
Effects of Gamma Amino Butyric Acid (GABA) on Nutrient Uptake of Loquat [Eriobotrya japonica (Thunb.) Lindl.] Seedlings

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Design
2.3. Determination of Indicators
2.4. Statistical Analysis
3. Results
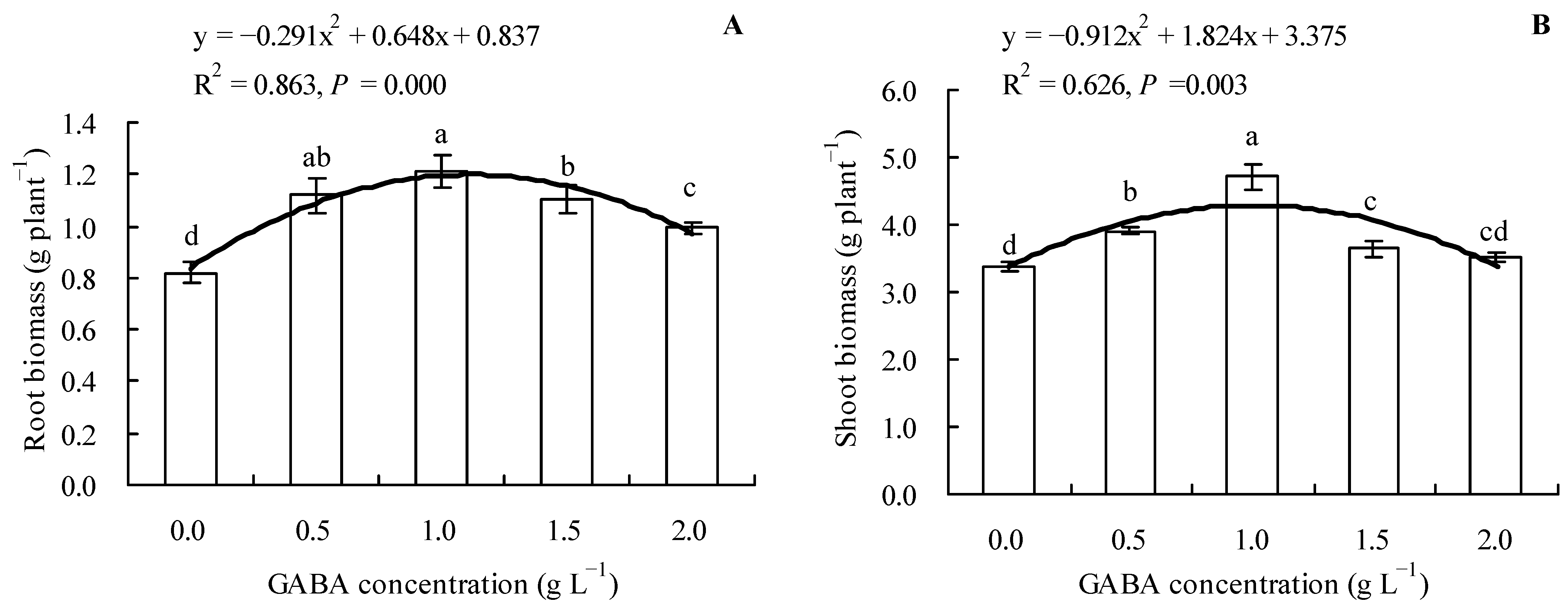
3.1. Effects of GABA on the Biomass (Dry Weight) of Loquat Seedlings
3.2. Effects of GABA on Photosynthetic Pigment Content in the Leaves of Loquat Seedlings
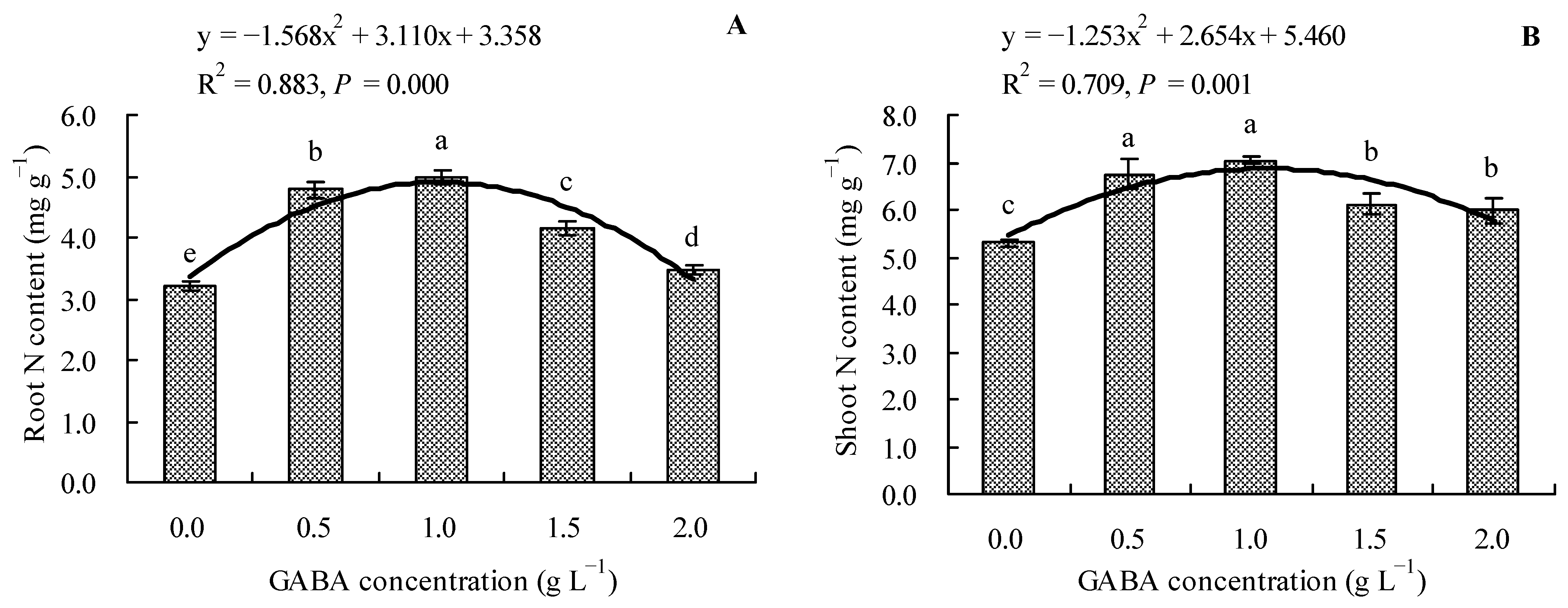
3.3. Effects of GABA on Total N Content in Loquat Seedlings
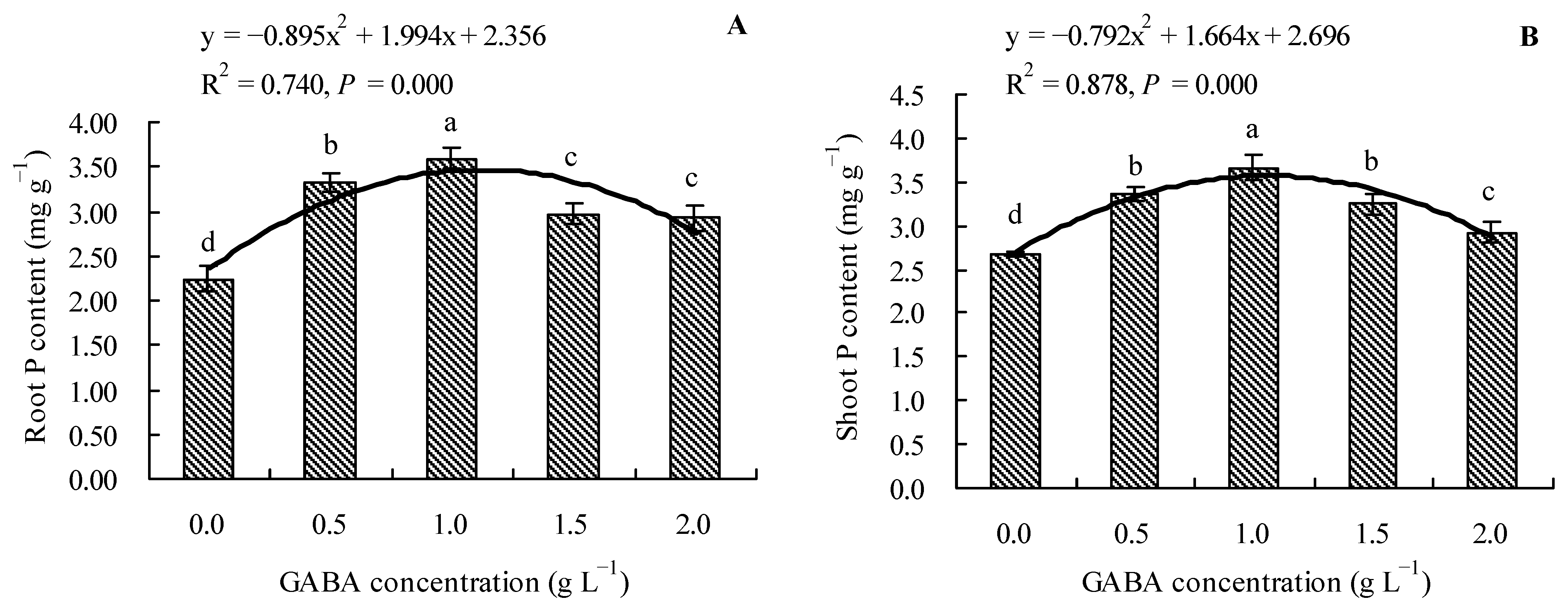
3.4. Effects of GABA on Total P Content in Loquat Seedlings
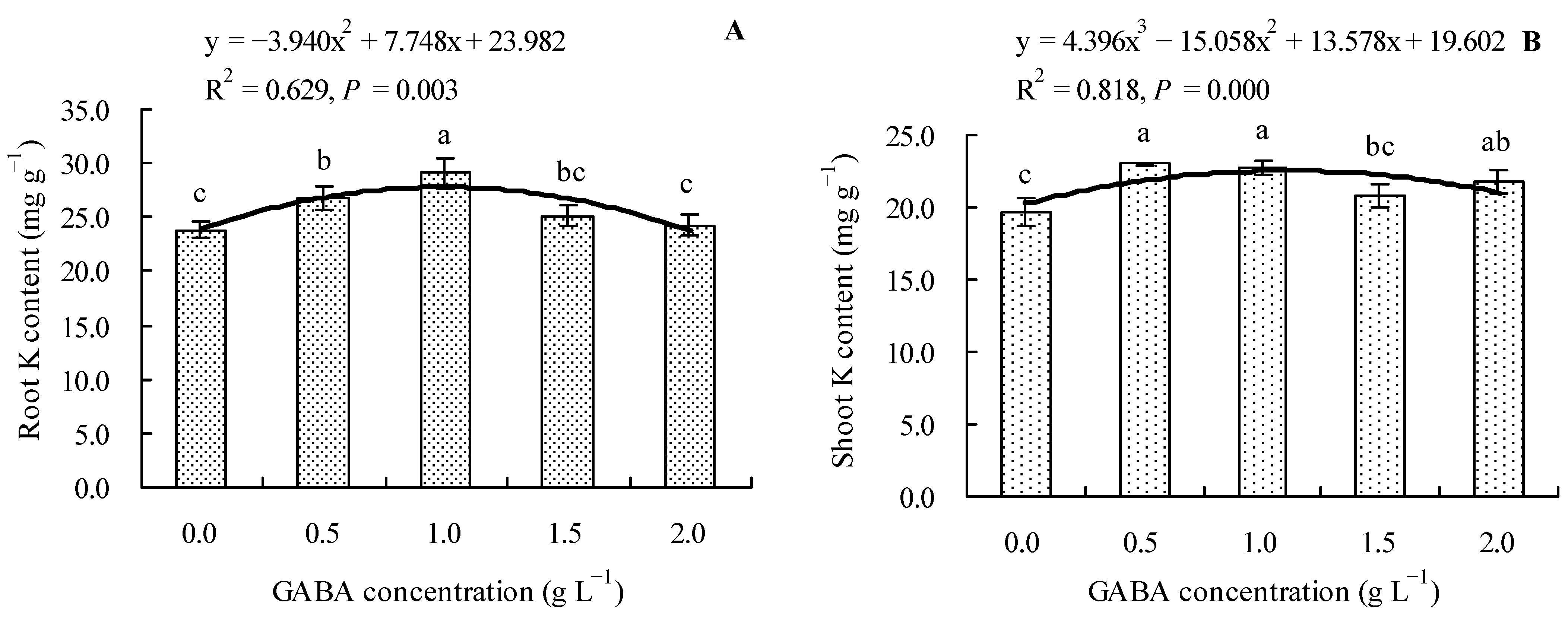
3.5. Effects of GABA on Total K Content in Loquat Seedlings
3.6. Effects of GABA on Soil pH Value and Available Nutrient Concentration
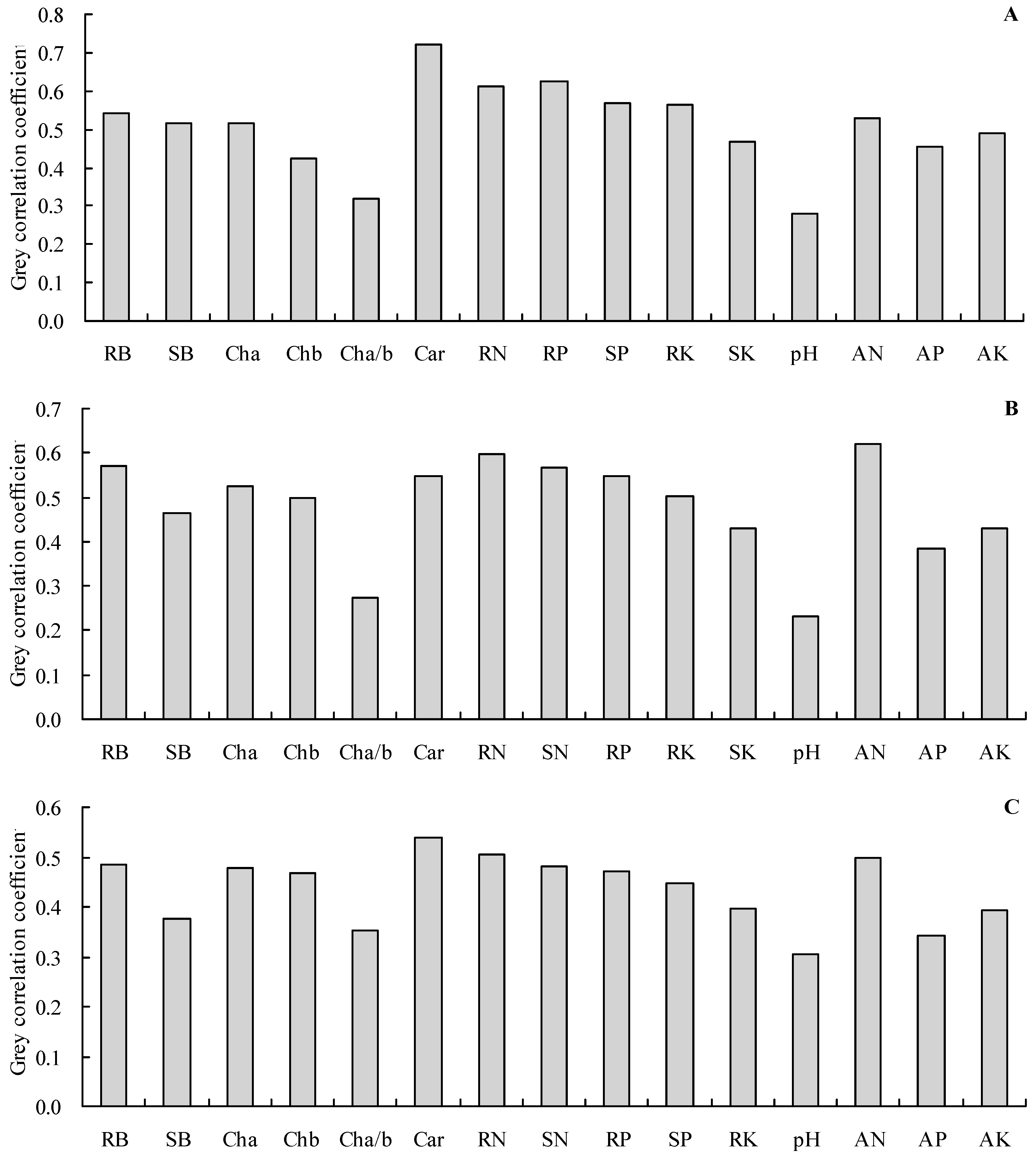
3.7. Correlation and Grey Relational Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Goldstein, G.; Bucci, S.J.; Scholz, F.G. Why do trees adjust water relations and hydraulic architecture in response to nutrient availability? Tree Physiol. 2013, 33, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Nath, M.; Tuteja, N. NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress. Protoplasma 2016, 253, 767–786. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Pan, H.L.; Pan, T.F.; Tang, H.R.; Wang, X.R.; Pan, D.M. Research progress on the interaction between scion and rootstock in fruit trees. Acta Hortic. Sin. 2017, 44, 1645–1657. [Google Scholar]
- Wu, T.R. Preliminary Study on the Potential of Different Loquat (Eriobotrya japonica) Resources as Rootstocks; Southwest University: Chongqing, China, 2020. [Google Scholar]
- Chen, X.; Li, N.; Liu, C.; Wang, H.; Li, Y.; Xie, Y.; Ma, F.; Liang, J.; Li, C. Exogenous GABA improves the resistance of apple seedlings to long-term drought stress by enhancing GABA shunt and secondary cell wall biosynthesis. Tree Physiol. 2022, 42, 2563–2577. [Google Scholar] [CrossRef]
- Wang, S.S.; Liu, X.J.; Hu, Y.; Ji, Y.L.; Bai, T.; Zhu, M.X.; Zhang, Y.H. Metabolism and enrichment mechanism of γ-aminobutyric acid in plants. J. Anhui Agric. Sci. 2020, 48, 9–12. [Google Scholar]
- Uzma Jalil, S.; Khan, M.I.R.; Ansari, M.I. Role of GABA transaminase in the regulation of development and senescence in Arabidopsis thaliana. Curr. Plant Biol. 2019, 19, 100119. [Google Scholar] [CrossRef]
- Fromm, H. GABA signaling in plants: Targeting the missing pieces of the puzzle. J. Exp. Bot. 2020, 71, 6238–6245. [Google Scholar] [CrossRef] [PubMed]
- Kaspal, M.; Kanapaddalagamage, M.H.; Ramesh, S.A. Emerging roles of γ aminobutyric acid (GABA) gated channels in plant stress tolerance. Plants 2021, 10, 2178. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.B.A.; Ashrafuzzaman, M.U.P.M.; Ismail, M.R.U.P.; Shahidullah, M.S.U.P.; Prodhan, A.K.M.A. Influence of foliar applied GABA on growth and yield contributing characters of white gourd (Benincasa hispida). Int. J. Agric. Biol. 2010, 12, 373–376. [Google Scholar]
- Hijaz, F.; Nehela, Y.; Killiny, N. Application of gamma-aminobutyric acid increased the level of phytohormones in Citrus sinensis. Planta 2018, 248, 909–918. [Google Scholar] [CrossRef]
- Xie, T.; Ji, J.; Chen, W.; Yue, J.; Du, C.; Sun, J.; Chen, L.; Jiang, Z.; Shi, S. GABA negatively regulates adventitious root development in poplar. J. Exp. Bot. 2020, 71, 1459–1474. [Google Scholar] [CrossRef]
- Renault, H.; Amrani, A.E.; Palanivelu, R.; Updegraff, E.P.; Yu, A.; Renou, J.; Preuss, D.; Bouchereau, A.; Deleu, C. GABA accumulation causes cell elongation defects and a decrease in expression of genes encoding secreted and cell wall-related proteins in Arabidopsis thaliana. Plant Cell Physiol. 2011, 52, 894–908. [Google Scholar] [CrossRef]
- Renault, H. Phylogeny and bioinformatics shed light on GABA functions in plants. Plant Signal Behav. 2013, 8, e24274. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, P.; Chen, Z.; Gu, Z.; Yang, R. GABA enhances physio-biochemical metabolism and antioxidant capacity of germinated hulless barley under NaCl stress. J. Plant Physiol. 2018, 231, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Batushansky, A.; Kirma, M.; Grillich, N.; Pham, P.A.; Rentsch, D.; Galili, G.; Fernie, A.R.; Fait, A. The transporter GAT1 plays an important role in GABA-mediated carbon-nitrogen interactions in Arabidopsis. Front. Plant Sci. 2015, 6, 785. [Google Scholar] [CrossRef] [PubMed]
- Wang, H. Response of Melatonin at Different Concentrations to Loquat Seedlings under Cold Stress; Southwest University: Chongqing, China, 2020. [Google Scholar]
- Liu, Y.; Zhang, W.; Xu, C.; Li, X. Biological activities of extracts from loquat (Eriobotrya japonica Lindl.): A review. Int. J. Mol. Sci. 2016, 17, 1983. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agricultural Chemistry Analysis; Chinese Agricultural Press: Beijing, China, 2000. [Google Scholar]
- Fan, H.; Zhang, Z.Y.; Tan, Y.; Zhou, X.F.; Xu, H.W. Effect of external GABA on agronomic traits and photosynthetic characteristics of maize seedlings. Shandong Agric. Sci. 2022, 54, 67–72, 86. [Google Scholar]
- Li, Y.; Liu, B.; Peng, Y.; Liu, C.; Zhang, X.; Zhang, Z.; Liang, W.; Ma, F.; Li, C. Exogenous GABA alleviates alkaline stress in Malus hupehensis by regulating the accumulation of organic acids. Sci. Hortic. 2020, 261, 108982. [Google Scholar] [CrossRef]
- Hao, Z.B.; Cang, J.; Xu, Z. Plant Physiology Experiment; Harbin Institute of Technology Press: Harbin, China, 2004. [Google Scholar]
- Li, Z.; Fan, R.; Peng, X.; Shu, J.; Liu, L.; Wang, J.; Lin, L. Salicylic acid alleviates selenium stress and promotes selenium uptake of grapevine. Physiol. Mol. Biol. Plants 2022, 28, 625–635. [Google Scholar] [CrossRef]
- Zhang, R.; Huang, Y.; Liu, Y.; Liu, Q.; Zhang, L.; Li, Z.; Xu, Y.; Lin, L.; Wang, L. Effects of mutual intercropping on cadmium accumulation of Solanum photeinocarpum Nakamura et Odashima and its post-grafting generations. Int. J. Phytoremediat. 2023, 25, 350–358. [Google Scholar] [CrossRef]
- Lin, L.; Li, Z.; Wang, J.; Liang, D.; Xia, H.; Lv, X.; Tang, Y.; Wang, X.; Deng, Q.; Liao, M. 24-epibrassinolide promotes selenium uptake in grapevine under selenium stress. Sci. Hortic. 2023, 308, 111564. [Google Scholar] [CrossRef]
- Shelp, B.J.; Mullen, R.T.; Waller, J.C. Compartmentation of GABA metabolism raises intriguing questions. Trends Plant Sci. 2012, 17, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Li, M.F.; Guo, S.J.; Yang, X.H.; Meng, Q.W.; Wei, X.J. Exogenous gamma-aminobutyric acid increases salt tolerance of wheat by improving photosynthesis and enhancing activities of antioxidant enzymes. Biol. Plant 2016, 60, 123–131. [Google Scholar] [CrossRef]
- Guo, Z.; Du, N.; Li, Y.; Zheng, S.; Shen, S.; Piao, F. Gamma-aminobutyric acid enhances tolerance to iron deficiency by stimulating auxin signaling in cucumber (Cucumis sativus L.). Ecotoxicol. Environ. Saf. 2020, 192, 110285. [Google Scholar] [CrossRef] [PubMed]
- Bashir, R.; Riaz, H.N.; Anwar, S.; Parveen, N.; Khalilzadeh, R.; Hussain, I.; Mahmood, S. Morpho-physiological changes in carrots by foliar γ-aminobutyric acid under drought stress. Rev. Bras. Dotânica 2020, 44, 57–68. [Google Scholar] [CrossRef]
- Zhang, S.S.; Yang, W.Z.; Kang, H.M.; Nuosu, N. Effects of light intensities and water conditions on growth and photosynthetic characteristics of Nyssa yun-nanensis seedlings. J. Northeast. For. Univ. 2018, 46, 16–23. [Google Scholar]
- Kalhor, M.S.; Aliniaeifard, S.; Seif, M.; Asayesh, E.J.; Bernard, F.; Hassani, B.; Li, T. Enhanced salt tolerance and photosynthetic performance: Implication of ɤ-amino butyric acid application in salt-exposed lettuce (Lactuca sativa L.) plants. Plant Physiol. Biochem. 2018, 130, 157–172. [Google Scholar] [CrossRef]
- Xiang, L.; Hu, L.; Xu, W.; Zhen, A.; Zhang, L.; Hu, X.; Shi, H. Exogenous γ-aminobutyric acid improves the structure and function of photosystem II in muskmelon seedlings exposed to salinity-alkalinity stress. PLoS ONE 2016, 11, e164847. [Google Scholar] [CrossRef]
- Salah, A.; Zhan, M.; Cao, C.; Han, Y.; Ling, L.; Liu, Z.; Li, P.; Ye, M.; Jiang, Y. γ-aminobutyric acid promotes chloroplast ultrastructure, antioxidant capacity, and growth of waterlogged maize seedlings. Sci. Rep. 2019, 9, 484. [Google Scholar] [CrossRef]
- Xia, Q.P.; Gao, H.B.; Li, J.R. Effects of gamma-aminobutyric acid on the photosynthesis and chlorophyll fluorescence parameters of muskmelon seedlings under hypoxia stress. Chin. J. Appl. Ecol. 2011, 22, 999–1006. [Google Scholar]
- Xiang, L.X. The Study of Exogenous γ-aminobutyric Acid Improves Structure and Function of Photosynthetic Apparatus in Muskmelon Seedlings Exposed to Salinity-Alkalinity Stress; Northwest A & F University: Yangling, China, 2016. [Google Scholar]
- Wang, J.; Xu, S.; Yan, T.; Ma, W.J.; Yan, Q.L. Effects of soil nutrients on seedling growth of major tree species in montane region of eastern Liaoning Province, China. Chin. J. Ecol. 2017, 36, 3148–3159. [Google Scholar]
- Lv, N.N.; Shen, Z.Z.; Wang, D.S.; Liu, H.J.; Xue, C.; Li, R.; Shen, Q.R. Effects of amino acid organic fertilizer on cucumber yield and soil biological characters. J. Nanjing Agric. Univ. 2018, 41, 456–464. [Google Scholar]
- Renault, H.; El, A.A.; Berger, A.; Mouille, G.; Soubigou-Taconnat, L.; Bouchereau, A.; Deleu, C. Gamma-aminobutyric acid transaminase deficiency impairs central carbon metabolism and leads to cell wall defects during salt stress in Arabidopsis roots. Plant Cell Environ. 2013, 36, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tian, Z.; Wu, X.; Lv, G.; Ma, W.; Zhang, Y.; Gao, H. Gamma-aminobutyric acid (GABA) modulates nitrate concentrations and metabolism in the leaves of pakchoi (Brassica campestris ssp. chinensis Makino) treated with a nitrogen-rich solution. Plant Mol. Biol. Report. 2018, 36, 530–542. [Google Scholar] [CrossRef]
- Wang, Y.C. Mechanism of Aminobutyric acid (GABA) Regulating Maize Seed Germination and Seedling Growth under Salt Stress; Northeast Agricultural University: Harbin, China, 2016. [Google Scholar]
- Carillo, P. GABA shunt in durum wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ma, X.X.; Jia, D.C.; Zhou, D.Y.; Peng, S.M.; Wang, Y.S. Progress in research on kiwifruit rootstocks. North. Hortic. 2022, 46, 125–133. [Google Scholar]
- Zhao, Y.; Song, X.; Zhong, D.B.; Yu, L.; Yu, X. Gamma-aminobutyric acid (GABA) regulates lipid production and cadmium uptake by Monoraphidium sp. QLY-1 under cadmium stress. Bioresour. Technol. 2020, 297, 122500. [Google Scholar] [CrossRef]
- Song, S.L.; Li, J.R.; Gao, H.B.; Li, Q.Y.; Yang, L.W.; Gong, R.J. Effects of exogenous γ-aminobutyric acid on inorganic nitrogen metabolism and mineral elements contents of melon seedling under hypoxia stress. Acta Hortic. Sin. 2012, 39, 695–704. [Google Scholar]
- Zhou, X. Physiological Effects of Salt Stress-Induced GABA Accumulation in Maize Seedlings; China Agricultural University: Beijing, China, 2004. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GABA Concentration (g L−1) | Chlorophyll a (mg g−1) | Chlorophyll b (mg g−1) | Total Chlorophyll (mg g−1) | Chlorophyll a/b | Carotenoid (mg g−1) |
|---|---|---|---|---|---|
| 0.0 | 0.317 ± 0.011 c | 0.175 ± 0.013 c | 0.491 ± 0.019 c | 1.816 ± 0.126 a | 0.041 ± 0.001 c |
| 0.5 | 0.379 ± 0.019 a | 0.246 ± 0.010 a | 0.625 ± 0.016 a | 1.543 ± 0.123 b | 0.063 ± 0.002 a |
| 1.0 | 0.386 ± 0.014 a | 0.222 ± 0.008 b | 0.607 ± 0.007 a | 1.743 ± 0.128 ab | 0.068 ± 0.003 a |
| 1.5 | 0.344 ± 0.009 b | 0.219 ± 0.006 b | 0.563 ± 0.005 b | 1.570 ± 0.079 b | 0.052 ± 0003 b |
| 2.0 | 0.322 ± 0.010 bc | 0.188 ± 0.006 c | 0.509 ± 0.005 c | 1.718 ± 0.104 ab | 0.050 ± 0.003 b |
| GABA Concentration (g L−1) | Soil pH Value | Alkali-Hydrolyzable N Concentration (mg kg−1) | Available P Concentration (mg kg−1) | Available K Concentration (mg kg−1) |
|---|---|---|---|---|
| 0.0 | 7.933 ± 0.055 a | 61.80 ± 1.45 c | 16.97 ± 0.96 b | 64.70 ± 1.09 a |
| 0.5 | 7.490 ± 0.079 c | 68.55 ± 0.58 a | 17.31 ± 0.17 b | 62.21 ± 0.82 ab |
| 1.0 | 7.263 ± 0.047 e | 70.29 ± 0.56 a | 22.23 ± 1.14 a | 58.10 ± 1.64 ab |
| 1.5 | 7.373 ± 0.055 d | 69.50 ± 1.65 a | 18.18 ± 0.56 b | 54.85 ± 0.82 b |
| 2.0 | 7.593 ± 0.021 b | 65.23 ± 1.46 b | 18.03 ± 1.51 b | 66.69 ± 0.91 ab |
| Indicators | RB | SB | Cha | Chb | Cha/b | Car | RN | SN | RP | SP | RK | SK | pH | AN | AP | AK |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RB | ||||||||||||||||
| SB | 0.757 ** | |||||||||||||||
| Cha | 0.833 ** | 0.761 ** | ||||||||||||||
| Chb | 0.740 ** | 0.562 * | 0.750 ** | |||||||||||||
| Cha/b | −0.309 | −0.093 | −0.128 | −0.747 ** | ||||||||||||
| Car | 0.876 ** | 0.852 ** | 0.911 ** | 0.776 ** | −0.269 | |||||||||||
| RN | 0.852 ** | 0.835 ** | 0.908 ** | 0.867 ** | −0.384 | 0.914 ** | ||||||||||
| SN | 0.885 ** | 0.836 ** | 0.870 ** | 0.762 ** | −0.299 | 0.976 ** | 0.900 ** | |||||||||
| RP | 0.895 ** | 0.821 ** | 0.819 ** | 0.770 ** | −0.353 | 0.943 ** | 0.884 ** | 0.945 ** | ||||||||
| SP | 0.904 ** | 0.878 ** | 0.848 ** | 0.786 ** | −0.345 | 0.880 ** | 0.929 ** | 0.889 ** | 0.897 ** | |||||||
| RK | 0.746 ** | 0.866 ** | 0.845 ** | 0.562 * | −0.008 | 0.881 ** | 0.843 ** | 0.897 ** | 0.837 ** | 0.837 ** | ||||||
| SK | 0.707 ** | 0.614 * | 0.695 ** | 0.703 ** | −0.381 | 0.818 ** | 0.760 ** | 0.830 ** | 0.802 ** | 0.662 ** | 0.630 * | |||||
| pH | −0.921 ** | −0.731 ** | −0.697 ** | −0.700 ** | 0.391 | −0.803 ** | −0.800 ** | −0.836 ** | −0.857 ** | −0.878 ** | −0.679 ** | −0.624 * | ||||
| AN | 0.885 ** | 0.684 ** | 0.718 ** | 0.764 ** | −0.451 | 0.797 ** | 0.829 ** | 0.815 ** | 0.826 ** | 0.893 ** | 0.717 ** | 0.578 * | −0.925 ** | |||
| AP | 0.608 * | 0.840 ** | 0.522 * | 0.238 | 0.136 | 0.630 * | 0.565 * | 0.641 * | 0.664 ** | 0.702 ** | 0.767 ** | 0.357 | −0.625 * | 0.521 * | ||
| AK | 0.754 ** | 0.597 * | 0.423 | 0.320 | −0.121 | 0.542 * | 0.501 | 0.594 * | 0.675 ** | 0.679 ** | 0.542 * | 0.420 | −0.828 ** | 0.740 ** | 0.698 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Xu, Y.; Song, P.; Li, X.; Zhou, J.; Lin, L.; Xia, H.; Liang, D.; Luo, X.; Zhang, H.; et al. Effects of Gamma Amino Butyric Acid (GABA) on Nutrient Uptake of Loquat [Eriobotrya japonica (Thunb.) Lindl.] Seedlings. Horticulturae 2023, 9, 196. https://doi.org/10.3390/horticulturae9020196
Yang Z, Xu Y, Song P, Li X, Zhou J, Lin L, Xia H, Liang D, Luo X, Zhang H, et al. Effects of Gamma Amino Butyric Acid (GABA) on Nutrient Uptake of Loquat [Eriobotrya japonica (Thunb.) Lindl.] Seedlings. Horticulturae. 2023; 9(2):196. https://doi.org/10.3390/horticulturae9020196
Chicago/Turabian StyleYang, Zhiwu, Yaxin Xu, Panhui Song, Xinyu Li, Jiayun Zhou, Lijin Lin, Hui Xia, Dong Liang, Xian Luo, Huifen Zhang, and et al. 2023. "Effects of Gamma Amino Butyric Acid (GABA) on Nutrient Uptake of Loquat [Eriobotrya japonica (Thunb.) Lindl.] Seedlings" Horticulturae 9, no. 2: 196. https://doi.org/10.3390/horticulturae9020196
APA StyleYang, Z., Xu, Y., Song, P., Li, X., Zhou, J., Lin, L., Xia, H., Liang, D., Luo, X., Zhang, H., Deng, Q., & Wang, Y. (2023). Effects of Gamma Amino Butyric Acid (GABA) on Nutrient Uptake of Loquat [Eriobotrya japonica (Thunb.) Lindl.] Seedlings. Horticulturae, 9(2), 196. https://doi.org/10.3390/horticulturae9020196