Identification of microRNAs and Their Expression in Leaf Tissues of Guava (Psidium guajava L.) under Salinity Stress

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
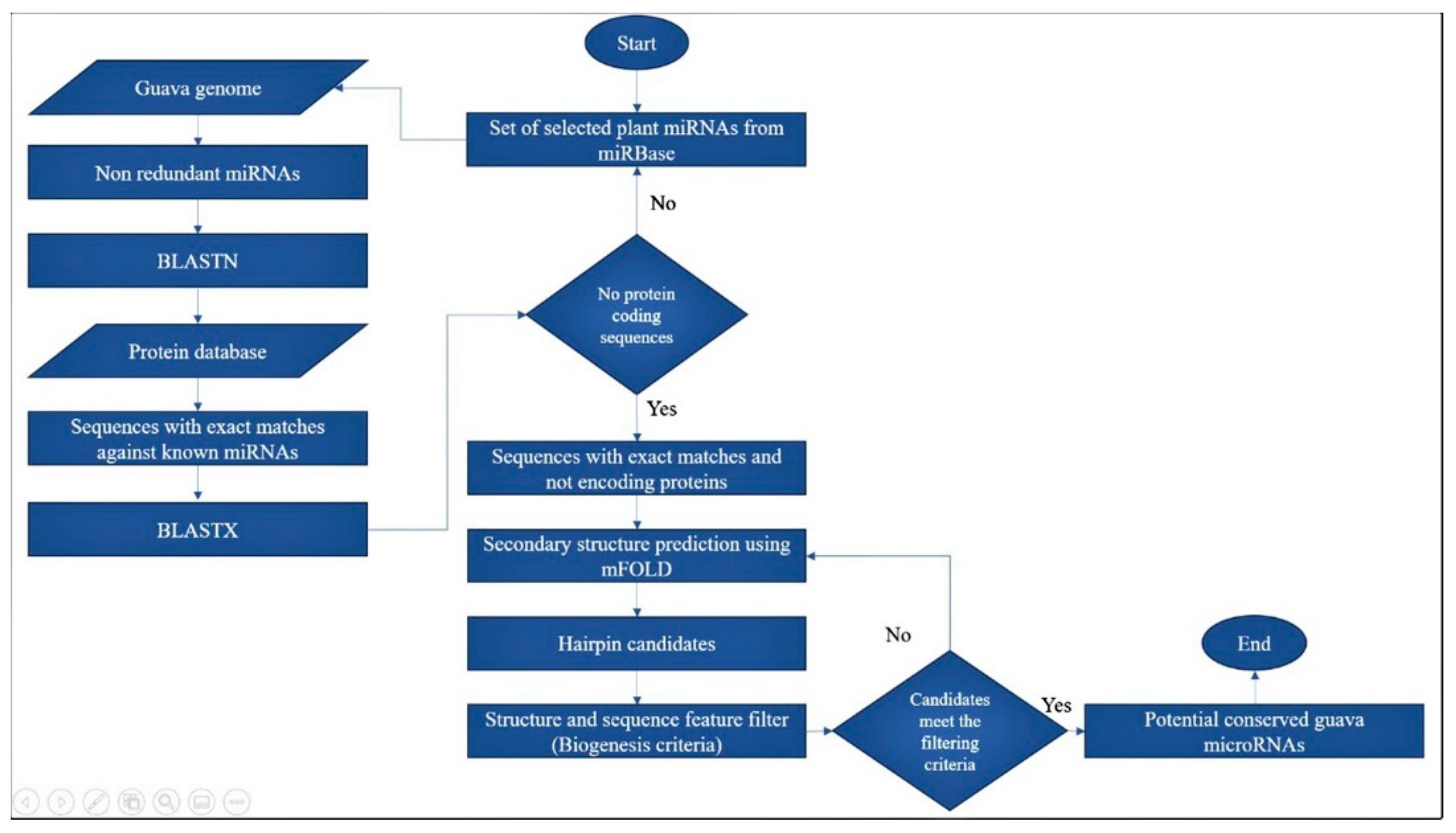
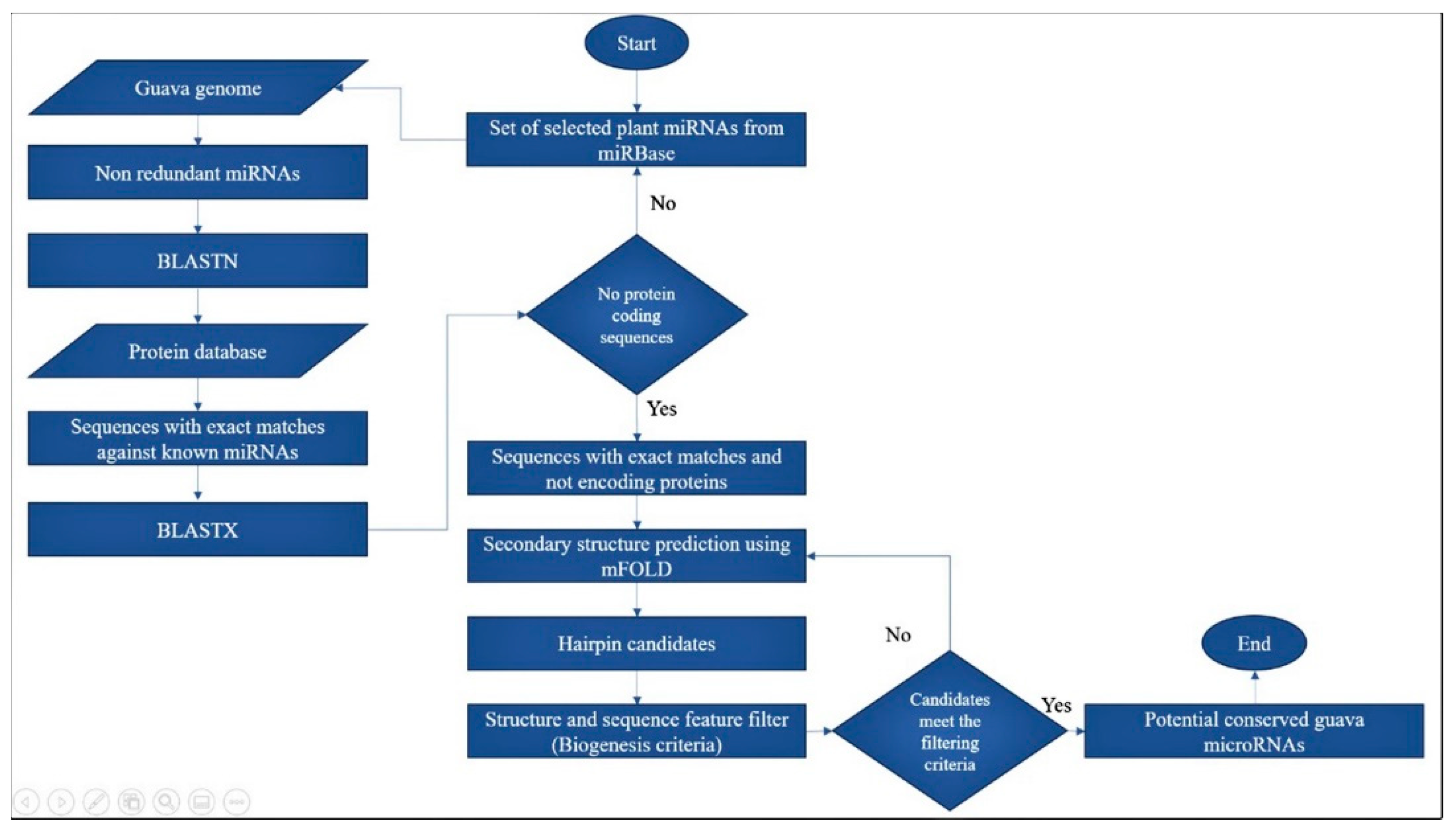
2.1. Computational Prediction of Potential Guava miRNAs and Their Pre-miRNA Candidates
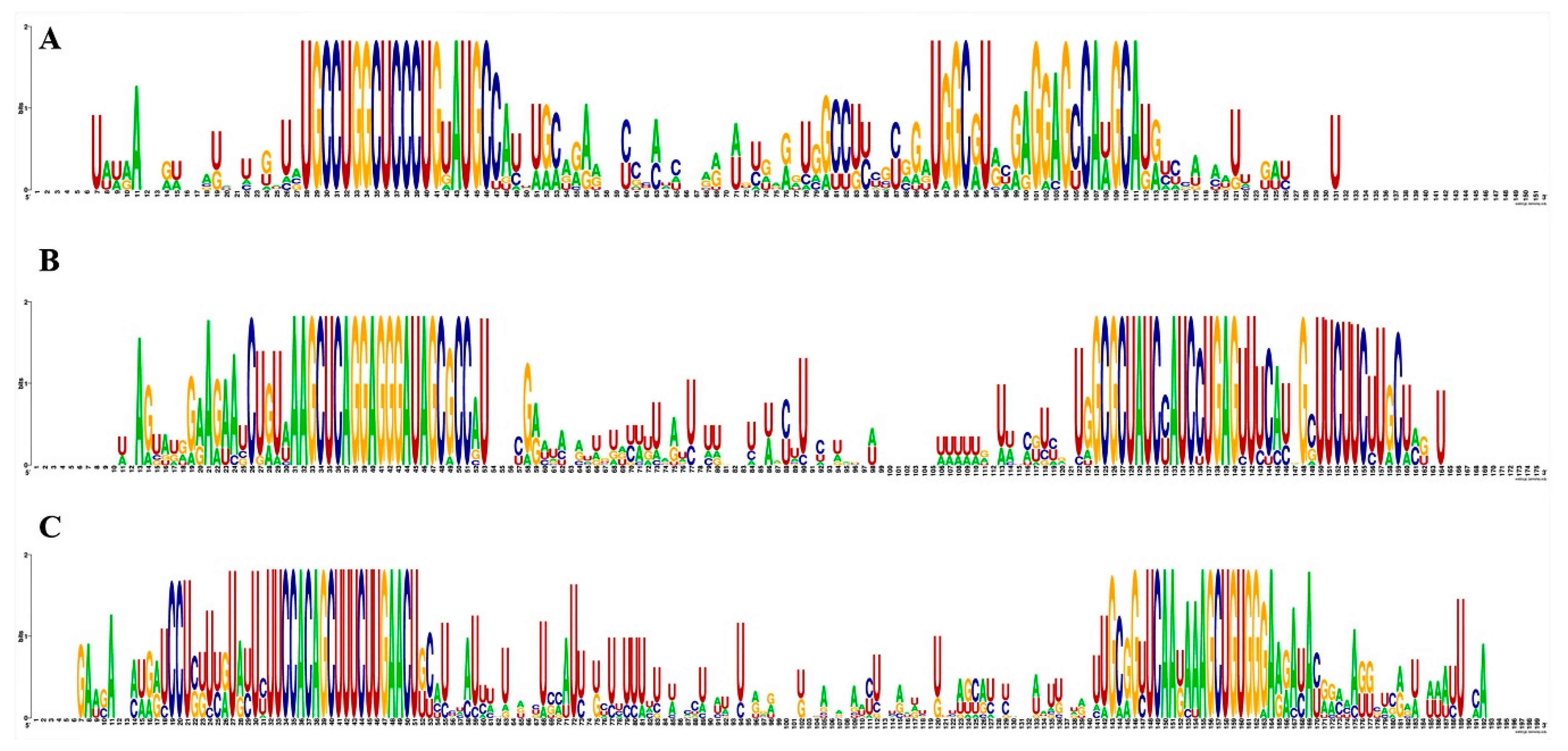
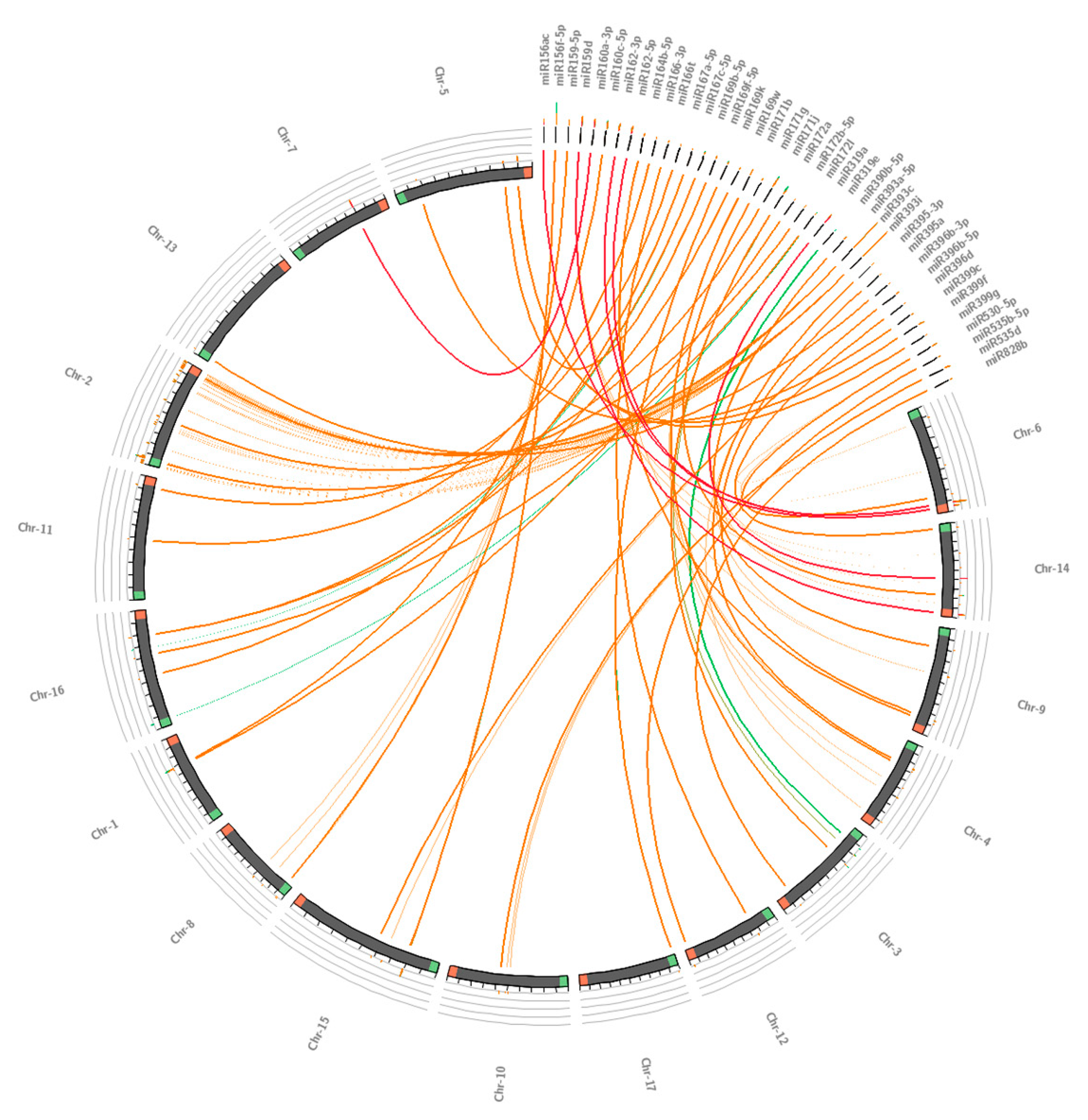
2.2. Phylogenetic and Conservation Analysis of Guava miRNA and Their Pre-miRNAs
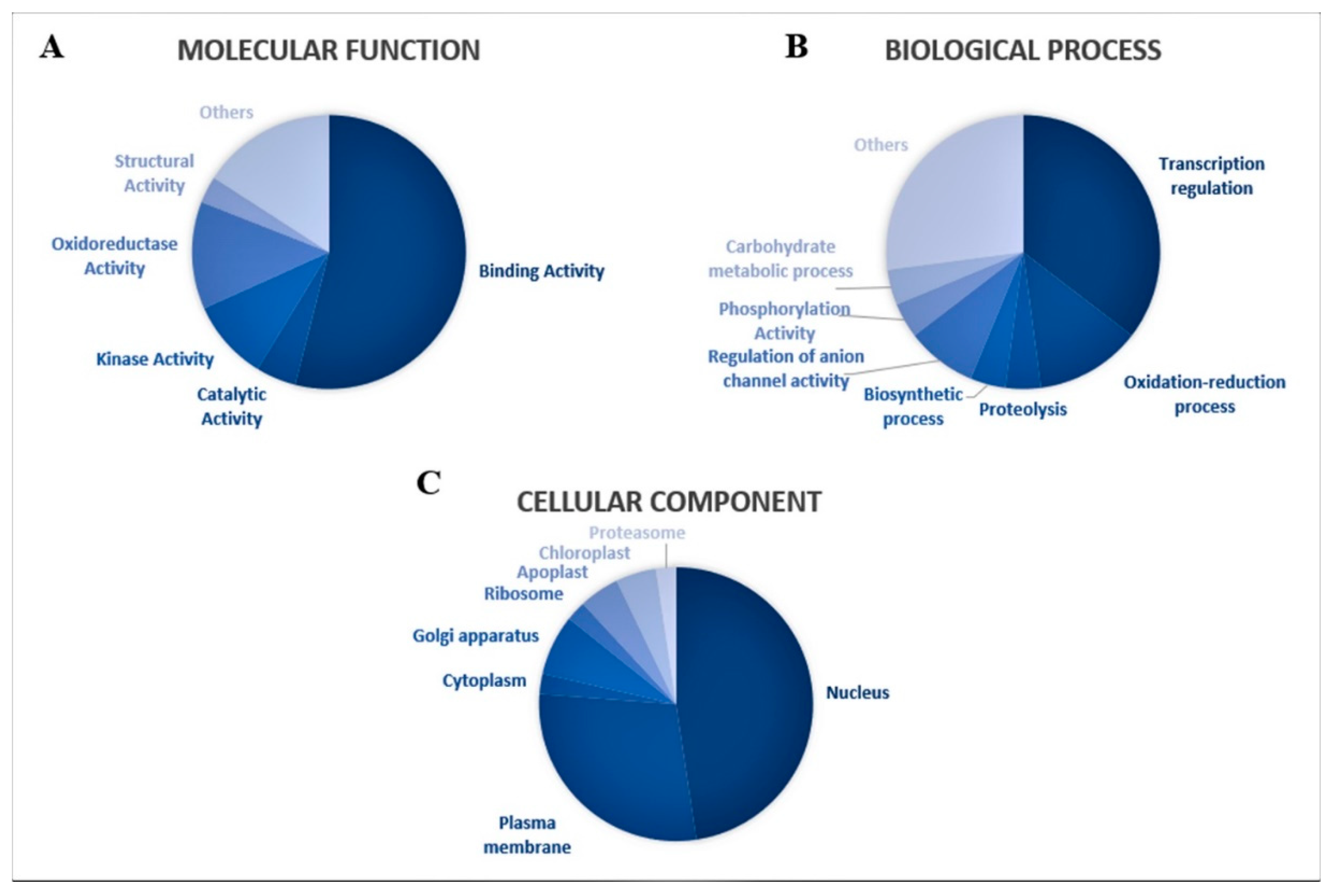
2.3. Target Prediction of Guava miRNAs and Their Functional Annotations
2.4. Plant Materials, Stress Treatment, RNA Extraction, and miRNA Expression Analysis
3. Results
3.1. Characterization of Guava miRNAs and Their Candidate Precursors
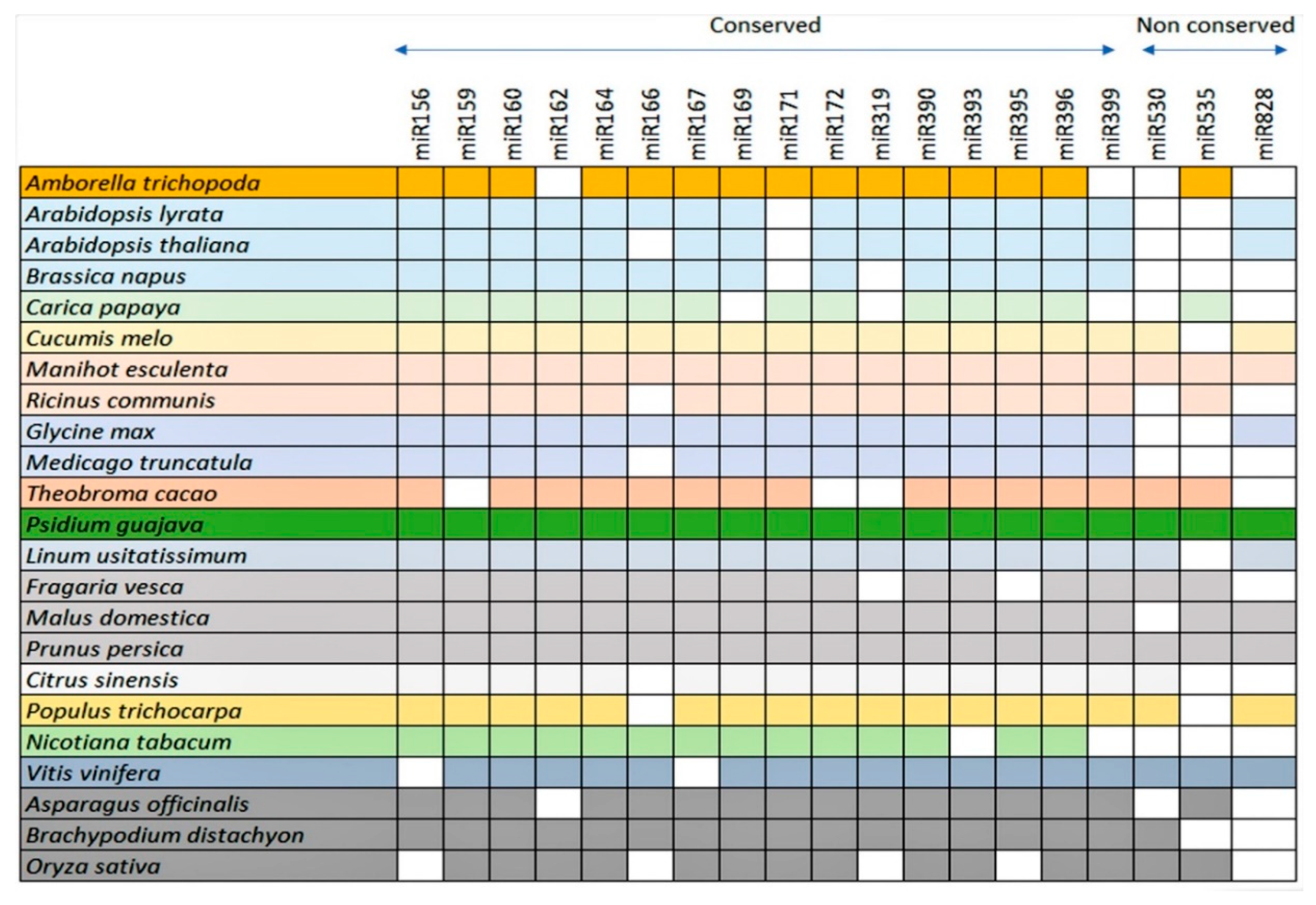
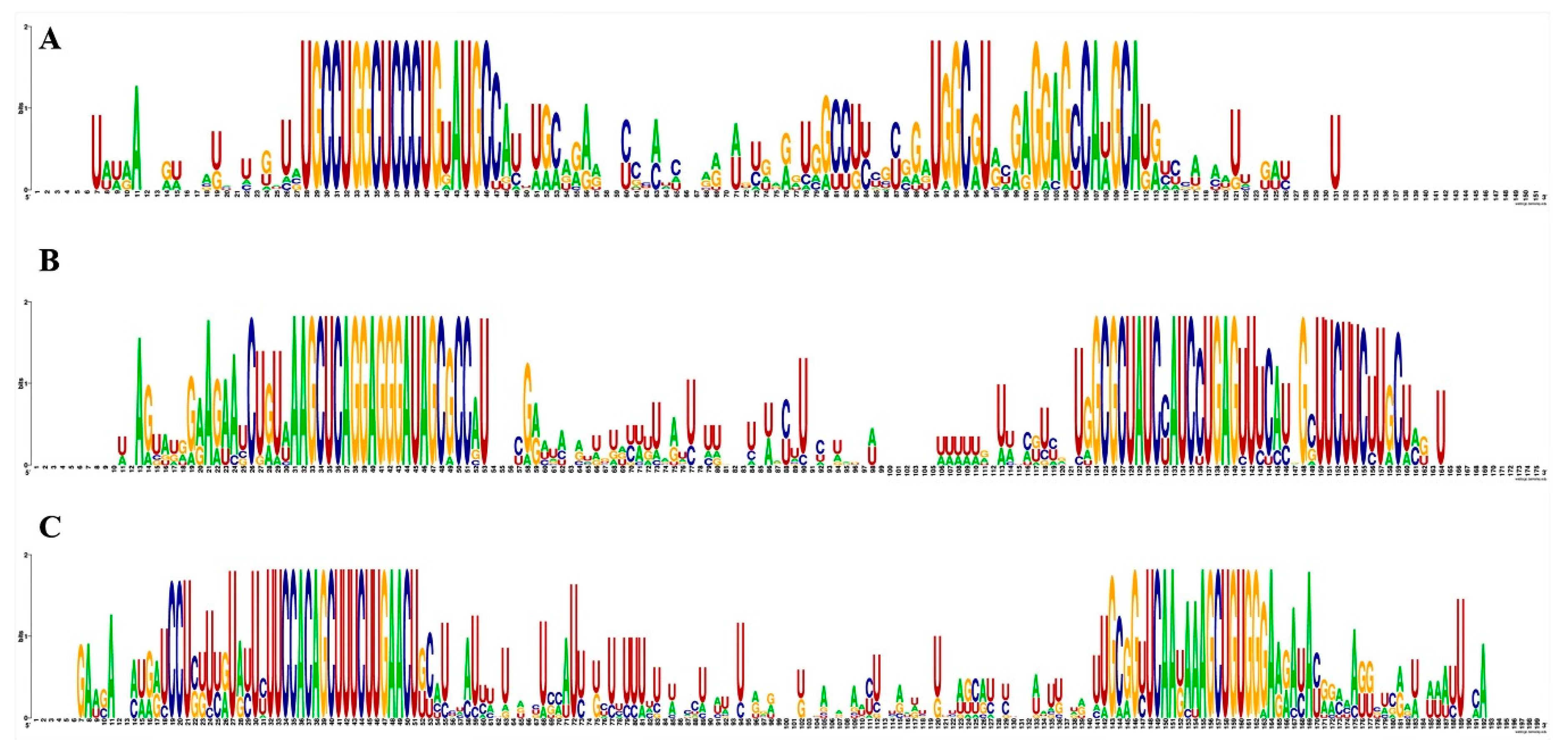
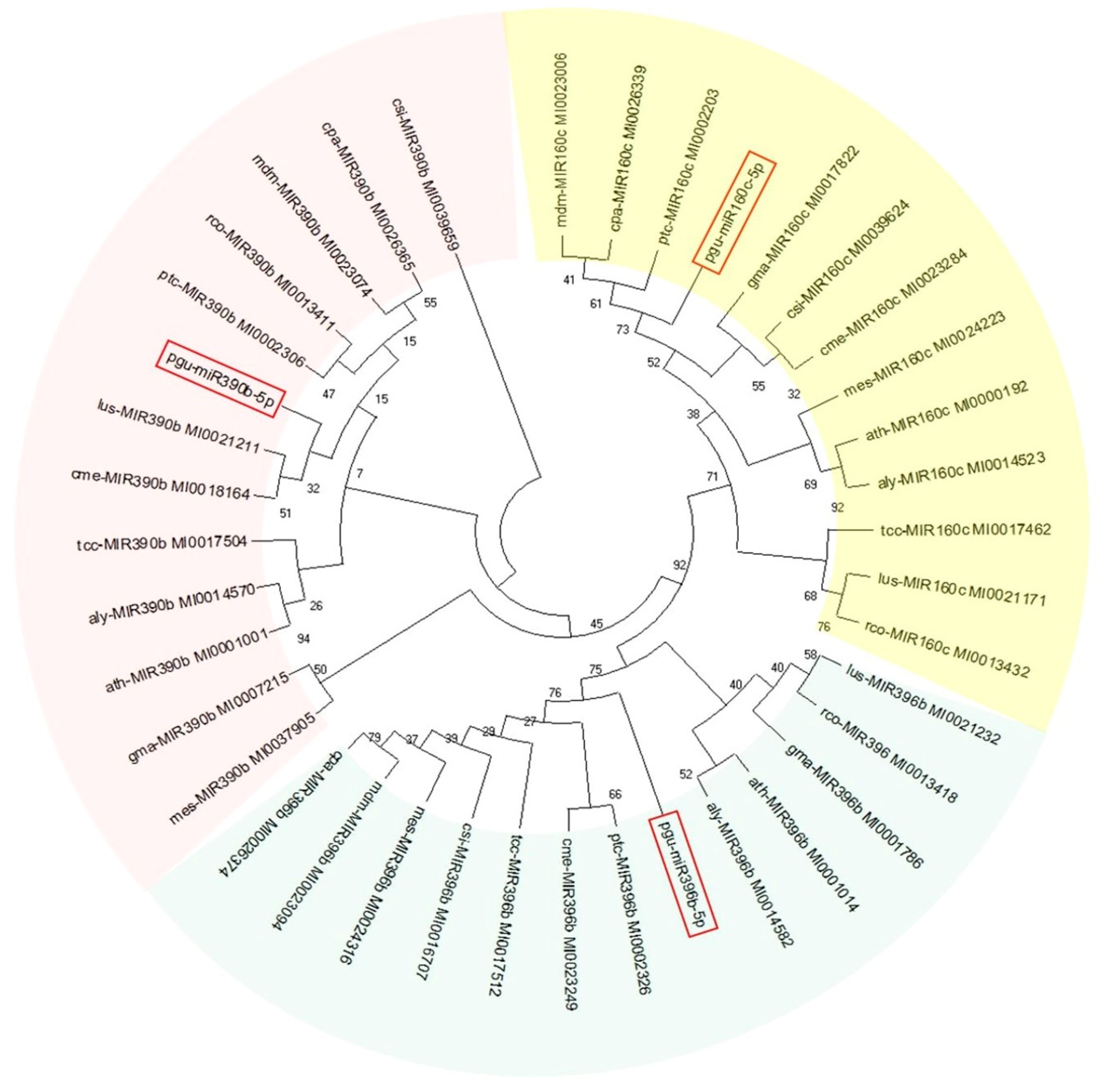
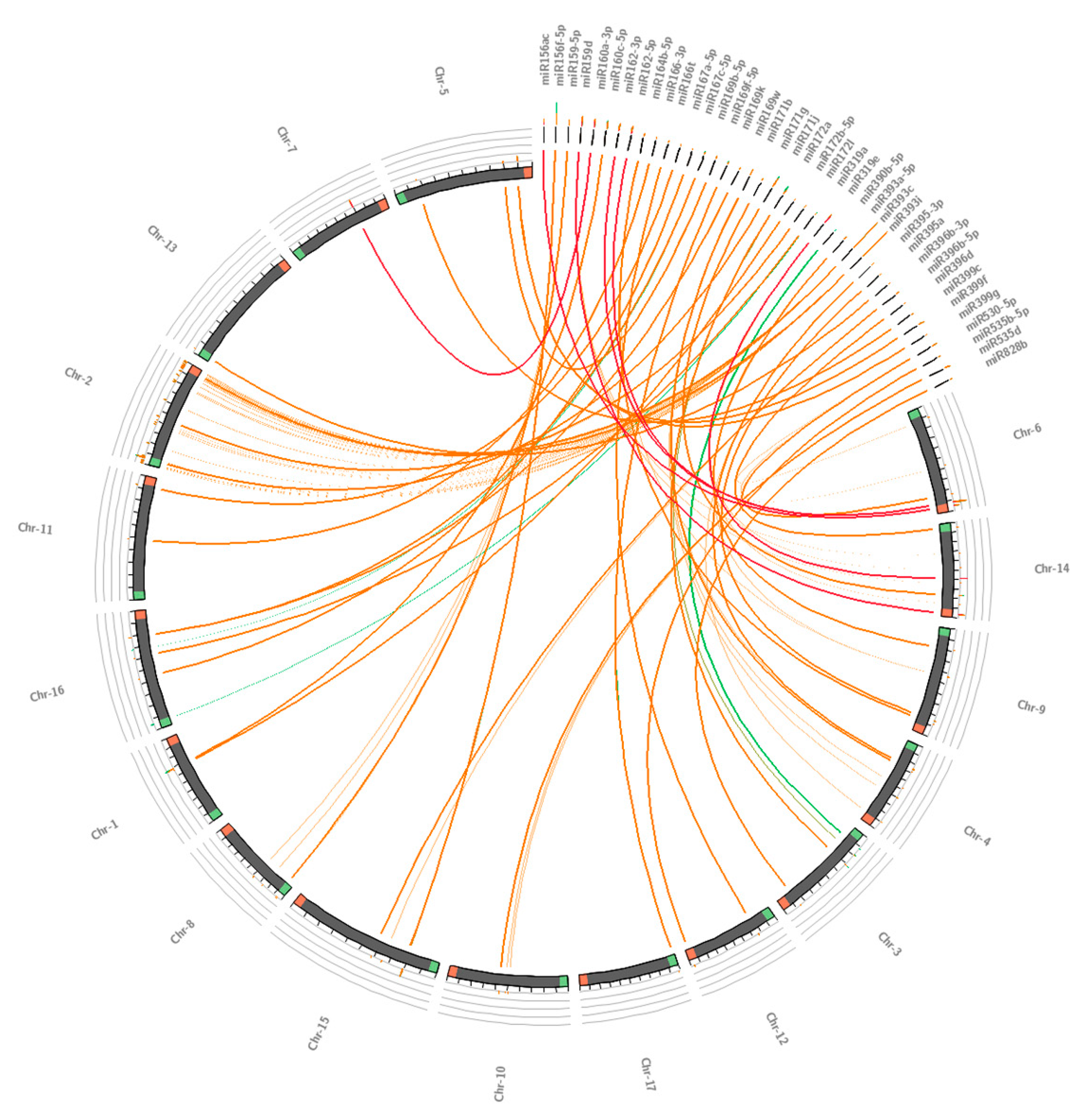
3.2. Conservation Analysis of Guava miRNAs and Their Potential Precursors
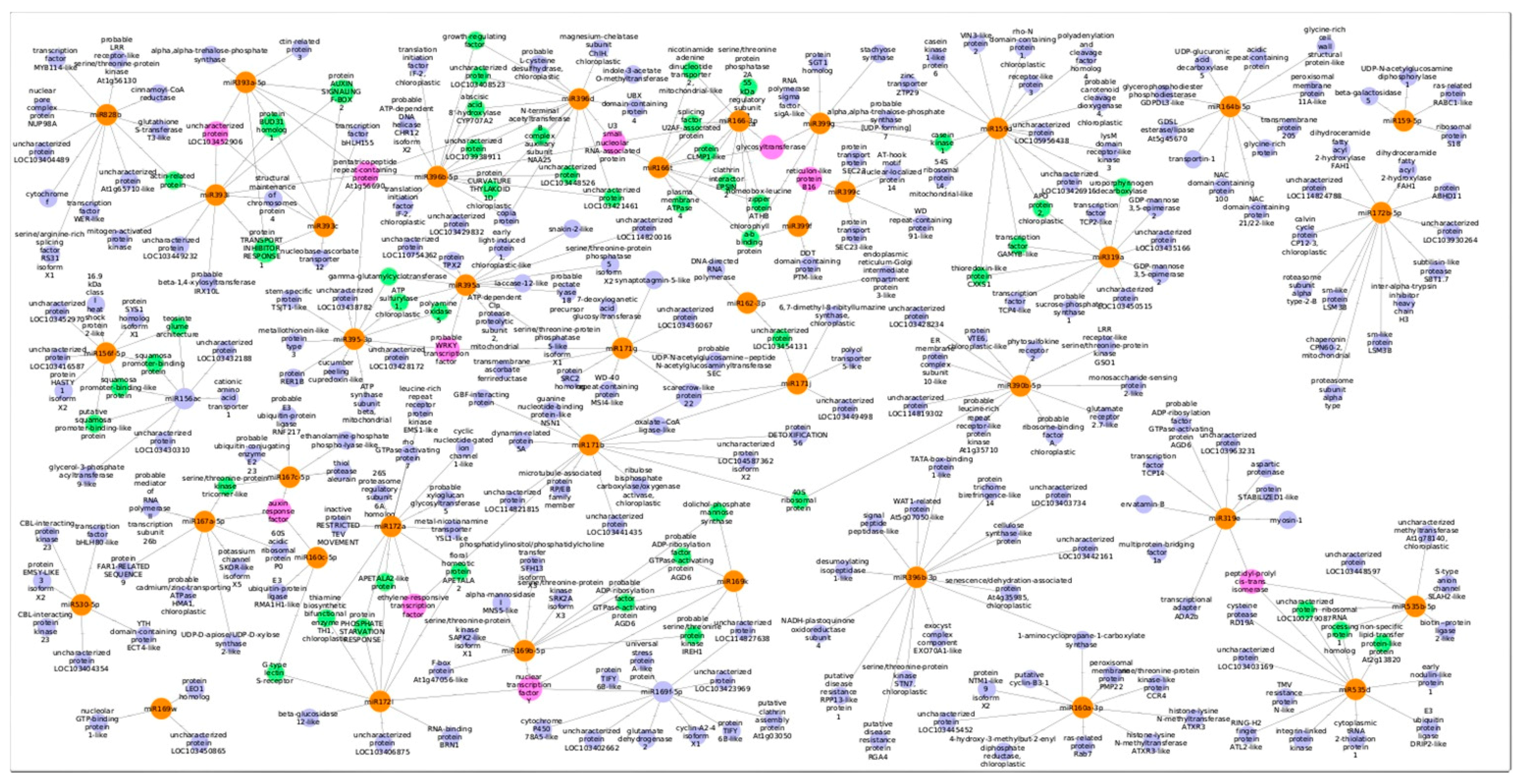
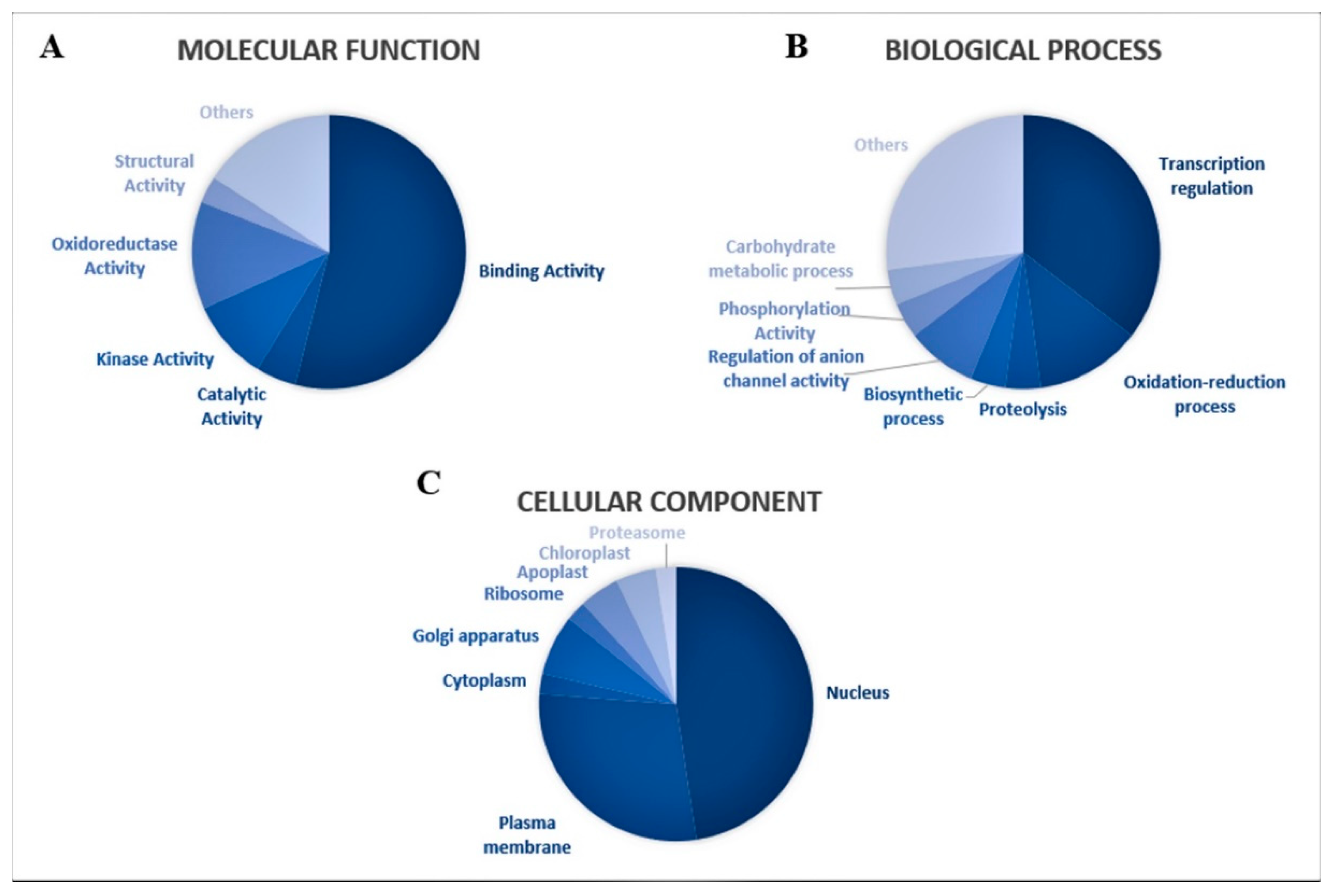
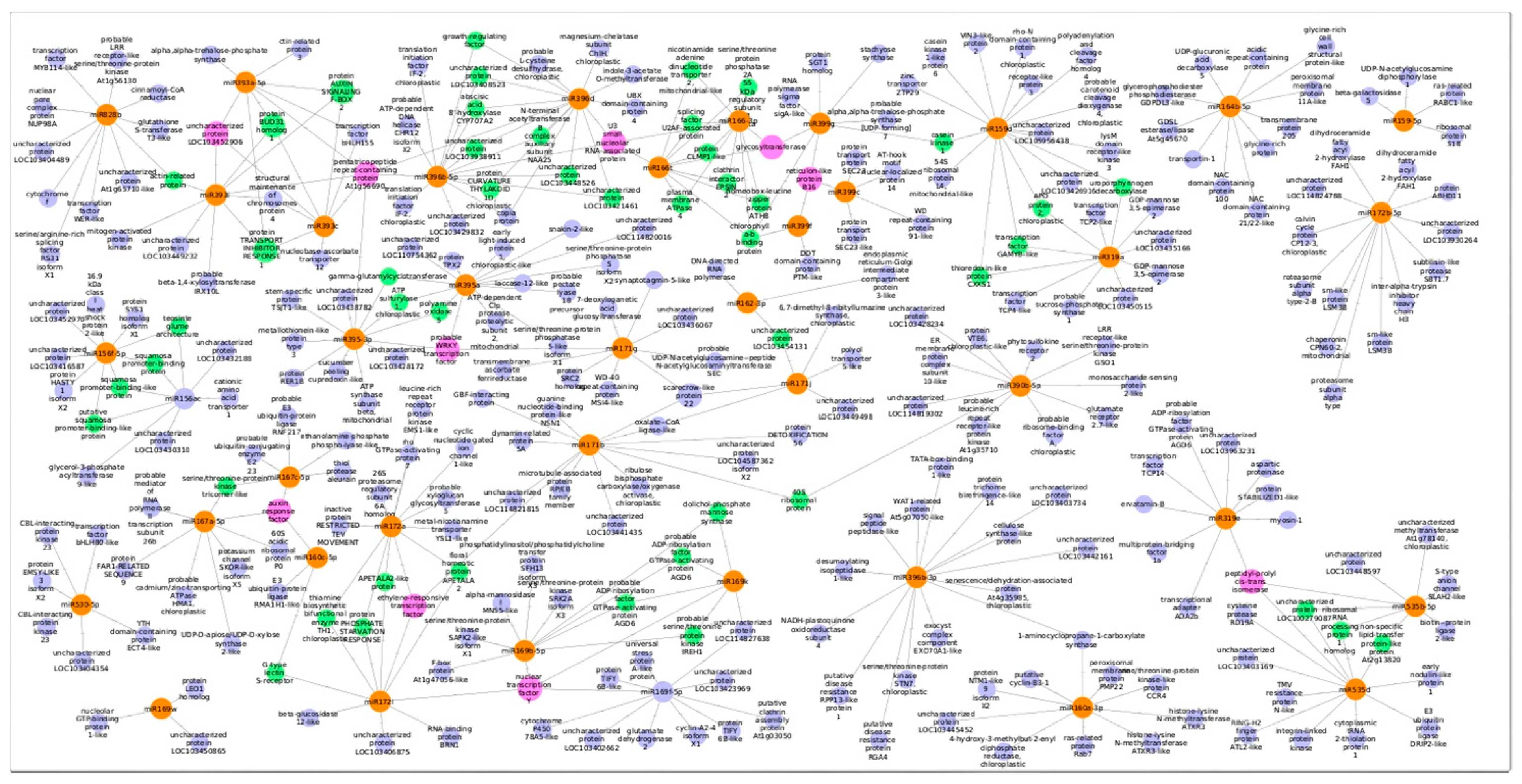
3.3. Predicted Targets for Guava miRNAs and Their Functional Annotations
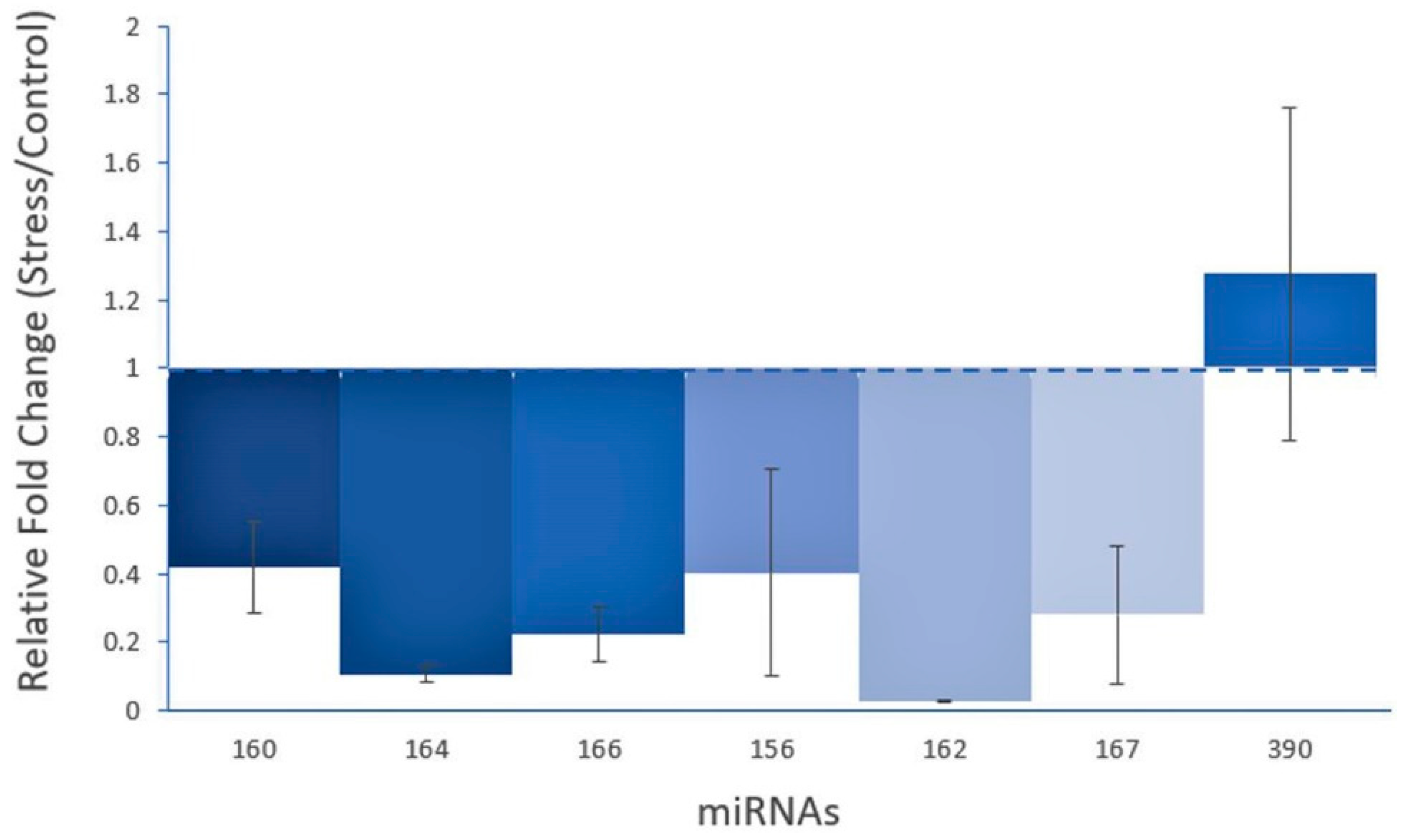
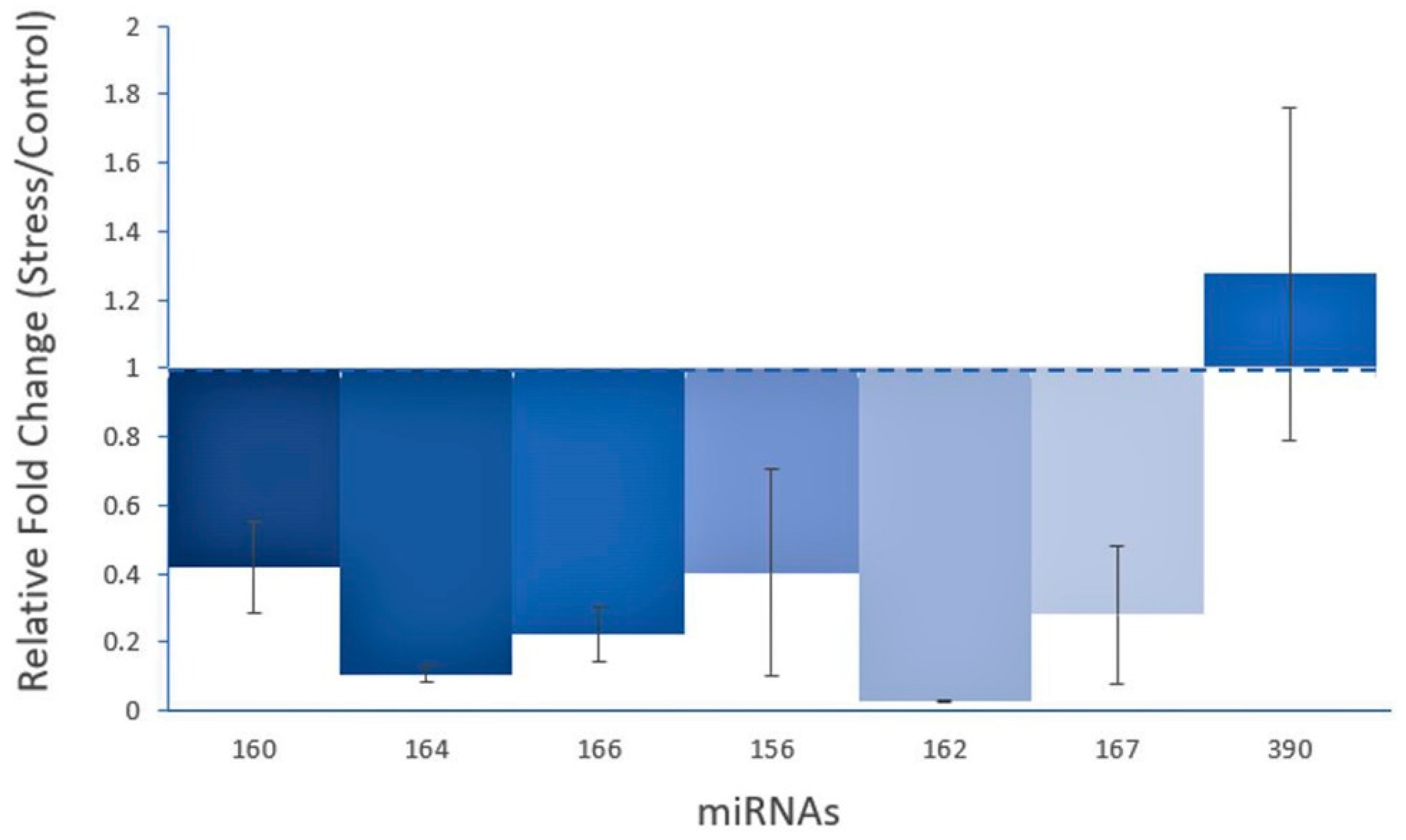
3.4. Expression Analysis of Guava miRNAs under Salinity Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Paul, S.; Kundu, A.; Pal, A. Identification and validation of conserved microRNAs along with their differential expression in roots of Vigna unguiculata grown under salt stress. Plant Cell. Tissue Organ Cult. 2011, 105, 233–242. [Google Scholar] [CrossRef]
- Fang, Z.; Jiang, W.; He, Y.; Ma, D.; Liu, Y.; Wang, S.; Zhang, Y.; Yin, J. Genome-wide identification, structure characterization, and expression profiling of Dof transcription factor gene family in wheat (Triticum aestivum L.). Agronomy 2020, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Cedillo-Jimenez, C.A.; Feregrino-Perez, A.A.; Guevara-González, R.G.; Cruz-Hernández, A. MicroRNA regulation during the tomato fruit development and ripening: A review. Sci. Hortic. 2020, 270, 109435. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Peragine, A.; Mee, Y.P.; Poethig, R.S. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes Dev. 2005, 19, 2164–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidner, C.A.; Martienssen, R.A. The developmental role of microRNA in plants. Curr. Opin. Plant Biol. 2005, 8, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Bejerano, P.I.A.; Maldonado, I.C.; Capote, M.d.D.; Madariaga-Navarrete, A.; Paul, S. Genome-wide computational prediction and experimental validation of quinoa (Chenopodium quinoa) micrornas. Can. J. Plant Sci. 2019, 99, 666–675. [Google Scholar] [CrossRef]
- Paul, S. Identification and characterization of microRNAs and their targets in high-altitude stress-adaptive plant maca (Lepidium meyenii Walp). 3 Biotech 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Kundu, A.; Pal, A. Identification and expression profiling of Vigna mungo microRNAs from leaf small RNA transcriptome by deep sequencing. J. Integr. Plant Biol. 2014, 56, 15–23. [Google Scholar] [CrossRef]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Xie, X.; Li, X.; Tian, Y.; Su, M.; Zhang, J.; Han, X.; Cui, Y.; Bian, S. Identification and characterization of microRNAs and their targets from expression sequence tags of Ribes nigrum. Can. J. Plant Sci. 2016, 96, 995–1001. [Google Scholar] [CrossRef]
- Zakeel, M.C.M.; Safeena, M.I.S.; Komathy, T. In silico identification of microRNAs and their target genes in watermelon (Citrullus lanatus). Sci. Hortic. 2019, 252, 55–60. [Google Scholar] [CrossRef]
- Paul, S.; de la Fuente-Jiménez, J.L.; Manriquez, C.G.; Sharma, A. Identification, characterization and expression analysis of passion fruit (Passiflora edulis) microRNAs. 3 Biotech 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury Paul, S.; Sharma, A.; Mehta, R.; Paul, S. In silico Characterization of microRNAs and Their Target Transcripts from Cranberry (Vaccinium macrocarpon). Cytol. Genet. 2020, 54, 82–90. [Google Scholar] [CrossRef]
- Wang, T.; Lu, J.; Xu, Z.; Yang, W.; Wang, J.; Cheng, T.; Zhang, Q. Selection of suitable reference genes for miRNA expression normalization by qRT-PCR during flower development and different genotypes of Prunus mume. Sci. Hortic. 2014, 169, 130–137. [Google Scholar] [CrossRef]
- Rosas Cárdenas, F.D.F.; Suárez, Y.R.; Rangel, R.M.C.; Garcia, V.L.; Aguilera, K.L.G.; Martínez, N.M.; De Folter, S. Effect of constitutive miR164 expression on plant morphology and fruit development in Arabidopsis and tomato. Agronomy 2017, 7, 48. [Google Scholar] [CrossRef] [Green Version]
- Kamath, J.; Rahul, N.; Ashok Kumar, C.; Lakshmi, S. Psidium guajava L: A review. Int. J. Green Pharm. 2008, 2, 9. [Google Scholar] [CrossRef]
- Gutierrez, L.; Bussell, J.D.; Pǎcurar, D.I.; Schwambach, J.; Pǎcurar, M.; Bellini, C. Phenotypic plasticity of adventitious rooting in arabidopsis is controlled by complex regulation of auxin response factor transcripts and microRNA abundance. Plant Cell 2009, 21, 3119–3132. [Google Scholar] [CrossRef] [Green Version]
- Joseph, B.; Priya, M. Review on nutritional, medicinal and pharmacological properties of guava (Psidium guajava Linn.). Int. J. Pharma Bio Sci. 2011, 2, 53–69. [Google Scholar] [CrossRef] [Green Version]
- Da Silva de Souza, T.; da Silva Ferreira, M.F.; Menini, L.; de Lima Souza, J.R.C.; Parreira, L.A.; Cecon, P.R.; Ferreira, A. Essential oil of Psidium guajava: Influence of genotypes and environment. Sci. Hortic. 2017, 216, 38–44. [Google Scholar] [CrossRef]
- Esfandiari Ghalati, R.; Shamili, M.; Homaei, A. Effect of putrescine on biochemical and physiological characteristics of guava (Psidium guajava L.) seedlings under salt stress. Sci. Hortic. 2020, 261, 108961. [Google Scholar] [CrossRef]
- Abbasi, J.A.; Gandahi, A.W.; Saleem, M.; Bhatti, S.M.; Bughio, Z.R. Growth and Development of Guava (Psidium guajava L.) Seedlings Against Sodium Chloride Salinized Water. Pakistan J. Agric. Eng. Vet. Sci. 2019, 1072, 81–85. [Google Scholar]
- Li, R.; Yuan, S.; He, Y.; Fan, J.; Zhou, Y.; Qiu, T.; Lin, X.; Yao, Y.; Liu, J.; Fu, S.; et al. Genome-wide identification and expression profiling analysis of the galactinol synthase gene family in cassava (Manihot esculenta Crantz). Agronomy 2018, 8, 250. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B. MicroRNA: A new target for improving plant tolerance to abiotic stress. J. Exp. Bot. 2015, 66, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- miRBase. Available online: http:/www.mirbase.org/cgi-bin/browse.pl (accessed on 7 May 2020).
- The UNAFold Web Server. Available online: http://unafold.rna.albany.edu/?q=mfold/RNA-Folding-Form (accessed on 8 May 2020).
- Zhang, B.; Pan, X.; Stellwag, E.J. Identification of soybean microRNAs and their targets. Planta 2008, 229, 161–182. [Google Scholar] [CrossRef]
- Yang, W.; Liu, X.; Zhang, J.; Feng, J.; Li, C.; Chen, J. Prediction and validation of conservative microRNAs of Solanum tuberosum L. Mol. Biol. Rep. 2010, 37, 3081–3087. [Google Scholar] [CrossRef]
- WebLogo. Available online: https://weblogo.berkeley.edu/logo.cgi (accessed on 22 June 2020).
- Darzentas, N. Circoletto: Visualizing sequence similarity with Circos. Bioinformatics 2010, 26, 2620–2621. [Google Scholar] [CrossRef]
- psRNATarget: A Plant Small RNA Target Analysis Server. Available online: http://plantgrn.noble.org/psRNATarget/ (accessed on 24 June 2020).
- Gene Ontology and GO Annotations. Available online: https://www.ebi.ac.uk/QuickGO/ (accessed on 13 July 2020).
- Cytoscape. Available online: https://cytoscape.org/release_notes_3_2_0.html (accessed on 18 August 2020).
- KAAS - KEEG Automatic Annotation Server. Available online: http:/www.genome.jp.tools/kaas/ (accessed on 20 August 2020).
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circ. Calif. Agric. Exp. Stn. 1950, 347, 32. [Google Scholar]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.L.; Tan, Y.C.; Yeoh, K.A.; Ghazali, A.K.; Yee, W.Y.; Hoh, C.C. De novo transcriptome analyses of host-fungal interactions in oil palm (Elaeis guineensis Jacq.). BMC Genom. 2016, 17, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.K.; Namasivayam, P.; Abdullah, J.O.; Ho, C.L. Transcriptome profiling of sulfate deprivation responses in two agarophytes Gracilaria changii and Gracilaria salicornia (Rhodophyta). Sci. Rep. 2017, 7, 46563. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.H.; Spriggs, A.; Matthew, L.; Fan, L.; Kennedy, G.; Gubler, F.; Helliwell, C. A diverse set of microRNAs and microRNA-like small RNAs in developing rice grains. Genome Res. 2008, 18, 1456–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, R.; Hu, Z.; Zhang, H. Identification of MicroRNAs in wild soybean (Glycine soja). J. Integr. Plant Biol. 2009, 51, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Frazier, T.P.; Xie, F.; Freistaedter, A.; Burklew, C.E.; Zhang, B. Identification and characterization of microRNAs and their target genes in tobacco (Nicotiana tabacum). Planta 2010, 232, 1289–1308. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Li, D.; Ohler, U.; Guan, J.; Zhou, S. Genome-wide search for miRNA-target interactions in Arabidopsis thaliana with an integrated approach. BMC Genom. 2012, 13, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Guo, G.; Du, J.; Guo, W.; Peng, H.; Ni, Z.; Sun, Q.; Yao, Y. Whole-genome discovery of miRNAs and their targets in wheat (Triticum aestivum L.). BMC Plant Biol. 2014, 14, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Pan, X.; Cannon, C.H.; Cobb, G.P.; Anderson, T.A. Conservation and divergence of plant microRNA genes. Plant J. 2006, 46, 243–259. [Google Scholar] [CrossRef]
- Barozai, M.Y.K.; Baloch, I.A.; Din, M. Identification of MicroRNAs and their targets in Helianthus. Mol. Biol. Rep. 2012, 39, 2523–2532. [Google Scholar] [CrossRef]
- Unver, T.; Budak, H. Conserved micrornas and their targets in model grass species brachypodium distachyon. Planta 2009, 230, 659–669. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.; Li, D.; Chen, H. Identification and characterization of maize microRNAs involved in the very early stage of seed germination. BMC Genom. 2011, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Neutelings, G.; Fénart, S.; Lucau-Danila, A.; Hawkins, S. Identification and characterization of miRNAs and their potential targets in flax. J. Plant Physiol. 2012, 169, 1754–1766. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Zhu, B.; Luan, F.; Zhu, H.; Shao, Y.; Chen, A.; Lu, C.; Luo, Y. Conserved miRNAs and their targets identified in lettuce (Lactuca) by EST analysis. Gene 2010, 463, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Grande, A.V.; Bujdoso, N.; Saedler, H.; Huijser, P. The microRNA regulated SBP-box genes SPL9 and SPL15 control shoot maturation in Arabidopsis. Plant Mol. Biol. 2008, 67, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, A.C.; Bartel, D.P.; Bartel, B. MicroRNA-directed regulation of Arabidopsis Auxin Response Factor17 is essential for proper development and modulates expression of early auxin response genes. Plant Cell 2005, 17, 1360–1375. [Google Scholar] [CrossRef] [Green Version]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Morishita, T.; Kojima, Y.; Maruta, T.; Nishizawa-Yokoi, A.; Yabuta, Y.; Shigeoka, S. Arabidopsis NAC transcription factor, ANAC078, regulates flavonoid biosynthesis under high-light. Plant Cell Physiol. 2009, 50, 2210–2222. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S.; Liu, Y.; Pan, L.; Hayward, A.; Wang, Y. Identification and characterization of miRNAs in ripening fruit of lycium barbarum L. Using high-throughput sequencing. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Peng, S.; Xian, Z.; Lin, D.; Hu, G.; Yang, L.; Ren, M.; Li, Z. Overexpression of a tomato miR171 target gene SlGRAS24 impacts multiple agronomical traits via regulating gibberellin and auxin homeostasis. Plant Biotechnol. J. 2017, 15, 472–488. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A. Plant Cellular and Molecular Responses to High Salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Bej, S.; Basak, J. MicroRNAs: The Potential Biomarkers in Plant Stress Response. Am. J. Plant Sci. 2014, 5, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Li, S. The Arabidopsis thaliana TCP transcription factors: A broadening horizon beyond development. Plant Signal. Behav. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, M.A.; Islas-Flores, T.; Ullah, H. Signaling through WD-Repeat Proteins in Plants. Front. Plant Sci. 2016, 7, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.H.; Tian, X.; Li, Y.J.; Wu, C.A.; Zheng, C.C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.; Zhang, L.; Wang, H.; Liu, Z.; Zhang, Z.; Zheng, Y. Differential expression of miRNAs in response to salt stress in maize roots. Ann. Bot. 2009, 103, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, B.; Ge, L.; Liang, R.; Li, W.; Ruan, K.; Lin, H.; Jin, Y. Members of miR-169 family are induced by high salinity and transiently inhibit the NF-YA transcription factor. BMC Mol. Biol. 2009, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Bai, X.; Yang, L.; Lv, D.; Pan, X.; Li, Y.; Cai, H.; Ji, W.; Chen, Q.; Zhu, Y. Osa-MIR393: A salinity- and alkaline stress-related microRNA gene. Mol. Biol. Rep. 2011, 38, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Li, Y.; Yu, J.; Liu, Y.; Li, C.; Han, X.; Shen, F. Difference in miRNA expression profiles between two cotton cultivars with distinct salt sensitivity. Mol. Biol. Rep. 2012, 39, 4961–4970. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, A.A.; Reyes, J.L. Post-transcriptional gene regulation of salinity and drought responses by plant microRNAs. Plant Cell Environ. 2010, 33, 481–489. [Google Scholar] [CrossRef]
- Kouhi, F.; Sorkheh, K.; Ercisli, S. MicroRNA expression patterns unveil differential expression of conserved miRNAs and target genes against abiotic stress in safflower. PLoS ONE 2020, 15, e0228850. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.S.; Xie, Q.; Fei, J.F.; Chua, N.H. MicroRNA directs mRNA cleavage of the transcription factor NAC1 to downregulate auxin signals for Arabidopsis lateral root development. Plant Cell 2005, 17, 1376–1386. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Fan, B.; Yu, Z.; Nie, L.; Zhao, Y.; Yu, X.; Sun, F.; Lei, X.; Ma, Y. Functional analysis of three miRNAs in agropyron mongolicum keng under drought stress. Agronomy 2019, 9, 661. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Zhu, P.; Kang, H.; Liu, L.; Cao, Q.; Sun, J.; Dong, T.; Zhu, M.; Li, Z.; Xu, T. High-throughput deep sequencing reveals the important role that microRNAs play in the salt response in sweet potato (Ipomoea batatas L.). BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohli, D.; Joshi, G.; Deokar, A.A.; Bhardwaj, A.R.; Agarwal, M.; Katiyar-Agarwal, S.; Srinivasan, R.; Jain, P.K. Identification and characterization of wilt and salt stress-responsive MicroRNAs in chickpea through high-throughput sequencing. PLoS ONE 2014, 9, e108851. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, D.P. Prediction of plant miRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Miura, E.; Robischon, M.; Martinez, C.; Groover, A. The populus class III HD ZIP transcription factor popcorona affects cell differentiation during secondary growth of woody stems. PLoS ONE 2011, 6, e17458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, P.; Carlsbecker, A.; Etchells, J.P.; Turner, S. Class III HD-ZIPs govern vascular cell fate: An HD view on patterning and differentiation. J. Exp. Bot. 2016, 68, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.G.; Shan, J.X.; Shi, M.; Gao, J.P.; Lin, H.X. The miR156-SPL9-DFR pathway coordinates the relationship between development and abiotic stress tolerance in plants. Plant J. 2014, 80, 1108–1117. [Google Scholar] [CrossRef]
- Arshad, M.; Gruber, M.Y.; Wall, K.; Hannoufa, A. An insight into microRNA156 role in salinity stress responses of alfalfa. Front. Plant Sci. 2017, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Cai, W.J.; Wang, S.; Shan, C.M.; Wang, L.J.; Chena, X.Y. Temporal control of trichome distribution by microRNA156-targeted SPL genes in Arabidopsis thaliana. Plant Cell 2010, 22, 2322–2335. [Google Scholar] [CrossRef] [Green Version]
- Ning, K.; Chen, S.; Huang, H.; Jiang, J.; Yuan, H.; Li, H. Molecular characterization and expression analysis of the SPL gene family with BpSPL9 transgenic lines found to confer tolerance to abiotic stress in Betula platyphylla Suk. Plant Cell. Tissue Organ Cult. 2017, 130, 469–481. [Google Scholar] [CrossRef]
- Bhardwaj, A.R.; Joshi, G.; Pandey, R.; Kukreja, B.; Goel, S.; Jagannath, A.; Kumar, A.; Katiyar-Agarwal, S.; Agarwal, M. A genome-wide perspective of miRNAome in response to high temperature, salinity and drought stresses in Brassica juncea (Czern) L. PLoS ONE 2014, 9, e0125644. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Sun, Z.; Li, J.; Cheng, D.; Luo, C.; Dai, C. Characterization of miRNA160/164 and Their Targets Expression of Beet (Beta vulgaris) Seedlings under the Salt Tolerance. Plant Mol. Biol. Report. 2018, 36, 790–799. [Google Scholar] [CrossRef]
- Jodder, J.; Das, R.; Sarkar, D.; Bhattacharjee, P.; Kundu, P. Distinct transcriptional and processing regulations control miR167a level in tomato during stress. RNA Biol. 2018, 15, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Pegler, J.L.; Nguyen, D.Q.; Grof, C.P.L.; Eamens, A.L. Profiling of the salt stress responsive MicroRNA landscape of C4 genetic model species Setaria viridis (l.) beauv. Agronomy 2020, 10, 837. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, J.; Wang, W.; Xu, L.A. ARF family identification in Tamarix chinensis reveals the salt responsive expression of TcARF6 targeted by miR167. PeerJ 2020, 2020, e8829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Kasschau, K.D.; Carrington, J.C. Negative Feedback Regulation of Dicer-Like1 in Arabidopsis by microRNA-Guided mRNA Degradation. Curr. Biol. 2003, 13, 784–789. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, Q.; Zhang, B. Response of miRNAs and their targets to salt and drought stresses in cotton (Gossypium hirsutum L.). Gene 2013, 530, 26–32. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhou, X.H.; Liu, J. Conserved miRNAs and their response to salt stress in wild eggplant Solanum linnaeanum roots. Int. J. Mol. Sci. 2014, 15, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Wang, Y.; Mou, F.; Tian, Y.; Chen, L.; Zhang, S.; Jiang, Q.; Li, X. Genome-wide small RNA analysis of soybean reveals auxin-responsive microRNAs that are differentially expressed in response to salt stress in root apex. Front. Plant Sci. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Morin, R.D.; Aksay, G.; Dolgosheina, E.; Ebhardt, H.A.; Magrini, V.; Mardis, E.R.; Sahinalp, S.C.; Unrau, P.J. Comparative analysis of the small RNA transcriptomes of Pinus contorta and Oryza sativa. Genome Res. 2008, 18, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Depuydt, S.; Rodriguez-Villalon, A.; Santuari, L.; Wyser-Rmili, C.; Ragni, L.; Hardtke, C.S. Suppression of Arabidopsis protophloem differentiation and root meristem growth by CLE45 requires the receptor-like kinase BAM3. Proc. Natl. Acad. Sci. USA 2013, 110, 7074–7079. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Li, W.; Yang, W.; Qi, L.; Han, S. Identification of microRNAs in Caragana intermedia by high-throughput sequencing and expression analysis of 12 microRNAs and their targets under salt stress. Plant Cell Rep. 2013, 32, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Feng, Z.; Liu, X.; Bian, L.; Xie, H.; Zhang, C.; Mysore, K.S.; Liang, J. MiR393 and miR390 synergistically regulate lateral root growth in rice under different conditions. BMC Plant Biol. 2018, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Trumbo, J.L.; Zhang, B.; Stewart, C.N. Manipulating microRNAs for improved biomass and biofuels from plant feedstocks. Plant Biotechnol. J. 2015, 13, 337–354. [Google Scholar] [CrossRef] [Green Version]
- Zafar, S.; Ashraf, M.Y.; Anwar, S.; Ali, Q.; Noman, A. Yield enhancement in wheat by soil and foliar fertilization of K and Zn under saline environment. Soil Environ. 2016, 35, 46–55. [Google Scholar]
- Noman, A.; Fahad, S.; Aqeel, M.; Ali, U.; Amanullah; Anwar, S.; Baloch, S.K.; Zainab, M. miRNAs: Major modulators for crop growth and development under abiotic stresses. Biotechnol. Lett. 2017, 39, 685–700. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, Q. MicroRNA-based biotechnology for plant improvement. J. Cell. Physiol. 2015, 230, 1–15. [Google Scholar] [CrossRef]
- Shriram, V.; Kumar, V.; Devarumath, R.M.; Khare, T.S.; Wani, S.H. Micrornas as potential targets for abiotic stress tolerance in plants. Front. Plant Sci. 2016, 7, 1–18. [Google Scholar] [CrossRef]
- Zhou, M.; Li, D.; Li, Z.; Hu, Q.; Yang, C.; Zhu, L.; Luo, H. Constitutive expression of a miR319 gene alters plant development and enhances salt and drought tolerance in transgenic creeping bentgrass. Plant Physiol. 2013, 161, 1375–1391. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Li, Z.; Li, D.; Yuan, N.; Hu, Q.; Luo, H. Constitutive expression of rice microRNA528 alters plant development and enhances tolerance to salinity stress and nitrogen starvation in creeping bentgrass. Plant Physiol. 2015, 169, 576–593. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Yuan, S.; Zhou, M.; Yuan, N.; Li, Z.; Hu, Q.; Bethea, F.G.; Liu, H.; Li, S.; Luo, H. Transgenic creeping bentgrass overexpressing Osa-miR393a exhibits altered plant development and improved multiple stress tolerance. Plant Biotechnol. J. 2019, 17, 233–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahito, Z.A.; Wang, L.; Sun, Z.; Yan, Q.; Zhang, X.; Jiang, Q.; Ullah, I.; Tong, Y.; Li, X. The miR172c-NNC1 module modulates root plastic development in response to salt in soybean. BMC Plant Biol. 2017, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identified miRNAs | LM * (nt) | Query miRNAs | miRNA Sequences | Accession | Strand | Location | LP * (nt) | MFEs (ΔG) | MFEI |
|---|---|---|---|---|---|---|---|---|---|
| pgu-miR156ac | 21 | mdm-miR156ac | UUGACAGAAGAUAGAGAGCAC | MI0022999 | +/− | 5′ | 86 | −47 | 1.18 |
| pgu-miR156f-5p | 20 | ath-miR156f-5p | UGACAGAAGAGAGUGAGCAC | MI0000183 | +/+ | 5′ | 86 | −50.8 | 1.1 |
| pgu-miR159-5p | 21 | eun-miR159-5p | AGCUGCUGGUCUAUGGAUCCC | MI0033013 | +/− | 5′ | 134 | −48.5 | 0.77 |
| pgu-miR159d | 21 | mdm-miR159d | UUUGGAUUGAAGGGAGCUCUA | MI0035594 | +/− | 3′ | 215 | −95.2 | 0.93 |
| pgu-miR160a-3p | 21 | ath-miR160a-3p | GCGUAUGAGGAGCCAUGCAUA | MI0000190 | +/+ | 3′ | 91 | −49.7 | 0.97 |
| pgu-miR160c-5p | 21 | ath-miR160c-5p | UGCCUGGCUCCCUGUAUGCCA | MI0000192 | +/+ | 5′ | 86 | −51.4 | 1.01 |
| pgu-miR162-3p | 21 | eun-miR162-3p | UCGAUAAACCUCUGCAUCCAG | MI0033015 | +/− | 3′ | 111 | −38.4 | 0.7 |
| pgu-miR164b-5p | 21 | ath-miR164b-5p | UGGAGAAGCAGGGCACGUGCA | MI0000198 | +/+ | 5′ | 91 | −43.4 | 0.9 |
| pgu-miR166-3p | 21 | eun-miR166-3p | UCGGACCAGGCUUCAUUCCCC | MI0033016 | +/− | 3′ | 123 | −51.3 | 0.99 |
| pgu-miR166t | 20 | gma-miR166t | UCGGACCAGGCUUCAUUCCC | MI0021696 | +/+ | 5′ | 111 | −47.7 | 0.75 |
| pgu-miR167a-5p | 21 | ath-miR167a-5p | UGAAGCUGCCAGCAUGAUCUA | MI0000208 | +/+ | 5′ | 111 | −50.4 | 1.01 |
| pgu-miR167c-5p | 22 | eun-miR167c-5p | UGAAGCUGCCAGCGUGAUCUCA | MI0033019 | +/− | 5′ | 86 | −33.1 | 0.74 |
| pgu-miR169b-5p | 21 | ath-miR169b-5p | CAGCCAAGGAUGACUUGCCGG | MI0000976 | +/+ | 5′ | 116 | −37.6 | 0.7 |
| pgu-miR169f-5p | 21 | ath-miR169f-5p | UGAGCCAAGGAUGACUUGCCG | MI0000980 | +/+ | 5′ | 96 | −41.5 | 0.86 |
| pgu-miR169k | 21 | mdm-miR169k | UAGCCAAGGAUGACUUGCCUG | MI0035667 | +/+ | 5′ | 103 | −45.5 | 1.08 |
| pgu-miR169w | 21 | gma-miR169w | CAAGGAUGACUUGCCGGCAUU | MI0033468 | +/+ | 5′ | 106 | −38.2 | 0.8 |
| pgu-miR171b | 21 | mdm-miR171b | UUGAGCCGCGUCAAUAUCUCC | MI0023043 | +/+ | 3′ | 116 | −46.1 | 0.85 |
| pgu-miR171g | 21 | mdm-miR171g | UGAUUGAGCCGUGCCAAUAUC | MI0023048 | +/+ | 3′ | 88 | −36.7 | 1.05 |
| pgu-miR171j | 21 | mdm-miR171j | UUGAGCCGCGCCAAUAUCACU | MI0023051 | +/+ | 3′ | 106 | −41.8 | 0.91 |
| pgu-miR172a | 21 | ath-miR172a | AGAAUCUUGAUGAUGCUGCAU | MI0000215 | +/+ | 3′ | 111 | −48.4 | 0.99 |
| pgu-miR172b-5p | 21 | eun-miR172b-5p | GCAGCAUCAUCAAGAUUCACA | MI0033022 | +/− | 5′ | 111 | −48.4 | 0.99 |
| pgu-miR172l | 21 | mdm-miR172l | GGAAUCUUGAUGAUGCUGCAG | MI0023067 | +/− | 3′ | 176 | −65.1 | 0.97 |
| pgu-miR319a | 21 | ath-miR319a | UUGGACUGAAGGGAGCUCCCU | MI0000544 | +/+ | 3′ | 201 | −92.4 | 0.9 |
| pgu-miR319e | 20 | mdm-miR319e | GAGCUUUCUUCAGUCCACUC | MI0035621 | +/+ | 5′ | 176 | −85.7 | 0.88 |
| pgu-miR390b-5p | 21 | ath-miR390b-5p | AAGCUCAGGAGGGAUAGCGCC | MI0001001 | +/+ | 5′ | 84 | −51.3 | 1.09 |
| pgu-miR393a-5p | 22 | ath-miR393a-5p | UCCAAAGGGAUCGCAUUGAUCC | MI0001003 | +/+ | 5′ | 96 | −40.7 | 0.93 |
| pgu-miR393c | 22 | mdm-miR393c | UCCAAAGGGAUCGCAUUGAUCU | MI0023081 | +/+ | 5′ | 111 | −44.4 | 0.91 |
| pgu-miR393i | 22 | gma-miR393i | UUCCAAAGGGAUCGCAUUGAUC | MI0021710 | +/+ | 5′ | 126 | −56.4 | 0.97 |
| pgu-miR395-3p | 21 | eun-miR395-3p | AUGAAGUGUUUGGGGGAACUC | MI0033024 | +/+ | 3′ | 72 | −47.5 | 1.36 |
| pgu-miR395a | 21 | ath-miR395a | CUGAAGUGUUUGGGGGAACUC | MI0001007 | +/+ | 3′ | 106 | −37.2 | 0.89 |
| pgu-miR396b-3p | 21 | ath-miR396b-3p | GCUCAAGAAAGCUGUGGGAAA | MI0001014 | +/+ | 3′ | 126 | −36.1 | 0.82 |
| pgu-miR396b-5p | 21 | eun-miR396b-5p | UUCCACAGCUUUCUUGAACUG | MI0033026 | +/+ | 5′ | 118 | −52.4 | 1.01 |
| pgu-miR396d | 21 | mdm-miR396d | UUCCACAGCUUUCUUGAACUU | MI0023096 | +/+ | 5′ | 96 | −50.6 | 0.9 |
| pgu-miR399c | 21 | mdm-miR399c | UGCCAAAGGAGAAUUGCCCUG | MI0023107 | +/− | 3′ | 96 | −50.4 | 1.12 |
| pgu-miR399f | 21 | ath-miR399f | UGCCAAAGGAGAUUUGCCCGG | MI0001025 | +/+ | 3′ | 111 | −50 | 0.98 |
| pgu-miR399g | 21 | mdm-miR399g | UGCCAAAGGAGAUUUGCUCGG | MI0023111 | +/+ | 3′ | 81 | −36.5 | 1.11 |
| pgu-miR530-5p | 21 | eun-miR530-5p | UCUGCAUUUGCACCUGCACCU | MI0033032 | +/+ | 5′ | 161 | −73.6 | 0.89 |
| pgu-miR535b-5p | 21 | eun-miR535b-5p | UGACAACGAGAGAGAGCACGC | MI0033034 | +/+ | 5′ | 81 | −41.6 | 1.01 |
| pgu-miR535d | 21 | mdm-miR535d | UGACGACGAGAGAGAGCACGC | MI0023131 | +/+ | 5′ | 101 | −49 | 1.02 |
| pgu-miR828b | 22 | mdm-miR828b | UCUUGCUCAAAUGAGUAUUCCA | MI0023134 | +/+ | 5′ | 99 | −33.8 | 0.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Ruiz-Manriquez, L.M.; Serrano-Cano, F.I.; Reyes-Pérez, P.R.; Tovar Alfaro, C.K.; Barrón Andrade, Y.E.; Hernández Aros, A.K.; Srivastava, A.; Paul, S. Identification of microRNAs and Their Expression in Leaf Tissues of Guava (Psidium guajava L.) under Salinity Stress. Agronomy 2020, 10, 1920. https://doi.org/10.3390/agronomy10121920
Sharma A, Ruiz-Manriquez LM, Serrano-Cano FI, Reyes-Pérez PR, Tovar Alfaro CK, Barrón Andrade YE, Hernández Aros AK, Srivastava A, Paul S. Identification of microRNAs and Their Expression in Leaf Tissues of Guava (Psidium guajava L.) under Salinity Stress. Agronomy. 2020; 10(12):1920. https://doi.org/10.3390/agronomy10121920
Chicago/Turabian StyleSharma, Ashutosh, Luis M. Ruiz-Manriquez, Francisco I. Serrano-Cano, Paula Roxana Reyes-Pérez, Cynthia Karina Tovar Alfaro, Yulissa Esmeralda Barrón Andrade, Ana Karen Hernández Aros, Aashish Srivastava, and Sujay Paul. 2020. "Identification of microRNAs and Their Expression in Leaf Tissues of Guava (Psidium guajava L.) under Salinity Stress" Agronomy 10, no. 12: 1920. https://doi.org/10.3390/agronomy10121920
APA StyleSharma, A., Ruiz-Manriquez, L. M., Serrano-Cano, F. I., Reyes-Pérez, P. R., Tovar Alfaro, C. K., Barrón Andrade, Y. E., Hernández Aros, A. K., Srivastava, A., & Paul, S. (2020). Identification of microRNAs and Their Expression in Leaf Tissues of Guava (Psidium guajava L.) under Salinity Stress. Agronomy, 10(12), 1920. https://doi.org/10.3390/agronomy10121920