Protective Effect of Prunus mume Fermented with Mixed Lactic Acid Bacteria in Dextran Sodium Sulfate-Induced Colitis

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Materials
2.2. Production of FP
2.3. Total Soluble Solids, pH and Cell Counting
2.4. Organic Acid and Free Amino Acid Contents
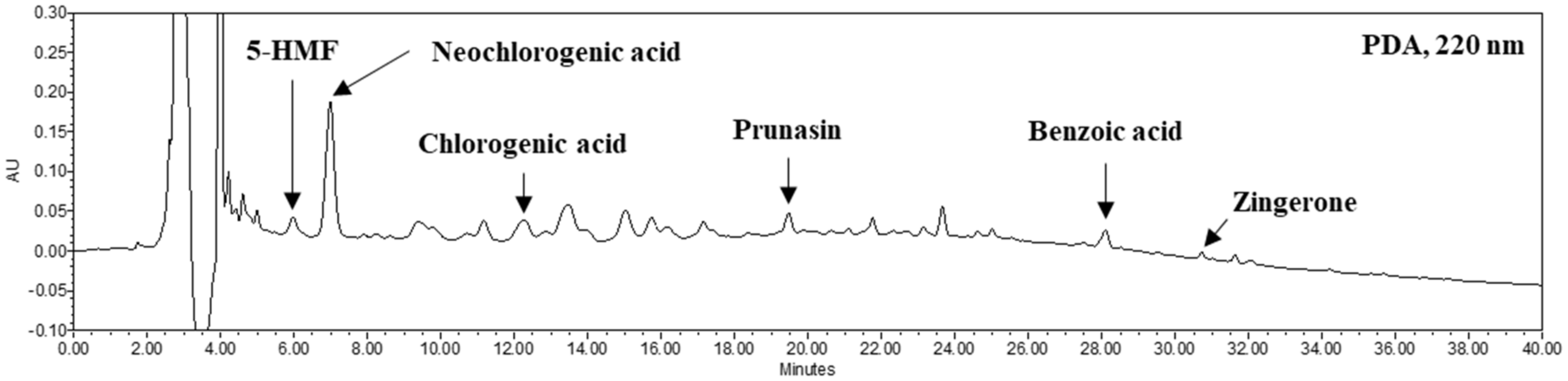
2.5. Identification of Phenolic Compounds in FP
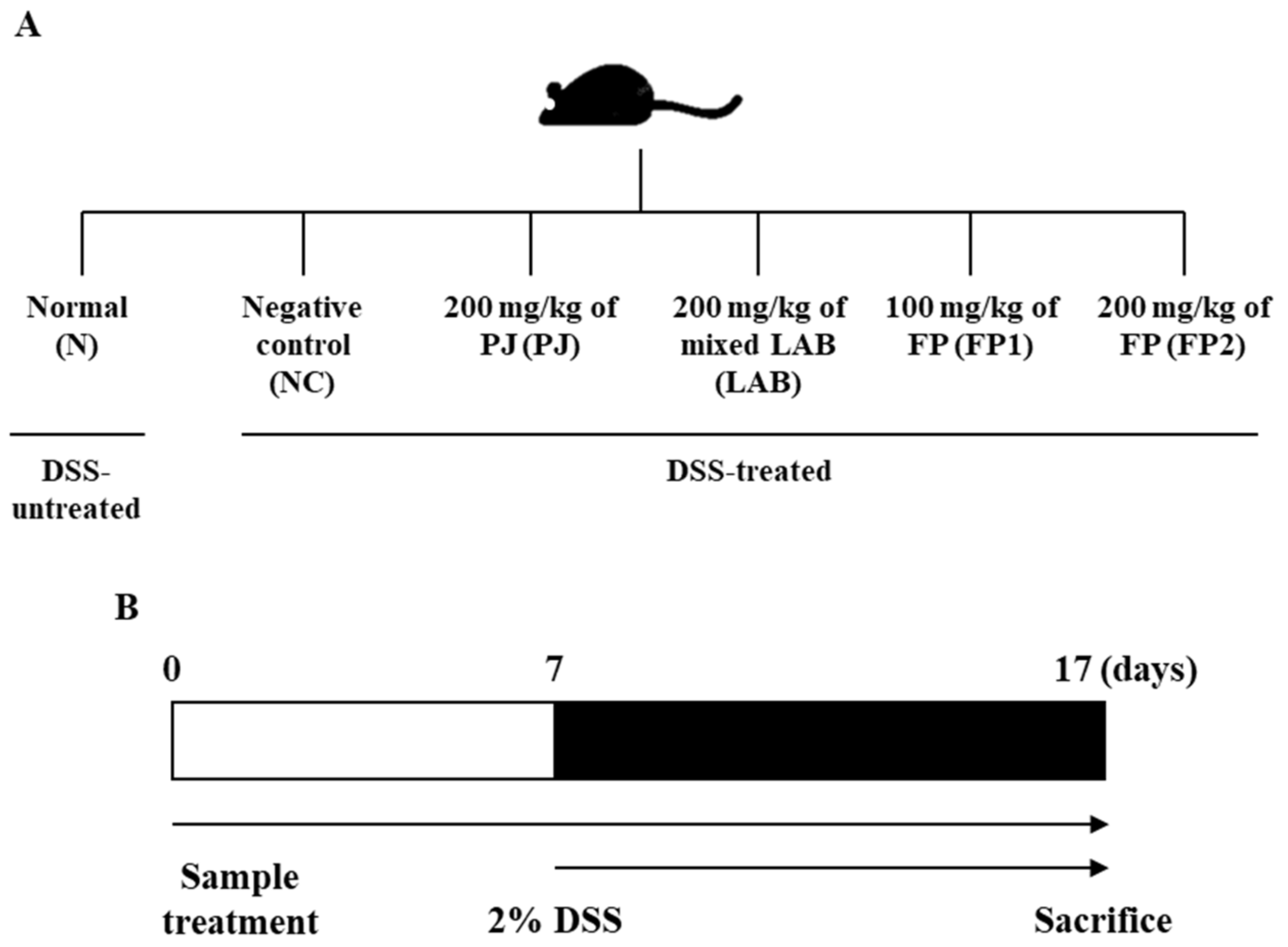
2.6. Experimental Design
2.7. Lipid Peroxidation
2.8. Hematoxylin and Eosin (H&E) Staining Assay
2.9. Evaluation of Inflammatory Cytokines
2.10. Determination of Apoptosis in Colon Tissue
2.11. Statistical Analysis
3. Results and Discussion
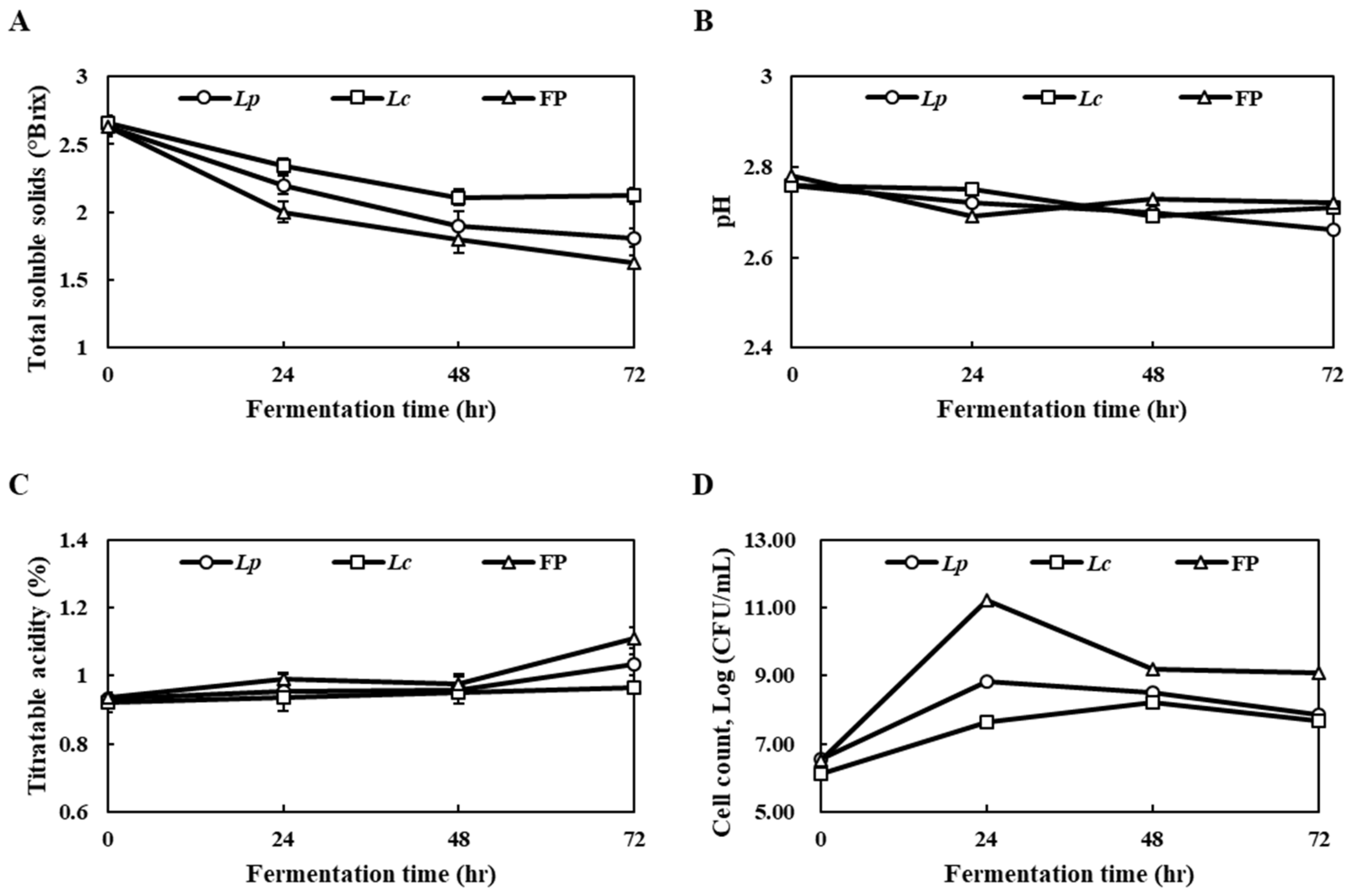
3.1. Changes in the Total Soluble Solids, pH and Cell Count
3.2. Contents of Organic Acids, Free Amino Acids and Phenolic Compounds of FP
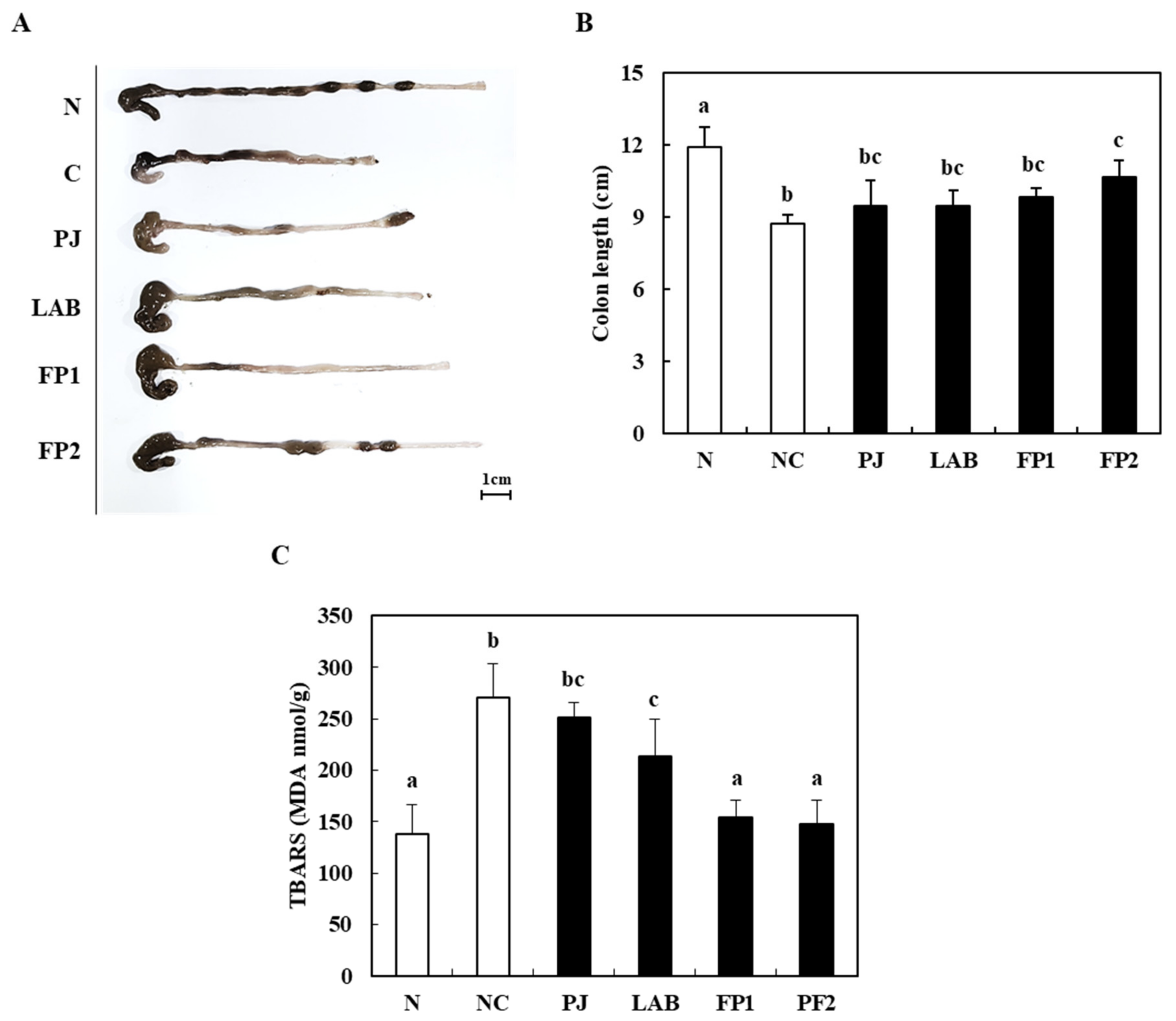
3.3. Effect of FP on the Colon Length and Lipid Peroxidation
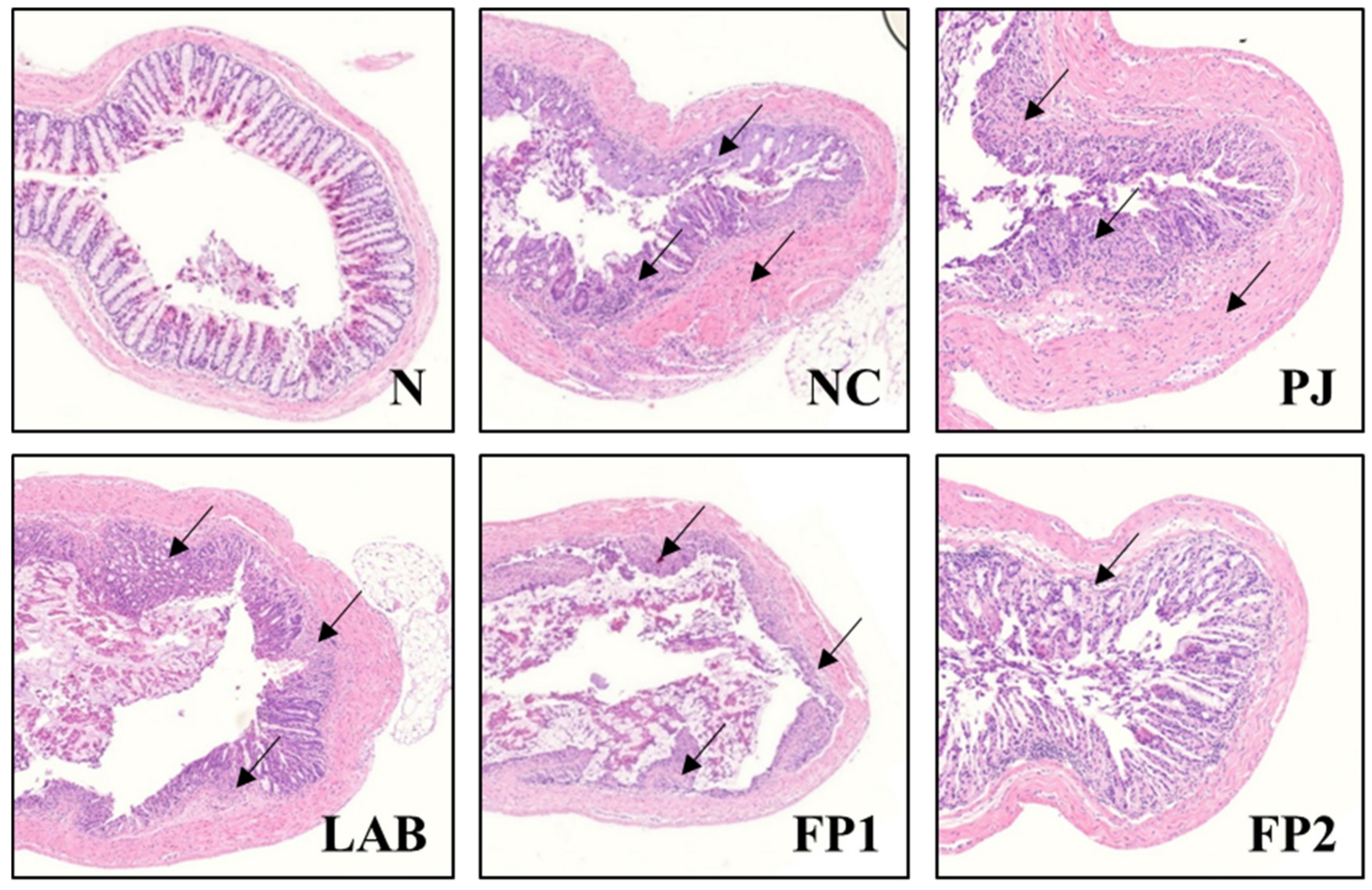
3.4. Effects of FP on Colonic Tissue Damage
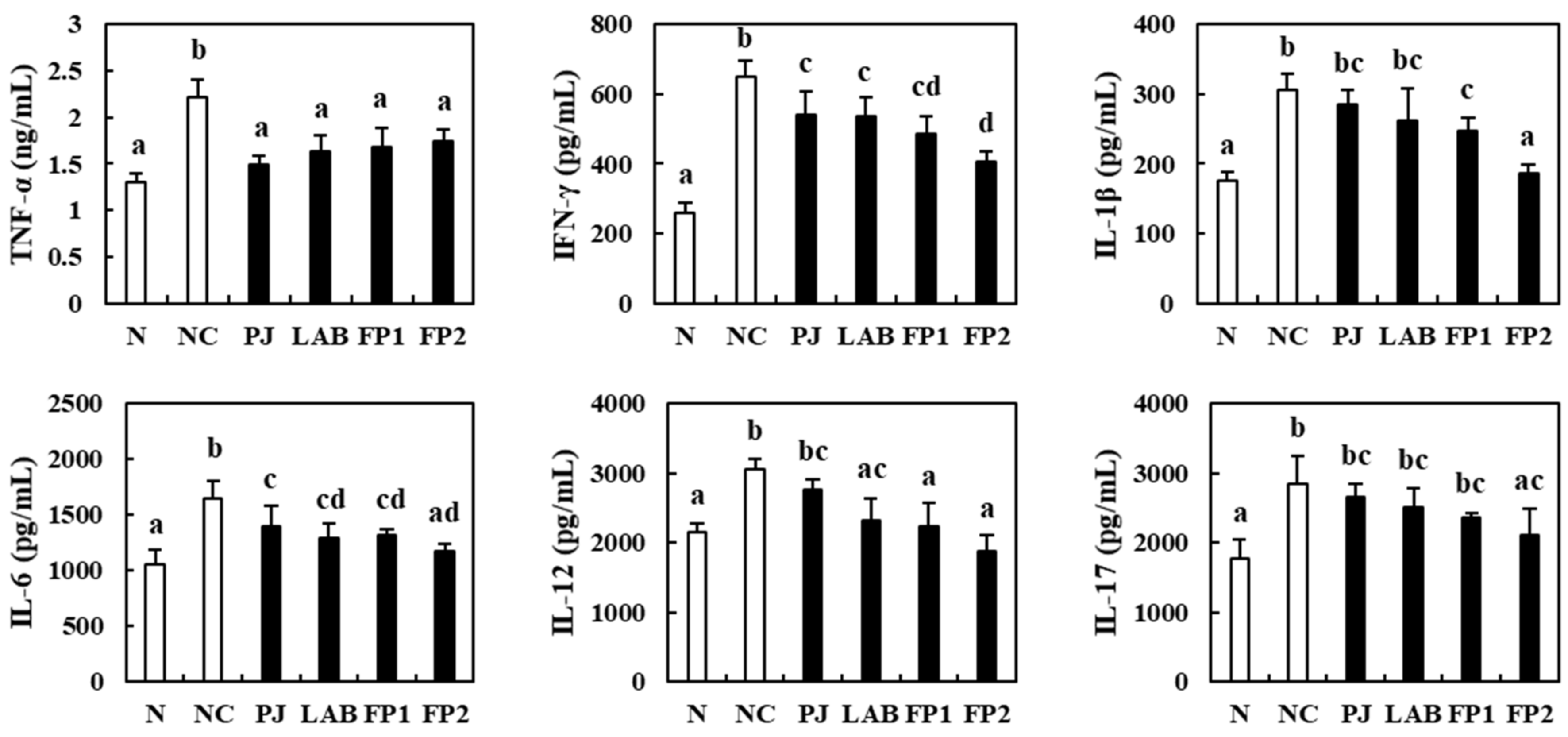
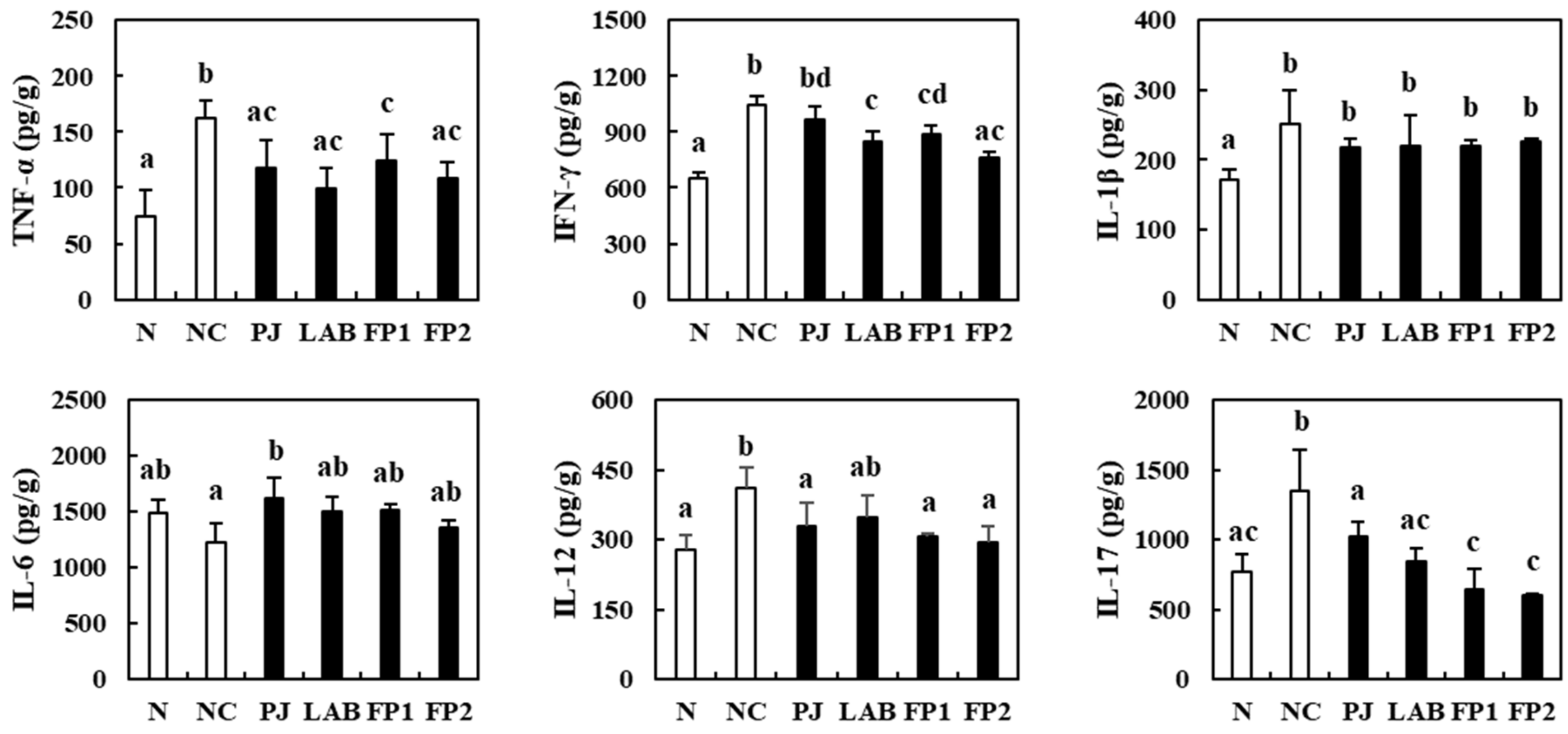
3.5. Effects of FP on the Serum Inflammatory Cytokines
3.6. Effects of FP on Colonic Inflammatory Cytokines
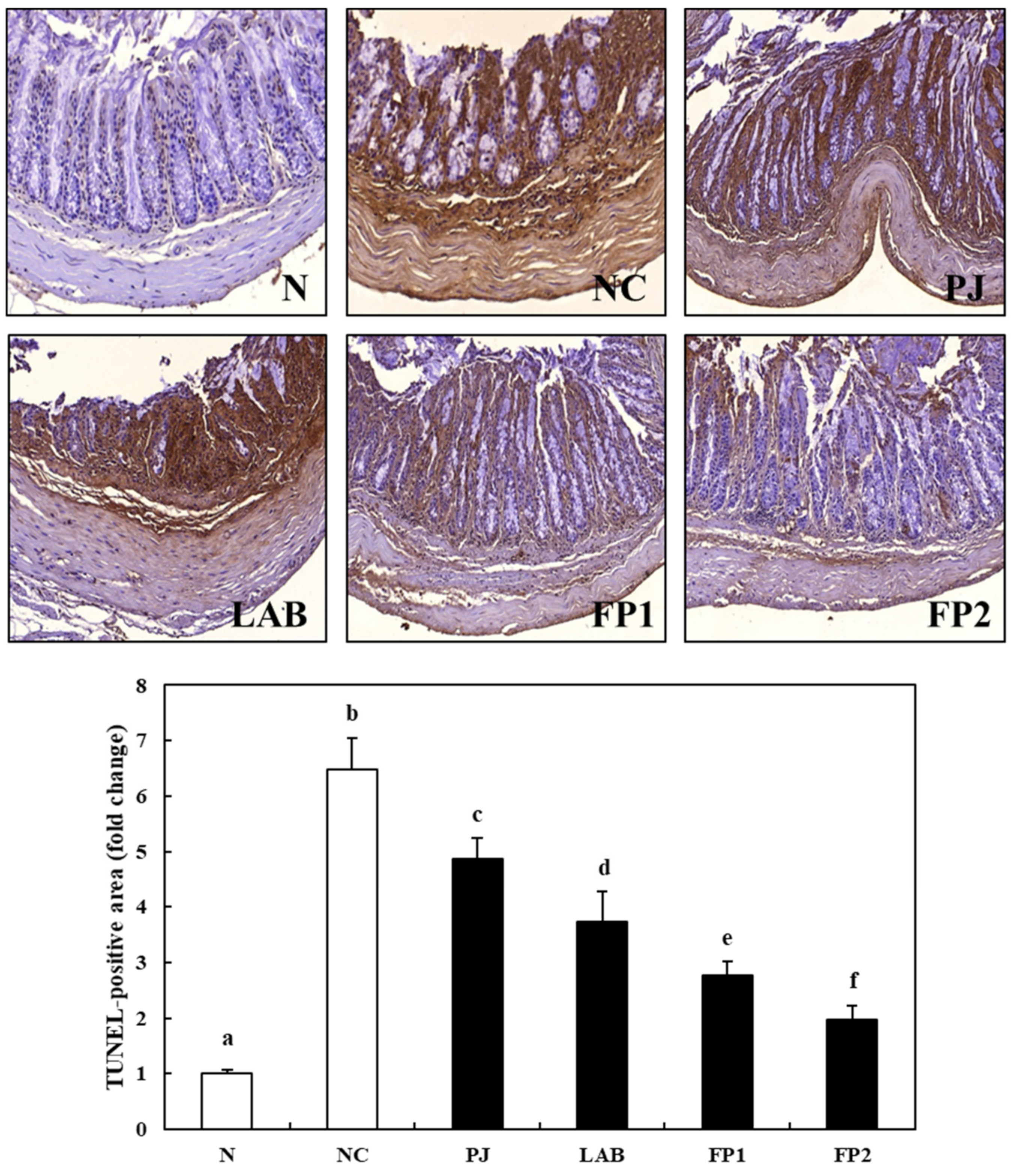
3.7. Effects of FP on Apoptosis in Colonic Tissue
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Kappleman, M.D.; Moore, K.R.; Allen, J.K.; Cook, S.F. Recent trends in the prevalence of Crohn’s disease and ulcerative colitis in a commercially insured US population. Dig. Dis. Sci. 2013, 58, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Sufit, A.J.D.; Wischmeyer, P.E. Glutamine therapy improves outcome of in vitro and in vivo experimental colitis model. J. Parenter. Enteral. Nutr. 2011, 35, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Vukelić, I.; Detel, D.; Pućar, L.B.; Potoćnjak, I.; Buljević, S.; Domitrović, R. Chlorogenic acid ameliorates experimental colitis in mice by suppressing signaling pathways involved in inflammatory response and apoptosis. Food Chem. Toxicol. 2018, 121, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Abraham, B.P.; Ahmed, T.; Ali, T. Inflammatory bowel disease: Pathophysiology and current therapeutic approaches. In Gastrointestinal Pharmacology; Springer: Boston, MA, USA, 2017; pp. 115–146. [Google Scholar]
- Na, J.R.; Oh, K.N.; Park, S.U.; Bae, D.H.; Choi, E.J.; Jung, M.A.; Choi, C.Y.; Lee, D.W.; Jun, W.J.; Lee, K.Y.; et al. The laxative effects of maesil (Prunus mume Siebold & Zucc.) on constipation induced by a low-fibre diet in a rat model. Int. J. Food Sci. Nutr. 2013, 64, 333–345. [Google Scholar]
- Hwang, J.Y.; Ham, J.W.; Nam, S.H. The antioxidant activity of maesil (Prunus mume). Korean J. Food Sci. Technol. 2004, 36, 461–464. [Google Scholar]
- Chuda, Y.; Ono, H.; Ohnishi-Kameyama, M.; Matsumoto, K.; Nagata, T.; Kikuchi, Y. Mumefural, citric acid derivate improving blood fluidity from fruit-juice concentrate of Japanese apricot (Prunus mume Sieb. et Zucc). J. Agric. Food Chem. 1999, 47, 828–831. [Google Scholar] [CrossRef]
- Debnath, T.; Kim, D.H.; Lim, B.O. Natural products as a source of anti-inflammatory agents associated with inflammatory bowel disease. Molecules 2013, 18, 7253–7270. [Google Scholar] [CrossRef]
- Kim, M.S.; Bang, J.H.; Lee, J.; Han, J.S.; Kang, H.W.; Jeon, W.K. Fructus mume ethanol extract prevents inflammation and normalized the septohippocampal cholinergic system in a rat model of chronic cerebral hypoperfusion. J. Med. Food 2016, 19, 196–204. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, S.J.; Hur, S.J. Effects of Prunus mume Sieb. et Zucc. Extract and its biopolymer encapsulation on a mouse model of colitis. J. Sci. Food Agric. 2017, 97, 686–692. [Google Scholar] [CrossRef]
- Nuraida, L. A review: Health promoting lactic acid bacteria in traditional Indonesian fermented foods. Food Sci. Hum. Wellness 2015, 4, 47–55. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, M.; Mujumdar, A.S.; Gao, Z. Recent research process of fermented plant extract: A review. Trends Food Sci. Technol. 2017, 65, 40–48. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Paparella, A.; Martuscelli, M.; Corsetti, A.; Tofalo, R.; Suzzi, G. Impact of microbial cultures on proteolysis and release of bioactive peptides in fermented milk. Food Microbiol. 2014, 42, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, R.; Bosco, F.; Mizrahi, I.; Bayer, E.A.; Pessione, E. Towards lactic acid bacteria-based biorefineries. Biotechnol. Adv. 2014, 32, 1216–1236. [Google Scholar] [CrossRef] [PubMed]
- Lizardo, R.C.M.; Cho, H.D.; Won, Y.S.; Seo, K.I. Fermentation with mono- and mixed cultures of Lactobacillus pllantarum and L. casei enhances the phytochemical content and biological activities of cherry silverberry (Elaeagnus multiflora Thunb.) fruit. J. Sci. Food Agric. 2020, 100, 3687–3696. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Liu, L.; Lai, T.; Zhang, R.; Wei, Z.; Xiao, J.; Deng, Y.; Zhang, M. Phenolic profile, free amino acids composition and antioxidant potential of dried longan fermented by lactic acid bacteria. J. Food Sci. Technol. 2018, 55, 4782–4791. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T.; Xu, H.; Ye, J.Z.; Wu, W.R.; Shi, D.; Fang, D.Q.; Liu, Y.; Li, L.J. Efficacy of Lactobacillus rhamnosus GG in treatment of acute pediatric diarrhea: A systematic review with meta-analysis. World J. Gastroenterol. 2019, 25, 4999–5016. [Google Scholar] [CrossRef]
- Yi, R.; Peng, P.; Zhang, J.; Du, M.; Lan, L.; Qian, Y.; Zhou, J.; Zhao, X. Lactobacillus plantarum CQPC02-fermented soybean milk improves loperamide-induced constipation in mice. J. Med. Food 2019, 22, 1208–1221. [Google Scholar] [CrossRef]
- The role of probiotic lactic acid bacteria and bifidobacteria in the prevention and treatment of inflammatory bowel disease and other related diseases: A systematic review of randomized human clinical trials. Biomed. Res. Int. 2015, 2015, 505878.
- Canon, F.; Nidelet, T.; Guédon, E.; Thierry, A.; Gagnaire, V. Understanding the mechanisms of positive microbial interactions that benefit lactic acid bacteria co-cultures. Front. Microbiol. 2020, 11, 2088. [Google Scholar] [CrossRef]
- Cho, H.D.; Kim, J.H.; Won, Y.S.; Moon, K.D.; Seo, K.I. Inhibitory effects of pectinase-treated Prunus mume fruit concentrate on colorectal cancer proliferation and angiogenesis of endothelial cells. J. Food Sci. 2019, 84, 3284–3295. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, H.D.; Won, Y.S.; Hong, S.M.; Moon, K.D.; Seo, K.I. Anti-fatigue effect of Prunus mume vinegar in high-intensity exercised rats. Nutrients 2020, 12, 1205. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.M.; Fillmann, H.S.; Martins, M.I.M.; Meurer, L.; Marroni, N.P. Boswellia serrata has beneficial anti-inflammatory and antioxidant properties in a model of experimental colitis. Phytother. Res. 2014, 28, 1392–1398. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.H.; Jacobson, K.A.; Rose, J.; Zeller, R. Hematoxylin and eosin staining of tissue and cell sections. CSH Protoc. 2008, 2008. [Google Scholar] [CrossRef]
- Elkhtab, E.; El-Alfy, M.; Shenana, M.; Mohamed, A.; Yousef, A.E. New potentially antihypertensive peptides liberated in milk during fermentation with selected lactic acid bacteria and kombucha cultures. J. Dairy Sci. 2017, 100, 9508–9520. [Google Scholar] [CrossRef]
- Cha, H.S.; Hwang, J.B.; Park, J.S.; Park, Y.K.; Jo, J.S. Changes in chemical composition of mume (Prunus mume Sieb. et Zucc) fruits during maturation. Korean J. Food Preserv. 1999, 12, 481–487. [Google Scholar]
- Mitani, T.; Horinishi, A.; Kishida, K.; Kawabata, T.; Yano, F.; Mimura, H.; Inaba, N.; Yamanishi, H.; Oe, T.; Negoro, K.; et al. Phenolics profile of mume, Japanese apricot (Prunus mume Sieb. et Zucc.) fruit. Biosci. Biotechnol. Biochem. 2013, 77, 1623–1627. [Google Scholar] [CrossRef]
- Chidi, B.S.; Bauer, F.F.; Rossouw, D. Organic acid metabolism and the impact of fermentation practices on wine acidity—A review. S. Afr. J. Enol. Cotoc. 2018, 39, 315–329. [Google Scholar] [CrossRef]
- Erbas, M.; Ertugay, M.F.; Erbas, M.O.; Certel, M. The effect of fermentation and storage on free amino acids of tarhana. Int. J. Food Sci. Nutr. 2005, 56, 349–359. [Google Scholar] [CrossRef]
- We, C.; Huang, J.; Zhou, R. Genomics of lactic acid bacteria: Current status and potential applications. Crit. Rev. Microbiol. 2017, 43, 393–404. [Google Scholar]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Shah, N.P. Lactic acid bacterial fermentation modified phenolic composition in tea extracts and enhanced their antioxidant activity and cellular uptake of phenolic compounds following in vitro digestion. J. Funct. Foods 2016, 20, 182–194. [Google Scholar] [CrossRef]
- Sangaraju, R.; Nalban, N.; Alavala, S.; Rajendran, V.; Jerald, M.K.; Sistla, R. Protective effect of galangin against dextran sulfate sodium (DSS)-induced ulcerative colitis in Balb/c mice. Inflamm. Res. 2019, 68, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Murano, M.; Maemura, K.; Hirata, K.; Toshina, K.; Nishikawa, T.; Hanamoto, N.; Sasaki, S.; Saitoh, O.; Katsu, K. Therapeutic effect of intracolonically administered nuclear factor κB (p65) antisense oligonucleotide on mouse dextran sulphate sodium (DSS)-induced colitis. Clin. Exp. Immunol. 2000, 120, 51–58. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2015, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef]
- Islam, J.; Koseki, T.; Watanabe, K.; Ardiansyah, A.; Budijanto, S.; Oikawa, A.; Alauddin, M.; Goto, T.; Aso, H.; Komai, M.; et al. Dietary supplementation of fermented rice bran effectively alleviates dextran sodium sulfate-induced colitis in mice. Nutrients 2017, 9, 747. [Google Scholar] [CrossRef]
- Dinarello, C.A. Proinflammatory cytokines. Chest 2000, 118, 503–508. [Google Scholar] [CrossRef]
- Mario, R.C.; Elizdath, M.G.; Alma, D.N.T.; Luvia, E.S.T.; Leticia, G.S.; Maria, M.C.M.; Luis, I.T.; Marco, A.R.M. Anti-inflammatory and antioxidant activities of the methanolic extract of Cyrtocarpa procera bark reduces the severity of ulcerative colitis in a chemically induced colitis model. Mediat. Inflamm. 2020, 2020, 5062506. [Google Scholar]
- Zhou, X.; Liu, H.; Zhang, J.; Mu, J.; Zalan, Z.; Hegyi, F.; Takacs, K.; Zhao, X.; Du, M. Protective effect of Lactobacillus fermentum CQPC04 on dextran sulfate sodium-induced colitis in mice is associated with modulation of the nuclear factor-κB signaling pathway. J. Dairy Sci. 2019, 102, 9570–9585. [Google Scholar] [CrossRef]
- Park, J.S.; Joe, I.S.; Rhee, P.D.; Jeong, C.S.; Jeong, G.J. A lactic acid bacterium isolated from kimchi ameliorates intestinal inflammation in DSS-induced colitis. J. Microbiol. 2017, 55, 304–310. [Google Scholar] [CrossRef]
- Cui, Y.; Wei, H.; Lu, F.; Liu, X.; Liu, D.; Gu, L.; Ouyang, C. Different effects of three selected Lactobacillus strains in dextran sulfate sodium-induced colitis in BALB/c mice. PLoS ONE 2016, 11, e0148241. [Google Scholar] [CrossRef] [PubMed]
- Nunzio, M.D.; Picone, G.; Pasini, F.; Chiarello, E.; Caboni, M.F.; Capozzi, F.; Gianotti, A.; Bordoni, A. Olive oil by-product as functional ingredient in bakery products. Influence of processing and evaluation of biological effects. Food Res. Int. 2020, 131, 108940. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Gu, P.Q.; Shen, H. Gallic acid improved inflammation via NF-κB pathway in TNBS-induced ulcerative colitis. Int. Immunopharmacol. 2019, 67, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Shen, W.; Wang, Y.; Cao, Y.; Nuerbulati, N.; Chen, W.; Lu, G.; Xiao, W.; Qi, R. Grape seed polyphenols ameliorated dextran sulfate sodium-induced colitis cia suppression of inflammation and apoptosis. Pharmacology 2020, 105, 9–18. [Google Scholar] [CrossRef]
- Shin, S.K.; Cho, J.H.; Kim, E.J.; Kim, E.K.; Park, D.K.; Kwon, K.A.; Chung, J.W.; Kim, K.O.; Kim, Y.J. Anti-inflammatory and anti-apoptotic effects of rosuvastatin by regulation of oxidative stress in a dextran sulfate sodium-induced colitis model. World J. Gastroenterol. 2017, 23, 4559–4568. [Google Scholar] [CrossRef]
- Chae, J.M.; Chang, M.H.; Heo, W.; Cho, H.T.; Lee, D.H.; Hwang, B.B.; Kim, J.W.; Yoon, S.M.; Yang, S.Y.; Lee, J.H.; et al. LB-9, novel probiotic lactic acid bacteria, ameliorates dextran sodium sulfate-induced colitis in mice by inhibiting TNF-α-mediated apoptosis of intestinal epithelial cells. J. Med. Food 2019, 22, 271–276. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Lp | Lc | FP |
|---|---|---|---|
| Lactobacillus plantarum | + | + | |
| Lactobacillus casei | + | + |
| Organic Acids | Contents (mg%) | Free Amino Acids | Contents (ppm) |
|---|---|---|---|
| Citric acid | 739.81 ± 17.61 | Aspartic acid | 33.32 ± 2.21 |
| Lactic acid | 184.27 ± 11.43 | Glutamic acid | 4.35 ± 0.74 |
| Acetic acid | 71.32 ± 4.52 | Alanine | 3.39 ± 0.35 |
| Malic acid | N.D. | Taurine | 2.29 ± 0.27 |
| Total organic acids | 996.07 ± 58.82 | Total free amino acid | 66.43 ± 4.25 |
| Body-Weight (g) | N | NC | PJ | LAB | FP1 | FP2 |
|---|---|---|---|---|---|---|
| Initial | 33.94 ± 0.81 | 34.11 ± 0.68 | 34.72 ± 1.04 | 34.71 ± 0.95 | 33.97 ± 1.65 | 34.61 ± 1.15 |
| Final | 36.26 ± 1.83 | 33.61 ± 1.82 | 34.69 ± 3.31 | 35.14 ± 2.32 | 34.59 ± 3.58 | 35.61 ± 2.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.-H.; Won, Y.-S.; Cho, H.-D.; Hong, S.-M.; Moon, K.-D.; Seo, K.-I. Protective Effect of Prunus mume Fermented with Mixed Lactic Acid Bacteria in Dextran Sodium Sulfate-Induced Colitis. Foods 2021, 10, 58. https://doi.org/10.3390/foods10010058
Kim J-H, Won Y-S, Cho H-D, Hong S-M, Moon K-D, Seo K-I. Protective Effect of Prunus mume Fermented with Mixed Lactic Acid Bacteria in Dextran Sodium Sulfate-Induced Colitis. Foods. 2021; 10(1):58. https://doi.org/10.3390/foods10010058
Chicago/Turabian StyleKim, Jeong-Ho, Yeong-Seon Won, Hyun-Dong Cho, Seong-Min Hong, Kwang-Deog Moon, and Kwon-Il Seo. 2021. "Protective Effect of Prunus mume Fermented with Mixed Lactic Acid Bacteria in Dextran Sodium Sulfate-Induced Colitis" Foods 10, no. 1: 58. https://doi.org/10.3390/foods10010058
APA StyleKim, J.-H., Won, Y.-S., Cho, H.-D., Hong, S.-M., Moon, K.-D., & Seo, K.-I. (2021). Protective Effect of Prunus mume Fermented with Mixed Lactic Acid Bacteria in Dextran Sodium Sulfate-Induced Colitis. Foods, 10(1), 58. https://doi.org/10.3390/foods10010058