, R.G. Systematic review of mathematical models exploring the epidemiological impact of future TB vaccines. Hum. Vaccin. Immunother. 2016, 12, 2813–2832. [Google Scholar] [CrossRef]
- Knight, G.M.; Griffiths, U.K.; Sumner, T.; Laurence, Y.V.; Gheorghe, A.; Vassall, A.; Glaziou, P.; White, R.G. Impact and cost-effectiveness of new tuberculosis vaccines in low- and middle-income countries. Proc. Natl. Acad. Sci. USA 2014, 111, 15520–15525. [Google Scholar] [CrossRef]
- World Health Organization. WHO Preferred Product Characteristics for New Tuberculosis Vaccines; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Schrager, L.K.; Harris, R.C.; Vekemans, J. Research and development of new tuberculosis vaccines: A review. F1000Research 2019, 7, 1732. [Google Scholar] [CrossRef]
- Kaufmann, S.H.E.; Weiner, J.; von Reyn, C.F. Novel approaches to tuberculosis vaccine development. Int. J. Infect. Dis. 2017, 56, 263–267. [Google Scholar] [CrossRef]
- Tameris, M.D.; Hatherill, M.; Landry, B.S.; Scriba, T.J.; Snowden, M.A.; Lockhart, S.; Shea, J.E.; McClain, J.B.; Hussey, G.D.; Hanekom, W.A.; et al. Safety and efficacy of MVA85A, a new tuberculosis vaccine, in infants previously vaccinated with BCG: A randomised, placebo-controlled phase 2b trial. Lancet 2013, 81, 1021–1028. [Google Scholar] [CrossRef]
- Macleod, M. Learning lessons from MVA85A, a failed booster vaccine for BCG. BMJ 2018, 360, k66. [Google Scholar] [CrossRef] [PubMed]
- Tameris, M.; McShane, H.; McClain, J.B.; Landry, B.; Lockhart, S.; Luabeya, A.K.; Geldenhuys, H.; Shea, J.; Hussey, G.; van der Merwe, L.; et al. Lessons learnt from the first efficacy trial of a new infant tuberculosis vaccine since BCG. Tuberculosis 2013, 93, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Aguilo, N.; Marinova, D.; Gonzalo-Asensio, J. Update on TB vaccine pipeline. Appl. Sci. 2020, 10, 2632. [Google Scholar] [CrossRef]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-Based Vaccines: Current Progress and Future Challenges. Chem. Rev. 2020, 120, 3210–3229. [Google Scholar] [CrossRef]
- Stylianou, E.; Harrington-Kandt, R.; Beglov, J.; Bull, N.; Pinpathomrat, N.; Swarbrick, G.M.; Lewinsohn, D.A.; Lewinsohn, D.M.; McShane, H. Identification and evaluation of novel protective antigens for the development of a candidate tuberculosis subunit vaccine. Infect. Immun. 2018, 86, e00014-18. [Google Scholar] [CrossRef]
- Bettencourt, P.; Müller, J.; Nicastri, A.; Cantillon, D.; Madhavan, M.; Charles, P.D.; Fotso, C.B.; Wittenberg, R.; Bull, N.; Pinpathomrat, N.; et al. Identification of antigens presented by MHC for vaccines against tuberculosis. NPJ Vaccines 2020, 5, 2. [Google Scholar] [CrossRef]
- Kapopoulou, A.; Lew, J.M.; Cole, S.T. The MycoBrowser portal: A comprehensive and manually annotated resource for mycobacterial genomes. Tuberculosis 2011, 91, 8–13. [Google Scholar] [CrossRef]
- Voss, G.; Casimiro, D.; Neyrolles, O.; Williams, A.; Kaufmann, S.; McShane, H.; Hatherill, M.; Fletcher, H.A. Progress and challenges in TB vaccine development. F1000Research 2018, 7, 199. [Google Scholar] [CrossRef]
- Uplekar, S.; Heym, B.; Friocourt, V.; Rougemont, J.; Cole, S.T. Comparative genomics of ESX genes from clinical isolates of Mycobacterium tuberculosis provides evidence for gene conversion and epitope variation. Infect. Immun. 2011, 79, 4042–4049. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.J. The enigmatic PE/PPE multigene family of mycobacteria and tuberculosis vaccination. Infect. Immun. 2017, 85, e00969-16. [Google Scholar] [CrossRef] [PubMed]
- Schrager, L.K.; Chandrasekaran, P.; Fritzell, B.H.; Hatherill, M.; Lambert, P.H.; McShane, H.; Tornieporth, N.; Vekemans, J. WHO preferred product characteristics for new vaccines against tuberculosis. Lancet Infect. Dis. 2018, 18, 828–829. [Google Scholar] [CrossRef]
- Lewinsohn, D.A.; Swarbrick, G.M.; Park, B.; Cansler, M.E.; Null, M.D.; Toren, K.G.; Baseke, J.; Zalwango, S.; Mayanja-Kizza, H.; Malone, L.L.; et al. Comprehensive definition of human immunodominant CD8 antigens in tuberculosis. NPJ Vaccines 2017, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Joosten, S.A.; Ottenhoff, T.H.M.; Lewinsohn, D.M.; Hoft, D.F.; Moody, D.B.; Seshadri, C. Harnessing donor unrestricted T-cells for new vaccines against tuberculosis. Vaccine 2019, 37, 3022–3030. [Google Scholar] [CrossRef]
- Zhang, L. Multi-epitope vaccines: A promising strategy against tumors and viral infections. Cell. Mol. Immunol. 2018, 15, 182–184. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef]
- Oleszycka, E.; McCluskey, S.; Sharp, F.A.; Muñoz-Wolf, N.; Hams, E.; Gorman, A.L.; Fallon, P.G.; Lavelle, E.C. The vaccine adjuvant alum promotes IL-10 production that suppresses Th1 responses. Eur. J. Immunol. 2018, 48, 705–715. [Google Scholar] [CrossRef]
- Khandhar, A.P.; Liang, H.; Simpson, A.C.; Reed, S.G.; Carter, D.; Fox, C.B.; Orr, M.T. Physicochemical structure of a polyacrylic acid stabilized nanoparticle alum (nanoalum) adjuvant governs TH1 differentiation of CD4+ T cells. Nanoscale 2020, 12, 2515–2523. [Google Scholar] [CrossRef]
- Orr, M.T.; Khandhar, A.P.; Seydoux, E.; Liang, H.; Gage, E.; Mikasa, T.; Beebe, E.L.; Rintala, N.D.; Persson, K.H.; Ahniyaz, A.; et al. Reprogramming the adjuvant properties of aluminum oxyhydroxide with nanoparticle technology. NPJ Vaccines 2019, 4, 1. [Google Scholar] [CrossRef]
- Davidsen, J.; Rosenkrands, I.; Christensen, D.; Vangala, A.; Kirby, D.; Perrie, Y.; Agger, E.M.; Andersen, P. Characterization of cationic liposomes based on dimethyldioctadecylammonium and synthetic cord factor from M. tuberculosis (trehalose 6,6′-dibehenate)—A novel adjuvant inducing both strong CMI and antibody responses. Biochim. Biophys. Acta 2005, 1718, 22–31. [Google Scholar] [CrossRef]
- Pedersen, G.K.; Andersen, P.; Christensen, D. Immunocorrelates of CAF family adjuvants. Semin. Immunol. 2018, 39, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Woodworth, J.S.; Christensen, D.; Cassidy, J.P.; Agger, E.M.; Mortensen, R.; Andersen, P. Mucosal boosting of H56:CAF01 immunization promotes lung-localized T cells and an accelerated pulmonary response to Mycobacterium tuberculosis infection without enhancing vaccine protection. Mucosal Immunol. 2019, 12, 816–826. [Google Scholar] [CrossRef]
- van Dissel, J.T.; Joosten, S.A.; Hoff, S.T.; Soonawala, D.; Prins, C.; Hokey, D.A.; O’Dee, D.M.; Graves, A.; Thierry-Carstensen, B.; Andreasen, L.V.; et al. A novel liposomal adjuvant system, CAF01, promotes long-lived Mycobacterium tuberculosis-specific T-cell responses in human. Vaccine 2014, 32, 7098–7107. [Google Scholar] [CrossRef] [PubMed]
- Coler, R.N.; Bertholet, S.; Moutaftsi, M.; Guderian, J.A.; Windish, H.P.; Baldwin, S.L.; Laughlin, E.M.; Duthie, M.S.; Fox, C.B.; Carter, D.; et al. Development and characterization of synthetic glucopyranosyl lipid adjuvant system as a vaccine adjuvant. PLoS ONE 2011, 6, e16333. [Google Scholar] [CrossRef] [PubMed]
- Penn-Nicholson, A.; Tameris, M.; Smit, E.; Day, T.A.; Musvosvi, M.; Jayashankar, L.; Vergara, J.; Mabwe, S.; Bilek, N.; TBVPX-114 Study Team; et al. Safety and immunogenicity of the novel tuberculosis vaccine ID93 + GLA-SE in BCG-vaccinated healthy adults in South Africa: A randomised, double-blind, placebo-controlled phase 1 trial. Lancet Respir. Med. 2018, 6, 287–298. [Google Scholar] [CrossRef]
- Coler, R.N.; Day, T.A.; Ellis, R.; Piazza, F.M.; Beckmann, A.M.; Vergara, J.; Rolf, T.; Lu, L.; Alter, G.; TBVPX-113 Study Team; et al. The TLR-4 agonist adjuvant, GLA-SE, improves magnitude and quality of immune responses elicited by the ID93 tuberculosis vaccine: First-in-human trial. NPJ Vaccines 2018, 3, 34. [Google Scholar] [CrossRef]
- Van Der Meeren, O.; Hatherill, M.; Nduba, V.; Wilkinson, R.J.; Muyoyeta, M.; Van Brakel, E.; Ayles, H.M.; Henostroza, G.; Thienemann, F.; Scriba, T.J.; et al. Phase 2b controlled trial of M72/AS01E vaccine to prevent tuberculosis. N. Engl. J. Med. 2018, 379, 1621–1634. [Google Scholar] [CrossRef]
- Tait, D.R.; Hatherill, M.; Van Der Meeren, O.; Ginsberg, A.M.; Van Brakel, E.; Salaun, B.; Scriba, T.J.; Akite, E.J.; Ayles, H.M.; Bollaerts, A.; et al. Final analysis of a trial of M72/AS01E vaccine to prevent tuberculosis. N. Engl. J. Med. 2019, 381, 2429–2439. [Google Scholar] [CrossRef]
- Marciani, D.J. Elucidating the Mechanisms of Action of Saponin-Derived Adjuvants. Trends Pharmacol. Sci. 2018, 39, 573–585. [Google Scholar] [CrossRef]
- Agger, E.M. Novel adjuvant formulations for delivery of anti-tuberculosis vaccine candidates. Adv. Drug Del. Rev. 2016, 102, 73–82. [Google Scholar] [CrossRef]
- Geldenhuys, H.; Mearns, H.; Miles, D.J.; Tameris, M.; Hokey, D.; Shi, Z.; Bennett, S.; Andersen, P.; Kromann, I.; Hoff, S.T.; et al. The tuberculosis vaccine H4: IC31 is safe and induces a persistent polyfunctional CD4 T cell response in South African adults: A randomized controlled trial. Vaccine 2015, 33, 3592–3599. [Google Scholar] [CrossRef]
- Suliman, S.; Luabeya, A.; Geldenhuys, H.; Tameris, M.; Hoff, S.T.; Shi, Z.; Tait, D.; Kromann, I.; Ruhwald, M.; H56-035 Trial Group; et al. Dose optimization of H56:IC31 vaccine for tuberculosis-endemic populations a double-blind, placebo-controlled, dose-selection trial. Am. J. Respir. Crit. Care Med. 2019, 199, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Vasina, D.V.; Kleymenov, D.A.; Manuylov, V.A.; Mazunina, E.P.; Koptev, E.Y.; Tukhovskaya, E.A.; Murashev, A.N.; Gintsburg, A.L.; Gushchin, V.A.; Tkachuk, A.P. First-in-human trials of gamtbvac, a recombinant subunit tuberculosis vaccine candidate: Safety and immunogenicity assessment. Vaccines 2019, 7, 166. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.; Triccas, J.A.; Petrovsky, N. Adjuvant strategies for more effective tuberculosis vaccine immunity. Microorganisms 2019, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Counoupas, C.; Pinto, R.; Nagalingam, G.; Britton, W.J.; Petrovsky, N.; Triccas, J.A. Delta inulin-based adjuvants promote the generation of polyfunctional CD4+ T cell responses and protection against Mycobacterium tuberculosis infection. Sci. Rep. 2017, 7, 8582. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Chandrudu, S.; Rigau-Planella, B.; Islam, M.T.; Cheong, Y.S.; Liu, G.; Wang, X.; Toth, I.; Hussein, W.M. Progress in the development of subunit vaccines against malaria. Vaccines 2020, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Azuar, A.; Jin, W.; Mukaida, S.; Hussein, W.M.; Toth, I.; Skwarczynski, M. Recent advances in the development of peptide vaccines and their delivery systems against group a streptococcus. Vaccines 2019, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Kalli, K.R.; Block, M.S.; Kasi, P.M.; Erskine, C.L.; Hobday, T.J.; Dietz, A.; Padley, D.; Gustafson, M.P.; Shreeder, B.; Puglisi-Knutson, D.; et al. Folate receptor alpha peptide vaccine generates immunity in breast and ovarian cancer patients. Clin. Cancer Res. 2018, 24, 3014–3025. [Google Scholar] [CrossRef]
- Mittendorf, E.A.; Lu, B.; Melisko, M.; Price Hiller, J.; Bondarenko, I.; Brunt, A.M.; Sergii, G.; Petrakova, K.; Peoples, G.E. Efficacy and safety analysis of nelipepimut-S vaccine to prevent breast cancer recurrence: A randomized, multicenter, phase III clinical trial. Clin. Cancer Res. 2019, 25, 4248–4254. [Google Scholar] [CrossRef]
- Dakappagari, N.K.; Douglas, D.B.; Triozzi, P.L.; Stevens, V.C.; Kaumaya, P.T.P. Prevention of mammary tumors with a chimeric HER-2 B-cell epitope peptide vaccine. Cancer Res. 2000, 60, 3782–3789, PMID:10919651. [Google Scholar] [PubMed]
- Purcell, A.W.; McCluskey, J.; Rossjohn, J. More than one reason to rethink the use of peptides in vaccine design. Nat. Rev. Drug Discov. 2007, 6, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Van Den Eeden, S.J.F.; Wilson, L.; Franken, K.L.M.C.; Ottenhoff, T.H.M.; Geluk, A. Synthetic long peptide derived from Mycobacterium tuberculosis latency antigen Rv1733c protects against tuberculosis. Clin. Vaccine Immunol. 2015, 22, 1060–1069. [Google Scholar] [CrossRef]
- Ashhurst, A.S.; McDonald, D.M.; Hanna, C.C.; Stanojevic, V.A.; Britton, W.J.; Payne, R.J. Mucosal Vaccination with a Self-Adjuvanted Lipopeptide Is Immunogenic and Protective against Mycobacterium tuberculosis. J. Med. Chem. 2019, 62, 8080–8089. [Google Scholar] [CrossRef] [PubMed]
- Gowthaman, U.; Singh, V.; Zeng, W.; Jain, S.; Siddiqui, K.F.; Chodisetti, S.B.; Gurram, R.K.; Parihar, P.; Gupta, P.; Gupta, U.D.; et al. Promiscuous peptide of 16 kDa antigen linked to Pam2Cys protects against mycobacterium tuberculosis by evoking enduring memory T-cell response. J. Infect. Dis. 2011, 204, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.K.; Chodisetti, S.B.; Zeng, W.; Nadeem, S.; Maurya, S.K.; Pahari, S.; Janmeja, A.K.; Jackson, D.C.; Agrewala, J.N. A lipidated peptide of Mycobacterium tuberculosis resuscitates the protective efficacy of BCG vaccine by evoking memory T cell immunity. J. Transl. Med. 2017, 15, 201. [Google Scholar] [CrossRef]
- Rai, P.K.; Chodisetti, S.B.; Nadeem, S.; Maurya, S.K.; Gowthaman, U.; Zeng, W.; Janmeja, A.K.; Jackson, D.C.; Agrewala, J.N. A novel therapeutic strategy of lipidated promiscuous peptide against Mycobacterium tuberculosis by eliciting Th1 and Th17 immunity of host. Sci. Rep. 2016, 6, 23917. [Google Scholar] [CrossRef]
- Chesson, C.B.; Huante, M.; Nusbaum, R.J.; Walker, A.G.; Clover, T.M.; Chinnaswamy, J.; Endsley, J.J.; Rudra, J.S. Nanoscale Peptide Self-assemblies Boost BCG-primed Cellular Immunity against Mycobacterium tuberculosis. Sci. Rep. 2018, 8, 12519. [Google Scholar] [CrossRef]
- Horváti, K.; Pályi, B.; Henczkó, J.; Balka, G.; Szabó, E.; Farkas, V.; Biri-Kovács, B.; Szeder, B.; Fodor, K. A convenient synthetic method to improve immunogenicity of mycobacterium tuberculosis related T-cell epitope peptides. Vaccines 2019, 7, 101. [Google Scholar] [CrossRef]
- Rai, P.K.; Chodisetti, S.B.; Maurya, S.K.; Nadeem, S.; Zeng, W.; Janmeja, A.K.; Jackson, D.C.; Agrewala, J.N. A lipidated bi-epitope vaccine comprising of MHC-I and MHC-II binder peptides elicits protective CD4 T cell and CD8 T cell immunity against Mycobacterium tuberculosis. J. Transl. Med. 2018, 16, 279. [Google Scholar] [CrossRef]
- Chatterjee, N.; Ojha, R.; Khatoon, N.; Prajapati, V.K. Scrutinizing Mycobacterium tuberculosis membrane and secretory proteins to formulate multiepitope subunit vaccine against pulmonary tuberculosis by utilizing immunoinformatic approaches. Int. J. Biol. Macromol. 2018, 118, 180–188. [Google Scholar] [CrossRef]
- Shah, P.; Mistry, J.; Reche, P.A.; Gatherer, D.; Flower, D.R. In silico design of Mycobacterium tuberculosis epitope ensemble vaccines. Mol. Immunol. 2018, 97, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; da Silva Antunes, R.; Sidney, J.; Lindestam Arlehamn, C.S.; Grifoni, A.; Dhanda, S.K.; Paul, S.; Peters, B.; Weiskopf, D.; Sette, A. A review on T Cell epitopes identified using prediction and cell-mediated immune models for mycobacterium tuberculosisand bordetella pertussis. Front. Immunol. 2018, 9, 2778. [Google Scholar] [CrossRef] [PubMed]
- Garnica, O.; Das, K.; Devasundaram, S.; Dhandayuthapani, S. Enhanced delivery of Mycobacterium tuberculosis antigens to antigen presenting cells using RVG peptide. Tuberculosis 2019, 116S, S34–S41. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.W.; Hansen, P.R.; Holm, A.; Andersen, P. Efficient protection against Mycobacterium tuberculosis by vaccination with a single subdominant epitope from the ESAT-6 antigen. Eur. J. Immunol. 2000, 30, 1724–1732. [Google Scholar] [CrossRef]
- Aagaard, C.S.; Hoang, T.T.; Vingsbo-Lundberg, C.; Dietrich, J.; Andersen, P. Quality and Vaccine Efficacy of CD4 + T Cell Responses Directed to Dominant and Subdominant Epitopes in ESAT-6 from Mycobacterium tuberculosis. J. Immunol. 2009, 183, 2659–2668. [Google Scholar] [CrossRef]
- Geluk, A.; van den Eeden, S.J.; van Meijgaarden, K.E.; Dijkman, K.; Franken, K.L.; Ottenhoff, T.H. A multistage-polyepitope vaccine protects against Mycobacterium tuberculosis infection in HLA-DR3 transgenic mice. Vaccine 2012, 30, 7513–7521. [Google Scholar] [CrossRef]
- Doi, T.; Yamada, H.; Yajima, T.; Wajjwalku, W.; Hara, T.; Yoshikai, Y. H2-M3-Restricted CD8 + T Cells Induced by Peptide-Pulsed Dendritic Cells Confer Protection against Mycobacterium tuberculosis. J. Immunol. 2007, 178, 3806–3813. [Google Scholar] [CrossRef]
- Covián, C.; Fernández-Fierro, A.; Retamal-Díaz, A.; Díaz, F.E.; Vasquez, A.E.; Lay, M.K.; Riedel, C.A.; González, P.A.; Bueno, S.M.; Kalergis, A.M. BCG-Induced Cross-Protection and Development of Trained Immunity: Implication for Vaccine Design. Front. Immunol. 2019, 10, 2806. [Google Scholar] [CrossRef]
- Covián, C.; Retamal-Díaz, A.; Bueno, S.M.; Kalergis, A.M. Could BCG Vaccination Induce Protective Trained Immunity for SARS-CoV-2? Front. Immunol. 2020, 11, 970. [Google Scholar] [CrossRef]
- Netea, M.G.; Giamarellos-Bourboulis, E.J.; Domínguez-Andrés, J.; Curtis, N.; van Crevel, R.; van de Veerdonk, F.L.; Bonten, M. Trained Immunity: A Tool for Reducing Susceptibility to and the Severity of SARS-CoV-2 Infection. Cell 2020, 181, 969–977. [Google Scholar] [CrossRef]
- Curtis, N.; Sparrow, A.; Ghebreyesus, T.A.; Netea, M.G. Considering BCG vaccination to reduce the impact of COVID-19. Lancet 2020, 395, 1545–1546. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Netea, M.G. BCG-induced trained immunity: Can it offer protection against COVID-19? Nat. Rev. Immunol. 2020, 20, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Klinger, D.; Blass, I.; Rappoport, N.; Linial, M. Significantly improved COVID-19 outcomes in countries with higher bcg vaccination coverage: A multivariable analysis. Vaccines 2020, 8, 378. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Khan, S.; Tiwari, R.; Sircar, S.; Bhat, S.; Malik, Y.S.; Singh, K.P.; Chaicumpa, W.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J. Coronavirus disease 2019–COVID-19. Clin. Microbiol. Rev. 2020, 33, e0028-20. [Google Scholar] [CrossRef]
- O’Connor, E.; Teh, J.; Kamat, A.M.; Lawrentschuk, N. Bacillus Calmette Guérin (BCG) vaccination use in the fight against COVID-19—What’s old is new again? Future Oncol. 2020, 16, 1323–1325. [Google Scholar] [CrossRef]
- Hamiel, U.; Kozer, E.; Youngster, I. SARS-CoV-2 Rates in BCG-Vaccinated and Unvaccinated Young Adults. JAMA 2020, 232, 2340–2341. [Google Scholar] [CrossRef]
- Hunter, R.; Actor, J. The pathogenesis of post-primary tuberculosis. A game for vaccine development. Tuberculosis 2019, 116S, S114–S117. [Google Scholar] [CrossRef]
- Lewinsohn, D.A.; Lewinsohn, D.M.; Scriba, T.J. Polyfunctional CD4+ T Cells as Targets for Tuberculosis Vaccination. Front. Immunol. 2017, 8, 1262. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).