


Photosynthesis Characteristics of Tomato Plants and Its’ Responses to Microclimate in New Solar Greenhouse in North China



Abstract
1. Introduction
2. Materials and Methods
2.1. Greenhouse and Tomato Cultivation Description
2.2. Plant Growth and Yield Measurement
2.3. Photosynthesis Characteristics Measurement
2.4. Microclimate Measurement
2.5. Data Treatment and Figure Preparation
3. Results
3.1. Tomato Growth and Yield
3.2. Microclimate Changes in the Greenhouse
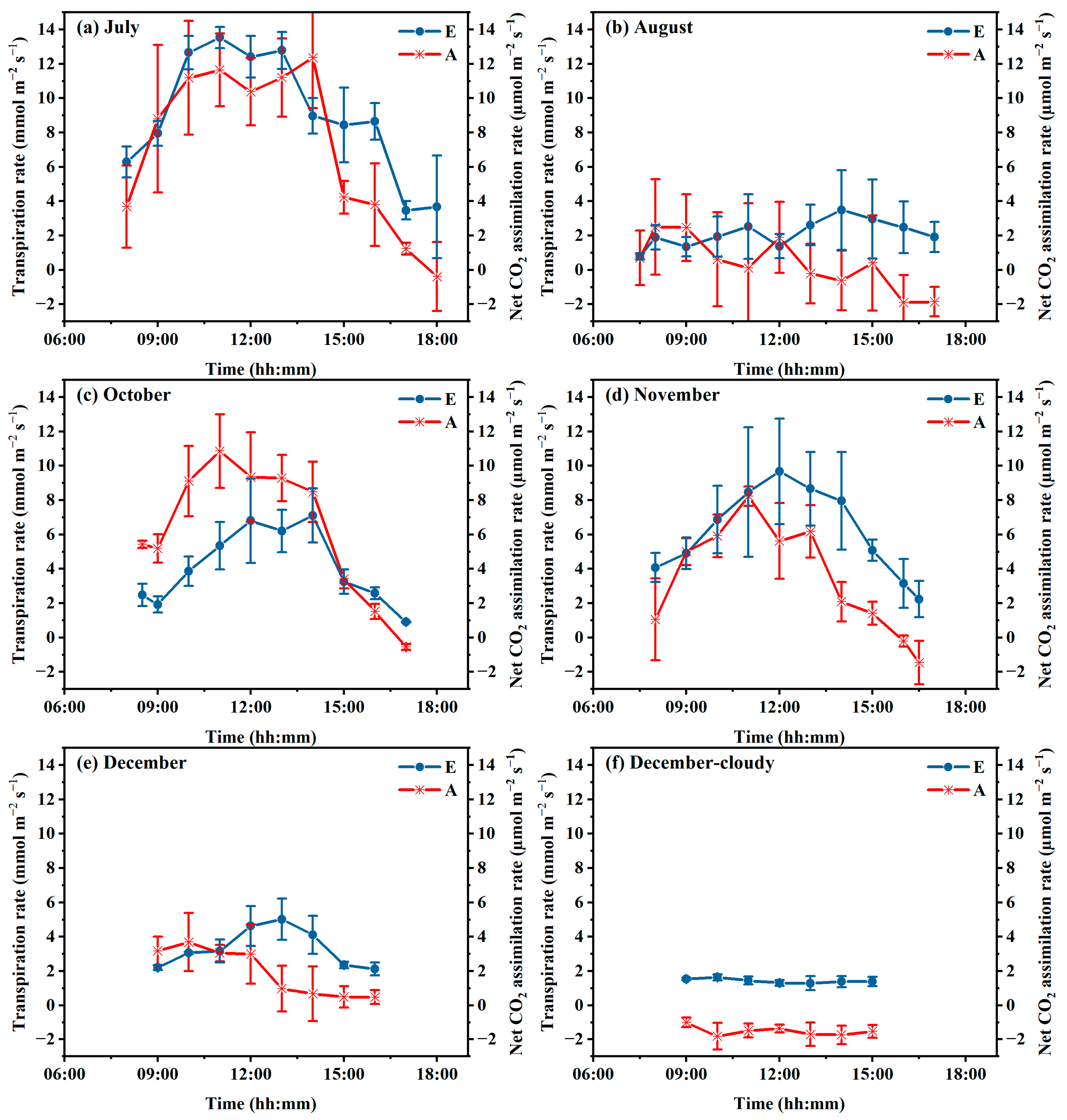
3.3. Photosynthesis Characteristic Changes
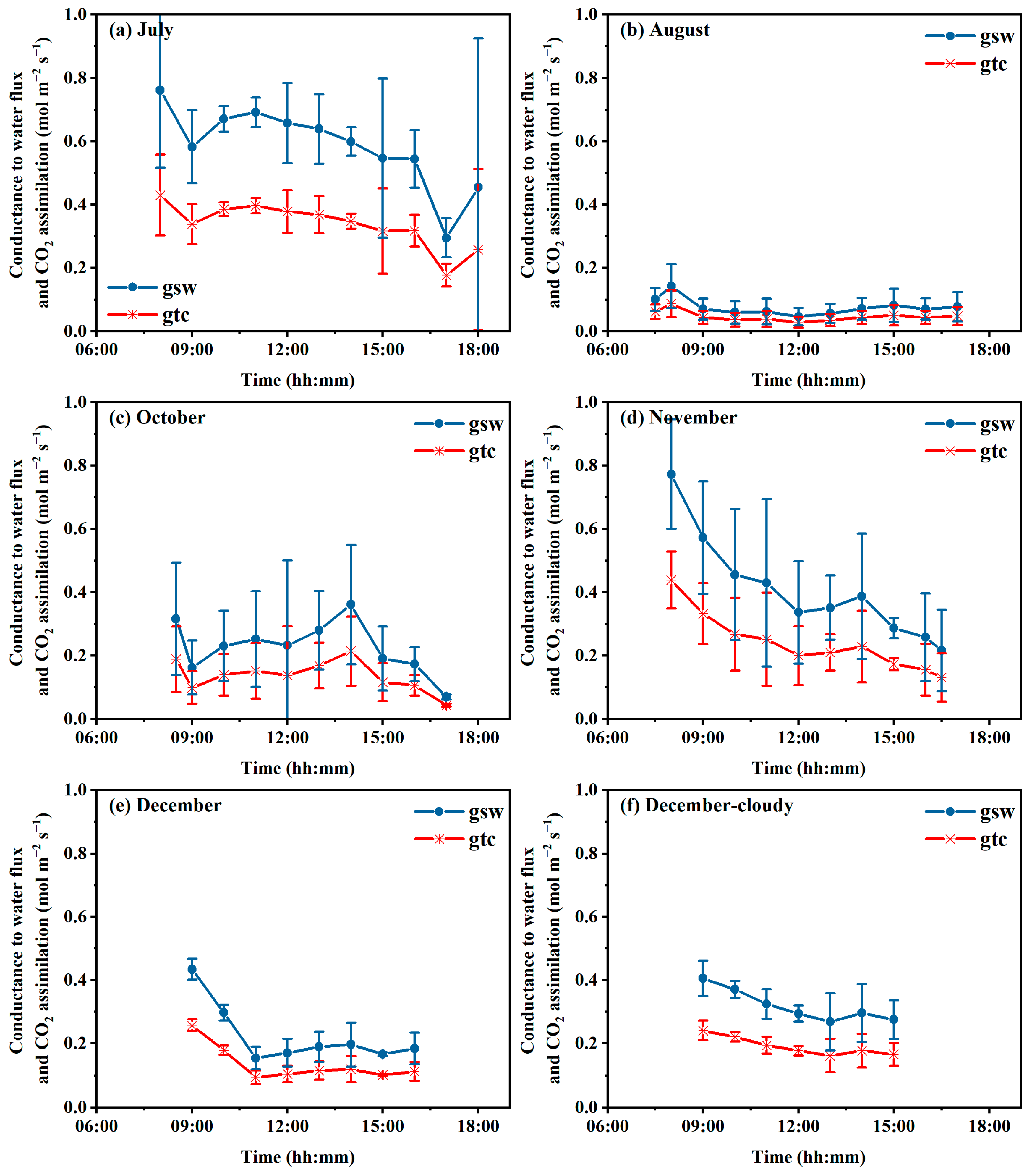
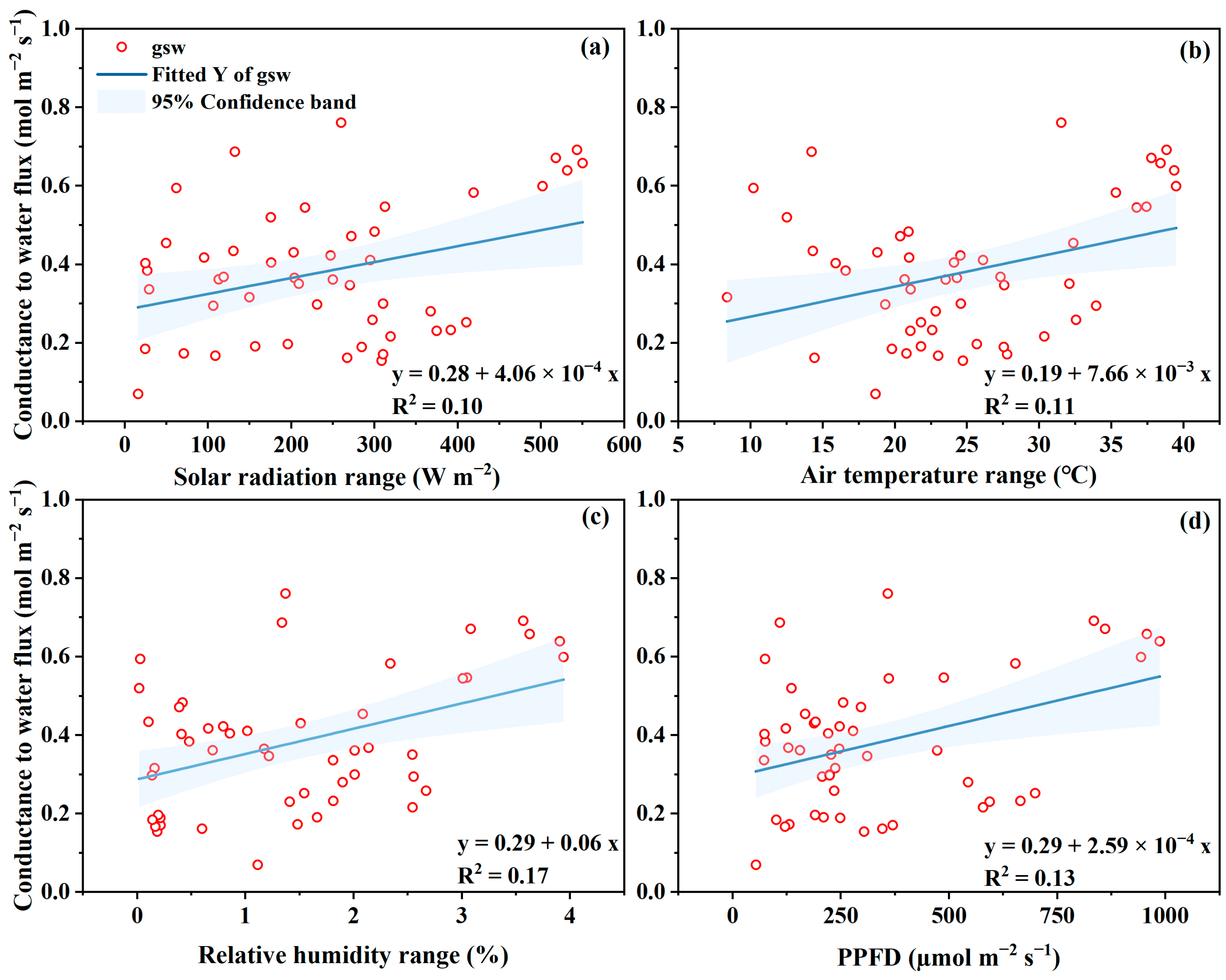
3.4. Conductance to Transpiration (gsw) and Photosynthesis (gtc)
4. Discussion
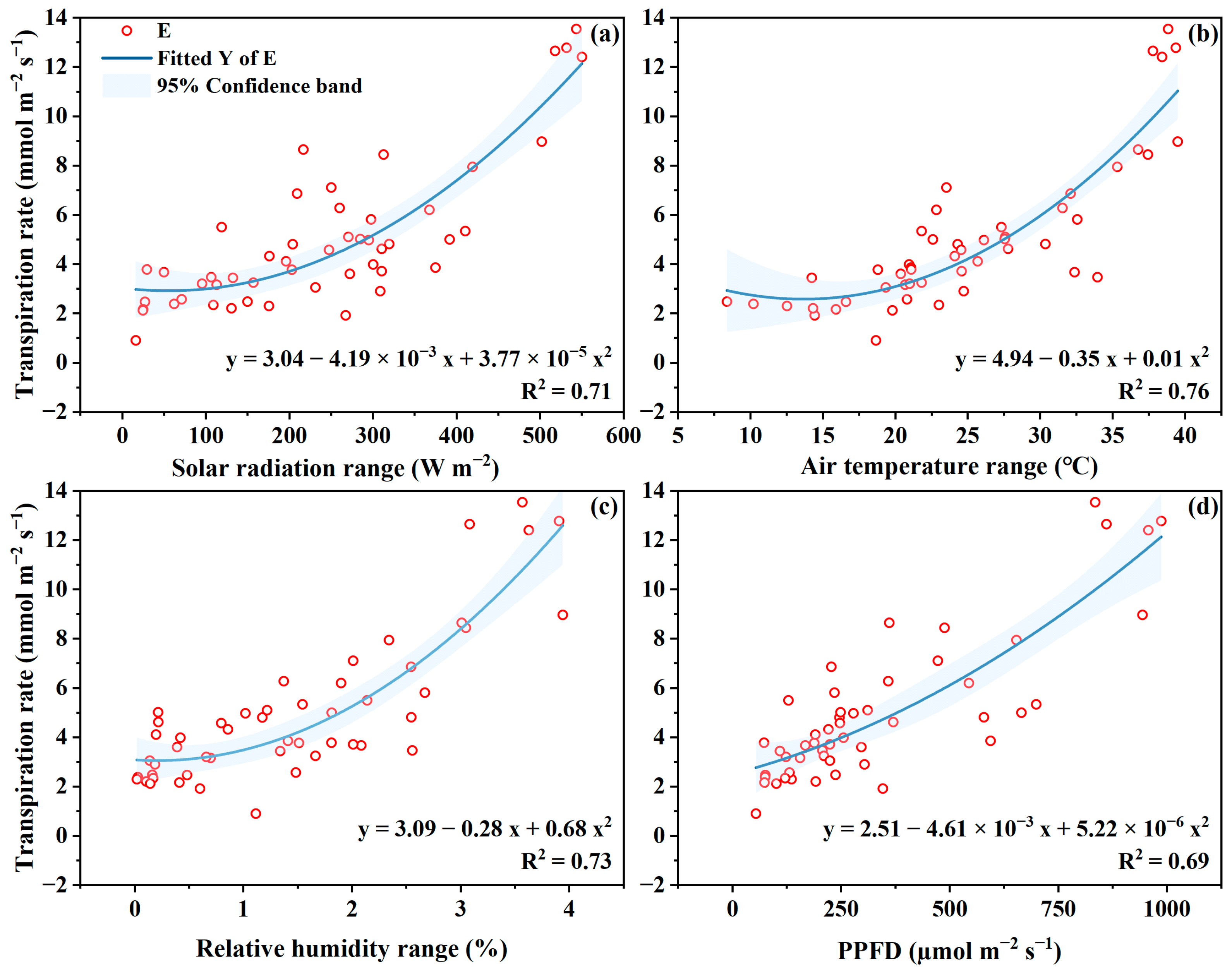
4.1. Transpiration Rate to Microclimate
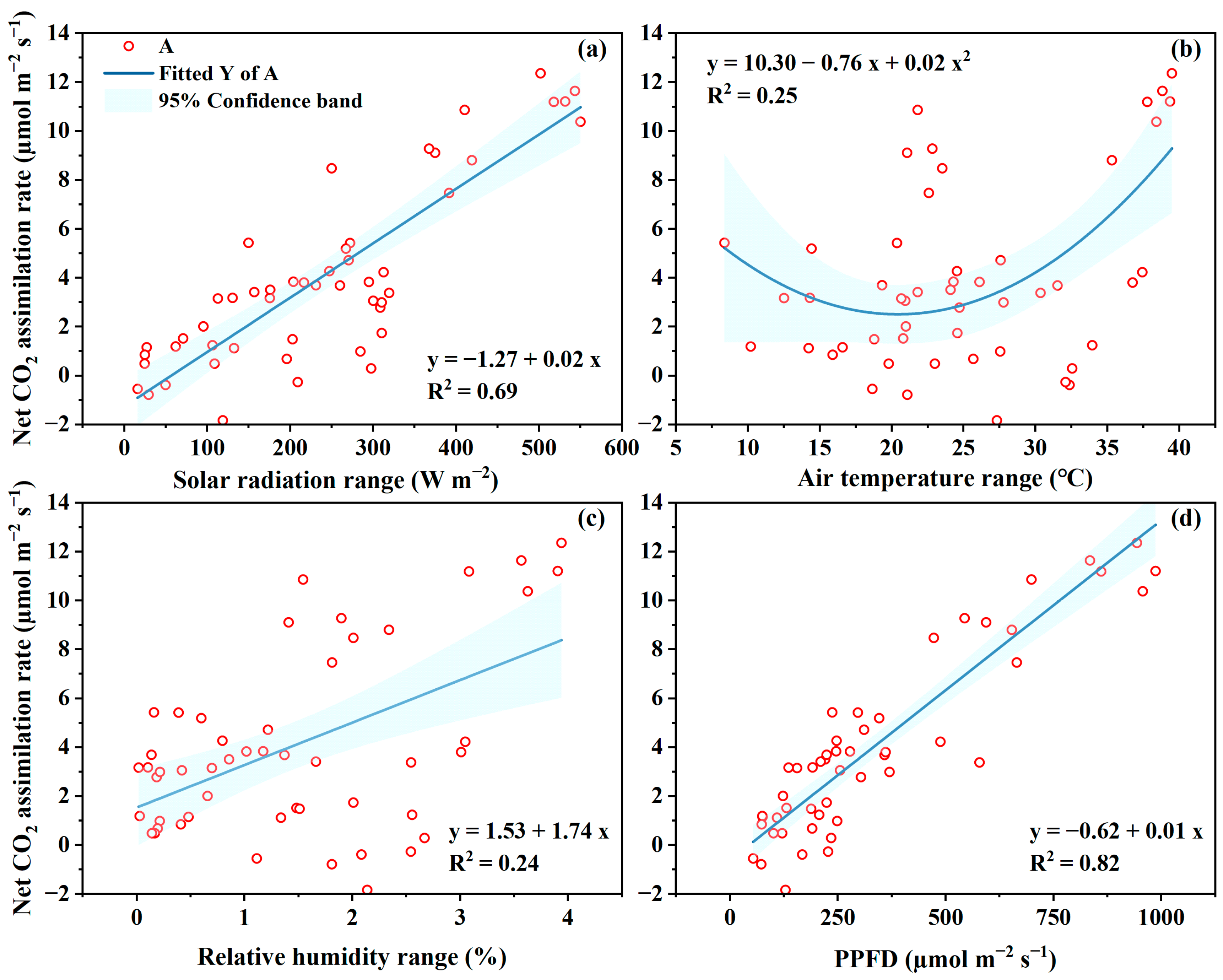
4.2. Photosynthesis Rate to Microclimate
4.3. Conductance of gsw and gtc to Microclimate
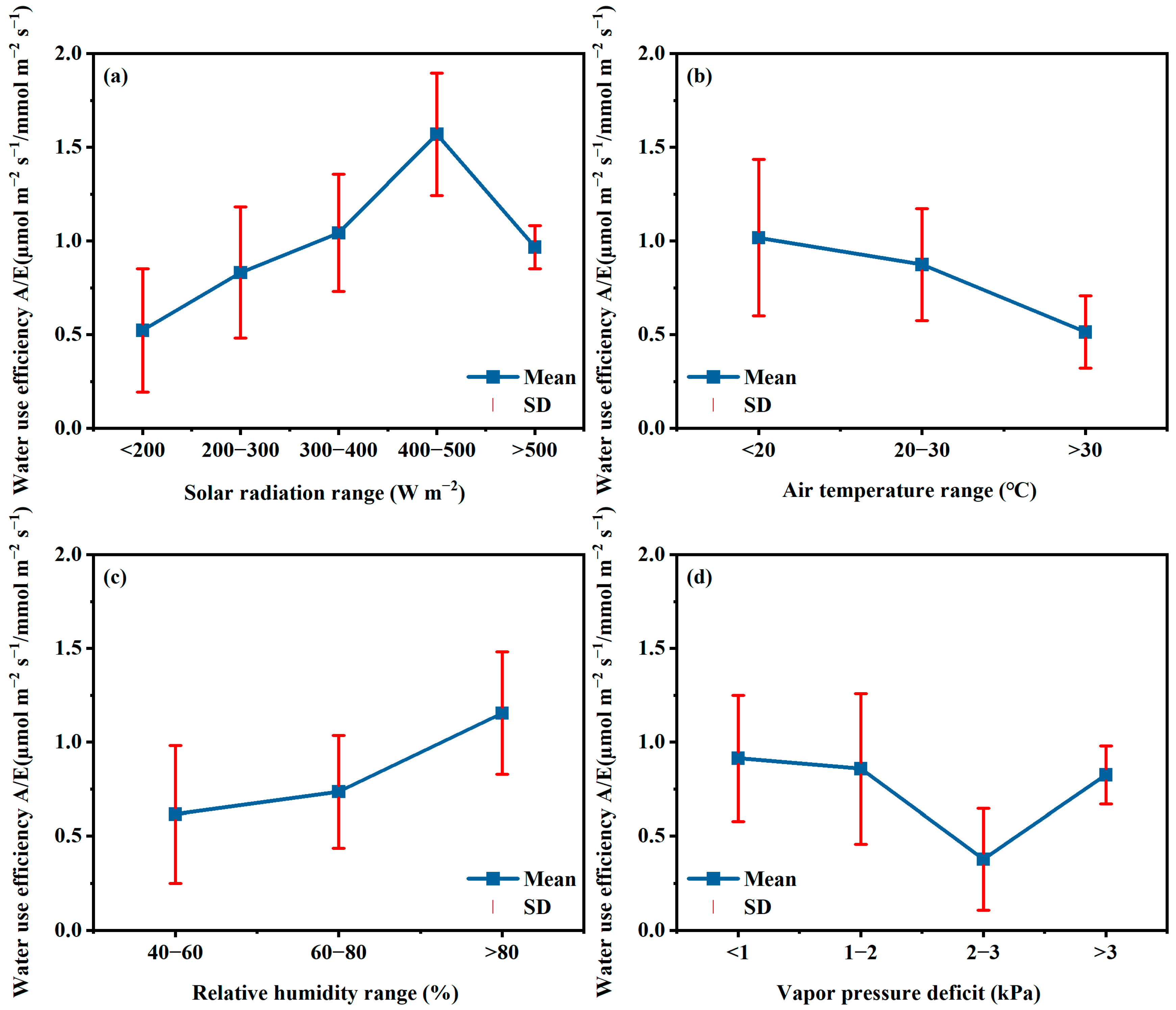
4.4. Microclimate Management for High Water Productivity in GH
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yeo, U.-H.; Lee, S.-Y.; Park, S.-J.; Kim, J.-G.; Choi, Y.-B.; Kim, R.-W.; Shin, J.H.; Lee, I.-B. Rooftop Greenhouse: (1) Design and Validation of a BES Model for a Plastic-Covered Greenhouse Considering the Tomato Crop Model and Natural Ventilation Characteristics. Agriculture 2022, 12, 903. [Google Scholar] [CrossRef]
- Bonachela, S.; Fernández, M.D.; Cabrera-Corral, F.J.; Granados, M.R. Salt and irrigation management of soil-grown Mediterranean greenhouse tomato crops drip-irrigated with moderately saline water. Agric. Water Manag. 2022, 262, 107433. [Google Scholar] [CrossRef]
- Sánchez-Hermosilla, J.; Pérez-Alonso, J.; Martínez-Carricondo, P.; Carvajal-Ramírez, F.; Agüera-Vega, F. Evaluation of Electrostatic Spraying Equipment in a Greenhouse Pepper Crop. Horticulturae 2022, 8, 541. [Google Scholar] [CrossRef]
- Hodge, C.; Rogers, M.; Handeen, D.; Schweser, G. Yield of leafy greens and microclimate in deep winter greenhouse production in Minnesota. Sustainability 2019, 11, 28. [Google Scholar] [CrossRef]
- Wacker, J.-D.; Verheul, M.J.; Righini, I.; Maessen, H.; Stanghellini, C. Optimisation of supplemental light systems in Norwegian tomato greenhouses—A simulation study. Biosyst. Eng. 2022, 215, 129–142. [Google Scholar] [CrossRef]
- Roonjho, S.J.; Kamal, R.M.; Roonjho, A.R. Modeling capillary wick irrigation system for greenhouse crop production. Agric. Water Manag. 2022, 274, 107927. [Google Scholar] [CrossRef]
- Ávila-Pozo, P.; Parrado, J.; Caballero, P.; Tejada, M. Use of a Biostimulant Obtained from Slaughterhouse Sludge in a Greenhouse Tomato Crop. Horticulturae 2022, 8, 622. [Google Scholar] [CrossRef]
- Yang, P.; Bai, J.; Yang, M.; Ma, E.; Yan, M.; Long, H.; Liu, J.; Li, L. Negative pressure irrigation for greenhouse crops in China: A review. Agric. Water Manag. 2022, 264, 107497. [Google Scholar] [CrossRef]
- Nikolaou, G.; Neocleous, D.; Christou, A.; Polycarpou, P.; Kitta, E.; Katsoulas, N. Energy and Water Related Parameters in Tomato and Cucumber Greenhouse Crops in Semiarid Mediterranean Regions. A Review, Part I: Increasing Energy Efficiency. Horticulturae 2021, 7, 521. [Google Scholar] [CrossRef]
- Nikolaou, G.; Neocleous, D.; Christou, A.; Polycarpou, P.; Kitta, E.; Katsoulas, N. Energy and Water Related Parameters in Tomato and Cucumber Greenhouse Crops in Semiarid Mediterranean Regions. A Review, Part II: Irrigation and Fertigation. Horticulturae 2021, 7, 548. [Google Scholar] [CrossRef]
- Liu, T.; Yuan, Q.; Wang, Y. Hierarchical optimization control based on crop growth model for greenhouse light environment. Comput. Electron. Agric. 2021, 180, 105854. [Google Scholar] [CrossRef]
- Liu, H.; Yin, C.; Hu, X.; Tanny, J.; Tang, X. Microclimate characteristics and evapotranspiration estimates of cucumber plants in a newly developed sunken solar greenhouse. Water 2020, 12, 2275. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Xu, C.; Wang, M.T.; Zhao, H.L.; Zheng, Y.J.; Huang, H.J.; Vuguziga, F.; Umutoni, M.A. Enhancing the thermotolerance of tomato seedlings by heat shock treatment. Photosynthetica 2019, 57, 1184–1192. [Google Scholar] [CrossRef]
- Xu, D.; Du, S.; van Willigenburg, L.G. Optimal control of Chinese solar greenhouse cultivation. Biosyst. Eng. 2018, 171, 205–219. [Google Scholar] [CrossRef]
- Shaheen, M.R.; Ayyub, C.M.; Amjad, M.; Waraich, E.A. Morpho-physiological evaluation of tomato genotypes under high temperature stress conditions: Evaluation of tomato genotypes under high temperature stress. J. Sci. Food Agric. 2016, 96, 2698–2704. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Y.; Tian, S.; Liang, Z.; Li, S.; Li, Y.; Wei, M.; Zhang, D. Effect of supplemental lighting on water transport, photosynthetic carbon gain and water use efficiency in greenhouse tomato. Sci. Hortic.-Amst. 2019, 256, 108630. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Angeles Morales, M.; Miguel Dell’Amico, J.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Alsamir, M.; Mahmood, T.; Trethowan, R.; Ahmad, N. An overview of heat stress in tomato (Solanum lycopersicum L.). Saudi J. Biol. Sci. 2021, 28, 1654–1663. [Google Scholar] [CrossRef]
- Alsamir, M.; Ahmad, N.; Mahmood, T.; Trethowan, R. Morpho-Physiological Traits Linked to High Temperature Stress Tolerance in Tomato (S. lycopersicum L.). Am. J. Plant Sci. 2017, 8, 2681–2694. [Google Scholar] [CrossRef]
- Buttaro, D.; Santamaria, P.; Signore, A.; Cantore, V.; Boari, F.; Montesano, F.F.; Parente, A. Irrigation management of greenhouse tomato and cucumber using tensiometer: Effects on yield, quality and water use. Agric. Agric. Sci. Procedia 2015, 4, 440–444. [Google Scholar] [CrossRef]
- Juárez-Maldonado, A.; Benavides-Mendoza, A.; de-Alba-Romenus, K.; Morales-Díaz, A.B. Estimation of the water requirements of greenhouse tomato crop using multiple regression models. Emir. J. Food Agric. 2014, 26, 885–897. [Google Scholar] [CrossRef]
- Du, Y.; Cao, H.; Liu, S.; Gu, X.; Cao, Y. Response of yield, quality, water and nitrogen use efficiency of tomato to different levels of water and nitrogen under drip irrigation in Northwestern China. J. Integr. Agric. 2017, 16, 1153–1161. [Google Scholar] [CrossRef]
- Ntinas, G.K.; Koukounaras, A.; Kotsopoulos, T. Effect of energy saving solar sleeves on characteristics of hydroponic tomatoes grown in a greenhouse. Sci. Hortic.-Amst. 2015, 194, 126–133. [Google Scholar] [CrossRef]
- Gourdo, L.; Fatnassi, H.; Bouharroud, R.; Ezzaeri, K.; Bazgaou, A.; Wifaya, A.; Demrati, H.; Bekkaoui, A.; Aharoune, A.; Poncet, C.; et al. Heating canarian greenhouse with a passive solar water–sleeve system: Effect on microclimate and tomato crop yield. Sol. Energy 2019, 188, 1349–1359. [Google Scholar] [CrossRef]
- Ni, M.; Lan, D.; Jahan, M.; Wang, J.; Guo, S. A pilot study on the microclimate of a multi-span solar energy greenhouse. Appl. Eng. Agric. 2019, 35, 601–616. [Google Scholar] [CrossRef]
- Li, A.; Huang, L.; Zhang, T. Field test and analysis of microclimate in naturally ventilated single-sloped greenhouses. Energy Build. 2017, 138, 479–489. [Google Scholar] [CrossRef]
- Liu, H.; Yuan, B.; Hu, X.; Yin, C.; Tang, X. Cucumber production and the economic revenues under various nitrogen applications in an unheated solar greenhouse on the North China Plain. Agron. J. 2021, 113, 3444–3459. [Google Scholar] [CrossRef]
- Fu, G.; Li, Z.; Liu, W.; Yang, Q. Improved root zone temperature buffer capacity enhancing sweet pepper yield via soil-ridged substrate-embedded cultivation in solar greenhouse. Int. J. Agric. Biol. Eng. 2018, 11, 41–47. [Google Scholar] [CrossRef]
- Li, Y.; Xue, X.; Xu, F.; Guo, W.; Duan, M.; Lin, S.; Li, Y.; Wang, Z. Negative-pressure irrigation improves water and fertilizer use efficiencies and fruit yield of greenhouse tomato on the North China Plain. Irrig. Drain. 2021, 70, 1027–1038. [Google Scholar] [CrossRef]
- Liu, H.; Yin, C.; Gao, Z.; Hou, L. Evaluation of cucumber yield, economic benefit and water productivity under different soil matric potentials in solar greenhouses in North China. Agric. Water Manag. 2021, 243, 106442. [Google Scholar] [CrossRef]
- Xu, K.; Guo, X.; He, J.; Yu, B.; Tan, J.; Guo, Y. A study on temperature spatial distribution of a greenhouse under solar load with considering crop transpiration and optical effects. Energy Convers. Manag. 2022, 254, 115277. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Ning, H.; Zhang, X.; Li, S.; Pang, J.; Wang, G.; Sun, J. Optimizing irrigation frequency and amount to balance yield, fruit quality and water use efficiency of greenhouse tomato. Agric. Water Manag. 2019, 226, 105787. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, Q.; Zheng, K.; Zhao, S.; Wang, P.; Cheng, J.; Zhang, X.; Chen, X. Effects of Diffuse Light on Microclimate of Solar Greenhouse, and Photosynthesis and Yield of Greenhouse-grown Tomatoes. HortScience 2020, 55, 1605–1613. [Google Scholar] [CrossRef]
- Yang, L.; Liu, H.; Cohen, S.; Gao, Z. Microclimate and Plant Transpiration of Tomato (Solanum lycopersicum L.) in a Sunken Solar Greenhouse in North China. Agriculture 2022, 12, 260. [Google Scholar] [CrossRef]
- Kimura, K.; Yasutake, D.; Koikawa, K.; Kitano, M. Spatiotemporal variability of leaf photosynthesis and its linkage with microclimates across an environment-controlled greenhouse. Biosyst. Eng. 2020, 195, 97–115. [Google Scholar] [CrossRef]
- Li, Q.; Wei, M.; Li, Y.; Feng, G.; Wang, Y.; Li, S.; Zhang, D. Effects of soil moisture on water transport, photosynthetic carbon gain and water use efficiency in tomato are influenced by evaporative demand. Agric. Water Manag. 2019, 226, 105818. [Google Scholar] [CrossRef]
- Islam, M.T. Effect of temperature on photosynthesis, yield attributes and yield of tomato genotypes. Int. J. Exp. Agric. 2011, 2, 8–11. [Google Scholar]
- Yokoyama, G.; Yasutake, D.; Tanizaki, T.; Kitano, M. Leaf wetting mitigates midday depression of photosynthesis in tomato plants. Photosynthetica 2019, 57, 740–747. [Google Scholar] [CrossRef]
- Shibuya, T.; Endo, R.; Yuba, T.; Kitaya, Y. The photosynthetic parameters of cucumber as affected by irradiances with different red:far-red ratios. Biol Plant 2015, 59, 198–200. [Google Scholar] [CrossRef]
- Shock, C.C.; Wang, F.X.; Flock, R.; Feibert, E.; Shock, C.A.; Pereira, A. Irrigation Monitoring Using Soil Water Tension; Oregon State University: Corvallis, OR, USA, 2013; pp. 1–9. [Google Scholar]
- Zheng, J.; Huang, G.; Jia, D.; Wang, J.; Mota, M.; Pereira, L.S.; Huang, Q.; Xu, X.; Liu, H. Responses of drip irrigated tomato (Solanum lycopersicum L.) yield, quality and water productivity to various soil matric potential thresholds in an arid region of Northwest China. Agric. Water Manag. 2013, 129, 181–193. [Google Scholar] [CrossRef]
- Wu, Y.S.; Gong, W.Z.; Wang, Y.M.; Yang, W.Y. Shading of mature leaves systemically regulates photosynthesis and leaf area of new developing leaves via hormones. Photosynthetica 2019, 57, 303–310. [Google Scholar] [CrossRef]
- Li, B.; Shi, B.; Yao, Z.; Kumar Shukla, M.; Du, T. Energy partitioning and microclimate of solar greenhouse under drip and furrow irrigation systems. Agric. Water Manag. 2020, 234, 106096. [Google Scholar] [CrossRef]
- Liu, F.; Cohen, Y.; Fuchs, M.; Plaut, Z.; Grava, A. The effect of vapor pressure deficit on leaf area and water transport in flower stems of soil-less culture rose. Agric. Water Manag. 2006, 81, 216–224. [Google Scholar] [CrossRef]
- Gong, X.; Qiu, R.; Sun, J.; Ge, J.; Li, Y.; Wang, S. Evapotranspiration and crop coefficient of tomato grown in a solar greenhouse under full and deficit irrigation. Agric. Water Manag. 2020, 235, 106154. [Google Scholar] [CrossRef]
- Liu, H.; Cohen, S.; Hugo, L.J.; Yair, I.; Josef, T. Sap flow, canopy conductance and microclimate in a banana screenhouse. Agric. For. Meteorol. 2015, 201, 165–175. [Google Scholar] [CrossRef]
- Mao, H.; Ikram, U.; Ni, J.; Qaiser, J.; Ahmad, A. Estimating tomato water consumption by sap flow measurement in response to water stress under greenhouse conditions. J. Plant Interact. 2017, 12, 402–413. [Google Scholar] [CrossRef]
- Li, L.; Chen, S.; Yang, C.; Meng, F.; Sigrimis, N. Prediction of plant transpiration from environmental parameters and relative leaf area index using the random forest regression algorithm. J. Clean. Prod. 2020, 261, 121136. [Google Scholar] [CrossRef]
- An, S.; Yang, F.; Yang, Y.; Huang, Y.; Zhangzhong, L.; Wei, X.; Yu, J. Water Demand Pattern and Irrigation Decision-Making Support Model for Drip-Irrigated Tomato Crop in a Solar Greenhouse. Agronomy 2022, 12, 1668. [Google Scholar] [CrossRef]
- Jarvis, P.G.; McNaughton, K.G. Stomatal Control of Transpiration: Scaling up from Leaf to Region. Adv. Ecol. Res. 1986, 15, 1–49. [Google Scholar]
- Farquhar, G.D.; Sharkey, T.D. Stomatal Conductance and Photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Motzo, R.; Pruneddu, G.; Giunta, F. The role of stomatal conductance for water and radiation use efficiency of durum wheat and triticale in a Mediterranean environment. Eur. J. Agron. 2013, 44, 87–97. [Google Scholar] [CrossRef]
- Siddiq, Z.; Chen, Y.-J.; Zhang, Y.-J.; Zhang, J.-L.; Cao, K.-F. More sensitive response of crown conductance to VPD and larger water consumption in tropical evergreen than in deciduous broadleaf timber trees. Agric. For. Meteorol. 2017, 247, 399–407. [Google Scholar] [CrossRef]
- Monje, O.; Bugbee, B. Radiometric Method for Determining Canopy Stomatal Conductance in Controlled Environments. Agronomy 2019, 9, 114. [Google Scholar] [CrossRef]
- Li, G.; Lin, L.; Dong, Y.; An, D.; Li, Y.; Luo, W.; Yin, X.; Li, W.; Shao, J.; Zhou, Y.; et al. Testing two models for the estimation of leaf stomatal conductance in four greenhouse crops cucumber, chrysanthemum, tulip and lilium. Agric. For. Meteorol. 2012, 165, 92–103. [Google Scholar] [CrossRef]
- Goto, K.; Yabuta, S.; Ssenyonga, P.; Tamaru, S.; Sakagami, J.-I. Response of leaf water potential, stomatal conductance and chlorophyll content under different levels of soil water, air vapor pressure deficit and solar radiation in chili pepper (Capsicum chinense). Sci. Hortic.-Amst. 2021, 281, 109943. [Google Scholar] [CrossRef]
- Flach, R.; Skalský, R.; Folberth, C.; Balkovič, J.; Jantke, K.; Schneider, U.A. Water productivity and footprint of major Brazilian rainfed crops—A spatially explicit analysis of crop management scenarios. Agric. Water Manag. 2020, 233, 105996. [Google Scholar] [CrossRef]
- Mekonnen, M.M.; Hoekstra, A.Y.; Neale, C.M.U.; Ray, C.; Yang, H.S. Water productivity benchmarks: The case of maize and soybean in Nebraska. Agric. Water Manag. 2020, 234, 106122. [Google Scholar] [CrossRef]
- Yaghi, T.; Arslan, A.; Naoum, F. Cucumber (Cucumis sativus, L.) water use efficiency (WUE) under plastic mulch and drip irrigation. Agric. Water Manag. 2013, 128, 149–157. [Google Scholar] [CrossRef]
- Liu, H.; Yuan, B.; Hu, X.; Yin, C. Drip irrigation enhances water use efficiency without losses in cucumber yield and economic benefits in greenhouses in North China. Irrig. Sci. 2022, 40, 135–149. [Google Scholar] [CrossRef]
- Jifon, J.L.; Syvertsen, J.P. Moderate shade can increase net gas exchange and reduce photoinhibition in citrus leaves. Tree Physiol. 2003, 23, 119–127. [Google Scholar] [CrossRef]
- Jutamanee, K.; Onnom, S. Improving photosynthetic performance and some fruit quality traits in mango trees by shading. Photosynthetica 2016, 54, 542–550. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Stommel, J.R. Pollen Viability and Fruit Set of Tomato Genotypes under Optimumand High-temperature Regimes. HortScience 1995, 30, 115–117. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Planting Seasons | Date | LAI | Plant Height (m) | Seasonal Yield (ton ha−1) |
|---|---|---|---|---|
| 2019SU | 27 June 2019 | 1.41 ± 0.06 | 1.02 ± 0.06 | 10.8 |
| 15 July 2019 | 2.07 ± 0.08 | 1.42 ± 0.10 | ||
| 26 July 2019 | 2.19 ± 0.13 | 1.50 ± 0.09 | ||
| 06 August 2019 | 2.27 ± 0.19 | 1.60 ± 0.09 | ||
| 16 August 2019 | 2.34 ± 0.23 | 1.70 ± 0.12 | ||
| 2019AW | 04 October 2019 | 0.92 ± 0.01 | 0.81 ± 0.02 | 56.3 |
| 26 October 2019 | 1.62 ± 0.21 | 1.19 ± 0.05 | ||
| 23 November 2019 | 1.55 ± 0.44 | 1.25 ± 0.06 | ||
| 21 December 2019 | 1.68 ± 0.38 | 1.23 ± 0.08 | ||
| 2020AW | 05 October 2020 | 2.19 ± 0.02 | 1.12 ± 0.02 | 83.6 |
| 10 November 2020 | 3.31 ± 0.46 | 1.54 ± 0.03 | ||
| 26 December 2020 | 2.19 ± 0.63 | 1.52 ± 0.04 |
| Planting Seasons | Month | Rs (MJ m−2 d−1) | Ta (°C) | RH (%) | VPD (kPa) | U (m s−1) |
|---|---|---|---|---|---|---|
| 2019SU | Inside | 10.93 ± 3.60 | 28.63 ± 2.50 | 77.07 ± 11.25 | 1.07 ± 0.47 | 0.39 ± 0.07 |
| Outside | 19.47 ± 6.23 | 27.36 ± 2.18 | 69.36 ± 16.48 | 0.97 ± 0.63 | 1.49 ± 0.98 | |
| 2019AW | Inside | 6.94 ± 3.60 | 17.84 ± 4.02 | 82.57 ± 9.02 | 0.57 ± 0.42 | 0.26 ± 0.06 |
| Outside | 9.95 ± 5.10 | 9.04 ± 8.63 | 73.16 ± 14.39 | 0.38 ± 0.28 | 0.93 ± 0.78 | |
| 2020AW | Inside | 6.49 ± 3.01 | 17.17 ± 2.19 | 82.70 ± 7.38 | 0.51 ± 0.23 | 0.15 ± 0.04 |
| Outside | 9.21 ± 3.73 | 6.50 ± 6.84 | 68.14 ± 14.74 | 0.38 ± 0.26 | 1.32 ± 0.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Shao, M.; Yang, L. Photosynthesis Characteristics of Tomato Plants and Its’ Responses to Microclimate in New Solar Greenhouse in North China. Horticulturae 2023, 9, 197. https://doi.org/10.3390/horticulturae9020197
Liu H, Shao M, Yang L. Photosynthesis Characteristics of Tomato Plants and Its’ Responses to Microclimate in New Solar Greenhouse in North China. Horticulturae. 2023; 9(2):197. https://doi.org/10.3390/horticulturae9020197
Chicago/Turabian StyleLiu, Haijun, Mengxuan Shao, and Li Yang. 2023. "Photosynthesis Characteristics of Tomato Plants and Its’ Responses to Microclimate in New Solar Greenhouse in North China" Horticulturae 9, no. 2: 197. https://doi.org/10.3390/horticulturae9020197
APA StyleLiu, H., Shao, M., & Yang, L. (2023). Photosynthesis Characteristics of Tomato Plants and Its’ Responses to Microclimate in New Solar Greenhouse in North China. Horticulturae, 9(2), 197. https://doi.org/10.3390/horticulturae9020197