
Neuroprotective Role of Lactoferrin during Early Brain Development and Injury through Lifespan


{kind=link}
{kind=link}
{kind=link}
Abstract
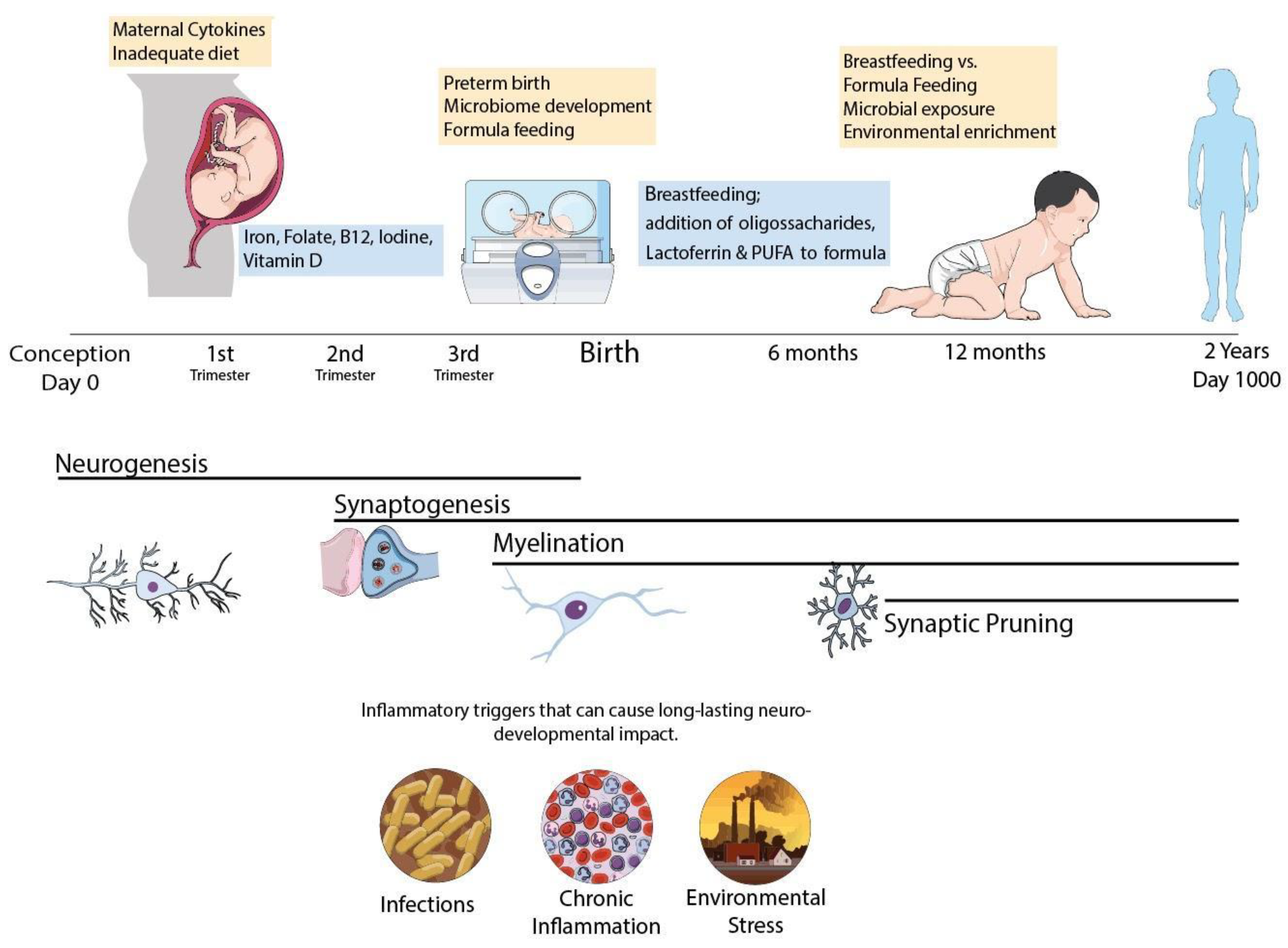
1. Fueling the Brain during Early Neurodevelopmental Stages
2. Nutritional Needs during Neurodevelopment
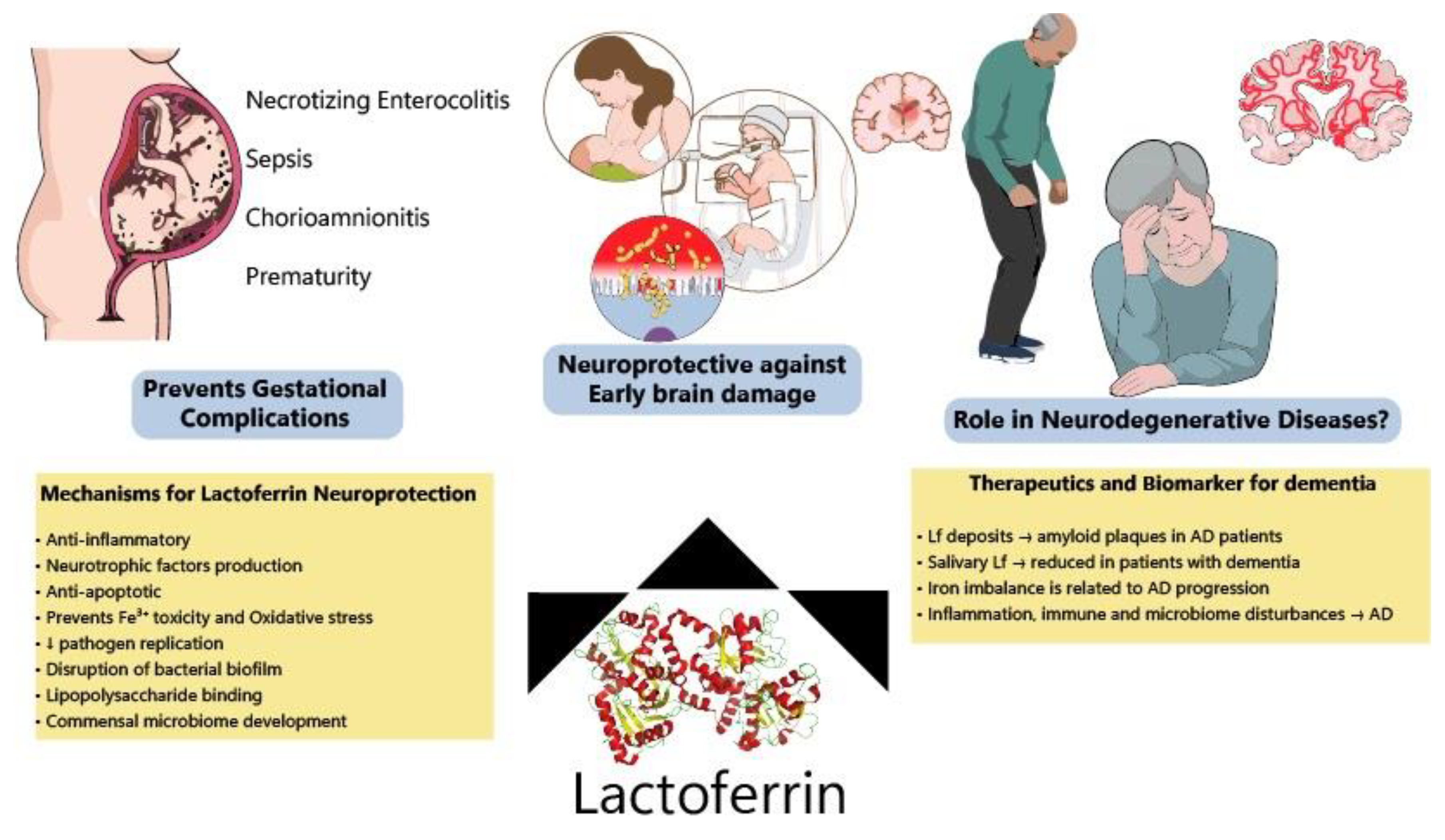
3. Lactoferrin for Preventing IUGR/Premature Delivery and Associated Brain Injury
4. Early Triggers of Neurodegeneration in Preterm Infants: Protective Roles of Lactoferrin
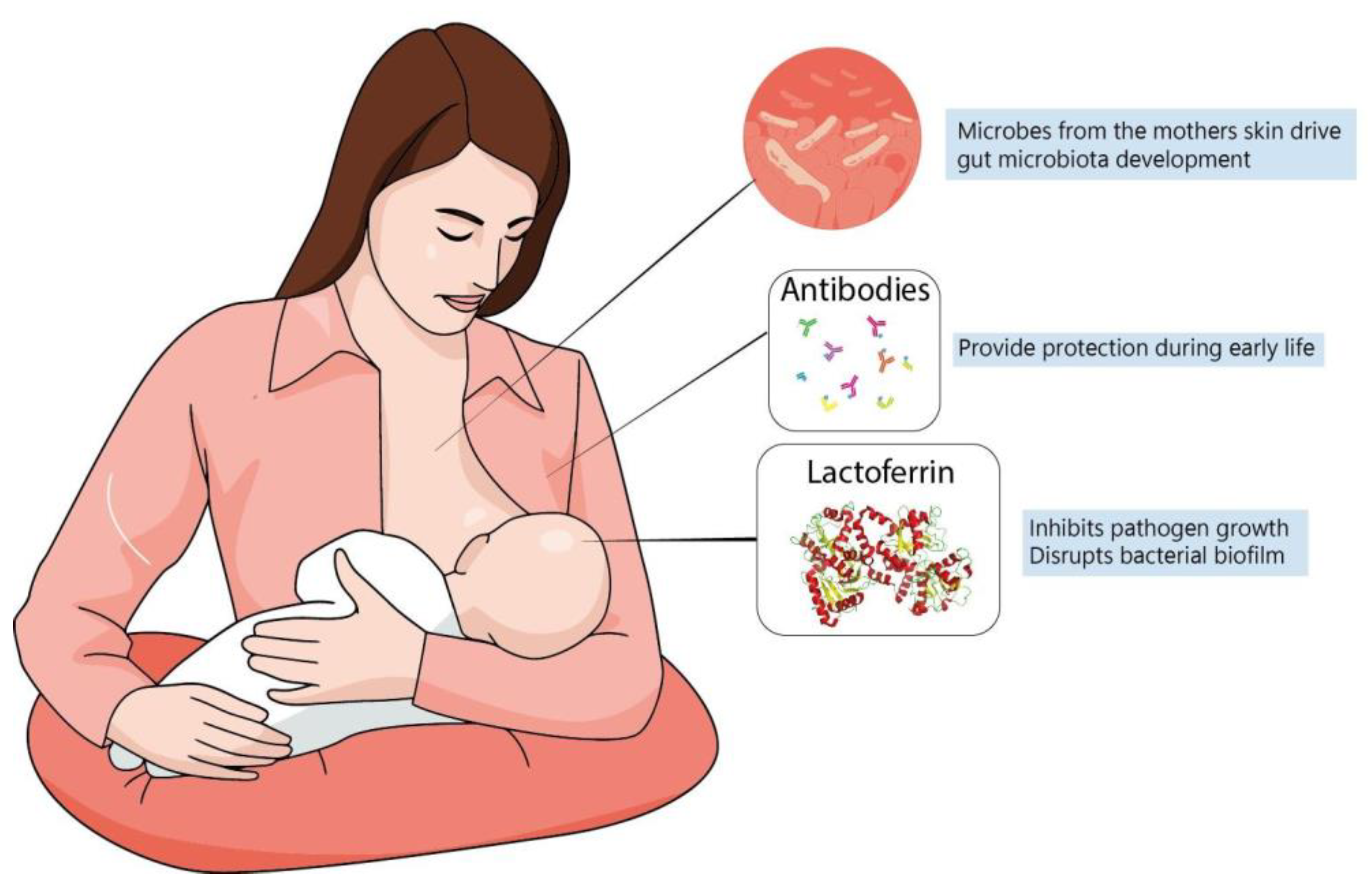
5. Lactoferrin and the Development of Infant Microbiome
6. Lactoferrin for Preventing Neurodegeneration: A Promising Molecule?
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mink, J.W.; Blumenschine, R.J.; Adams, D.B. Ratio of Central Nervous System to Body Metabolism in Vertebrates: Its Constancy and Functional Basis. Am. J. Physiol. 1981, 241, R203–R212. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, A.L.; Babbitt, C.C. The Appropriation of Glucose through Primate Neurodevelopment. J. Hum. Evol. 2014, 77, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Bordone, M.P.; Salman, M.M.; Titus, H.E.; Amini, E.; Andersen, J.V.; Chakraborti, B.; Diuba, A.V.; Dubouskaya, T.G.; Ehrke, E.; Espindola de Freitas, A.; et al. The Energetic Brain—A Review from Students to Students. J. Neurochem. 2019, 151, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Kuzawa, C.W.; Chugani, H.T.; Grossman, L.I.; Lipovich, L.; Muzik, O.; Hof, P.R.; Wildman, D.E.; Sherwood, C.C.; Leonard, W.R.; Lange, N. Metabolic Costs and Evolutionary Implications of Human Brain Development. Proc. Natl. Acad. Sci. USA 2014, 111, 13010–13015. [Google Scholar] [CrossRef] [PubMed]
- Marder, E.; Goaillard, J.-M. Variability, Compensation and Homeostasis in Neuron and Network Function. Nat. Rev. Neurosci. 2006, 7, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Chugani, H.T. Imaging Brain Metabolism in the Newborn. J. Child Neurol. 2018, 33, 851–860. [Google Scholar] [CrossRef]
- Goyal, M.S.; Raichle, M.E. Glucose Requirements of the Developing Human Brain. J. Pediatr. Gastroenterol. Nutr. 2018, 66 (Suppl. 3), S46–S49. [Google Scholar] [CrossRef]
- Dubois, J.; Alison, M.; Counsell, S.J.; Hertz-Pannier, L.; Hüppi, P.S.; Benders, M.J.N.L. MRI of the Neonatal Brain: A Review of Methodological Challenges and Neuroscientific Advances. J. Magn. Reson. Imaging 2021, 53, 1318–1343. [Google Scholar] [CrossRef]
- Bale, T.L. Epigenetic and Transgenerational Reprogramming of Brain Development. Nat. Rev. Neurosci. 2015, 16, 332–344. [Google Scholar] [CrossRef]
- Georgieff, M.K.; Ramel, S.E.; Cusick, S.E. Nutritional Influences on Brain Development. Acta Paediatr. 2018, 107, 1310–1321. [Google Scholar] [CrossRef]
- Blüml, S.; Wisnowski, J.L.; Nelson, M.D., Jr.; Paquette, L.; Gilles, F.H.; Kinney, H.C.; Panigrahy, A. Metabolic Maturation of the Human Brain from Birth through Adolescence: Insights from in Vivo Magnetic Resonance Spectroscopy. Cereb. Cortex 2013, 23, 2944–2955. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenberg, S.J.; Georgieff, M.K. COMMITTEE ON NUTRITION Advocacy for Improving Nutrition in the First 1000 Days to Support Childhood Development and Adult Health. Pediatrics 2018, 141, e20173716. [Google Scholar] [CrossRef] [PubMed]
- Ehrenkranz, R.A. Nutrition, Growth and Clinical Outcomes. World Rev. Nutr. Diet. 2014, 110, 11–26. [Google Scholar] [PubMed]
- Wachs, T.D.; Georgieff, M.; Cusick, S.; McEwen, B.S. Issues in the Timing of Integrated Early Interventions: Contributions from Nutrition, Neuroscience, and Psychological Research. In Prenatal and Childhood Nutrition: Evaluating the Neurocognitive Connections; Croft, C., Ed.; Apple Academic Press: Waretown, NJ, USA, 2015; Volume 418, pp. 363–397. [Google Scholar]
- Bordeleau, M.; Fernández de Cossío, L.; Chakravarty, M.M.; Tremblay, M.-È. From Maternal Diet to Neurodevelopmental Disorders: A Story of Neuroinflammation. Front. Cell Neurosci. 2020, 14, 612705. [Google Scholar] [CrossRef]
- Bryce, J.; Coitinho, D.; Darnton-Hill, I.; Pelletier, D.; Pinstrup-Andersen, P. Maternal and Child Undernutrition Study Group Maternal and Child Undernutrition: Effective Action at National Level. Lancet 2008, 371, 510–526. [Google Scholar] [CrossRef]
- Prado, E.L.; Dewey, K.G. Nutrition and Brain Development in Early Life. Nutr. Rev. 2014, 72, 267–284. [Google Scholar] [CrossRef]
- Tau, G.Z.; Peterson, B.S. Normal Development of Brain Circuits. Neuropsychopharmacology 2010, 35, 147–168. [Google Scholar] [CrossRef]
- Hüppi, P.S.; Warfield, S.; Kikinis, R.; Barnes, P.D.; Zientara, G.P.; Jolesz, F.A.; Tsuji, M.K.; Volpe, J.J. Quantitative Magnetic Resonance Imaging of Brain Development in Premature and Mature Newborns. Ann. Neurol. 1998, 43, 224–235. [Google Scholar] [CrossRef]
- Harding, J.E.; Cormack, B.E.; Alexander, T.; Alsweiler, J.M.; Bloomfield, F.H. Advances in Nutrition of the Newborn Infant. Lancet 2017, 389, 1660–1668. [Google Scholar] [CrossRef]
- Liberman, N.; Wang, S.Y.; Greer, E.L. Transgenerational Epigenetic Inheritance: From Phenomena to Molecular Mechanisms. Curr. Opin. Neurobiol. 2019, 59, 189–206. [Google Scholar] [CrossRef]
- Bodnar, L.M.; Wisner, K.L. Nutrition and Depression: Implications for Improving Mental Health among Childbearing-Aged Women. Biol. Psychiatry 2005, 58, 679–685. [Google Scholar] [CrossRef]
- McNamara, J.P.; Huber, K. Metabolic and Endocrine Role of Adipose Tissue during Lactation. Annu. Rev. Anim. Biosci. 2018, 6, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Monk, C.; Georgieff, M.K.; Osterholm, E.A. Research Review: Maternal Prenatal Distress and Poor Nutrition—Mutually Influencing Risk Factors Affecting Infant Neurocognitive Development. J. Child Psychol. Psychiatry 2013, 54, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Casal, M.N.; Estevez, D.; De-Regil, L.M. Multiple Micronutrient Supplements in Pregnancy: Implementation Considerations for Integration as Part of Quality Services in Routine Antenatal Care. Objectives, Results, and Conclusions of the Meeting. Matern. Child Nutr. 2018, 14 (Suppl. 5), e12704. [Google Scholar] [CrossRef] [PubMed]
- Kominiarek, M.A.; Rajan, P. Nutrition Recommendations in Pregnancy and Lactation. Med. Clin. N. Am. 2016, 100, 1199–1215. [Google Scholar] [CrossRef]
- Ares Segura, S.; Arena Ansótegui, J.; Marta Díaz-Gómez, N. The Importance of Maternal Nutrition during Breastfeeding: Do Breastfeeding Mothers Need Nutritional Supplements? An. Pediatría 2016, 84, 347.e1–347.e7. [Google Scholar] [CrossRef]
- Martin, J.C.; Zhou, S.J.; Flynn, A.C.; Malek, L.; Greco, R.; Moran, L. The Assessment of Diet Quality and Its Effects on Health Outcomes Pre-Pregnancy and during Pregnancy. Semin. Reprod. Med. 2016, 34, 83–92. [Google Scholar]
- Georgieff, M.K. Nutrition and the Developing Brain: Nutrient Priorities and Measurement. Am. J. Clin. Nutr. 2007, 85, 614S–620S. [Google Scholar]
- Buyken, A.E.; Goletzke, J.; Joslowski, G.; Felbick, A.; Cheng, G.; Herder, C.; Brand-Miller, J.C. Association between Carbohydrate Quality and Inflammatory Markers: Systematic Review of Observational and Interventional Studies. Am. J. Clin. Nutr. 2014, 99, 813–833. [Google Scholar] [CrossRef]
- Nielsen, S.D.; Beverly, R.L.; Underwood, M.A.; Dallas, D.C. Differences and Similarities in the Peptide Profile of Preterm and Term Mother’s Milk, and Preterm and Term Infant Gastric Samples. Nutrients 2020, 12, 2825. [Google Scholar] [CrossRef]
- Bolton, J.L.; Bilbo, S.D. Developmental Programming of Brain and Behavior by Perinatal Diet: Focus on Inflammatory Mechanisms. Dialogues Clin. Neurosci. 2014, 16, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Gordon, J.I. The Core Gut Microbiome, Energy Balance and Obesity. J. Physiol. 2009, 587, 4153–4158. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial Ecology: Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota-Gut-Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.E.; Lallier, S.W.; Waidyaratne, G.; Thompson, M.D.; Tipple, T.E.; Hester, M.E.; Trask, A.J.; Rogers, L.K. Maternal High Fat Diet Exposure Is Associated with Increased Hepcidin Levels, Decreased Myelination, and Neurobehavioral Changes in Male Offspring. Brain Behav. Immun. 2016, 58, 369–378. [Google Scholar] [CrossRef]
- Castanon, N.; Luheshi, G.; Layé, S. Role of Neuroinflammation in the Emotional and Cognitive Alterations Displayed by Animal Models of Obesity. Front. Neurosci. 2015, 9, 229. [Google Scholar] [CrossRef]
- Kentner, A.C.; Bilbo, S.D.; Brown, A.S.; Hsiao, E.Y.; McAllister, A.K.; Meyer, U.; Pearce, B.D.; Pletnikov, M.V.; Yolken, R.H.; Bauman, M.D. Maternal Immune Activation: Reporting Guidelines to Improve the Rigor, Reproducibility, and Transparency of the Model. Neuropsychopharmacology 2019, 44, 245–258. [Google Scholar] [CrossRef]
- Mousa, A.; Naqash, A.; Lim, S. Macronutrient and Micronutrient Intake during Pregnancy: An Overview of Recent Evidence. Nutrients 2019, 11, 443. [Google Scholar] [CrossRef]
- Hadley, K.B.; Ryan, A.S.; Forsyth, S.; Gautier, S.; Salem, N., Jr. The Essentiality of Arachidonic Acid in Infant Development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef]
- Kawakita, E.; Hashimoto, M.; Shido, O. Docosahexaenoic Acid Promotes Neurogenesis in Vitro and in Vivo. Neuroscience 2006, 139, 991–997. [Google Scholar] [CrossRef]
- Tokuda, H.; Kontani, M.; Kawashima, H.; Kiso, Y.; Shibata, H.; Osumi, N. Differential Effect of Arachidonic Acid and Docosahexaenoic Acid on Age-Related Decreases in Hippocampal Neurogenesis. Neurosci. Res. 2014, 88, 58–66. [Google Scholar] [CrossRef] [PubMed]
- de la Owens, S.P.; de la Presa Owens, S.; Innis, S.M. Docosahexaenoic and Arachidonic Acid Prevent a Decrease in Dopaminergic and Serotoninergic Neurotransmitters in Frontal Cortex Caused by a Linoleic and α-Linolenic Acid Deficient Diet in Formula-Fed Piglets. J. Nutr. 1999, 129, 2088–2093. [Google Scholar] [CrossRef]
- Darios, F.; Davletov, B. Omega-3 and Omega-6 Fatty Acids Stimulate Cell Membrane Expansion by Acting on Syntaxin 3. Nature 2006, 440, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Paoli, A.; Rubini, A.; Volek, J.S.; Grimaldi, K.A. Beyond Weight Loss: A Review of the Therapeutic Uses of Very-Low-Carbohydrate (Ketogenic) Diets. Eur. J. Clin. Nutr. 2013, 67, 789–796. [Google Scholar] [CrossRef]
- Spiegler, E.; Kim, Y.-K.; Wassef, L.; Shete, V.; Quadro, L. Maternal-Fetal Transfer and Metabolism of Vitamin A and Its Precursor β-Carotene in the Developing Tissues. Biochim. Biophys. Acta 2012, 1821, 88–98. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, L.M.; Teixeira, F.M.E.; Sato, M.N. Impact of Retinoic Acid on Immune Cells and Inflammatory Diseases. Mediators Inflamm. 2018, 2018, 3067126. [Google Scholar] [CrossRef]
- Kang, B.Y.; Chung, S.W.; Kim, S.H.; Kang, S.N.; Choe, Y.K.; Kim, T.S. Retinoid-Mediated Inhibition of Interleukin-12 Production in Mouse Macrophages Suppresses Th1 Cytokine Profile in CD4(+) T Cells. Br. J. Pharmacol. 2000, 130, 581–586. [Google Scholar] [CrossRef][Green Version]
- Alatshan, A.; Kovács, G.E.; Aladdin, A.; Czimmerer, Z.; Tar, K.; Benkő, S. All-Trans Retinoic Acid Enhances Both the Signaling for Priming and the Glycolysis for Activation of NLRP3 Inflammasome in Human Macrophage. Cells 2020, 9, 1591. [Google Scholar] [CrossRef]
- Császár, E.; Lénárt, N.; Cserép, C.; Környei, Z.; Fekete, R.; Pósfai, B.; Balázsfi, D.; Hangya, B.; Schwarcz, A.D.; Szabadits, E.; et al. Microglia Modulate Blood Flow, Neurovascular Coupling, and Hypoperfusion via Purinergic Actions. J. Exp. Med. 2022, 219, e20211071. [Google Scholar] [CrossRef]
- Stamm, R.A.; Houghton, L.A. Nutrient Intake Values for Folate during Pregnancy and Lactation Vary Widely around the World. Nutrients 2013, 5, 3920–3947. [Google Scholar] [CrossRef]
- De-Regil, L.M.; Fernández-Gaxiola, A.C.; Dowswell, T.; Peña-Rosas, J.P. Effects and Safety of Periconceptional Folate Supplementation for Preventing Birth Defects. Cochrane Database Syst. Rev. 2015, 2015, CD007950. [Google Scholar]
- Parisi, F.; Laoreti, A.; Cetin, I. Multiple Micronutrient Needs in Pregnancy in Industrialized Countries. Ann. Nutr. Metab. 2014, 65, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Naninck, E.F.G.; Stijger, P.C.; Brouwer-Brolsma, E.M. The Importance of Maternal Folate Status for Brain Development and Function of Offspring. Adv. Nutr. 2019, 10, 502–519. [Google Scholar] [CrossRef] [PubMed]
- Ford, T.C.; Downey, L.A.; Simpson, T.; McPhee, G.; Oliver, C.; Stough, C. The Effect of a High-Dose Vitamin B Multivitamin Supplement on the Relationship between Brain Metabolism and Blood Biomarkers of Oxidative Stress: A Randomized Control Trial. Nutrients 2018, 10, 1860. [Google Scholar] [CrossRef] [PubMed]
- Khaire, A.; Rathod, R.; Kale, A.; Joshi, S. Vitamin B12 and Omega-3 Fatty Acids Together Regulate Lipid Metabolism in Wistar Rats. Prostaglandins Leukot. Essent. Fatty Acids 2015, 99, 7–17. [Google Scholar] [CrossRef]
- Guillot, X.; Semerano, L.; Saidenberg-Kermanac’h, N.; Falgarone, G.; Boissier, M.-C. Vitamin D and Inflammation. Joint Bone Spine 2010, 77, 552–557. [Google Scholar] [CrossRef]
- Kleinewietfeld, M.; Manzel, A.; Titze, J.; Kvakan, H.; Yosef, N.; Linker, R.A.; Muller, D.N.; Hafler, D.A. Sodium Chloride Drives Autoimmune Disease by the Induction of Pathogenic TH17 Cells. Nature 2013, 496, 518–522. [Google Scholar] [CrossRef]
- Faraco, G.; Hochrainer, K.; Segarra, S.G.; Schaeffer, S.; Santisteban, M.M.; Menon, A.; Jiang, H.; Holtzman, D.M.; Anrather, J.; Iadecola, C. Dietary Salt Promotes Cognitive Impairment through Tau Phosphorylation. Nature 2019, 574, 686–690. [Google Scholar] [CrossRef]
- Zhang, W.-C.; Zheng, X.-J.; Du, L.-J.; Sun, J.-Y.; Shen, Z.-X.; Shi, C.; Sun, S.; Zhang, Z.; Chen, X.-Q.; Qin, M.; et al. High Salt Primes a Specific Activation State of Macrophages, M(Na). Cell Res. 2015, 25, 893–910. [Google Scholar] [CrossRef]
- Guo, H.-X.; Ye, N.; Yan, P.; Qiu, M.-Y.; Zhang, J.; Shen, Z.-G.; He, H.-Y.; Tian, Z.-Q.; Li, H.-L.; Li, J.-T. Sodium Chloride Exacerbates Dextran Sulfate Sodium-Induced Colitis by Tuning Proinflammatory and Antiinflammatory Lamina Propria Mononuclear Cells through P38/MAPK Pathway in Mice. World J. Gastroenterol. 2018, 24, 1779–1794. [Google Scholar] [CrossRef]
- Xiao, Z.X.; Hu, X.; Zhang, X.; Chen, Z.; Wang, J.; Jin, K.; Cao, F.L.; Sun, B.; Bellanti, J.A.; Olsen, N.; et al. High Salt Diet Accelerates the Progression of Murine Lupus through Dendritic Cells via the P38 MAPK and STAT1 Signaling Pathways. Signal Transduct. Target. Ther. 2020, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Aceves, C.; Anguiano, B.; Delgado, G. The Extrathyronine Actions of Iodine as Antioxidant, Apoptotic, and Differentiation Factor in Various Tissues. Thyroid 2013, 23, 938–946. [Google Scholar] [CrossRef]
- Berbel, P.; Obregón, M.J.; Bernal, J.; Escobar del Rey, F.; Morreale de Escobar, G. Iodine Supplementation during Pregnancy: A Public Health Challenge. Trends Endocrinol. Metab. 2007, 18, 338–343. [Google Scholar] [CrossRef] [PubMed]
- Skeaff, S.A. Iodine Deficiency in Pregnancy: The Effect on Neurodevelopment in the Child. Nutrients 2011, 3, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Stoltzfus, R.J. Iron Deficiency: Global Prevalence and Consequences. Food Nutr. Bull. 2003, 24, S99–S103. [Google Scholar] [CrossRef] [PubMed]
- Gambling, L.; Charania, Z.; Hannah, L.; Antipatis, C.; Lea, R.G.; McArdle, H.J. Effect of Iron Deficiency on Placental Cytokine Expression and Fetal Growth in the Pregnant Rat. Biol. Reprod. 2002, 66, 516–523. [Google Scholar] [CrossRef]
- Donangelo, C.M.; King, J.C. Maternal Zinc Intakes and Homeostatic Adjustments during Pregnancy and Lactation. Nutrients 2012, 4, 782–798. [Google Scholar] [CrossRef]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and Its Importance for Human Health: An Integrative Review. J. Res. Med. Sci. 2013, 18, 144–157. [Google Scholar]
- Sauer, A.K.; Grabrucker, A.M. Zinc Deficiency During Pregnancy Leads to Altered Microbiome and Elevated Inflammatory Markers in Mice. Front. Neurosci. 2019, 13, 1295. [Google Scholar] [CrossRef]
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and Anti-Inflammatory Effects of Zinc. Zinc-Dependent NF-ΚB Signaling. Inflammopharmacology 2017, 25, 11–24. [Google Scholar] [CrossRef]
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc Status Is Associated with Inflammation, Oxidative Stress, Lipid, and Glucose Metabolism. J. Physiol. Sci. 2018, 68, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Grabrucker, S.; Jannetti, L.; Eckert, M.; Gaub, S.; Chhabra, R.; Pfaender, S.; Mangus, K.; Reddy, P.P.; Rankovic, V.; Schmeisser, M.J.; et al. Zinc Deficiency Dysregulates the Synaptic ProSAP/Shank Scaffold and Might Contribute to Autism Spectrum Disorders. Brain 2014, 137, 137–152. [Google Scholar] [CrossRef] [PubMed]
- van de Looij, Y.; Larpin, C.; Cabungcal, J.-H.; Sanches, E.F.; Toulotte, A.; Do, K.Q.; Sizonenko, S.V. Nutritional Intervention for Developmental Brain Damage: Effects of Lactoferrin Supplementation in Hypocaloric Induced Intrauterine Growth Restriction Rat Pups. Front. Endocrinol. 2019, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Parks, W.T.; Simhan, H.N.; Bertolet, M.; Catov, J.M. Early Pregnancy Immune Profile and Preterm Birth Classified According to Uteroplacental Lesions. Placenta 2020, 89, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, T.J.; Sizonenko, S.V. Lactoferrin and Prematurity: A Promising Milk Protein? Biochem. Cell Biol. 2017, 95, 22–30. [Google Scholar] [CrossRef]
- Inder, T.E.; Warfield, S.K.; Wang, H.; Hüppi, P.S.; Volpe, J.J. Abnormal Cerebral Structure Is Present at Term in Premature Infants. Pediatrics 2005, 115, 286–294. [Google Scholar] [CrossRef]
- Khwaja, O.; Volpe, J.J. Pathogenesis of Cerebral White Matter Injury of Prematurity. Arch. Dis. Child Fetal Neonatal Ed. 2008, 93, F153–F161. [Google Scholar] [CrossRef]
- Ronayne de Ferrer, P.A.; Baroni, A.; Sambucetti, M.E.; López, N.E.; Ceriani Cernadas, J.M. Lactoferrin Levels in Term and Preterm Milk. J. Am. Coll. Nutr. 2000, 19, 370–373. [Google Scholar] [CrossRef]
- Sandomirsky, B.P.; Galchenko, S.E.; Galchenko, K.S. Antioxidative Properties of Lactoferrin from Bovine Colostrum before and after Its Lyophilization. Cryo Lett. 2003, 24, 275–280. [Google Scholar]
- Satué-Gracia, M.T.; Frankel, E.N.; Rangavajhyala, N.; German, J.B. Lactoferrin in Infant Formulas: Effect on Oxidation. J. Agric. Food Chem. 2000, 48, 4984–4990. [Google Scholar] [CrossRef]
- Takayama, Y. Lactoferrin and Its Role in Wound Healing; Springer: Dordrecht, The Netherlands, 2012; ISBN 9789400724662. [Google Scholar]
- Mikulic, N.; Uyoga, M.A.; Mwasi, E.; Stoffel, N.U.; Zeder, C.; Karanja, S.; Zimmermann, M.B. Iron Absorption Is Greater from Apo-Lactoferrin and Is Similar Between Holo-Lactoferrin and Ferrous Sulfate: Stable Iron Isotope Studies in Kenyan Infants. J. Nutr. 2020, 150, 3200–3207. [Google Scholar] [CrossRef] [PubMed]
- Sreedhara, A.; Flengsrud, R.; Langsrud, T.; Kaul, P.; Prakash, V.; Vegarud, G.E. Structural Characteristic, PH and Thermal Stabilities of Apo and Holo Forms of Caprine and Bovine Lactoferrins. Biometals 2010, 23, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Hagberg, H.; Mallard, C.; Jacobsson, B. Role of Cytokines in Preterm Labour and Brain Injury. BJOG 2005, 112 (Suppl. 1), 16–18. [Google Scholar] [CrossRef] [PubMed]
- Metz-Boutigue, M.H.; Jollès, J.; Mazurier, J.; Schoentgen, F.; Legrand, D.; Spik, G.; Montreuil, J.; Jollès, P. Human Lactotransferrin: Amino Acid Sequence and Structural Comparisons with Other Transferrins. Eur. J. Biochem. 1984, 145, 659–676. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.D.; Tan, S. Perinatal Infections, Prematurity and Brain Injury. Curr. Opin. Pediatr. 2006, 18, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, A.; Otsuki, K.; Sasaki, Y.; Sawada, M.; Mitsukawa, K.; Chiba, H.; Nagatsuka, M.; Okai, T.; Kato, A. Preventive Effect of Recombinant Human Lactoferrin in a Rabbit Preterm Delivery Model. Am. J. Obstet. Gynecol. 2005, 192, 1038–1043. [Google Scholar] [CrossRef]
- Lang, J.; Yang, N.; Deng, J.; Liu, K.; Yang, P.; Zhang, G.; Jiang, C. Inhibition of SARS Pseudovirus Cell Entry by Lactoferrin Binding to Heparan Sulfate Proteoglycans. PLoS ONE 2011, 6, e23710. [Google Scholar] [CrossRef]
- Wotring, J.W.; Fursmidt, R.; Ward, L.; Sexton, J.Z. Evaluating the in Vitro Efficacy of Bovine Lactoferrin Products against SARS-CoV-2 Variants of Concern. J. Dairy Sci. 2022, 105, 2791–2802. [Google Scholar] [CrossRef]
- Mitsuhashi, Y.; Otsuki, K.; Yoda, A.; Shimizu, Y.; Saito, H.; Yanaihara, T. Effect of Lactoferrin on Lipopolysaccharide (LPS) Induced Preterm Delivery in Mice. Acta Obstet. Gynecol. Scand. 2000, 79, 355–358. [Google Scholar]
- Sasaki, Y.; Otsuki, K.; Hasegawa, A.; Sawada, M.; Chiba, H.; Negishi, M.; Nagatsuka, M.; Okai, T. Preventive Effect of Recombinant Human Lactoferrin on Lipopolysaccharide-Induced Preterm Delivery in Mice. Acta Obstet. Gynecol. Scand. 2004, 83, 1035–1038. [Google Scholar] [CrossRef]
- Otsuki, K.; Yakuwa, K.; Sawada, M.; Hasegawa, A.; Sasaki, Y.; Mitsukawa, K.; Chiba, H.; Nagatsuka, M.; Saito, H.; Okai, T. Recombinant Human Lactoferrin Has Preventive Effects on Lipopolysaccharide-Induced Preterm Delivery and Production of Inflammatory Cytokines in Mice. J. Perinat. Med. 2005, 33, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Paesano, R.; Pietropaoli, M.; Berlutti, F.; Valenti, P. Bovine Lactoferrin in Preventing Preterm Delivery Associated with Sterile Inflammation. Biochem. Cell Biol. 2012, 90, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Locci, M.; Nazzaro, G.; Miranda, M.; Salzano, E.; Montagnani, S.; Castaldo, C.; De Placido, G. Vaginal Lactoferrin in Asymptomatic Patients at Low Risk for Pre-Term Labour for Shortened Cervix: Cervical Length and Interleukin-6 Changes. J. Obstet. Gynaecol. 2013, 33, 144–148. [Google Scholar] [CrossRef]
- Ginet, V.; van de Looij, Y.; Petrenko, V.; Toulotte, A.; Kiss, J.; Hüppi, P.S.; Sizonenko, S.V. Lactoferrin during Lactation Reduces Lipopolysaccharide-Induced Brain Injury. Biofactors 2016, 42, 323–336. [Google Scholar] [PubMed]
- van de Looij, Y.; Ginet, V.; Chatagner, A.; Toulotte, A.; Somm, E.; Hüppi, P.S.; Sizonenko, S.V. Lactoferrin during Lactation Protects the Immature Hypoxic-Ischemic Rat Brain. Ann. Clin. Transl. Neurol. 2014, 1, 955–967. [Google Scholar] [CrossRef]
- Zakharova, E.T.; Kostevich, V.A.; Sokolov, A.V.; Vasilyev, V.B. Human Apo-Lactoferrin as a Physiological Mimetic of Hypoxia Stabilizes Hypoxia-Inducible Factor-1 Alpha. Biometals 2012, 25, 1247–1259. [Google Scholar] [CrossRef]
- Sanches, E.; van de Looij, Y.; Sow, S.; Toulotte, A.; da Silva, A.; Modernell, L.; Sizonenko, S. Dose-Dependent Neuroprotective Effects of Bovine Lactoferrin Following Neonatal Hypoxia-Ischemia in the Immature Rat Brain. Nutrients 2021, 13, 3880. [Google Scholar] [CrossRef]
- Sokolov, A.V.; Dubrovskaya, N.M.; Kostevich, V.A.; Vasilev, D.S.; Voynova, I.V.; Zakharova, E.T.; Runova, O.L.; Semak, I.V.; Budevich, A.I.; Nalivaeva, N.N.; et al. Lactoferrin Induces Erythropoietin Synthesis and Rescues Cognitive Functions in the Offspring of Rats Subjected to Prenatal Hypoxia. Nutrients 2022, 14, 1399. [Google Scholar] [CrossRef]
- Kaufman, D.A.; Berenz, A.; Itell, H.L.; Conaway, M.; Blackman, A.; Nataro, J.P.; Permar, S.R. Dose Escalation Study of Bovine Lactoferrin in Preterm Infants: Getting the Dose Right. Biochem. Cell Biol. 2021, 99, 7–13. [Google Scholar] [CrossRef]
- Chen, K.; Jin, S.; Chen, H.; Cao, Y.; Dong, X.; Li, H.; Zhou, Z.; Liu, C. Dose Effect of Bovine Lactoferrin Fortification on Diarrhea and Respiratory Tract Infections in Weaned Infants with Anemia: A Randomized, Controlled Trial. Nutrition 2021, 90, 111288. [Google Scholar] [CrossRef]
- Li, W.; Fu, K.; Lv, X.; Wang, Y.; Wang, J.; Li, H.; Tian, W.; Cao, R. Lactoferrin Suppresses Lipopolysaccharide-Induced Endometritis in Mice via down-Regulation of the NF-ΚB Pathway. Int. Immunopharmacol. 2015, 28, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, B.; Yang, C.; Shi, Y.; Dong, Z.; Troy, F.A., 2nd. Functional Correlates and Impact of Dietary Lactoferrin Intervention and Its Concentration-Dependence on Neurodevelopment and Cognition in Neonatal Piglets. Mol. Nutr. Food Res. 2021, 65, e2001099. [Google Scholar] [CrossRef] [PubMed]
- Dobryk, D.; Dobryk, O.; Dobryanskyy, D. The Effect of Enteral Lactoferrin Supplementation in Prevention of Morbidity Associated with Immature Digestive Tract in Premature Infants: Prospective Cohort Study. Georgian Med. News 2022, 323, 94–101. [Google Scholar]
- Leung, M.P.; Thompson, B.; Black, J.; Dai, S.; Alsweiler, J.M. The Effects of Preterm Birth on Visual Development. Clin. Exp. Optom. 2018, 101, 4–12. [Google Scholar] [CrossRef]
- Batalle, D.; Hughes, E.J.; Zhang, H.; Tournier, J.-D.; Tusor, N.; Aljabar, P.; Wali, L.; Alexander, D.C.; Hajnal, J.V.; Nosarti, C.; et al. Early Development of Structural Networks and the Impact of Prematurity on Brain Connectivity. Neuroimage 2017, 149, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Back, S.A.; Luo, N.L.; Borenstein, N.S.; Levine, J.M.; Volpe, J.J.; Kinney, H.C. Late Oligodendrocyte Progenitors Coincide with the Developmental Window of Vulnerability for Human Perinatal White Matter Injury. J. Neurosci. 2001, 21, 1302–1312. [Google Scholar] [CrossRef]
- Bassan, H.; Limperopoulos, C.; Visconti, K.; Mayer, D.L.; Feldman, H.A.; Avery, L.; Benson, C.B.; Stewart, J.; Ringer, S.A.; Soul, J.S.; et al. Neurodevelopmental Outcome in Survivors of Periventricular Hemorrhagic Infarction. Pediatrics 2007, 120, 785–792. [Google Scholar] [CrossRef]
- Özek, E.; Kersin, S.G. Intraventricular Hemorrhage in Preterm Babies. Turk Pediatri Ars. 2020, 55, 215–221. [Google Scholar]
- Foix-L’helias, L.; Baud, O.; Lenclen, R.; Kaminski, M.; Lacaze-Masmonteil, T. Benefit of Antenatal Glucocorticoids According to the Cause of Very Premature Birth. Arch. Dis. Child Fetal Neonatal Ed. 2005, 90, F46–F48. [Google Scholar] [CrossRef]
- Adams-Chapman, I.; Heyne, R.J.; DeMauro, S.B.; Duncan, A.F.; Hintz, S.R.; Pappas, A.; Vohr, B.R.; McDonald, S.A.; Das, A.; Newman, J.E.; et al. Neurodevelopmental Impairment Among Extremely Preterm Infants in the Neonatal Research Network. Pediatrics 2018, 141, e20173091. [Google Scholar] [CrossRef]
- Stoll, B.J.; Hansen, N.I.; Adams-Chapman, I.; Fanaroff, A.A.; Hintz, S.R.; Vohr, B.; Higgins, R.D. National Institute of Child Health and Human Development Neonatal Research Network Neurodevelopmental and Growth Impairment among Extremely Low-Birth-Weight Infants with Neonatal Infection. JAMA 2004, 292, 2357–2365. [Google Scholar] [CrossRef] [PubMed]
- Pammi, M.; Suresh, G. Enteral Lactoferrin Supplementation for Prevention of Sepsis and Necrotizing Enterocolitis in Preterm Infants. Cochrane Database Syst. Rev. 2020, 3, CD007137. [Google Scholar] [CrossRef] [PubMed]
- Mwaniki, M.K.; Atieno, M.; Lawn, J.E.; Newton, C.R. Long-Term Neurodevelopmental Outcomes after Intrauterine and Neonatal Insults: A Systematic Review. Lancet 2012, 379, 445–452. [Google Scholar] [CrossRef]
- Horváth-Puhó, E.; van Kassel, M.N.; Gonçalves, B.P.; de Gier, B.; Procter, S.R.; Paul, P.; van der Ende, A.; Søgaard, K.K.; Hahné, S.J.M.; Chandna, J.; et al. Mortality, Neurodevelopmental Impairments, and Economic Outcomes after Invasive Group B Streptococcal Disease in Early Infancy in Denmark and the Netherlands: A National Matched Cohort Study. Lancet Child Adolesc. Health 2021, 5, 398–407. [Google Scholar] [CrossRef]
- Bilbo, S.D.; Block, C.L.; Bolton, J.L.; Hanamsagar, R.; Tran, P.K. Beyond Infection—Maternal Immune Activation by Environmental Factors, Microglial Development, and Relevance for Autism Spectrum Disorders. Exp. Neurol. 2018, 299, 241–251. [Google Scholar] [CrossRef]
- Hollander, J.A.; Cory-Slechta, D.A.; Jacka, F.N.; Szabo, S.T.; Guilarte, T.R.; Bilbo, S.D.; Mattingly, C.J.; Moy, S.S.; Haroon, E.; Hornig, M.; et al. Beyond the Looking Glass: Recent Advances in Understanding the Impact of Environmental Exposures on Neuropsychiatric Disease. Neuropsychopharmacology 2020, 45, 1086–1096. [Google Scholar] [CrossRef]
- Williamson, L.L.; Chao, A.; Bilbo, S.D. Environmental Enrichment Alters Glial Antigen Expression and Neuroimmune Function in the Adult Rat Hippocampus. Brain Behav. Immun. 2012, 26, 500–510. [Google Scholar] [CrossRef]
- Smith, C.J.; Kingsbury, M.A.; Dziabis, J.E.; Hanamsagar, R.; Malacon, K.E.; Tran, J.N.; Norris, H.A.; Gulino, M.; Bordt, E.A.; Bilbo, S.D. Neonatal Immune Challenge Induces Female-Specific Changes in Social Behavior and Somatostatin Cell Number. Brain Behav. Immun. 2020, 90, 332–345. [Google Scholar] [CrossRef]
- Bilbo, S.D.; Biedenkapp, J.C.; Der-Avakian, A.; Watkins, L.R.; Rudy, J.W.; Maier, S.F. Neonatal Infection-Induced Memory Impairment after Lipopolysaccharide in Adulthood Is Prevented via Caspase-1 Inhibition. J. Neurosci. 2005, 25, 8000–8009. [Google Scholar] [CrossRef]
- Cao, P.; Chen, C.; Liu, A.; Shan, Q.; Zhu, X.; Jia, C.; Peng, X.; Zhang, M.; Farzinpour, Z.; Zhou, W.; et al. Early-Life Inflammation Promotes Depressive Symptoms in Adolescence via Microglial Engulfment of Dendritic Spines. Neuron 2021, 109, 2573–2589.e9. [Google Scholar] [CrossRef]
- Sanches, E.F.; Carvalho, A.S.; van de Looij, Y.; Toulotte, A.; Wyse, A.T.; Netto, C.A.; Sizonenko, S.V. Experimental Cerebral Palsy Causes Microstructural Brain Damage in Areas Associated to Motor Deficits but No Spatial Memory Impairments in the Developing Rat. Brain Res. 2021, 1761, 147389. [Google Scholar] [CrossRef] [PubMed]
- Posillico, C.K.; Garcia-Hernandez, R.E.; Tronson, N.C. Sex Differences and Similarities in the Neuroimmune Response to Central Administration of Poly I:C. J. Neuroinflamm. 2021, 18, 193. [Google Scholar] [CrossRef] [PubMed]
- Seki, D.; Mayer, M.; Hausmann, B.; Pjevac, P.; Giordano, V.; Goeral, K.; Unterasinger, L.; Klebermaß-Schrehof, K.; De Paepe, K.; Van de Wiele, T.; et al. Aberrant Gut-Microbiota-Immune-Brain Axis Development in Premature Neonates with Brain Damage. Cell Host Microbe 2021, 29, 1558–1572.e6. [Google Scholar] [CrossRef] [PubMed]
- Ratsika, A.; Codagnone, M.C.; O’Mahony, S.; Stanton, C.; Cryan, J.F. Priming for Life: Early Life Nutrition and the Microbiota-Gut-Brain Axis. Nutrients 2021, 13, 423. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between Microbiota and Immunity in Health and Disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Walker, W.A. The Importance of Appropriate Initial Bacterial Colonization of the Intestine in Newborn, Child, and Adult Health. Pediatr. Res. 2017, 82, 387–395. [Google Scholar] [CrossRef]
- De Palma, G.; Blennerhassett, P.; Lu, J.; Deng, Y.; Park, A.J.; Green, W.; Denou, E.; Silva, M.A.; Santacruz, A.; Sanz, Y.; et al. Microbiota and Host Determinants of Behavioural Phenotype in Maternally Separated Mice. Nat. Commun. 2015, 6, 7735. [Google Scholar] [CrossRef]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Kim, H.-J.; Cho, M.-H.; Shim, W.H.; Kim, J.K.; Jeon, E.-Y.; Kim, D.-H.; Yoon, S.-Y. Deficient Autophagy in Microglia Impairs Synaptic Pruning and Causes Social Behavioral Defects. Mol. Psychiatry 2016, 22, 1576–1584. [Google Scholar] [CrossRef]
- Dziabis, J.E.; Bilbo, S.D. Microglia and Sensitive Periods in Brain Development; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–24. [Google Scholar]
- Eltokhi, A.; Janmaat, I.E.; Genedi, M.; Haarman, B.C.M.; Sommer, I.E.C. Dysregulation of Synaptic Pruning as a Possible Link between Intestinal Microbiota Dysbiosis and Neuropsychiatric Disorders. J. Neurosci. Res. 2020, 98, 1335–1369. [Google Scholar] [CrossRef]
- Medawar, E.; Haange, S.-B.; Rolle-Kampczyk, U.; Engelmann, B.; Dietrich, A.; Thieleking, R.; Wiegank, C.; Fries, C.; Horstmann, A.; Villringer, A.; et al. Gut Microbiota Link Dietary Fiber Intake and Short-Chain Fatty Acid Metabolism with Eating Behavior. Transl. Psychiatry 2021, 11, 500. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Masujima, Y.; Ushiroda, C.; Mizushima, R.; Taira, S.; Ohue-Kitano, R.; Kimura, I. Dietary Short-Chain Fatty Acid Intake Improves the Hepatic Metabolic Condition via FFAR3. Sci. Rep. 2019, 9, 16574. [Google Scholar] [CrossRef] [PubMed]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of Short-Chain Fatty Acids and Their Receptors in Inflammation and Carcinogenesis. Pharmacol. Ther. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, M.; Wicksteed, B.; Schiltz, G.E.; Gilchrist, A.; Layden, B.T. SCFA Receptors in Pancreatic β Cells: Novel Diabetes Targets? Trends Endocrinol. Metab. 2016, 27, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef]
- Sandall, J.; Tribe, R.M.; Avery, L.; Mola, G.; Visser, G.H.; Homer, C.S.; Gibbons, D.; Kelly, N.M.; Kennedy, H.P.; Kidanto, H.; et al. Short-Term and Long-Term Effects of Caesarean Section on the Health of Women and Children. Lancet 2018, 392, 1349–1357. [Google Scholar] [CrossRef]
- Fettweis, J.M.; Serrano, M.G.; Brooks, J.P.; Edwards, D.J.; Girerd, P.H.; Parikh, H.I.; Huang, B.; Arodz, T.J.; Edupuganti, L.; Glascock, A.L.; et al. The Vaginal Microbiome and Preterm Birth. Nat. Med. 2019, 25, 1012–1021. [Google Scholar] [CrossRef]
- Bashiardes, S.; Thaiss, C.A.; Elinav, E. It’s in the Milk: Feeding the Microbiome to Promote Infant Growth. Cell Metab. 2016, 23, 393–394. [Google Scholar] [CrossRef]
- Charbonneau, M.R.; O’Donnell, D.; Blanton, L.V.; Totten, S.M.; Davis, J.C.C.; Barratt, M.J.; Cheng, J.; Guruge, J.; Talcott, M.; Bain, J.R.; et al. Sialylated Milk Oligosaccharides Promote Microbiota-Dependent Growth in Models of Infant Undernutrition. Cell 2016, 164, 859–871. [Google Scholar] [CrossRef]
- Zou, L.; Pande, G.; Akoh, C.C. Infant Formula Fat Analogs and Human Milk Fat: New Focus on Infant Developmental Needs. Annu. Rev. Food Sci. Technol. 2016, 7, 139–165. [Google Scholar] [CrossRef]
- Ahern, G.J.; Hennessy, A.A.; Ryan, C.A.; Ross, R.P.; Stanton, C. Advances in Infant Formula Science. Annu. Rev. Food Sci. Technol. 2019, 10, 75–102. [Google Scholar]
- Chernikova, D.A.; Madan, J.C.; Housman, M.L.; Zain-Ul-Abideen, M.; Lundgren, S.N.; Morrison, H.G.; Sogin, M.L.; Williams, S.M.; Moore, J.H.; Karagas, M.R.; et al. The Premature Infant Gut Microbiome during the First 6 Weeks of Life Differs Based on Gestational Maturity at Birth. Pediatr. Res. 2018, 84, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Baranowski, J.R.; Claud, E.C. Necrotizing Enterocolitis and the Preterm Infant Microbiome. Adv. Exp. Med. Biol. 2019, 1125, 25–36. [Google Scholar]
- Cristofalo, E.A.; Schanler, R.J.; Blanco, C.L.; Sullivan, S.; Trawoeger, R.; Kiechl-Kohlendorfer, U.; Dudell, G.; Rechtman, D.J.; Lee, M.L.; Lucas, A.; et al. Randomized Trial of Exclusive Human Milk versus Preterm Formula Diets in Extremely Premature Infants. J. Pediatr. 2013, 163, 1592–1595.e1. [Google Scholar] [CrossRef]
- Sprockett, D.; Fukami, T.; Relman, D.A. Role of Priority Effects in the Early-Life Assembly of the Gut Microbiota. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 197–205. [Google Scholar]
- Henrick, B.M.; Rodriguez, L.; Lakshmikanth, T.; Pou, C.; Henckel, E.; Arzoomand, A.; Olin, A.; Wang, J.; Mikes, J.; Tan, Z.; et al. Bifidobacteria-Mediated Immune System Imprinting Early in Life. Cell 2021, 184, 3884–3898.e11. [Google Scholar] [CrossRef]
- Gopalakrishna, K.P.; Hand, T.W. Influence of Maternal Milk on the Neonatal Intestinal Microbiome. Nutrients 2020, 12, 823. [Google Scholar] [CrossRef]
- Ziemka-Nalecz, M.; Jaworska, J.; Sypecka, J.; Polowy, R.; Filipkowski, R.K.; Zalewska, T. Sodium Butyrate, a Histone Deacetylase Inhibitor, Exhibits Neuroprotective/Neurogenic Effects in a Rat Model of Neonatal Hypoxia-Ischemia. Mol. Neurobiol. 2017, 54, 5300–5318. [Google Scholar] [CrossRef]
- Jaworska, J.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The Potential Neuroprotective Role of a Histone Deacetylase Inhibitor, Sodium Butyrate, after Neonatal Hypoxia-Ischemia. J. Neuroinflamm. 2017, 14, 34. [Google Scholar] [CrossRef]
- Erny, D.; Prinz, M. Microbiology: Gut Microbes Augment Neurodegeneration. Nature 2017, 544, 304–305. [Google Scholar] [CrossRef] [PubMed]
- Masson, P.L.; Heremans, J.F.; Schonne, E. Lactoferrin, an Iron-Binding Protein in Neutrophilic Leukocytes. J. Exp. Med. 1969, 130, 643–658. [Google Scholar] [CrossRef] [PubMed]
- Wandersman, C.; Delepelaire, P. Bacterial Iron Sources: From Siderophores to Hemophores. Annu. Rev. Microbiol. 2004, 58, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Fainstein, N.; Elad, A.; Lachish, M.; Goldfarb, S.; Einstein, O.; Ben-Hur, T. Microbial Pathogens Induce Neurodegeneration in Alzheimer’s Disease Mice: Protection by Microglial Regulation. J. Neuroinflamm. 2022, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Parsek, M.R.; Greenberg, E.P.; Welsh, M.J. A Component of Innate Immunity Prevents Bacterial Biofilm Development. Nature 2002, 417, 552–555. [Google Scholar] [CrossRef]
- Paulsson, M.A.; Svensson, U.; Kishore, A.R.; Naidu, A.S. Thermal Behavior of Bovine Lactoferrin in Water and Its Relation to Bacterial Interaction and Antibacterial Activity. J. Dairy Sci. 1993, 76, 3711–3720. [Google Scholar] [CrossRef]
- William Hutchens, T.; Lonnerdal, B.; Rumball, S.V. Lactoferrin: Structure and Function; Springer: New York, NY, USA, 2012; ISBN 9781461360872. [Google Scholar]
- Mastromarino, P.; Capobianco, D.; Campagna, G.; Laforgia, N.; Drimaco, P.; Dileone, A.; Baldassarre, M.E. Correlation between Lactoferrin and Beneficial Microbiota in Breast Milk and Infant’s Feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef]
- Xu, G.; Xiong, W.; Hu, Q.; Zuo, P.; Shao, B.; Lan, F.; Lu, X.; Xu, Y.; Xiong, S. Lactoferrin-Derived Peptides and Lactoferricin Chimera Inhibit Virulence Factor Production and Biofilm Formation in Pseudomonas Aeruginosa. J. Appl. Microbiol. 2010, 109, 1311–1318. [Google Scholar] [CrossRef]
- Ando, K.; Hasegawa, K.; Shindo, K.-I.; Furusawa, T.; Fujino, T.; Kikugawa, K.; Nakano, H.; Takeuchi, O.; Akira, S.; Akiyama, T.; et al. Human Lactoferrin Activates NF-KappaB through the Toll-like Receptor 4 Pathway While It Interferes with the Lipopolysaccharide-Stimulated TLR4 Signaling. FEBS J. 2010, 277, 2051–2066. [Google Scholar] [CrossRef]
- Niño, D.F.; Zhou, Q.; Yamaguchi, Y.; Martin, L.Y.; Wang, S.; Fulton, W.B.; Jia, H.; Lu, P.; Prindle, T., Jr.; Zhang, F.; et al. Cognitive Impairments Induced by Necrotizing Enterocolitis Can Be Prevented by Inhibiting Microglial Activation in Mouse Brain. Sci. Transl. Med. 2018, 10, eaan0237. [Google Scholar] [CrossRef]
- Okubo, K.; Kamiya, M.; Urano, Y.; Nishi, H.; Herter, J.M.; Mayadas, T.; Hirohama, D.; Suzuki, K.; Kawakami, H.; Tanaka, M.; et al. Lactoferrin Suppresses Neutrophil Extracellular Traps Release in Inflammation. EBioMedicine 2016, 10, 204–215. [Google Scholar] [CrossRef] [PubMed]
- Li, K.-J.; Lu, M.-C.; Hsieh, S.-C.; Wu, C.-H.; Yu, H.-S.; Tsai, C.-Y.; Yu, C.-L. Release of Surface-Expressed Lactoferrin from Polymorphonuclear Neutrophils after Contact with CD4+ T Cells and Its Modulation on Th1/Th2 Cytokine Production. J. Leukoc. Biol. 2006, 80, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Legrand, D.; Elass, E.; Carpentier, M.; Mazurier, J. Interactions of Lactoferrin with Cells Involved in Immune Function. Biochem. Cell Biol. 2006, 84, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Kruzel, M.L.; Zimecki, M.; Actor, J.K. Lactoferrin in a Context of Inflammation-Induced Pathology. Front. Immunol. 2017, 8, 1438. [Google Scholar] [CrossRef] [PubMed]
- Lutaty, A.; Soboh, S.; Schif-Zuck, S.; Zeituni-Timor, O.; Rostoker, R.; Podolska, M.J.; Schauer, C.; Herrmann, M.; Muñoz, L.E.; Ariel, A. A 17-KDa Fragment of Lactoferrin Associates With the Termination of Inflammation and Peptides Within Promote Resolution. Front. Immunol. 2018, 9, 644. [Google Scholar] [CrossRef]
- Rosa, L.; Cutone, A.; Lepanto, M.S.; Scotti, M.J.; Conte, M.P.; Paesano, R.; Valenti, P. Physico-Chemical Properties Influence the Functions and Efficacy of Commercial Bovine Lactoferrins. Biometals 2018, 31, 301–312. [Google Scholar] [CrossRef]
- Park, S.-H.; Lee, J.H.; Shin, J.; Kim, J.-S.; Cha, B.; Lee, S.; Kwon, K.S.; Shin, Y.W.; Choi, S.H. Cognitive Function Improvement after Fecal Microbiota Transplantation in Alzheimer’s Dementia Patient: A Case Report. Curr. Med. Res. Opin. 2021, 37, 1739–1744. [Google Scholar] [CrossRef]
- Hazan, S. Rapid Improvement in Alzheimer’s Disease Symptoms Following Fecal Microbiota Transplantation: A Case Report. J. Int. Med. Res. 2020, 48, 300060520925930. [Google Scholar] [CrossRef]
- Sun, J.; Xu, J.; Ling, Y.; Wang, F.; Gong, T.; Yang, C.; Ye, S.; Ye, K.; Wei, D.; Song, Z.; et al. Fecal Microbiota Transplantation Alleviated Alzheimer’s Disease-like Pathogenesis in APP/PS1 Transgenic Mice. Transl. Psychiatry 2019, 9, 189. [Google Scholar] [CrossRef]
- Xiang, S.; Ji, J.-L.; Li, S.; Cao, X.-P.; Xu, W.; Tan, L.; Tan, C.-C. Efficacy and Safety of Probiotics for the Treatment of Alzheimer’s Disease, Mild Cognitive Impairment, and Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2022, 14, 730036. [Google Scholar] [CrossRef]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kruzel, M.; Aronowski, J. Lactoferrin and Hematoma Detoxification after Intracerebral Hemorrhage. Biochem. Cell Biol. 2021, 99, 97–101. [Google Scholar] [CrossRef]
- Zhao, X.; Kruzel, M.; Ting, S.-M.; Sun, G.; Savitz, S.I.; Aronowski, J. Optimized Lactoferrin as a Highly Promising Treatment for Intracerebral Hemorrhage: Pre-Clinical Experience. J. Cereb. Blood Flow Metab. 2021, 41, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ke, W.; Han, L.; Liu, Y.; Shao, K.; Jiang, C.; Pei, Y. Lactoferrin-Modified Nanoparticles Could Mediate Efficient Gene Delivery to the Brain in Vivo. Brain Res. Bull. 2010, 81, 600–604. [Google Scholar] [CrossRef]
- Mao, H.; Li, L.; Fan, Q.; Angelini, A.; Saha, P.K.; Coarfa, C.; Rajapakshe, K.; Perera, D.; Cheng, J.; Wu, H.; et al. Endothelium-Specific Depletion of LRP1 Improves Glucose Homeostasis through Inducing Osteocalcin. Nat. Commun. 2021, 12, 5296. [Google Scholar] [CrossRef]
- Fillebeen, C.; Descamps, L.; Dehouck, M.P.; Fenart, L.; Benaïssa, M.; Spik, G.; Cecchelli, R.; Pierce, A. Receptor-Mediated Transcytosis of Lactoferrin through the Blood-Brain Barrier. J. Biol. Chem. 1999, 274, 7011–7017. [Google Scholar] [CrossRef]
- Agrawal, M.; Ajazuddin; Tripathi, D.K.; Saraf, S.; Saraf, S.; Antimisiaris, S.G.; Mourtas, S.; Hammarlund-Udenaes, M.; Alexander, A. Recent Advancements in Liposomes Targeting Strategies to Cross Blood-Brain Barrier (BBB) for the Treatment of Alzheimer’s Disease. J. Control Release 2017, 260, 61–77. [Google Scholar] [CrossRef]
- Belaidi, A.A.; Masaldan, S.; Southon, A.; Kalinowski, P.; Acevedo, K.; Appukuttan, A.T.; Portbury, S.; Lei, P.; Agarwal, P.; Leurgans, S.E.; et al. Apolipoprotein E Potently Inhibits Ferroptosis by Blocking Ferritinophagy. Mol. Psychiatry 2022. [Google Scholar] [CrossRef]
- Stockwell, B.R. Ferroptosis Turns 10: Emerging Mechanisms, Physiological Functions, and Therapeutic Applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef]
- Guo, C.; Yang, Z.-H.; Zhang, S.; Chai, R.; Xue, H.; Zhang, Y.-H.; Li, J.-Y.; Wang, Z.-Y. Intranasal Lactoferrin Enhances α-Secretase-Dependent Amyloid Precursor Protein Processing via the ERK1/2-CREB and HIF-1α Pathways in an Alzheimer’s Disease Mouse Model. Neuropsychopharmacology 2017, 42, 2504–2515. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M.; Saraf, S.; Saraf, S.; Antimisiaris, S.G.; Chougule, M.B.; Shoyele, S.A.; Alexander, A. Nose-to-Brain Drug Delivery: An Update on Clinical Challenges and Progress towards Approval of Anti-Alzheimer Drugs. J. Control Release 2018, 281, 139–177. [Google Scholar] [CrossRef] [PubMed]
- Reseco, L.; Atienza, M.; Fernandez-Alvarez, M.; Carro, E.; Cantero, J.L. Salivary Lactoferrin Is Associated with Cortical Amyloid-Beta Load, Cortical Integrity, and Memory in Aging. Alzheimers. Res. Ther. 2021, 13, 150. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Pareja, F.; Del Ser, T.; Valentí, M.; de la Fuente, M.; Bartolome, F.; Carro, E. Salivary Lactoferrin as Biomarker for Alzheimer’s Disease: Brain-Immunity Interactions. Alzheimers. Dement. 2020, 16, 1196–1204. [Google Scholar] [CrossRef]
- Carro, E.; Bartolomé, F.; Bermejo-Pareja, F.; Villarejo-Galende, A.; Molina, J.A.; Ortiz, P.; Calero, M.; Rabano, A.; Cantero, J.L.; Orive, G. Early Diagnosis of Mild Cognitive Impairment and Alzheimer’s Disease Based on Salivary Lactoferrin. Alzheimer’s Dement. 2017, 8, 131–138. [Google Scholar] [CrossRef]
- Antequera, D.; Moneo, D.; Carrero, L.; Bartolome, F.; Ferrer, I.; Proctor, G.; Carro, E. Salivary Lactoferrin Expression in a Mouse Model of Alzheimer’s Disease. Front. Immunol. 2021, 12, 749468. [Google Scholar] [CrossRef]
- Gleerup, H.S.; Jensen, C.S.; Høgh, P.; Hasselbalch, S.G.; Simonsen, A.H. Lactoferrin in Cerebrospinal Fluid and Saliva Is Not a Diagnostic Biomarker for Alzheimer’s Disease in a Mixed Memory Clinic Population. EBioMedicine 2021, 67, 103361. [Google Scholar] [CrossRef]
- Prinz, M.; Priller, J. The Role of Peripheral Immune Cells in the CNS in Steady State and Disease. Nat. Neurosci. 2017, 20, 136–144. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation, Metaflammation and Immunometabolic Disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- González-Sánchez, M.; Bartolome, F.; Antequera, D.; Puertas-Martín, V.; González, P.; Gómez-Grande, A.; Llamas-Velasco, S.; Herrero-San Martín, A.; Pérez-Martínez, D.; Villarejo-Galende, A.; et al. Decreased Salivary Lactoferrin Levels Are Specific to Alzheimer’s Disease. EBioMedicine 2020, 57, 102834. [Google Scholar] [CrossRef]
- Wang, J.; Bi, M.; Liu, H.; Song, N.; Xie, J. The Protective Effect of Lactoferrin on Ventral Mesencephalon Neurons against MPP+ Is Not Connected with Its Iron Binding Ability. Sci. Rep. 2015, 5, 10729. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wang, J.; Rogers, J.; Xie, J. Brain Iron Metabolism Dysfunction in Parkinson’s Disease. Mol. Neurobiol. 2017, 54, 3078–3101. [Google Scholar] [CrossRef]
- Xu, S.-F.; Zhang, Y.-H.; Wang, S.; Pang, Z.-Q.; Fan, Y.-G.; Li, J.-Y.; Wang, Z.-Y.; Guo, C. Lactoferrin Ameliorates Dopaminergic Neurodegeneration and Motor Deficits in MPTP-Treated Mice. Redox Biol. 2019, 21, 101090. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, H.; Zhu, N.; Xu, Z.; Wang, Y.; Qu, Y.; Wang, J. Lactoferrin Protects against Iron Dysregulation, Oxidative Stress, and Apoptosis in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP)-Induced Parkinson’s Disease in Mice. J. Neurochem. 2020, 152, 397–415. [Google Scholar] [CrossRef] [PubMed]
- Harach, T.; Marungruang, N.; Duthilleul, N.; Cheatham, V.; Mc Coy, K.D.; Frisoni, G.; Neher, J.J.; Fåk, F.; Jucker, M.; Lasser, T.; et al. Reduction of Abeta Amyloid Pathology in APPPS1 Transgenic Mice in the Absence of Gut Microbiota. Sci. Rep. 2017, 7, 41802. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. The Endotoxin Hypothesis of Neurodegeneration. J. Neuroinflamm. 2019, 16, 180. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schirmbeck, G.H.; Sizonenko, S.; Sanches, E.F. Neuroprotective Role of Lactoferrin during Early Brain Development and Injury through Lifespan. Nutrients 2022, 14, 2923. https://doi.org/10.3390/nu14142923
Schirmbeck GH, Sizonenko S, Sanches EF. Neuroprotective Role of Lactoferrin during Early Brain Development and Injury through Lifespan. Nutrients. 2022; 14(14):2923. https://doi.org/10.3390/nu14142923
Chicago/Turabian StyleSchirmbeck, Gabriel Henrique, Stéphane Sizonenko, and Eduardo Farias Sanches. 2022. "Neuroprotective Role of Lactoferrin during Early Brain Development and Injury through Lifespan" Nutrients 14, no. 14: 2923. https://doi.org/10.3390/nu14142923
APA StyleSchirmbeck, G. H., Sizonenko, S., & Sanches, E. F. (2022). Neuroprotective Role of Lactoferrin during Early Brain Development and Injury through Lifespan. Nutrients, 14(14), 2923. https://doi.org/10.3390/nu14142923