Coastal Sand Dunes Monitoring by Low Vegetation Cover Classification and Digital Elevation Model Improvement Using Synchronized Hyperspectral and Full-Waveform LiDAR Remote Sensing

 ,
,  ,
, 
Abstract
:
1. Introduction
2. Study Areas, Field Measurements, and Data Acquisition
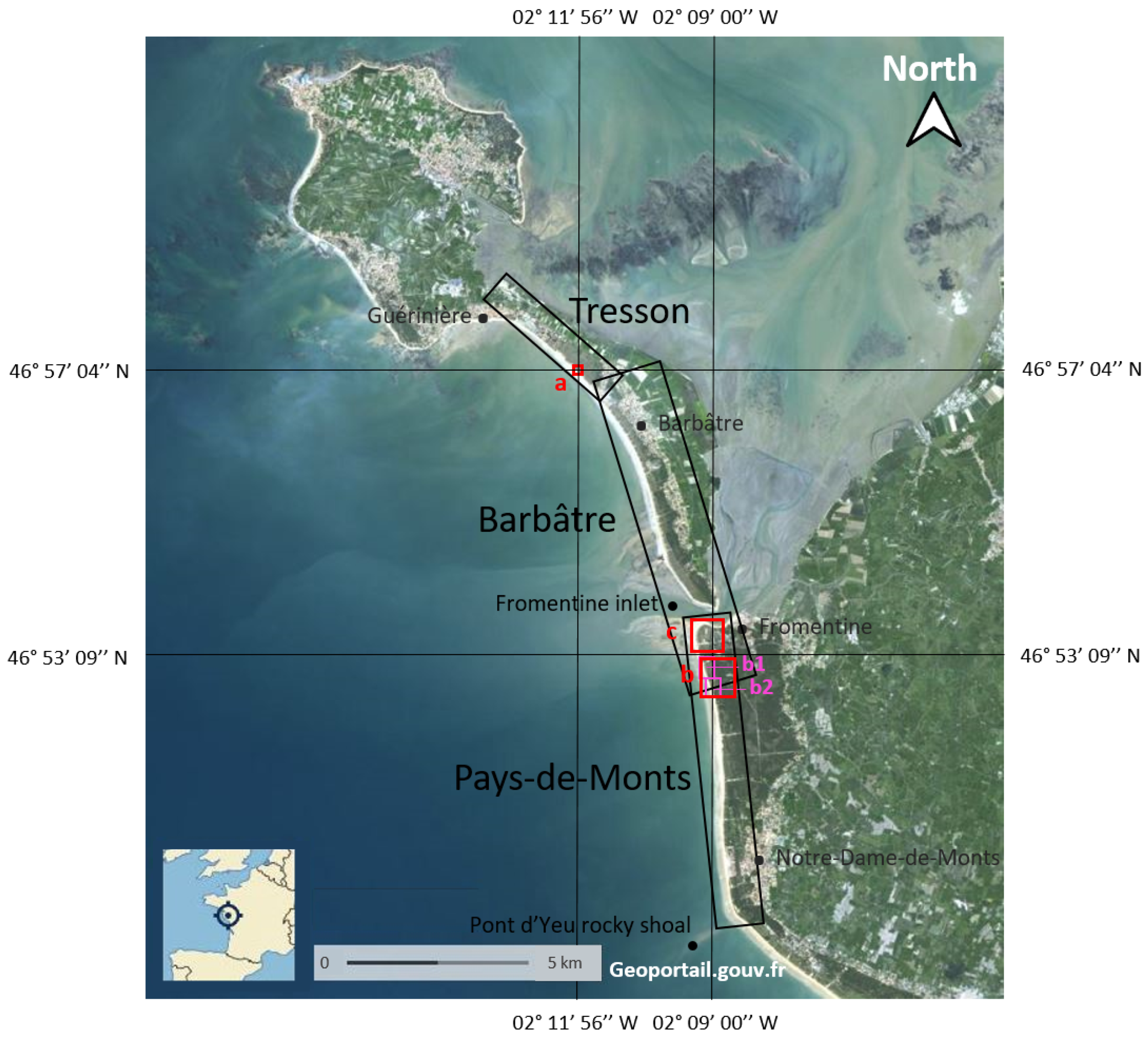
2.1. Study Areas
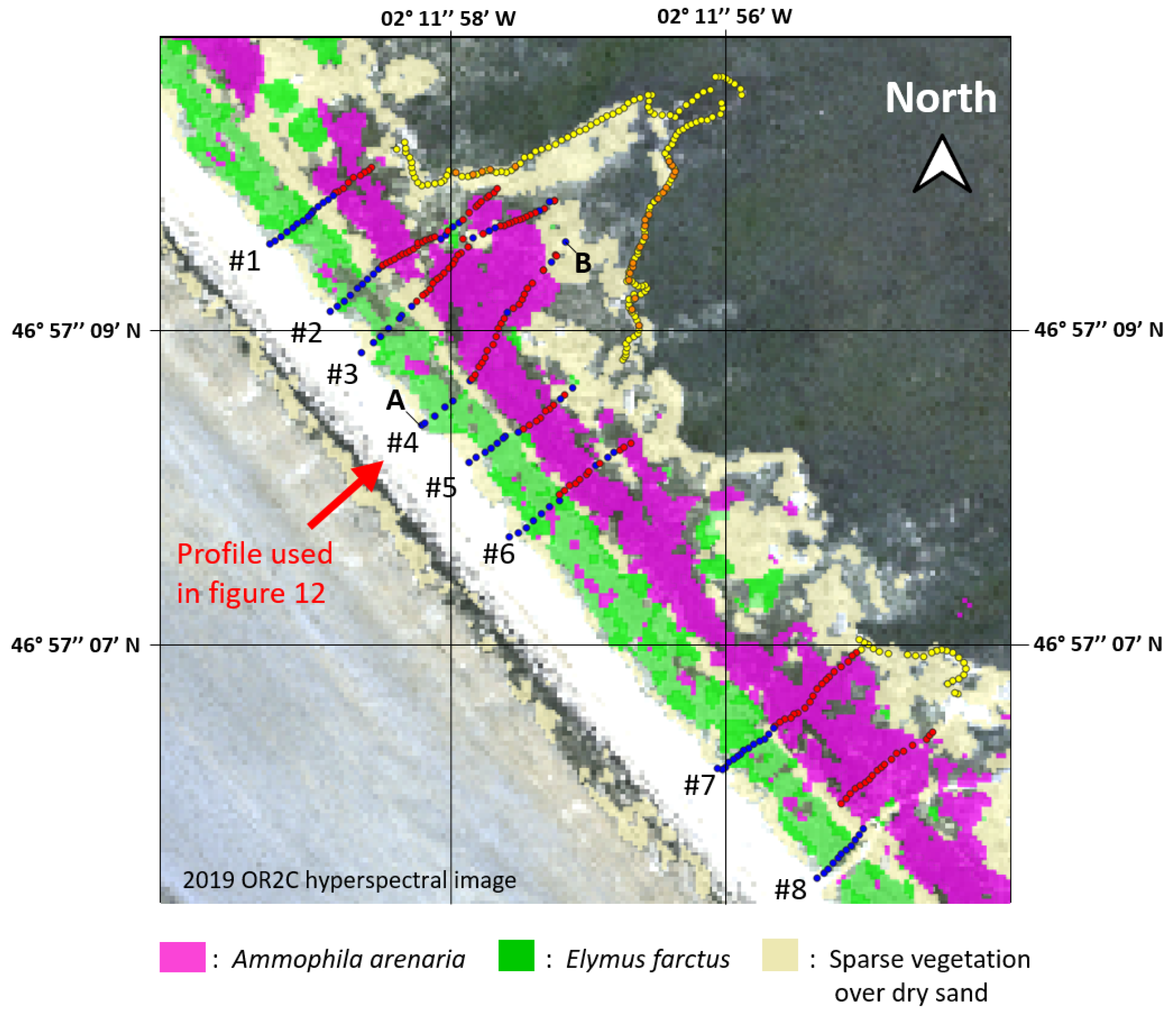
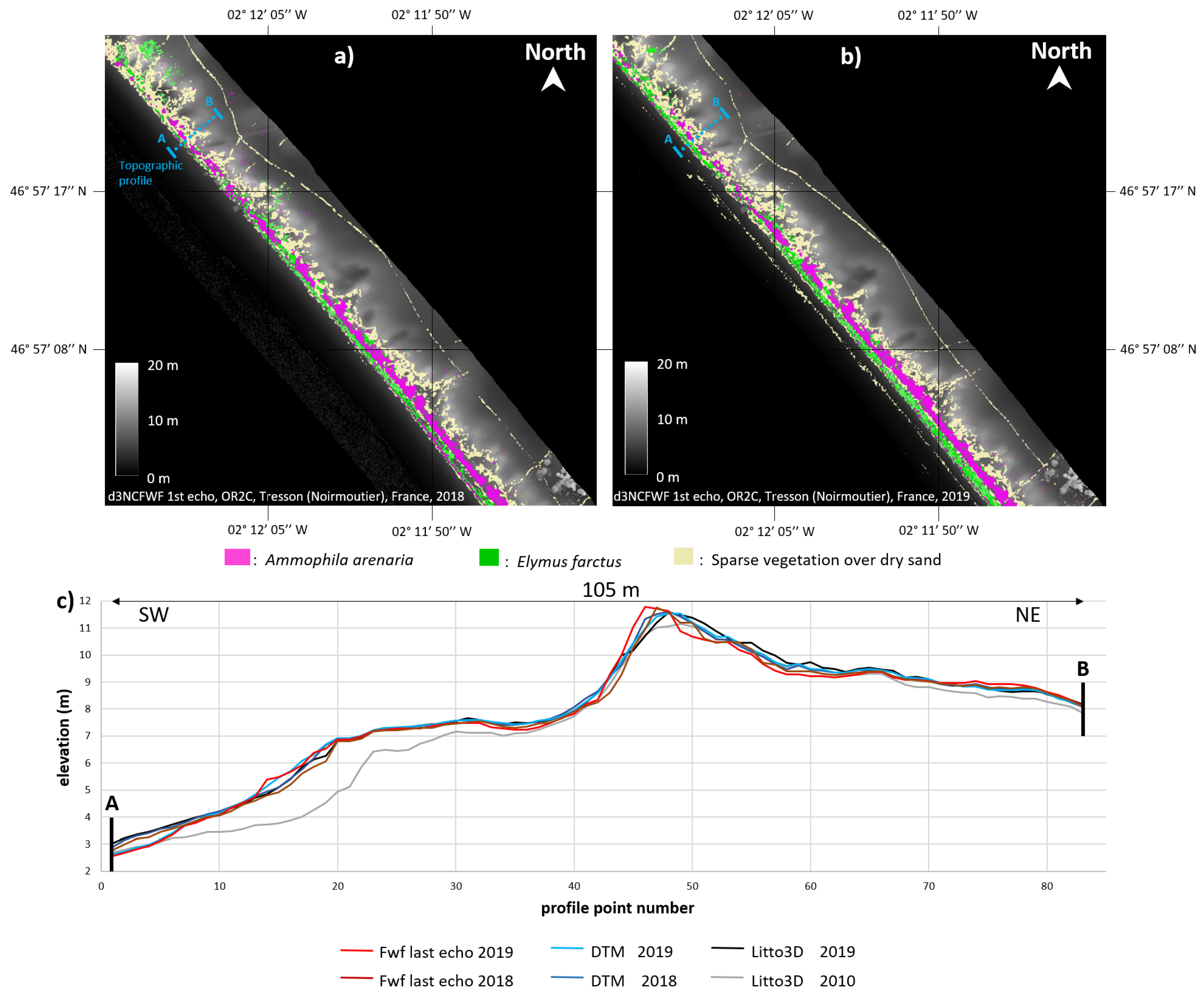
2.1.1. Training Area: Tresson
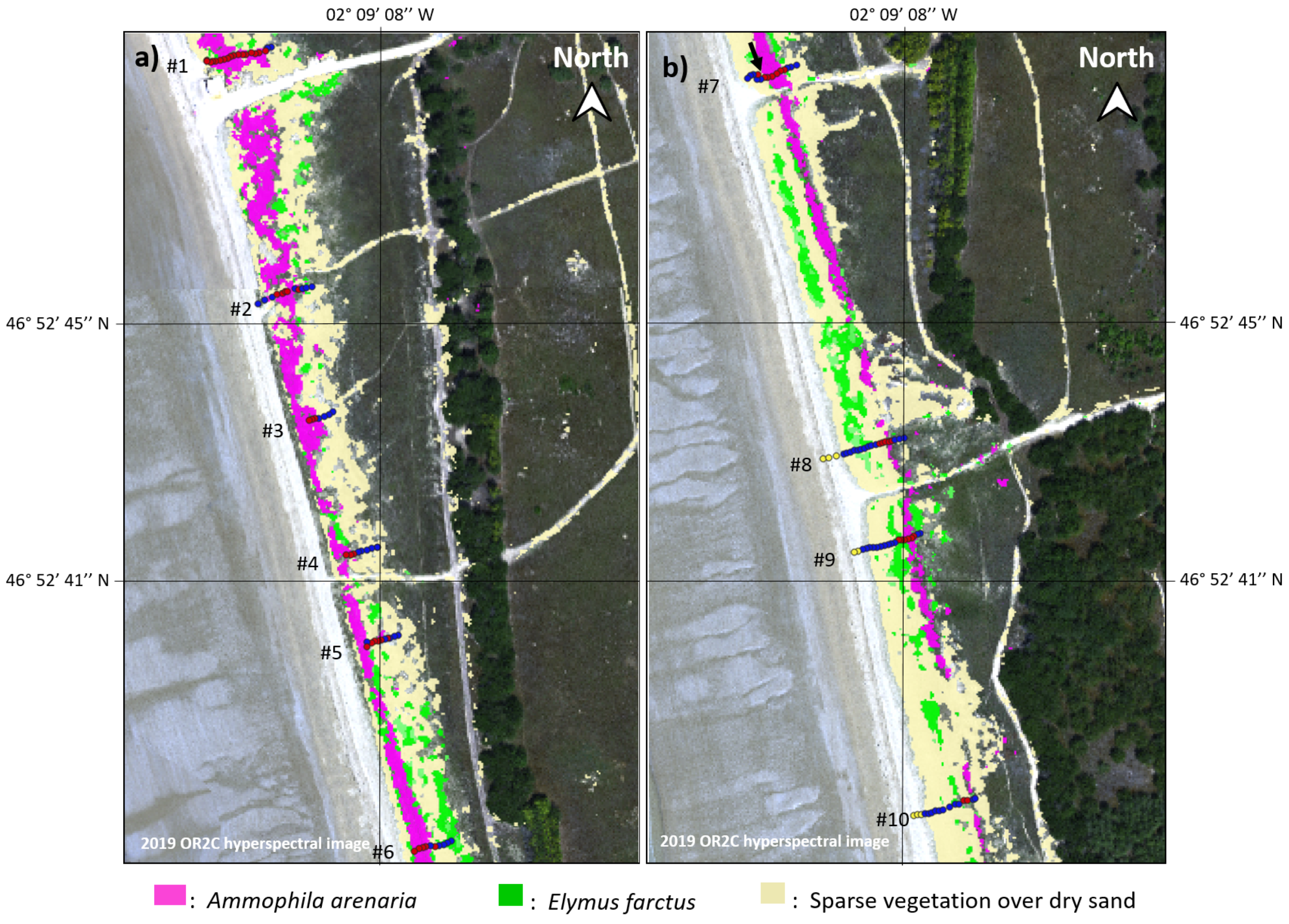
2.1.2. Validation Areas
Barbâtre
Pays-de-Monts
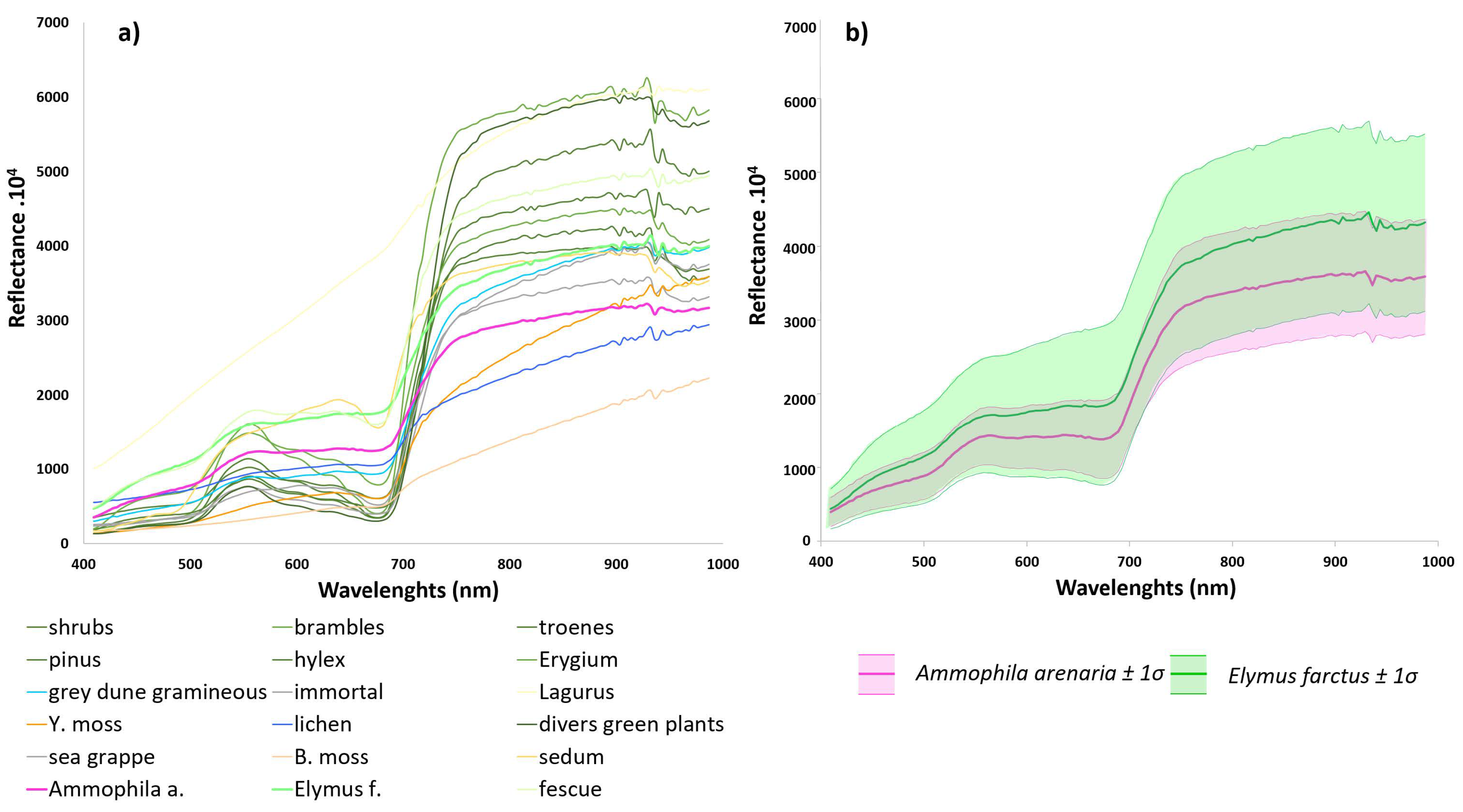
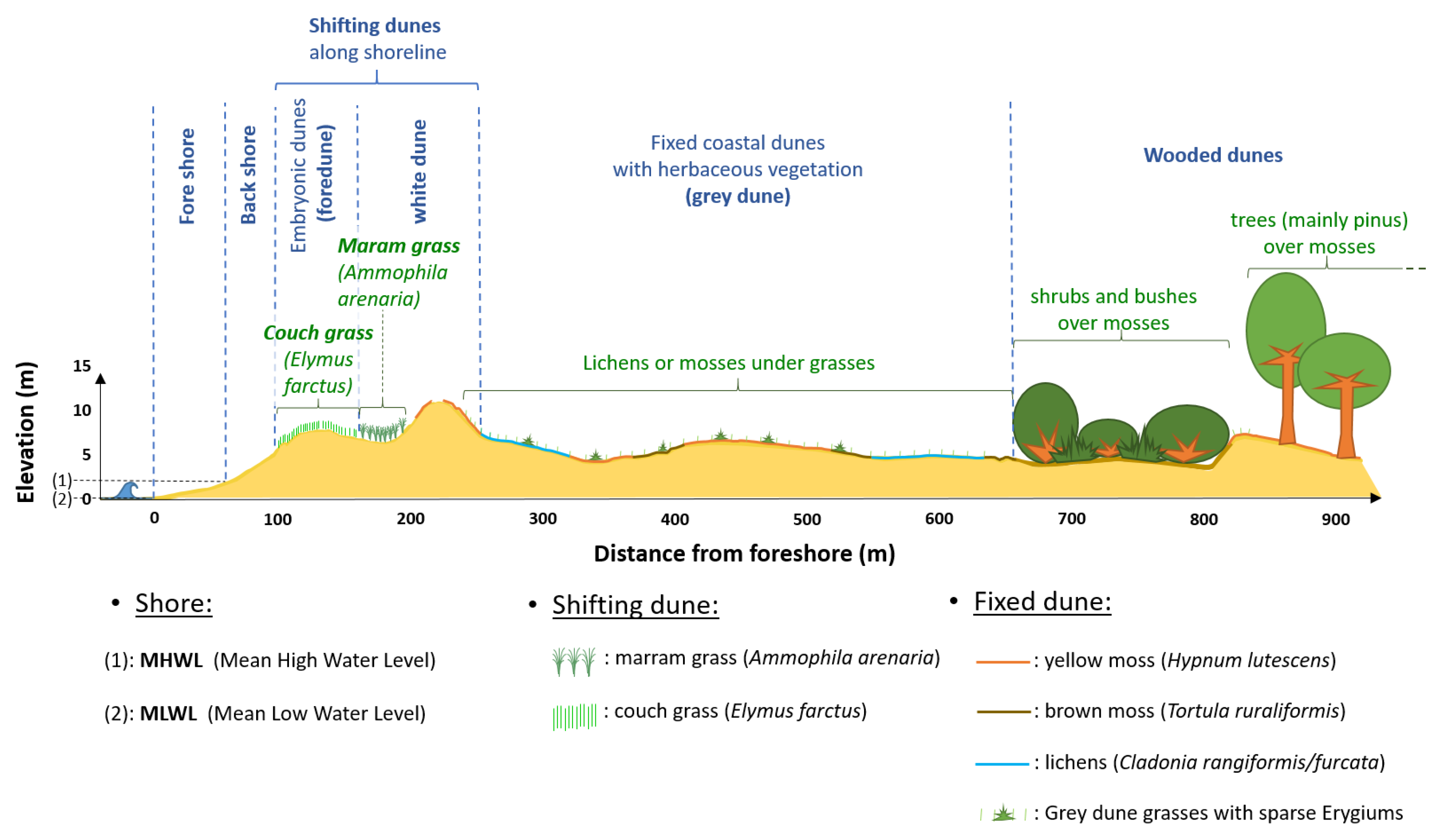
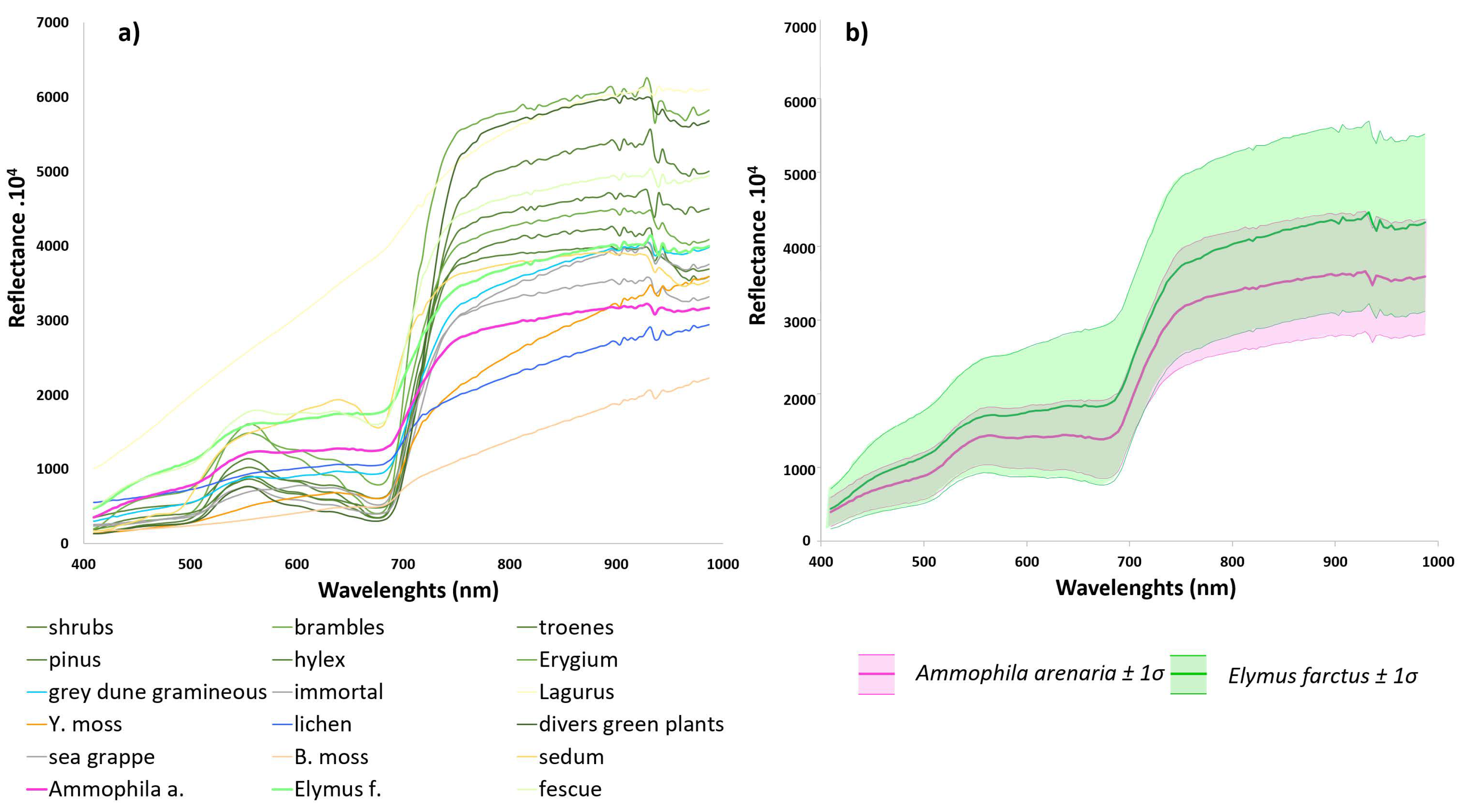
2.1.3. Description of Typical Vegetation Cover
2.2. Material
2.3. Field Measurements
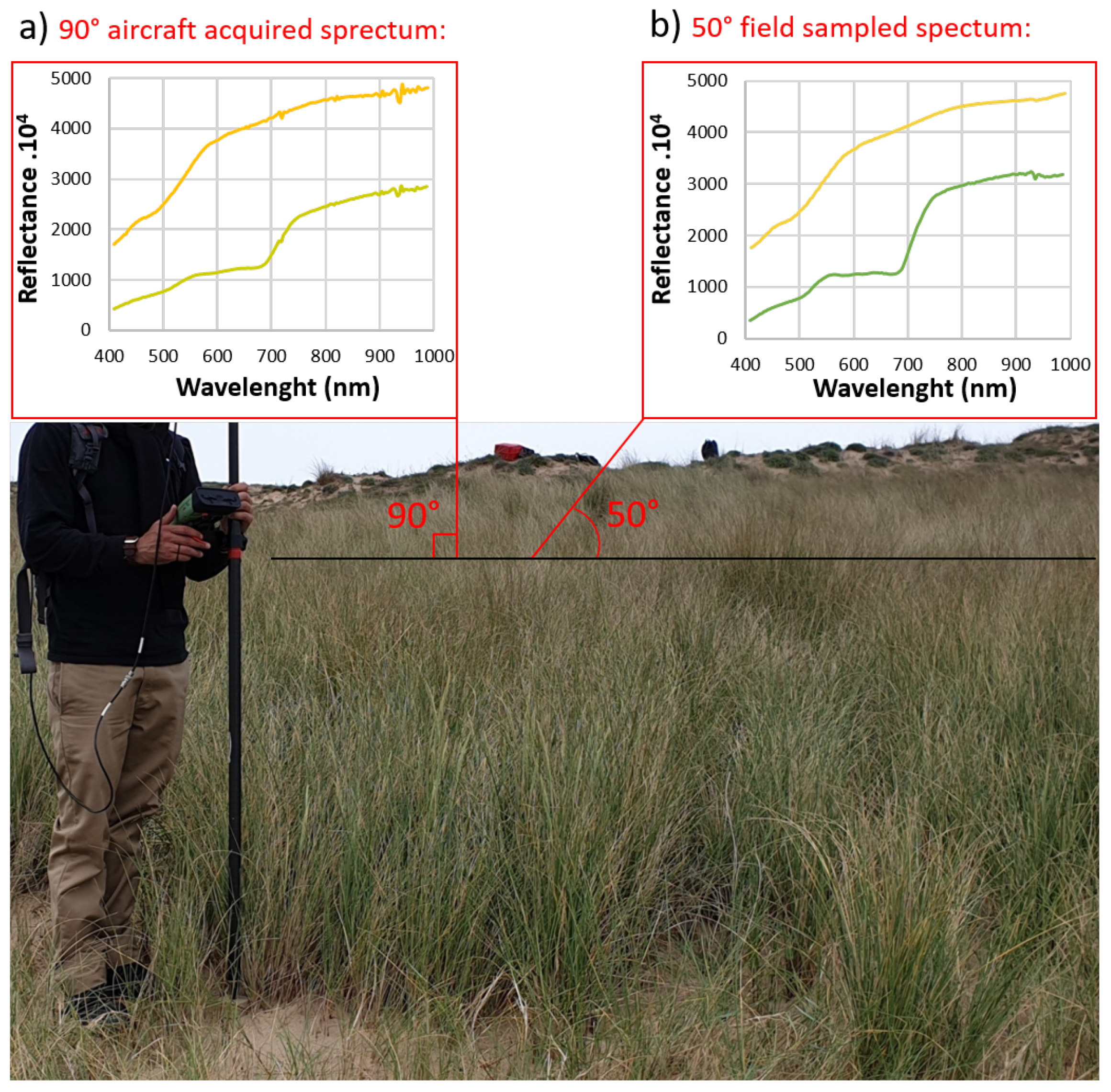
2.3.1. Spectral Field Measurements
2.3.2. dGPS Field Measurements
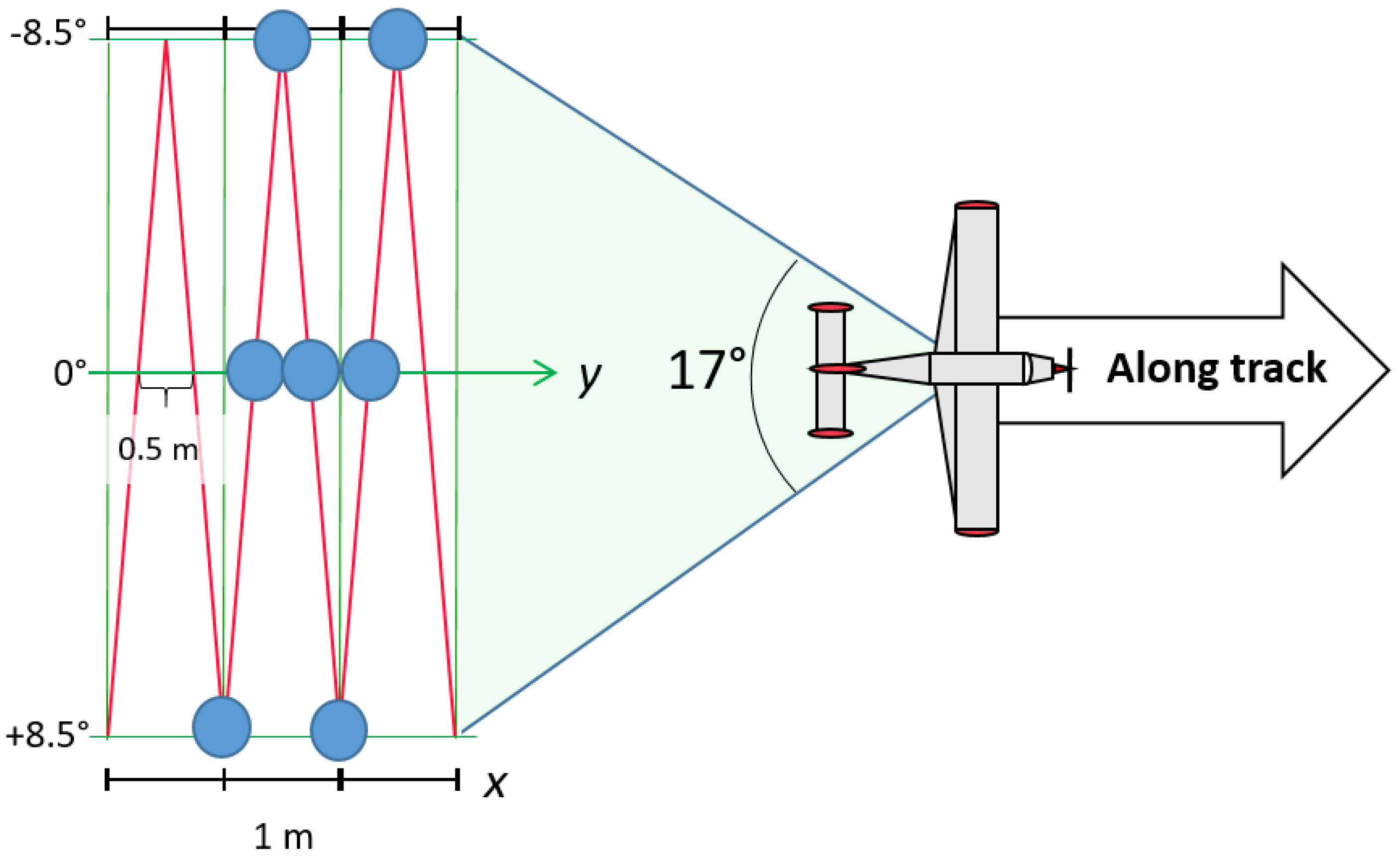
2.4. Airborne Acquisitions
3. Methodology
3.1. LiDAR Processing
3.1.1. Discrete LiDAR
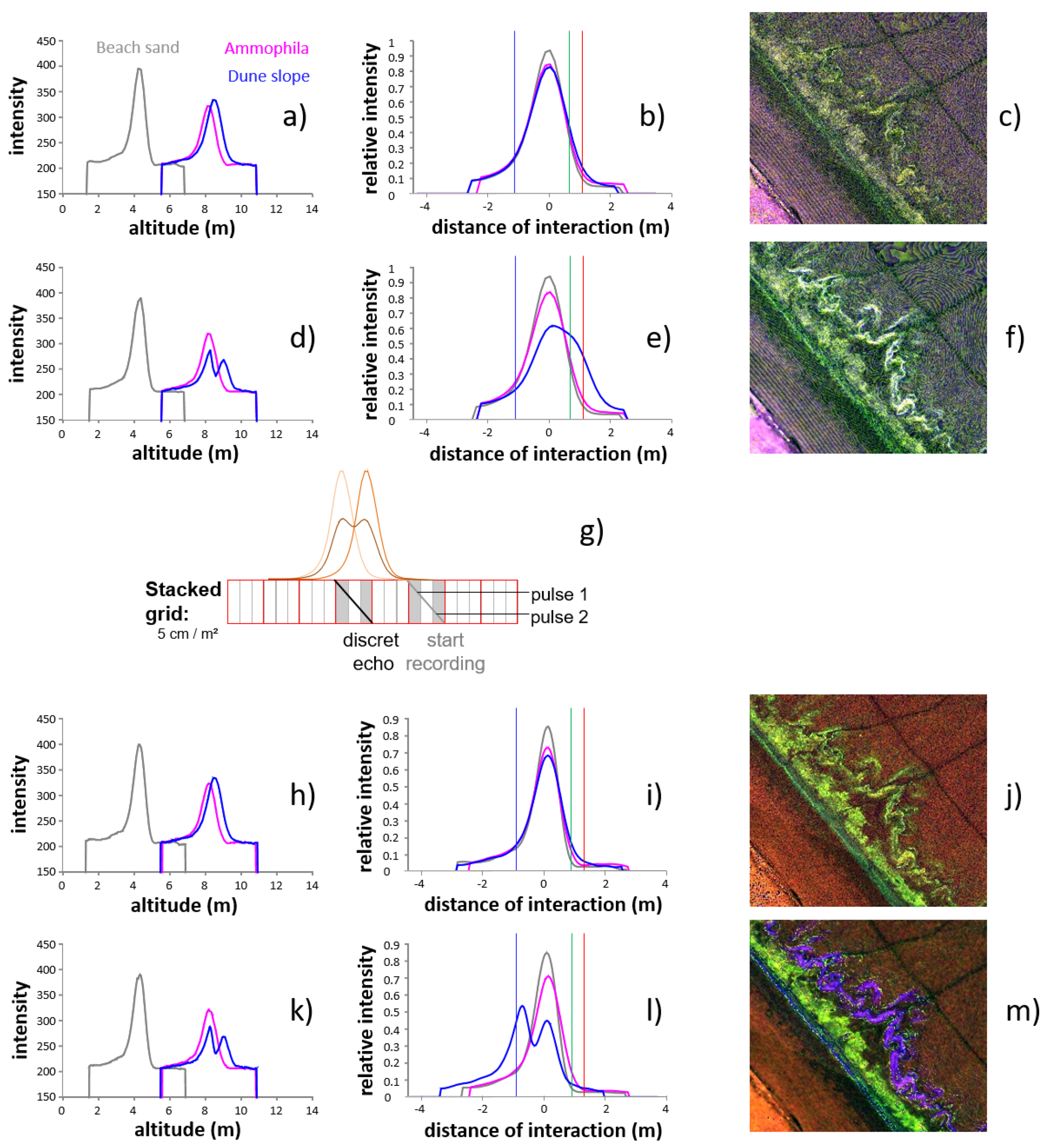
3.1.2. FWF LiDAR
3.2. Hyperspectral Processing
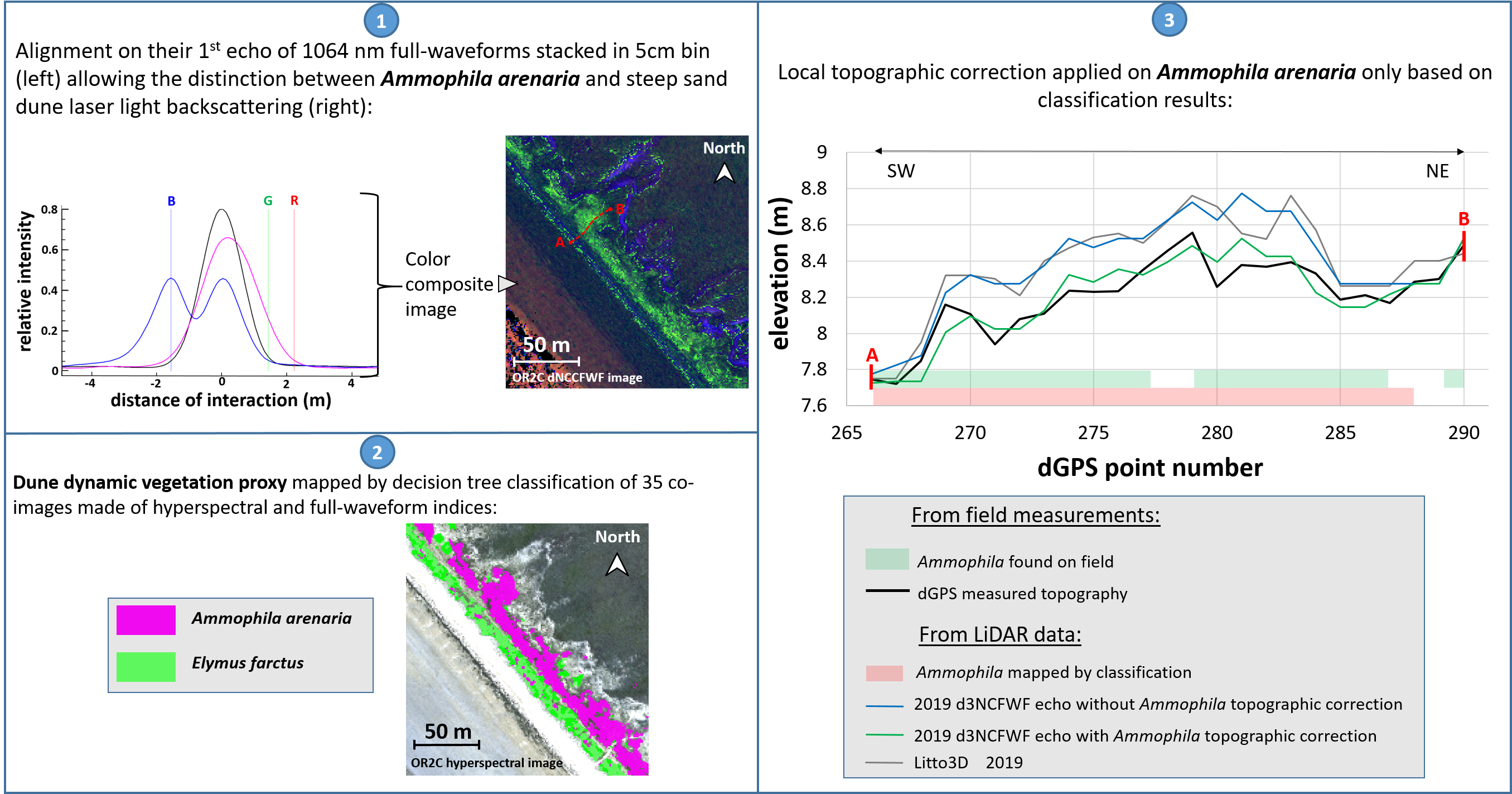
3.3. Classification of Main Dune Vegetation Proxy
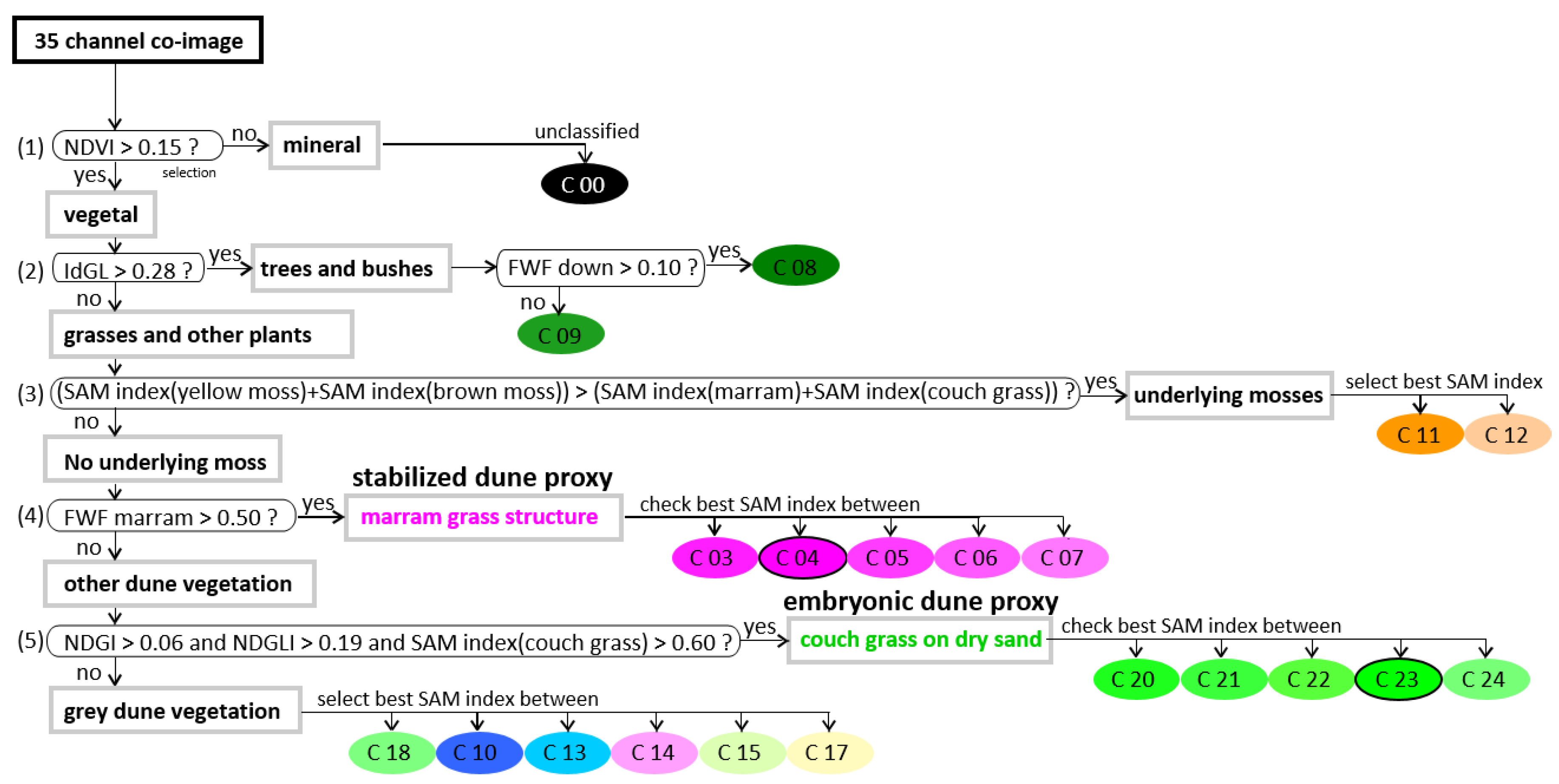
3.3.1. Straightforward Hierarchical Classification of Dune Proxies by Combination of FWF and HSI Data
3.3.2. Classification Validity Assessment
3.4. Topographic Analysis Methodology
4. Results
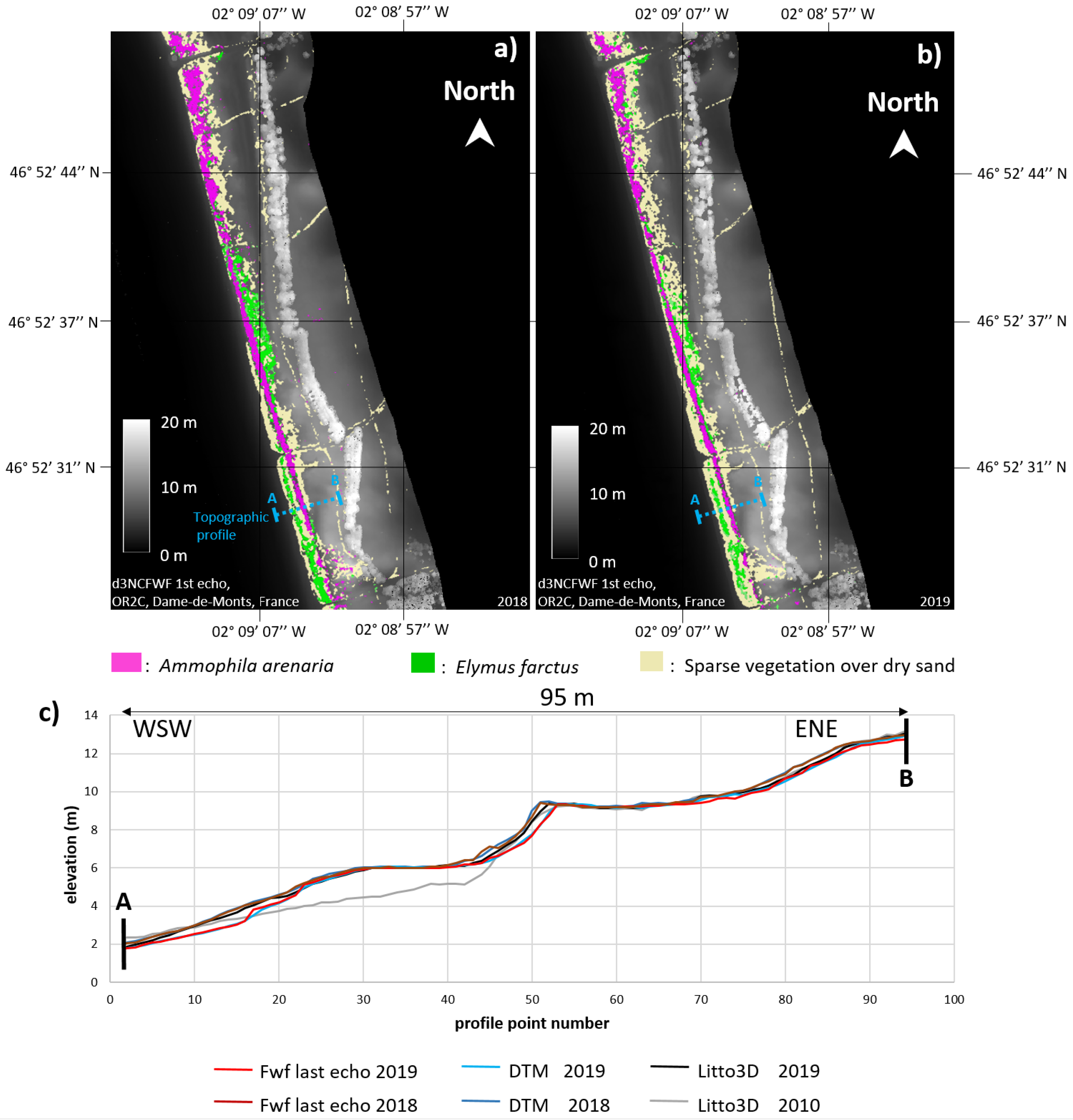
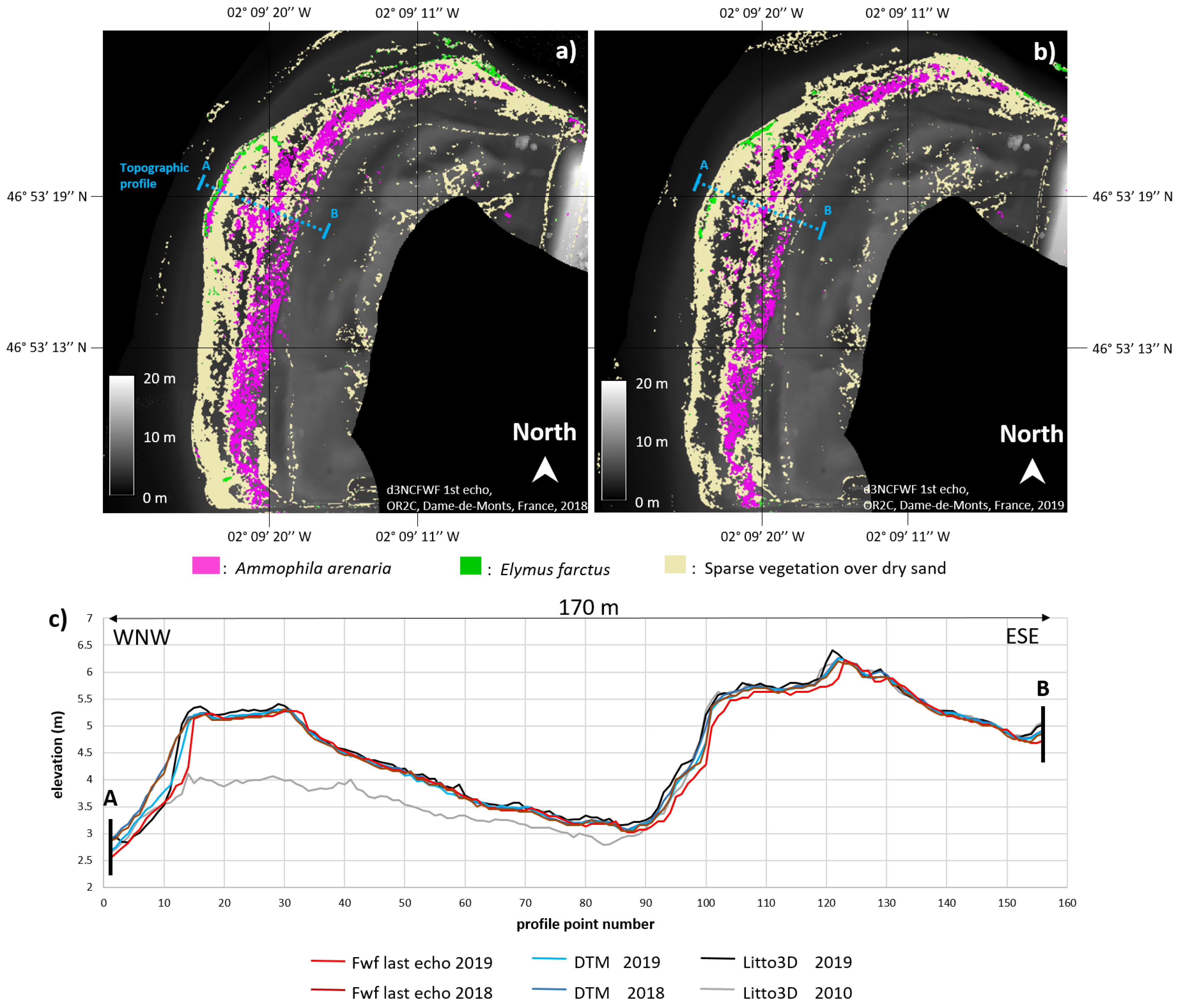
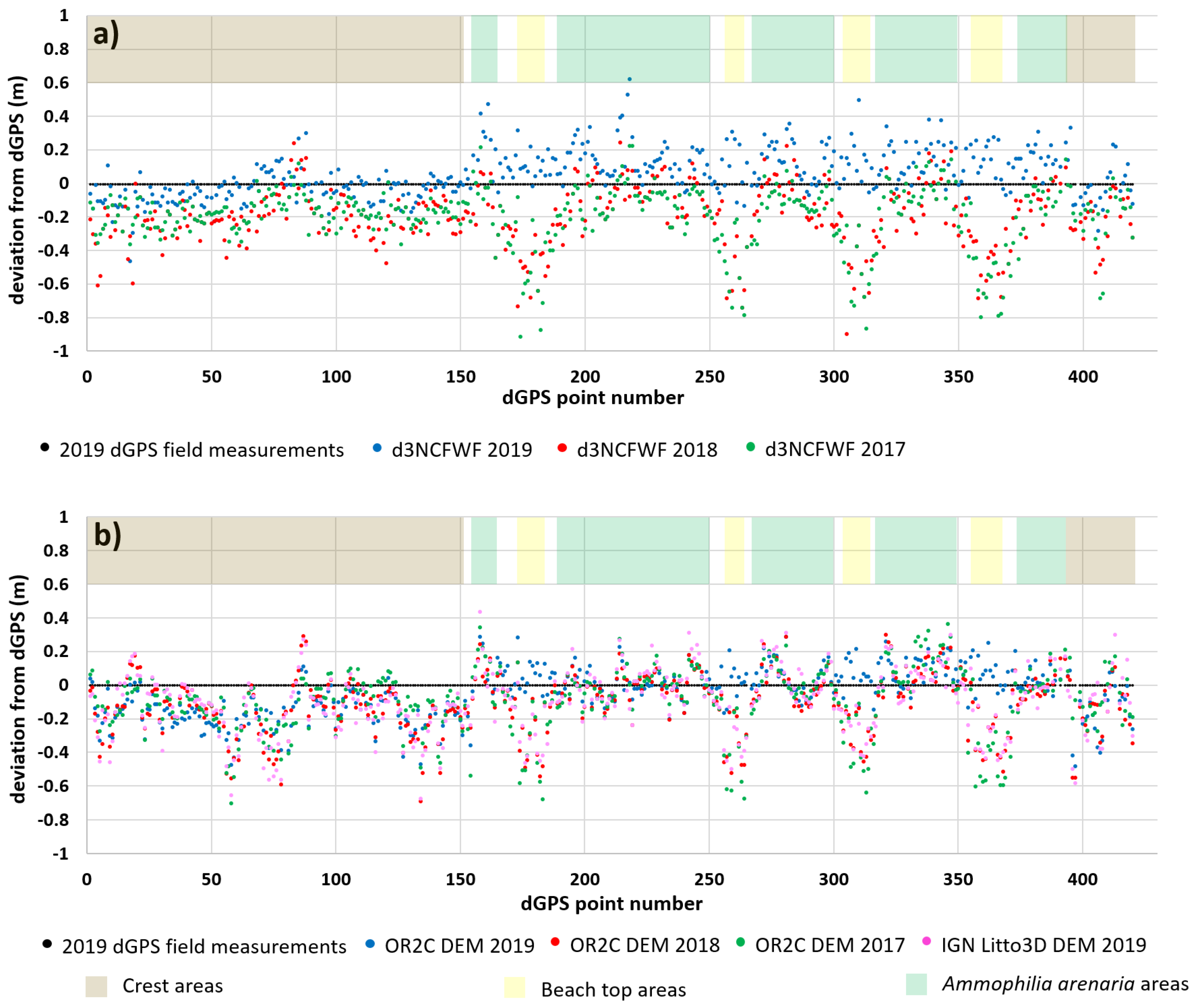
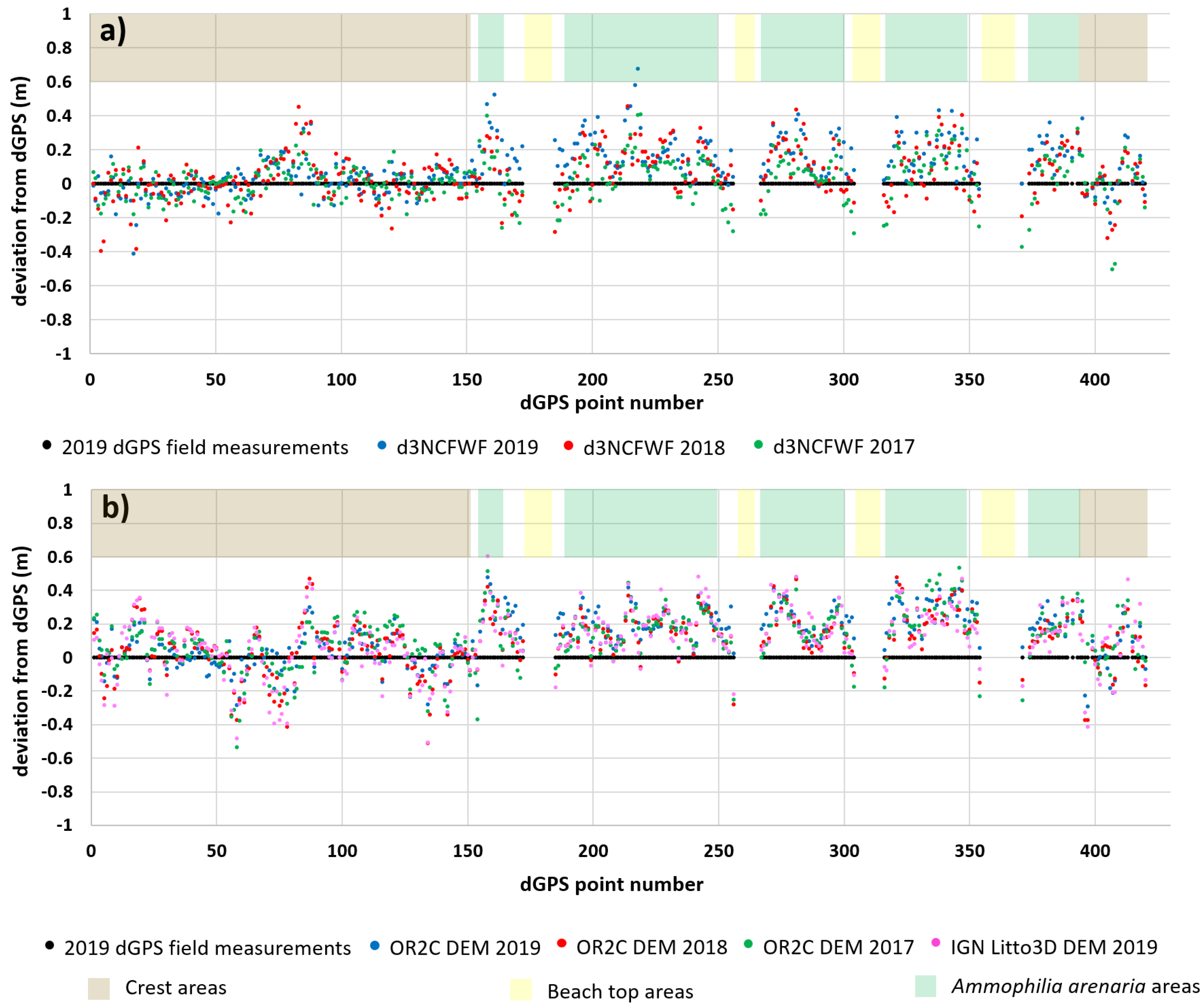
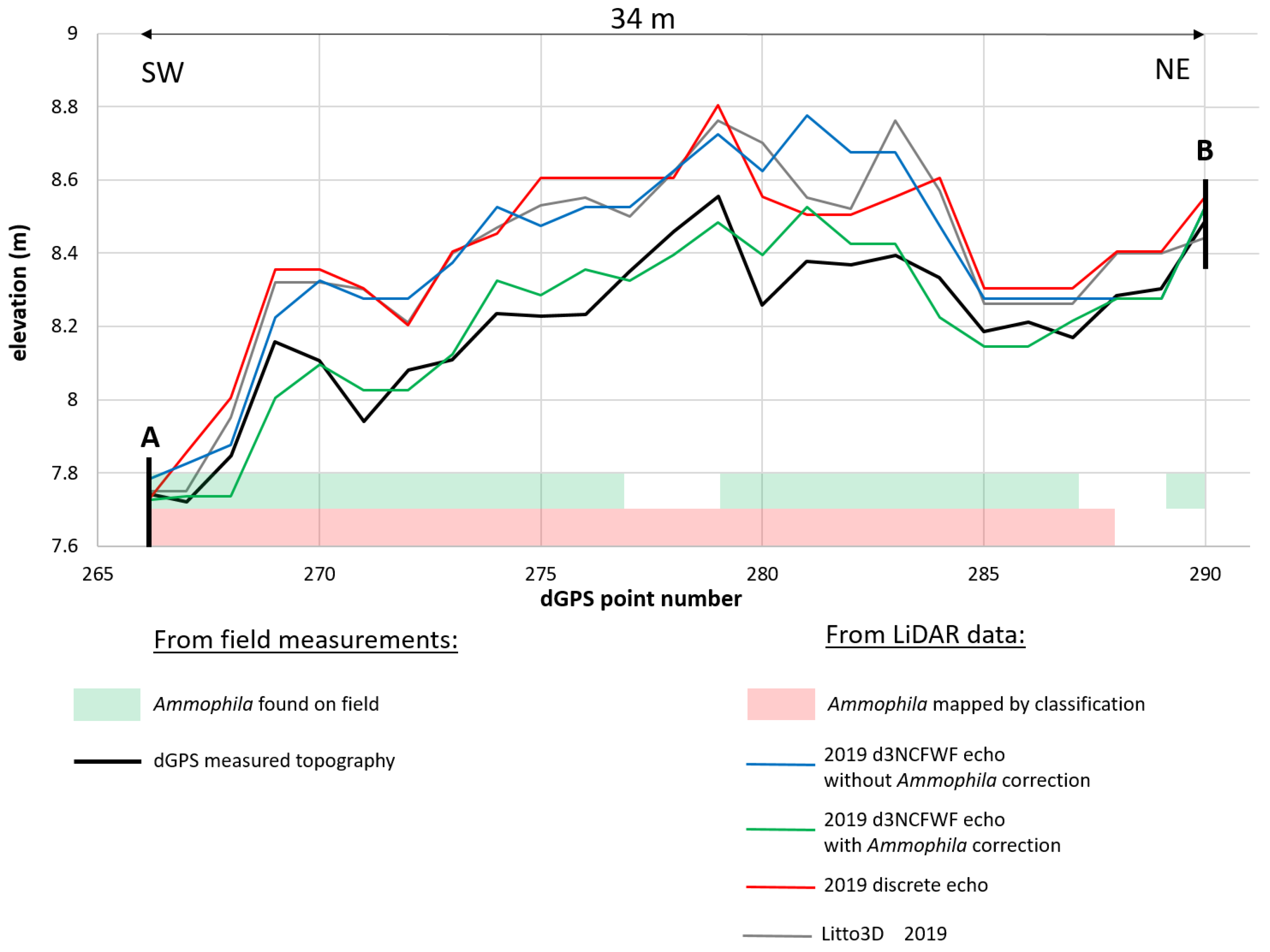
4.1. Topographic Analysis: Comparison between LiDAR Data and dGPS Field Measurements
4.2. Training Area
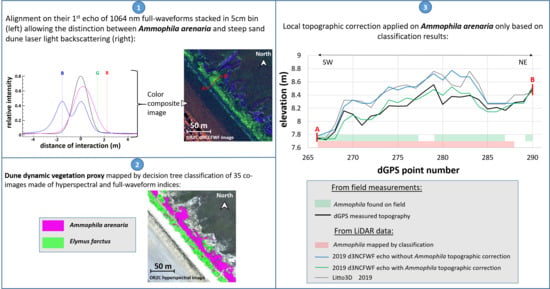
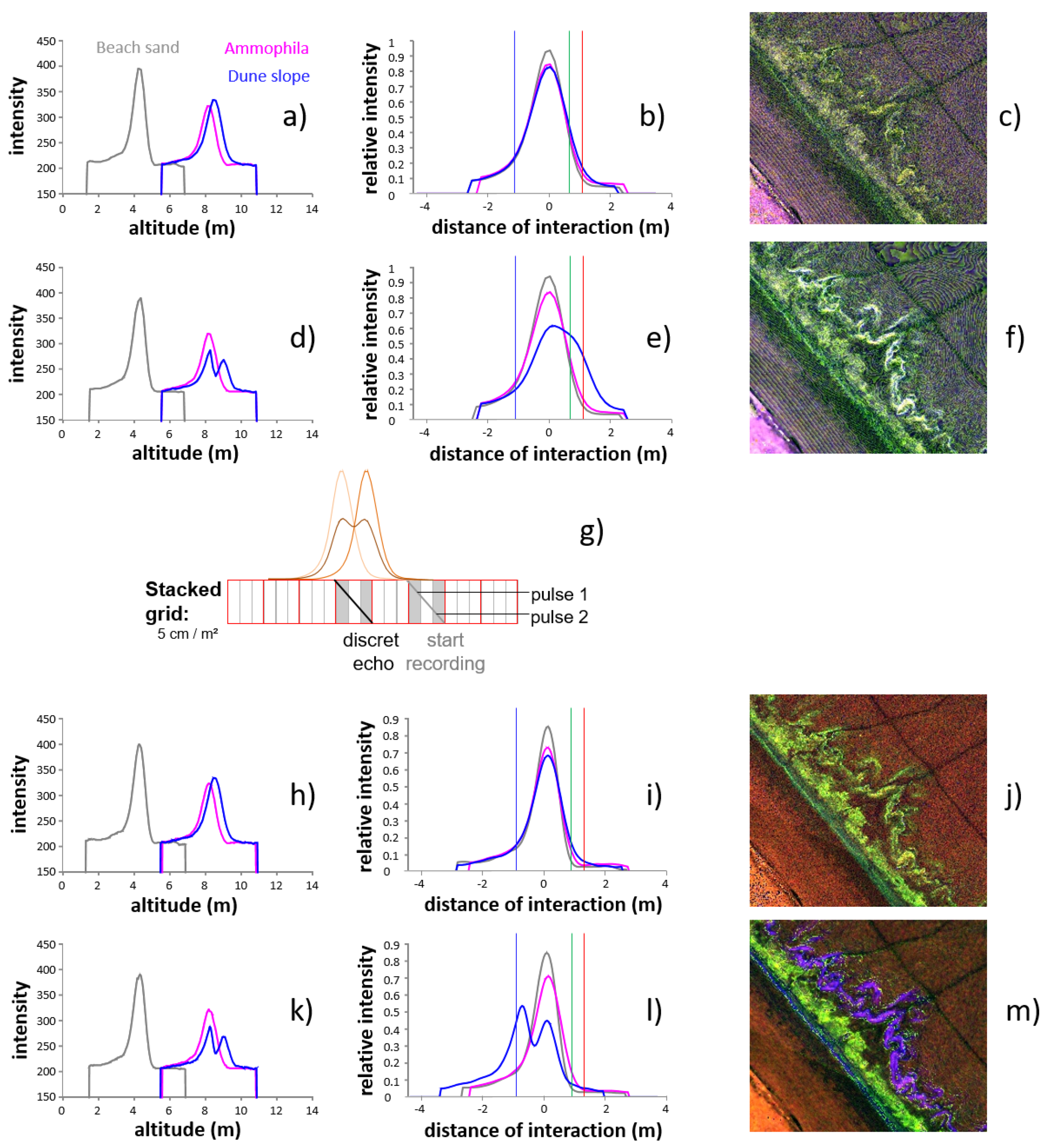
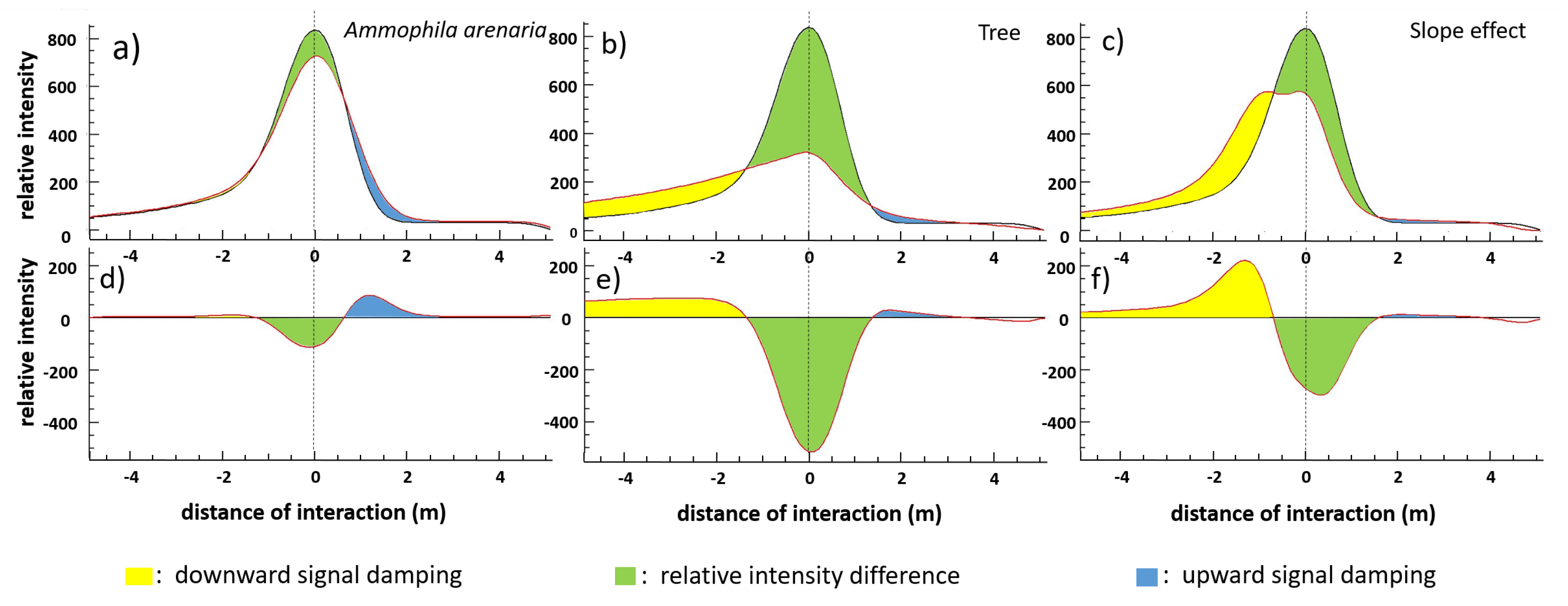
4.2.1. Ammophila arenaria Selection by FWF
4.2.2. Ammophila arenaria Topographic Correction
4.3. Validation Area
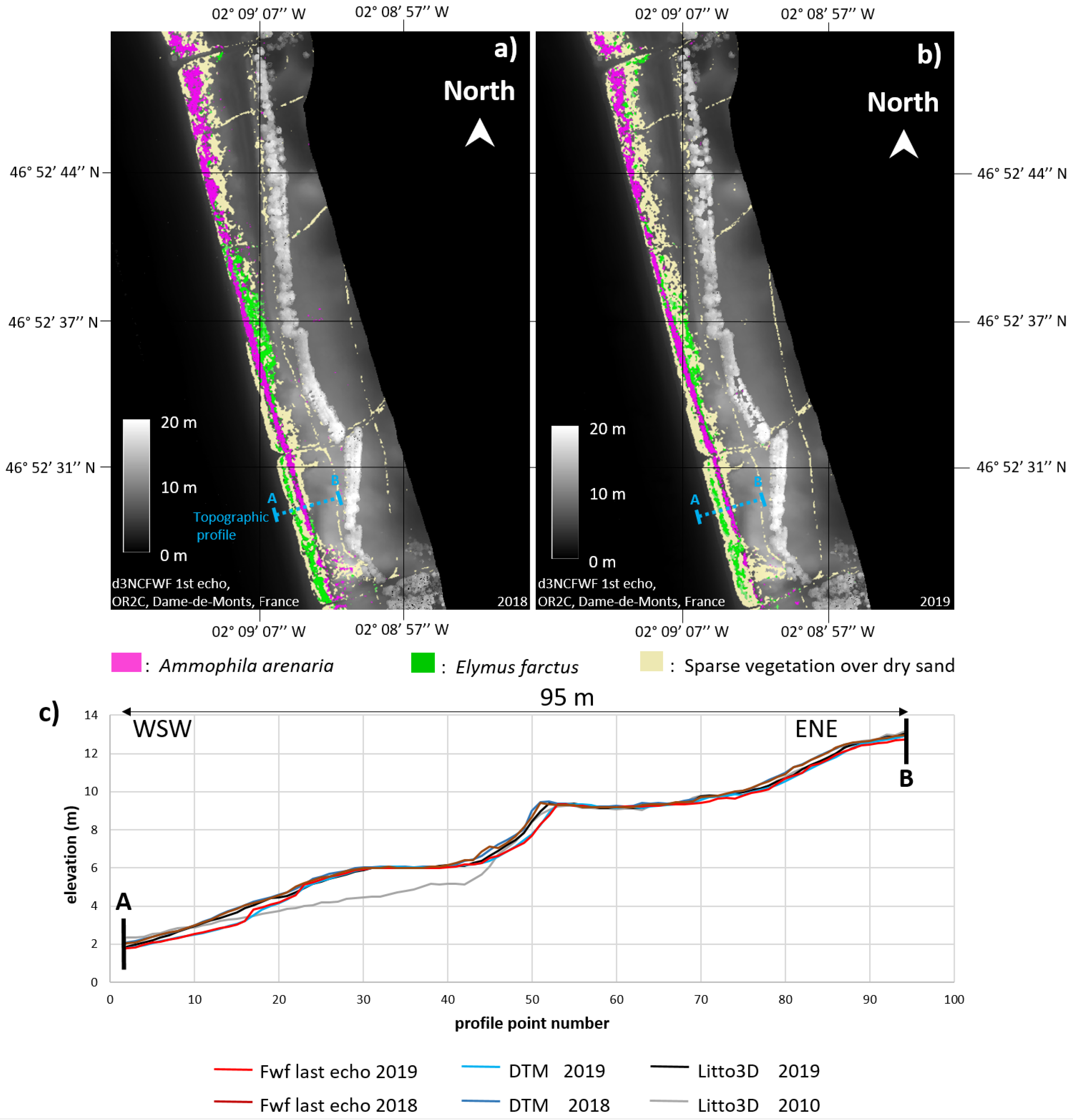
4.3.1. Notre-Dame-de-Monts
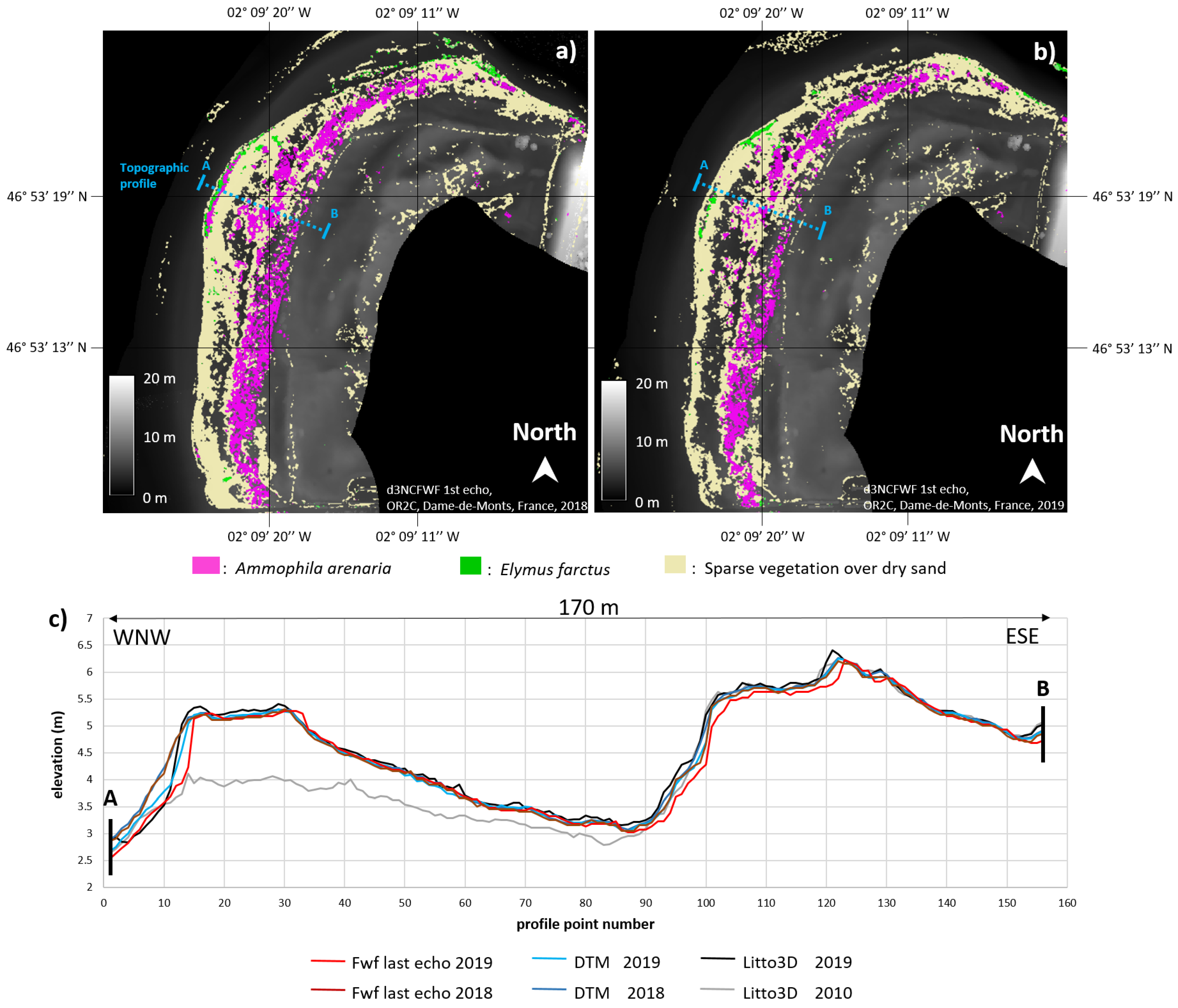
4.3.2. Barre-de-Monts
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dolan, R.; Hayden, B.P.; May, P.; May, S. The reliability of shoreline change measurements from aerial photographs. Shore Beach 1980, 48, 22–29. [Google Scholar]
- James, R.J. From beaches to beach environments: Linking the ecology, human-use and management of beaches in Australia. Ocean Coast. Manag. 2000, 43, 495–514. [Google Scholar]
- Kandrot, S. Beach-dune morphological relationships at Youghal Beach, Cork. In Bridging the Geographic Information Sciences: International AGILE’2012 Conference, Avignon, France, 24–27 April 2012; Gensel, J., Josselin, D., Vandenbroucke, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 367–390. [Google Scholar] [CrossRef]
- Arkema, K.K.; Guannel, G.; Verutes, G.; Wood, S.A.; Guerry, A.; Ruckelshaus, M.; Kareiva, P.; Lacayo, M.; Silver, J.M. Coastal habitats shield people and property from sea-level rise and storms. Nat. Clim. Chang. 2013, 3, 913–918. [Google Scholar]
- Juigner, M.; Robin, M.; Fattal, P.; Maanan, M.; Debaine, F.; Le Guern, C.; Gouguet, L.; Baudouin, V. Cinématique d’un trait de côte sableux en Vendée entre 1920 et 2010. Méthode et analyse. Dynamiques Environnementales-Journal international des géosciences et de l’environnement. L’homme Dyn. Littorale Maîtrise Adapt. 2013, 30, 29–39. [Google Scholar]
- Masselink, G.; Short, A.D. The effect of tide range on beach morphodynamics and morphology: A conceptual beach model. J. Coast. Res. 1993, 9, 785–800. [Google Scholar]
- Masselink, G. Alongshore variation in beach cusp morphology in a coastal embayment. Earth Surf. Process. Landf. J. Br. Geomorphol. Res. Group 1999, 24, 335–347. [Google Scholar]
- Ruessink, B.; Terwindt, J. The behaviour of nearshore bars on the time scale of years: A conceptual model. Mar. Geol. 2000, 163, 289–302. [Google Scholar]
- Quartel, S.; Kroon, A.; Ruessink, B. Seasonal accretion and erosion patterns of a microtidal sandy beach. Mar. Geol. 2008, 250, 19–33. [Google Scholar]
- Ortega-Sánchez, M.; Bramato, S.; Quevedo, E.; Mans, C.; Losada, M. Atmospheric-hydrodynamic coupling in the nearshore. Geophys. Res. Lett. 2008, 35. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.h.; Nicholls, R.J.; Birkemeier, W.A. Storm-driven variability of the beach-nearshore profile at Duck, North Carolina, USA, 1981–1991. Mar. Geol. 1998, 148, 163–177. [Google Scholar]
- Del Río, L.; Plomaritis, T.A.; Benavente, J.; Valladares, M.; Ribera, P. Establishing storm thresholds for the Spanish Gulf of Cádiz coast. Geomorphology 2012, 143, 13–23. [Google Scholar]
- Ruggiero, P. Impacts of climate change on coastal erosion and flood probability in the US Pacific Northwest. In Solutions to Coastal Disasters 2008; American Society of Civil Engineers: Reston, VA, USA, 2008; pp. 158–169. [Google Scholar]
- Zhang, K.; Douglas, B.C.; Leatherman, S.P. Global warming and coastal erosion. Clim. Chang. 2004, 64, 41. [Google Scholar]
- Jackson, N.L.; Nordstrom, K.F.; Feagin, R.A.; Smith, W.K. Coastal geomorphology and restoration. Geomorphology 2013, 199, 1–7. [Google Scholar]
- Mann, T.; Westphal, H. Multi-decadal shoreline changes on Takú Atoll, Papua New Guinea: Observational evidence of early reef island recovery after the impact of storm waves. Geomorphology 2016, 257, 75–84. [Google Scholar]
- Luijendijk, A.; Hagenaars, G.; Ranasinghe, R.; Baart, F.; Donchyts, G.; Aarninkhof, S. The state of the world’s beaches. Sci. Rep. 2018, 8, 6641. [Google Scholar]
- Corlay, J.P. Géographie sociale, géographie du littoral. Norois 1995, 165, 247–265. [Google Scholar]
- McGranahan, G.; Balk, D.; Anderson, B. The rising tide: Assessing the risks of climate change and human 97 settlements in low elevation coastal zones. Environ. Urban. 2007, 19, 17–37. [Google Scholar]
- Crowell, M.; Edelman, S.; Coulton, K.; McAfee, S. How many people live in coastal areas? J. Coast. Res. 2007, 235, iii–vi. [Google Scholar]
- Le Berre, I.; Maulpoix, A.; Thériault, M.; Gourmelon, F. A probabilistic model of residential urban development along the French Atlantic coast between 1968 and 2008. Land Use Policy 2016, 50, 461–478. [Google Scholar]
- Meur-Férec, C.; Deboudt, P.; Morel, V. Coastal risks in France: An integrated method for evaluating vulnerability. J. Coast. Res. 2008, 24, 178–189. [Google Scholar]
- Douglas, B.C.; Crowell, M. Long-term shoreline position prediction and error propagation. J. Coast. Res. 2000, 16, 145–152. [Google Scholar]
- Deboudt, P. Towards coastal risk management in France. Ocean Coast. Manag. 2010, 53, 366–378. [Google Scholar] [CrossRef]
- Kerguillec, R.; Audère, M.; Baltzer, A.; Debaine, F.; Fattal, P.; Juigner, M.; Launeau, P.; Le Mauff, B.; Luquet, F.; Maanan, M.; et al. Monitoring and management of coastal hazards: Creation of a regional observatory of coastal erosion and storm surges in the pays de la Loire region (Atlantic coast, France). Ocean Coast. Manag. 2019, 181, 104904. [Google Scholar] [CrossRef]
- Boak, E.H.; Turner, I.L. Shoreline definition and detection: A review. J. Coast. Res. 2005, 21, 688–703. [Google Scholar] [CrossRef] [Green Version]
- Hesp, P. Foredunes and blowouts: Initiation, geomorphology and dynamics. Geomorphology 2002, 48, 245–268. [Google Scholar] [CrossRef]
- Barbour, M.G.; De Jong, T.M.; Pavlik, B.M. Marine beach and dune plant communities. In Physiological Ecology of North American Plant Communities; Springer: Berlin/Heidelberg, Germany, 1985; pp. 296–322. [Google Scholar]
- Duffaud, M.H. Végétation des dunes littorales atlantiques de l’Adour a Noirmoutier. Rev. For. Française 1998, 50, 328–348. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Yu, K.B. A conceptual model of coastal dune ecology synthesizing spatial gradients of vegetation, soil, and geomorphology. Plant Ecol. 2009, 202, 135. [Google Scholar] [CrossRef]
- Durán, O.; Moore, L.J. Vegetation controls on the maximum size of coastal dunes. Proc. Natl. Acad. Sci. USA 2013, 110, 17217–17222. [Google Scholar] [CrossRef] [Green Version]
- Maun, A.; Maun, M.A. The Biology of Coastal Sand Dunes; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Hart, A.T.; Hilton, M.J.; Wakes, S.J.; Dickinson, K.J. The impact of Ammophila arenaria foredune development on downwind aerodynamics and parabolic dune development. J. Coast. Res. 2012, 28, 112–122. [Google Scholar] [CrossRef]
- Hewett, D. The colonization of sand dunes after stabilization with marram grass (Ammophila arenaria). J. Ecol. 1970, 58, 653–668. [Google Scholar] [CrossRef]
- Hertling, U.; Lubke, R. Use of Ammophila arenaria for dune stabilization in South Africa and its current distribution—Perceptions and problems. Environ. Manag. 1999, 24, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Psuty, N. The coastal foredune: A morphological basis for regional coastal dune development. In Coastal Dunes; Springer: Berlin, Germany, 2008; pp. 11–27. [Google Scholar]
- Schmid, K.A.; Hadley, B.C.; Wijekoon, N. Vertical accuracy and use of topographic LIDAR data in coastal marshes. J. Coast. Res. 2011, 27, 116–132. [Google Scholar] [CrossRef]
- Spaete, L.P.; Glenn, N.F.; Derryberry, D.R.; Sankey, T.T.; Mitchell, J.J.; Hardegree, S.P. Vegetation and slope effects on accuracy of a LiDAR-derived DEM in the sagebrush steppe. Remote Sens. Lett. 2011, 2, 317–326. [Google Scholar] [CrossRef]
- Bonn, F.; Rochon, G. Précis de télédétection, Volume 1: Principes et Méthodes; PUQ/AUPELF Sainte-Foy; Presses de l’Université du Québec: Quebec, QC, Canada, 1992. [Google Scholar]
- Ustin, S.L.; DiPietro, D.; Olmstead, K.; Underwood, E.; Scheer, G.J. Hyperspectral remote sensing for invasive species detection and mapping. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, Toronto, ON, Canada, 24–28 June 2002; Volume 3, pp. 1658–1660. [Google Scholar]
- Govender, M.; Chetty, K.; Bulcock, H. A review of hyperspectral remote sensing and its application in vegetation and water resource studies. Water Sa 2007, 33. [Google Scholar] [CrossRef] [Green Version]
- Hirano, A.; Madden, M.; Welch, R. Hyperspectral image data for mapping wetland vegetation. Wetlands 2003, 23, 436–448. [Google Scholar] [CrossRef]
- Adam, E.; Mutanga, O.; Rugege, D. Multispectral and hyperspectral remote sensing for identification and mapping of wetland vegetation: A review. Wetl. Ecol. Manag. 2010, 18, 281–296. [Google Scholar] [CrossRef]
- Schmidtlein, S.; Sassin, J. Mapping of continuous floristic gradients in grasslands using hyperspectral imagery. Remote Sens. Environ. 2004, 92, 126–138. [Google Scholar] [CrossRef]
- Treitz, P.M.; Howarth, P.J. Hyperspectral remote sensing for estimating biophysical parameters of forest ecosystems. Prog. Phys. Geogr. 1999, 23, 359–390. [Google Scholar] [CrossRef]
- Clark, M.L.; Roberts, D.A.; Clark, D.B. Hyperspectral discrimination of tropical rain forest tree species at leaf to crown scales. Remote Sens. Environ. 2005, 96, 375–398. [Google Scholar] [CrossRef]
- Ramdani, F. Urban vegetation mapping from fused hyperspectral image and LiDAR data with application to monitor urban tree heights. J. Geogr. Inf. Syst. 2013, 4, 404–408. [Google Scholar] [CrossRef]
- Launeau, P.; Kassouk, Z.; Debaine, F.; Roy, R.; Mestayer, P.G.; Boulet, C.; Rouaud, J.M.; Giraud, M. Airborne hyperspectral mapping of trees in an urban area. Int. J. Remote Sens. 2017, 38, 1277–1311. [Google Scholar] [CrossRef]
- Schmidt, K.; Skidmore, A.; Kloosterman, E.; Van Oosten, H.; Kumar, L.; Janssen, J. Mapping coastal vegetation using an expert system and hyperspectral imagery. Photogramm. Eng. Remote Sens. 2004, 70, 703–715. [Google Scholar] [CrossRef]
- Baltsavias, E.P. Airborne laser scanning: Basic relations and formulas. ISPRS J. Photogramm. Remote Sens. 1999, 54, 199–214. [Google Scholar] [CrossRef]
- Persson, Å.; Söderman, U.; Töpel, J.; Ahlberg, S. Visualization and analysis of full-waveform airborne laser scanner data. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2005, 36, 103–109. [Google Scholar]
- Mallet, C.; Chauve, A.; Bretar, F. Analyse et traitement d’ondes lidar pour la cartographie et la reconnaissance de formes: Application au milieu urbain. Reconnaiss. Formes Intell. Artif. RFIA 2008. Available online: https://hal.archives-ouvertes.fr/hal-02384672/ (accessed on 16 December 2020).
- Mallet, C.; Bretar, F. Full-waveform topographic lidar: State-of-the-art. ISPRS J. Photogramm. Remote Sens. 2009, 64, 1–16. [Google Scholar] [CrossRef]
- Launeau, P.; Giraud, M.; Ba, A.; Moussaoui, S.; Robin, M.; Debaine, F.; Lague, D.; Le Menn, E. Full-Waveform LiDAR Pixel Analysis for Low-Growing Vegetation Mapping of Coastal Foredunes in Western France. Remote Sens. 2018, 10, 669. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, T.; Steinvall, O.; Letalick, D. Signature Simulation and Signal Analysis for 3-D Laser Radar; Swedish Defence Research Agency: Stockholm, Sweden, 2001; pp. 7–8.
- Jutzi, B.; Stilla, U. Range determination with waveform recording laser systems using a Wiener Filter. ISPRS J. Photogramm. Remote Sens. 2006, 61, 95–107. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Kennedy-Bowdoin, T.; Jones, M.O.; Martin, R.E.; Boardman, J.W.; Field, C.B. Carnegie airborne observatory: In-flight fusion of hyperspectral imaging and waveform light detection and ranging for three-dimensional studies of ecosystems. J. Appl. Remote Sens. 2007, 1, 013536. [Google Scholar] [CrossRef]
- Koetz, B.; Sun, G.; Morsdorf, F.; Ranson, K.; Kneubühler, M.; Itten, K.; Allgöwer, B. Fusion of imaging spectrometer and LIDAR data over combined radiative transfer models for forest canopy characterization. Remote Sens. Environ. 2007, 106, 449–459. [Google Scholar] [CrossRef]
- Dalponte, M.; Bruzzone, L.; Gianelle, D. Fusion of hyperspectral and LIDAR remote sensing data for classification of complex forest areas. IEEE Trans. Geosci. Remote Sens. 2008, 46, 1416–1427. [Google Scholar] [CrossRef] [Green Version]
- Brell, M.; Segl, K.; Guanter, L.; Bookhagen, B. Hyperspectral and lidar intensity data fusion: A framework for the rigorous correction of illumination, anisotropic effects, and cross calibration. IEEE Trans. Geosci. Remote Sens. 2017, 55, 2799–2810. [Google Scholar] [CrossRef] [Green Version]
- Brell, M.; Segl, K.; Guanter, L.; Bookhagen, B. 3D hyperspectral point cloud generation: Fusing airborne laser scanning and hyperspectral imaging sensors for improved object-based information extraction. ISPRS J. Photogramm. Remote Sens. 2019, 149, 200–214. [Google Scholar] [CrossRef]
- Wang, H.; Glennie, C. Fusion of waveform LiDAR data and hyperspectral imagery for land cover classification. ISPRS J. Photogramm. Remote Sens. 2015, 108, 1–11. [Google Scholar] [CrossRef]
- Le Mauff, B.; Juigner, M.; Ba, A.; Robin, M.; Launeau, P.; Fattal, P. Coastal monitoring solutions of the geomorphological response of beach-dune systems using multi-temporal LiDAR datasets (Vendée coast, France). Geomorphology 2018, 304, 121–140. [Google Scholar] [CrossRef]
- Le Cornec, E.; Fiere, M.; Grunnet, N.; Peeters, P. Etude de Connaissance des Phénomènes D’érosion sur le Littoral Vendéen; Rapport de la Danish Hydraulics Institute France pour la DDE85; GEOS & DHI: Hørsholm, Denmark, 2008. [Google Scholar]
- Debaine, F.; Debaine, F.; Robin, M.; Roze, F.; Favennec, J.; Gouguet, L.; Prat, M.C. Aide à la Gestion Multifonctionnelle des Dunes Littorales Atlantiques par l’évaluation Cartographiée de Leur état de Conservation. Programme «Multidune», Rapport de Synthèse. 2012. Available online: https://www1.liteau.net/uploads/projet_documents/LITEAU_III_2007_Debaine_Synthese_long.pdf (accessed on 16 December 2020).
- Hapke, B. Bidirectional reflectance spectroscopy: 1. Theory. J. Geophys. Res. Solid Earth 1981, 86, 3039–3054. [Google Scholar] [CrossRef]
- Kerguillec, R.; Robin, P.F. Observatoire Regional des Risques Cotiers en Pays de la Loire. 2016. Available online: https://or2c.osuna.univ-nantes.fr/ (accessed on 16 December 2020).
- Kilian, J.; Haala, N.; Englich, M. Capture and evaluation of airborne laser scanner data. Int. Arch. Photogramm. Remote Sens. 1996, 31, 383–388. [Google Scholar]
- ENVI-LIDAR. Exelis Visual Information Solutions. Available online: https://www.harrisgeospatial.com/docs/using_envi_lidar_Home.html (accessed on 16 December 2020).
- Grilli, E.; Menna, F.; Remondino, F. A review of point clouds segmentation and classification algorithms. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2017, 42, 339. [Google Scholar] [CrossRef] [Green Version]
- Lohani, B.; Ghosh, S. Airborne LiDAR technology: A review of data collection and processing systems. Proc. Natl. Acad. Sci. India Sect. A Phys. Sci. 2017, 87, 567–579. [Google Scholar] [CrossRef]
- Wagner, W.; Ullrich, A.; Melzer, T.; Briese, C.; Kraus, K. From Single-Pulse to Full-Waveform Airborne Laser Scanners: Potential and Practical Challenges; Elsevier: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Wang, H.; Glennie, C.; Prasad, S. Voxelization of full waveform LiDAR data for fusion with hyperspectral imagery. In Proceedings of the 2013 IEEE International Geoscience and Remote Sensing Symposium-IGARSS, Melbourne, VIC, Australia, 21–26 July 2013; pp. 3407–3410. [Google Scholar]
- Hermosilla, T.; Coops, N.; Ruiz, L.; Moskal, L. Deriving pseudo-vertical waveforms from small-footprint full-waveform LiDAR data. Remote Sens. Lett. 2014, 5, 332–341. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Coops, N.C.; Innes, J.L.; Dai, J.; Ruan, H.; She, G. Tree species classification in subtropical forests using small-footprint full-waveform LiDAR data. Int. J. Appl. Earth Obs. Geoinf. 2016, 49, 39–51. [Google Scholar] [CrossRef]
- Jutzi, B.; Stilla, U. Laser pulse analysis for reconstruction and classification of urban objects. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2003, 34, 151–156. [Google Scholar]
- Bhabatosh, C. Digital Image Processing and Analysis; PHI Learning Pvt. Ltd.: New Delhi, Delhi, India, 2011. [Google Scholar]
- Despan, D.; Bedidi, A.; Cervelle, B.; Rudant, J.P. Bidirectional reflectance of Gaussian random surfaces and its scaling properties. Math. Geol. 1998, 30, 873–888. [Google Scholar] [CrossRef]
- Parrish, C.E.; Rogers, J.N.; Calder, B.R. Assessment of waveform features for lidar uncertainty modeling in a coastal salt marsh environment. IEEE Geosci. Remote Sens. Lett. 2013, 11, 569–573. [Google Scholar] [CrossRef]
- Hovi, A.; Korhonen, L.; Vauhkonen, J.; Korpela, I. LiDAR waveform features for tree species classification and their sensitivity to tree-and acquisition related parameters. Remote Sens. Environ. 2016, 173, 224–237. [Google Scholar] [CrossRef]
- Ba, A.; Patrick, L.; Robin, M.; Moussaoui, S.; Cyril, M.; Giraud, M.; Le Menn, E. Apport du LiDAR dans le géoréférencement d’images hyperspectrales en vue d’un couplage LiDAR/Hyperspectral. HAL 2015. Available online: https://hal-cea.archives-ouvertes.fr/LETG/hal-01271965v1 (accessed on 16 December 2020).
- Richter, R.; Schläpfer, D. Atmospheric and Topographic Correction (ATCOR Theoretical Background Document). DLR IB 2019, 564-03. [Google Scholar]
- Green, A.A.; Berman, M.; Switzer, P.; Craig, M.D. A transformation for ordering multispectral data in terms of image quality with implications for noise removal. IEEE Trans. Geosci. Remote Sens. 1988, 26, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Brantley, S.T.; Zinnert, J.C.; Young, D.R. Application of hyperspectral vegetation indices to detect variations in high leaf area index temperate shrub thicket canopies. Remote Sens. Environ. 2011, 115, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J.; Haas, R.; Deering, D.; Schell, J.; Harlan, J. Monitoring the vernal advancement and retrogradation (green wave effect) of natural vegetation, greenbelt. In MD: NASA/GSFC Type III Final Report; Remote Sensing Center, Texas A&M Univ.: College Station, TX, USA, 1974; p. 371. [Google Scholar]
- Wang, J.; Rich, P.M.; Price, K.P. Temporal responses of NDVI to precipitation and temperature in the central Great Plains, USA. Int. J. Remote Sens. 2003, 24, 2345–2364. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Kassouk, Z.; Launeau, P.; Roy, R.; Mestayer, P.; Rouaud, J.; Giraud, M. Urban Mapping Using Hyperspectral Hyspex Images over Nantes City, France; International Association for Spectral Imaging (IASIM): Dublin, Ireland, 2010. [Google Scholar]
- Kruse, F.A.; Lefkoff, A.; Boardman, J.; Heidebrecht, K.; Shapiro, A.; Barloon, P.; Goetz, A. The spectral image processing system (SIPS)-interactive visualization and analysis of imaging spectrometer data. In AIP Conference Proceedings; American Institute of Physics: Melville, NY, USA, 1993; Volume 283, pp. 192–201. [Google Scholar]
- Launeau, P.; Méléder, V.; Verpoorter, C.; Barillé, L.; Kazemipour-Ricci, F.; Giraud, M.; Jesus, B.; Le Menn, E. Microphytobenthos biomass and diversity mapping at different spatial scales with a hyperspectral optical model. Remote Sens. 2018, 10, 716. [Google Scholar] [CrossRef] [Green Version]
- Louvart, L.; Grateau, C. The Litto3D project. In Proceedings of the Europe Oceans 2005, Brest, France, 20–23 June 2005; Volume 2, pp. 1244–1251. [Google Scholar]
- Pastol, Y. Use of Airborne LIDAR Bathymetry for Coastal Hydrographic Surveying: The French Experience. J. Coast. Res. 2011, 2011, 6–18. [Google Scholar] [CrossRef]
- IGN, SHOM. Litto3D®—v 1.0: Spécifications Techniques; IGN, SHOM: Brest, France, 2015. [Google Scholar]
- Bertin, X.; Li, K.; Roland, A.; Breil, J.F.; Zhang, Y.; Chaumillon, E. Simulation numérique de la submersion marine associée à la tempête Xynthia (février 2010). In XIII Journées Nationales Génie Côtier–Génie Civil; Paralia: Nantes, France, 2012. [Google Scholar]
- Robin, M.; Juigner, M.; Luquet, F.; Audère, M. Assessing surface changes between shorelines from 1950 to 2011: The case of a 169-km sandy coast, Pays de la Loire (W France). J. Coast. Res. 2019, 88, 122–134. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Formula | FWHM/Sampling Interval (nm) | Authors |
|---|---|---|---|
| NDVI | − + | 70–110 | Rouse et al. (1974) [86] |
| NDVI | − + | 4.5/3.7 | Launeau et al. (2017) [48] |
| NDGLI | − + | 4.5/3.7 | Kassouk et al. (2010) [89] |
| NDGI | − + | 4.5/3.7 | Kassouk et al. (2010) [89] |
| IdGL | + − 1 | 4.5/3.7 | Kassouk et al. (2010) [89] |
| SAM index | 4.5/3.7 | Launeau et al. (2017) [48] |
| Year | 2018 | 2019 |
|---|---|---|
| pixels | 18,489 | 17,779 |
| C3: grey dune grasses | 2% | 1% |
| C4: Ammophila arenaria | 45% | 35% |
| C5: festuca | 6% | 4% |
| C6: Elymus farctus | 35% | 48% |
| C7: sparsed vegetation over exposed dry sand | 11% | 11% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frati, G.; Launeau, P.; Robin, M.; Giraud, M.; Juigner, M.; Debaine, F.; Michon, C. Coastal Sand Dunes Monitoring by Low Vegetation Cover Classification and Digital Elevation Model Improvement Using Synchronized Hyperspectral and Full-Waveform LiDAR Remote Sensing. Remote Sens. 2021, 13, 29. https://doi.org/10.3390/rs13010029
Frati G, Launeau P, Robin M, Giraud M, Juigner M, Debaine F, Michon C. Coastal Sand Dunes Monitoring by Low Vegetation Cover Classification and Digital Elevation Model Improvement Using Synchronized Hyperspectral and Full-Waveform LiDAR Remote Sensing. Remote Sensing. 2021; 13(1):29. https://doi.org/10.3390/rs13010029
Chicago/Turabian StyleFrati, Giovanni, Patrick Launeau, Marc Robin, Manuel Giraud, Martin Juigner, Françoise Debaine, and Cyril Michon. 2021. "Coastal Sand Dunes Monitoring by Low Vegetation Cover Classification and Digital Elevation Model Improvement Using Synchronized Hyperspectral and Full-Waveform LiDAR Remote Sensing" Remote Sensing 13, no. 1: 29. https://doi.org/10.3390/rs13010029
APA StyleFrati, G., Launeau, P., Robin, M., Giraud, M., Juigner, M., Debaine, F., & Michon, C. (2021). Coastal Sand Dunes Monitoring by Low Vegetation Cover Classification and Digital Elevation Model Improvement Using Synchronized Hyperspectral and Full-Waveform LiDAR Remote Sensing. Remote Sensing, 13(1), 29. https://doi.org/10.3390/rs13010029