High-Resolution Crystal Structure of Chloroplastic Ribose-5-Phosphate Isomerase from Chlamydomonas reinhardtii—An Enzyme Involved in the Photosynthetic Calvin-Benson Cycle



Abstract
1. Introduction
2. Results
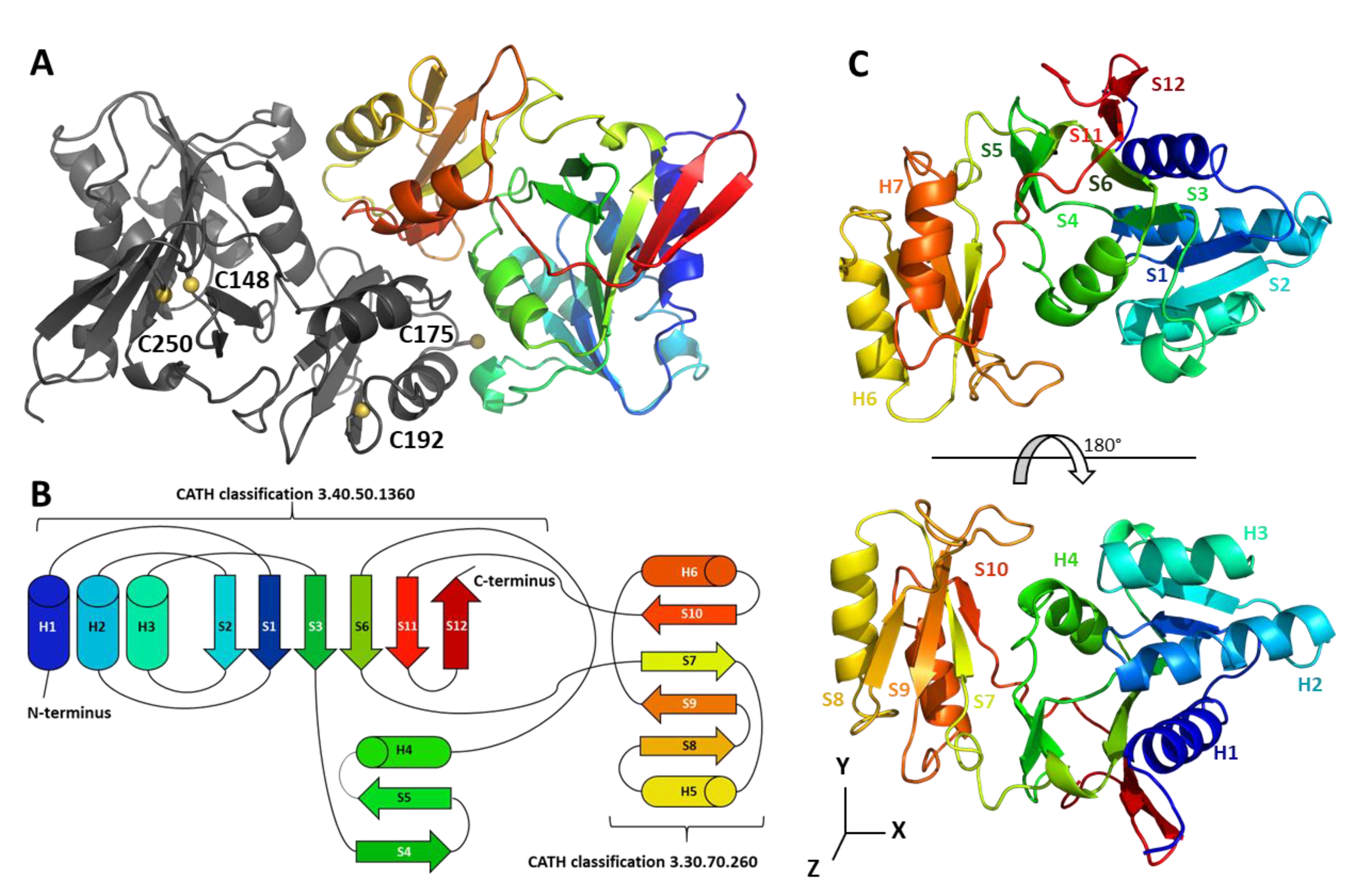
2.1. CrRPI1 Structural Folds
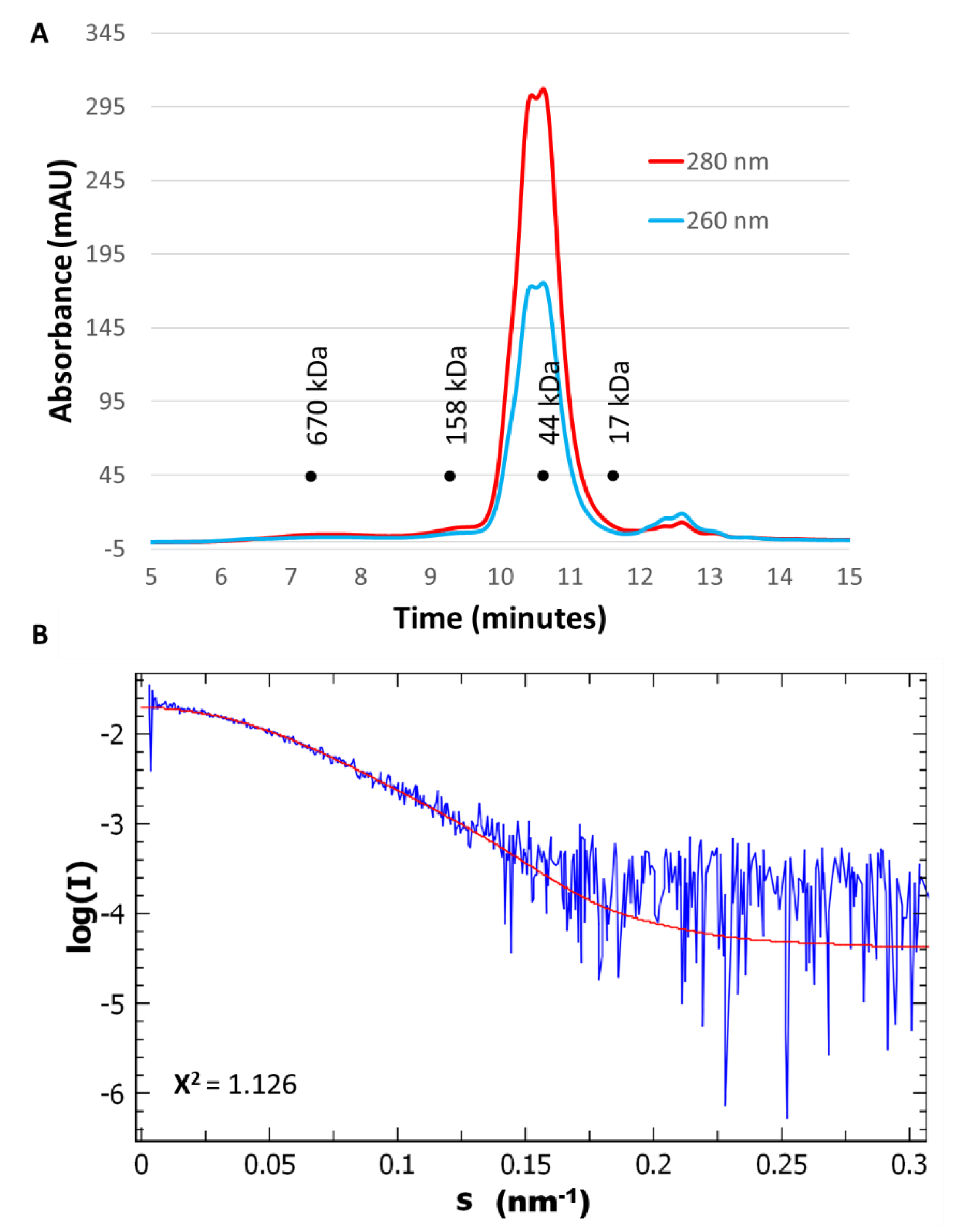
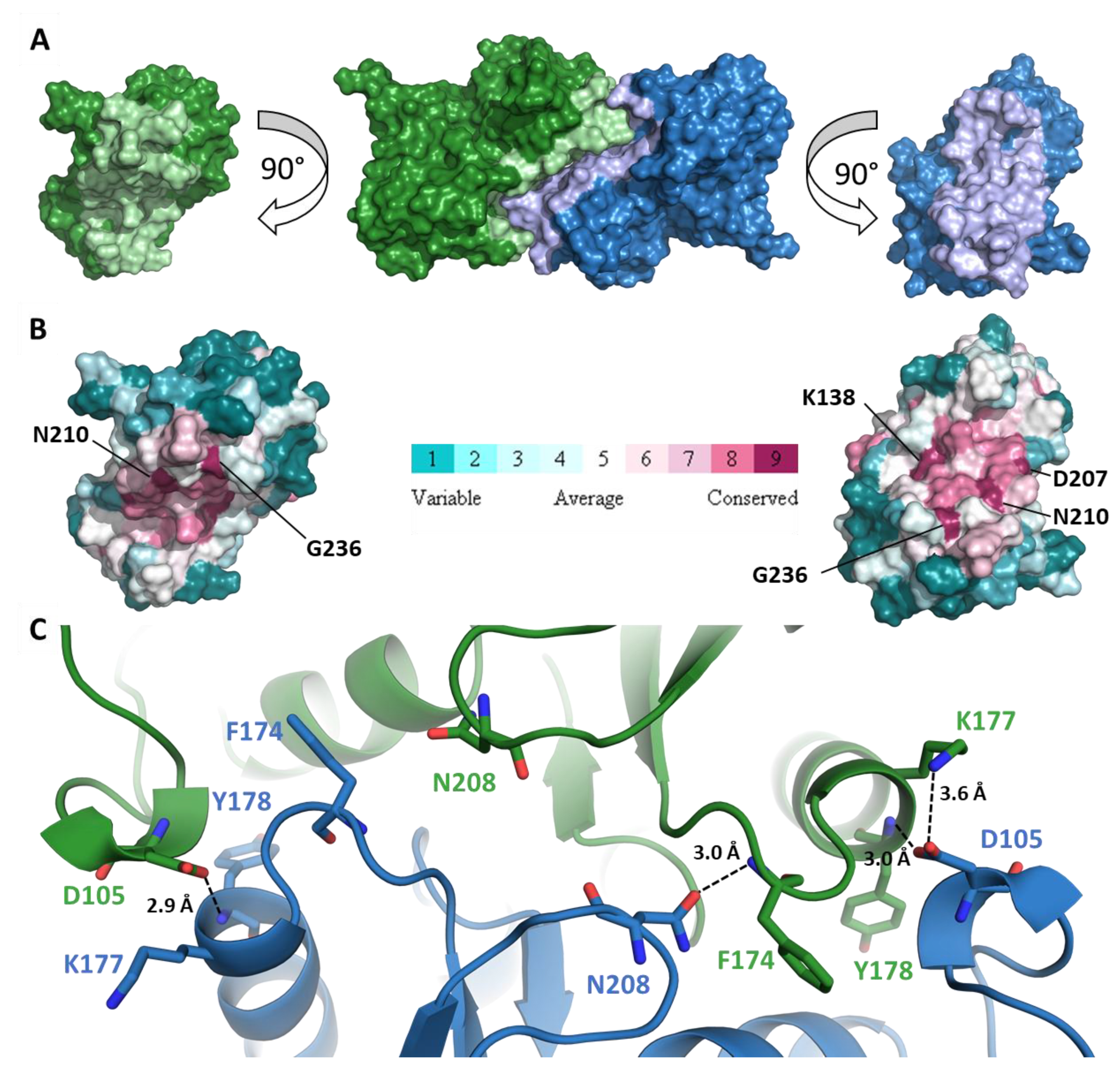
2.2. CrRPI1 is a Homodimer
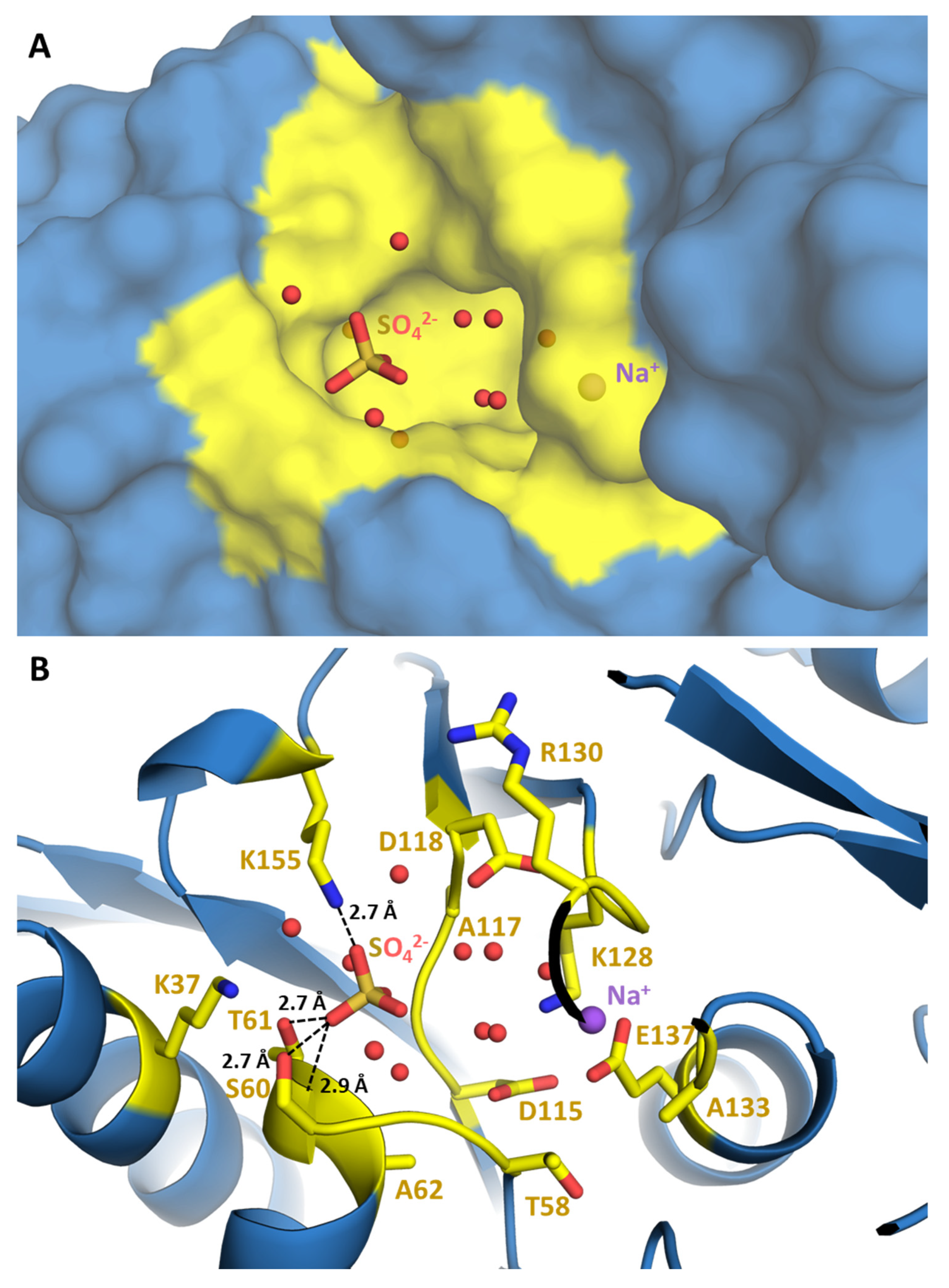
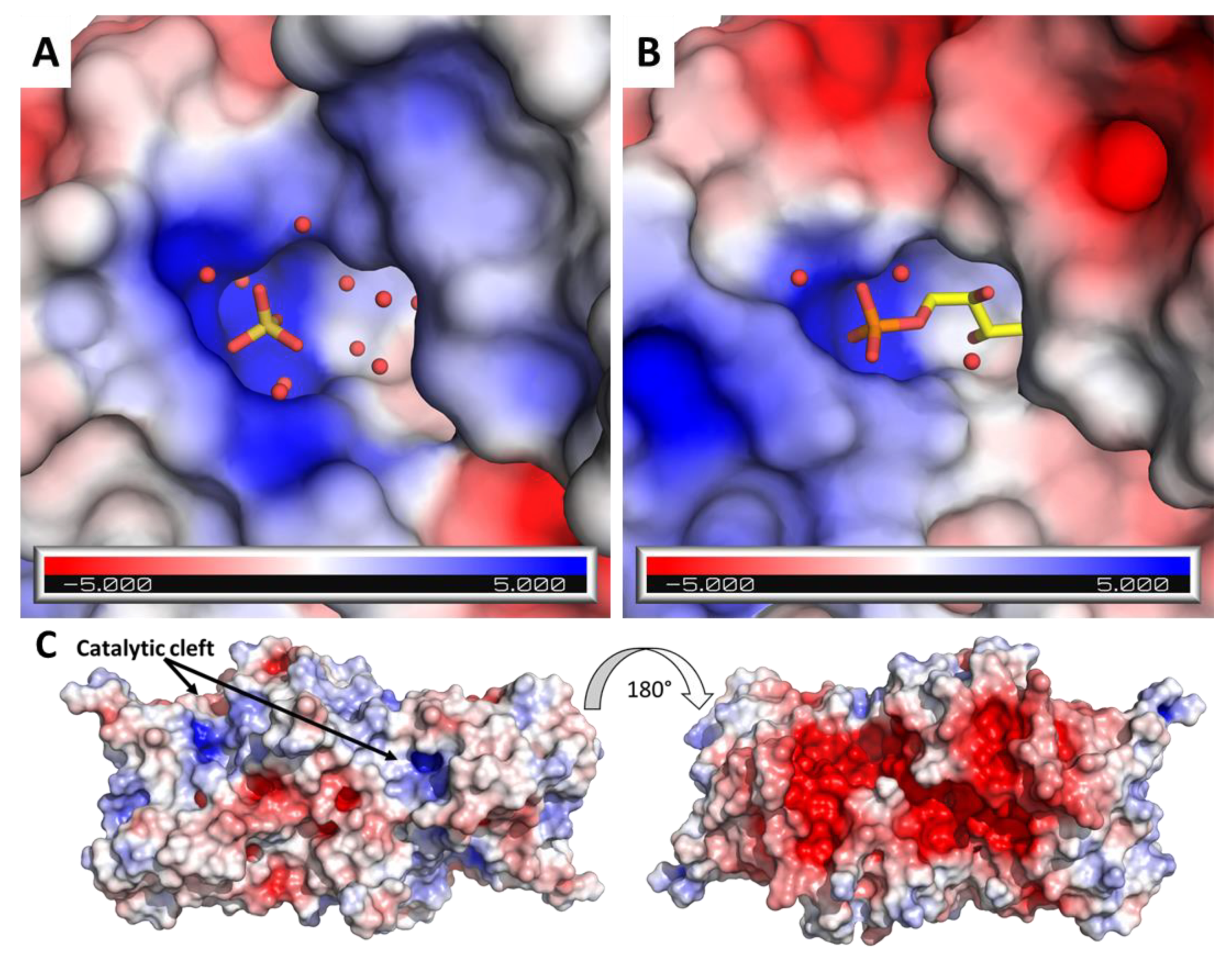
2.3. RPI1 Catalytic Site
2.4. CrRPI1 Sites of Post-Translational Modifications
3. Discussion
4. Materials and Methods
4.1. Cloning
4.2. Protein Expression and Purification
4.3. Protein Crystallization and Structure Determination
4.4. SEC-SAXS Analysis
4.5. Structural Data
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pfannschmidt, T.; Yang, C. The hidden function of photosynthesis: A sensing system for environmental conditions that regulates plant acclimation responses. Protoplasma 2012, 249 (Suppl 2), S125–S136. [Google Scholar] [CrossRef]
- Benson, A.A.; Bassham, J.A.; Calvin, M.; Hall, A.G.; Hirsch, H.E.; Kawaguchi, S.; Lynch, V.; Tolbert, N.E. The path of carbon in photosynthesis. XV. Ribulose and sedoheptulose. J. Biol. Chem. 1952, 196, 703–716. [Google Scholar] [PubMed]
- Zhang, R.; Andersson, C.E.; Savchenko, A.; Skarina, T.; Evdokimova, E.; Beasley, S.; Arrowsmith, C.H.; Edwards, A.M.; Joachimiak, A.; Mowbray, S.L. Structure of Escherichia coli ribose-5-phosphate isomerase: A ubiquitous enzyme of the pentose phosphate pathway and the Calvin cycle. Structure 2003, 11, 31–42. [Google Scholar] [CrossRef]
- Chen, J.; Wu, H.; Zhang, W.; Mu, W. Ribose-5-phosphate isomerases: Characteristics, structural features, and applications. Appl. Microbiol. Biotechnol. 2020, 104, 6429–6441. [Google Scholar] [CrossRef] [PubMed]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Gruning, N.M.; Kruger, A.; Tauqeer Alam, M.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. Camb. Philos. Soc. 2015, 90, 927–963. [Google Scholar] [CrossRef]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. From molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. eLife 2018, 7, e39233. [Google Scholar] [CrossRef] [PubMed]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Marechal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Tardif, M.; Atteia, A.; Specht, M.; Cogne, G.; Rolland, N.; Brugiere, S.; Hippler, M.; Ferro, M.; Bruley, C.; Peltier, G.; et al. PredAlgo: A new subcellular localization prediction tool dedicated to green algae. Mol. Biol. Evol. 2012, 29, 3625–3639. [Google Scholar] [CrossRef] [PubMed]
- Johnson, X.; Alric, J. Central carbon metabolism and electron transport in Chlamydomonas reinhardtii: Metabolic constraints for carbon partitioning between oil and starch. Eukaryot. Cell 2013, 12, 776–793. [Google Scholar] [CrossRef]
- Klein, U. Compartmentation of glycolysis and of the oxidative pentose-phosphate pathway in Chlamydomonas reinhardii. Planta 1986, 167, 81–86. [Google Scholar] [CrossRef]
- Strenkert, D.; Schmollinger, S.; Gallaher, S.D.; Salome, P.A.; Purvine, S.O.; Nicora, C.D.; Mettler-Altmann, T.; Soubeyrand, E.; Weber, A.P.M.; Lipton, M.S.; et al. Multiomics resolution of molecular events during a day in the life of Chlamydomonas. Proc. Natl. Acad. Sci. USA 2019, 116, 2374–2383. [Google Scholar] [CrossRef] [PubMed]
- Hammel, A.; Sommer, F.; Zimmer, D.; Stitt, M.; Mühlhaus, T.; Schroda, M. Overexpression of sedoheptulose-1,7-bisphosphatase enhances photosynthesis in Chlamydomonas reinhardtii and has no effect on the abundance of other Calvin-Benson cycle enzymes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mettler, T.; Muhlhaus, T.; Hemme, D.; Schottler, M.A.; Rupprecht, J.; Idoine, A.; Veyel, D.; Pal, S.K.; Yaneva-Roder, L.; Winck, F.V.; et al. Systems Analysis of the Response of Photosynthesis, Metabolism, and Growth to an Increase in Irradiance in the Photosynthetic Model Organism Chlamydomonas reinhardtii. Plant Cell 2014, 26, 2310–2350. [Google Scholar] [CrossRef] [PubMed]
- Scheibe, R. Redox-modulation of chloroplast enzymes: A common principle for individual control. Plant Physiol. 1991, 96, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Michelet, L.; Zaffagnini, M.; Morisse, S.; Sparla, F.; Perez-Perez, M.E.; Francia, F.; Danon, A.; Marchand, C.H.; Fermani, S.; Trost, P.; et al. Redox regulation of the Calvin-Benson cycle: Something old, something new. Front. Plant Sci. 2013, 4, 470. [Google Scholar] [CrossRef]
- Morisse, S.; Zaffagnini, M.; Gao, X.H.; Lemaire, S.D.; Marchand, C.H. Insight into protein S-nitrosylation in Chlamydomonas reinhardtii. Antioxid. Redox. Signal. 2014, 21, 1271–1284. [Google Scholar] [CrossRef]
- Michelet, L.; Zaffagnini, M.; Vanacker, H.; Le Marechal, P.; Marchand, C.; Schroda, M.; Lemaire, S.D.; Decottignies, P. In vivo targets of S-thiolation in Chlamydomonas reinhardtii. J. Biol. Chem. 2008, 283, 21571–21578. [Google Scholar] [CrossRef]
- Huppe, H.C.; Buchanan, B.B. Activation of a chloroplast type of fructose bisphosphatase from Chlamydomonas reinhardtii by light-mediated agents. Z. Naturforsch. C J. Biosci. 1989, 44, 487–494. [Google Scholar] [CrossRef]
- Pasquini, M.; Fermani, S.; Tedesco, D.; Sciabolini, C.; Crozet, P.; Naldi, M.; Henri, J.; Vothknecht, U.; Bertucci, C.; Lemaire, S.D.; et al. Structural basis for the magnesium-dependent activation of transketolase from Chlamydomonas reinhardtii. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2132–2145. [Google Scholar] [CrossRef]
- Perez-Perez, M.E.; Mauries, A.; Maes, A.; Tourasse, N.J.; Hamon, M.; Lemaire, S.D.; Marchand, C.H. The Deep Thioredoxome in Chlamydomonas reinhardtii: New Insights into Redox Regulation. Mol. Plant 2017, 10, 1107–1125. [Google Scholar] [CrossRef]
- Wang, H.; Gau, B.; Slade, W.O.; Juergens, M.; Li, P.; Hicks, L.M. The global phosphoproteome of Chlamydomonas reinhardtii reveals complex organellar phosphorylation in the flagella and thylakoid membrane. Mol. Cell. Proteom. 2014, 13, 2337–2353. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.A.; Buckner, F.S.; Van Voorhis, W.C.; Verlinde, C.L.; Mehlin, C.; Boni, E.; DeTitta, G.; Luft, J.; Lauricella, A.; Anderson, L.; et al. Structure of ribose 5-phosphate isomerase from Plasmodium falciparum. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Essenberg, M.K.; Cooper, R.A. Two ribose-5-phosphate isomerases from Escherichia coli K12: Partial characterisation of the enzymes and consideration of their possible physiological roles. Eur. J. Biochem. 1975, 55, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Rutner, A.C. Spinach 5-phosphoribose isomerase. Purification and properties of the enzyme. Biochemistry 1970, 9, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]
- Landau, M.; Mayrose, I.; Rosenberg, Y.; Glaser, F.; Martz, E.; Pupko, T.; Ben-Tal, N. ConSurf 2005: The projection of evolutionary conservation scores of residues on protein structures. Nucleic Acids Res. 2005, 33, W299–W302. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K.; et al. Improvements to the APBS biomolecular solvation software suite. Protein Sci. Publ. Protein Soc. 2018, 27, 112–128. [Google Scholar] [CrossRef]
- Dolinsky, T.J.; Czodrowski, P.; Li, H.; Nielsen, J.E.; Jensen, J.H.; Klebe, G.; Baker, N.A. PDB2PQR: Expanding and upgrading automated preparation of biomolecular structures for molecular simulations. Nucleic Acids Res. 2007, 35, W522–W525. [Google Scholar] [CrossRef]
- Hamada, K.; Ago, H.; Sugahara, M.; Nodake, Y.; Kuramitsu, S.; Miyano, M. Oxyanion hole-stabilized stereospecific isomerization in ribose-5-phosphate isomerase (Rpi). J. Biol. Chem. 2003, 278, 49183–49190. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Bedhomme, M.; Groni, H.; Marchand, C.H.; Puppo, C.; Gontero, B.; Cassier-Chauvat, C.; Decottignies, P.; Lemaire, S.D. Glutathionylation in the photosynthetic model organism Chlamydomonas reinhardtii: A proteomic survey. Mol. Cell. Proteom. 2012, 11, M111.014142. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Morisse, S.; Michelet, L.; Bedhomme, M.; Marchand, C.H.; Calvaresi, M.; Trost, P.; Fermani, S.; Zaffagnini, M.; Lemaire, S.D. Thioredoxin-dependent redox regulation of chloroplastic phosphoglycerate kinase from Chlamydomonas reinhardtii. J. Biol. Chem. 2014, 289, 30012–30024. [Google Scholar] [CrossRef] [PubMed]
- Marri, L.; Zaffagnini, M.; Collin, V.; Issakidis-Bourguet, E.; Lemaire, S.D.; Pupillo, P.; Sparla, F.; Miginiac-Maslow, M.; Trost, P. Prompt and easy activation by specific thioredoxins of calvin cycle enzymes of Arabidopsis thaliana associated in the GAPDH/CP12/PRK supramolecular complex. Mol. Plant 2009, 2, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, S.D.; Tedesco, D.; Crozet, P.; Michelet, L.; Fermani, S.; Zaffagnini, M.; Henri, J. Crystal Structure of Chloroplastic Thioredoxin f2 from Chlamydomonas reinhardtii Reveals Distinct Surface Properties. Antioxidants 2018, 7, 171. [Google Scholar] [CrossRef] [PubMed]
- Gurrieri, L.; Del Giudice, A.; Demitri, N.; Falini, G.; Pavel, N.V.; Zaffagnini, M.; Polentarutti, M.; Crozet, P.; Marchand, C.H.; Henri, J.; et al. Arabidopsis and Chlamydomonas phosphoribulokinase crystal structures complete the redox structural proteome of the Calvin-Benson cycle. Proc. Natl. Acad. Sci. USA 2019, 116, 8048–8053. [Google Scholar] [CrossRef]
- Graille, M.; Meyer, P.; Leulliot, N.; Sorel, I.; Janin, J.; Van Tilbeurgh, H.; Quevillon-Cheruel, S. Crystal structure of the S. cerevisiae D-ribose-5-phosphate isomerase: Comparison with the archaeal and bacterial enzymes. Biochimie 2005, 87, 763–769. [Google Scholar] [CrossRef]
- Ishikawa, K.; Matsui, I.; Payan, F.; Cambillau, C.; Ishida, H.; Kawarabayasi, Y.; Kikuchi, H.; Roussel, A. A hyperthermostable D-ribose-5-phosphate isomerase from Pyrococcus horikoshii characterization and three-dimensional structure. Structure 2002, 10, 877–886. [Google Scholar] [CrossRef]
- Dickens, F. Recent advances in knowledge of the hexose monophosphate shunt. Ann. N. Y. Acad. Sci. 1958, 75, 71–94. [Google Scholar] [CrossRef]
- Kuken, A.; Sommer, F.; Yaneva-Roder, L.; Mackinder, L.C.; Hohne, M.; Geimer, S.; Jonikas, M.C.; Schroda, M.; Stitt, M.; Nikoloski, Z.; et al. Effects of microcompartmentation on flux distribution and metabolic pools in Chlamydomonas reinhardtii chloroplasts. eLife 2018, 7, e37960. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef]
- Emanuelsson, O.; Nielsen, H.; von Heijne, G. ChloroP, a neural network-based method for predicting chloroplast transit peptides and their cleavage sites. Protein Sci. Publ. Protein Soc. 1999, 8, 978–984. [Google Scholar] [CrossRef]
- Asamizu, E.; Nakamura, Y.; Miura, K.; Fukuzawa, H.; Fujiwara, S.; Hirono, M.; Iwamoto, K.; Matsuda, Y.; Minagawa, J.; Shimogawara, K.; et al. Establishment of publicly available cDNA material and information resource of Chlamydomonas reinhardtii (Chlorophyta) to facilitate gene function analysis. Phycologia 2004, 43, 722–726. [Google Scholar] [CrossRef]
- Lesley, S.A.; Wilson, I.A. Protein production and crystallization at the joint center for structural genomics. J. Struct. Funct. Genom. 2005, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Coati, A.; Chavas, L.M.G.; Fontaine, P.; Foos, N.; Guimaraes, B.; Gourhant, P.; Legrand, P.; Itie, J.P.; Fertey, P.; Shepard, W.; et al. Status of the crystallography beamlines at synchrotron SOLEIL. Eur. Phys. J. Plus 2017, 132, 174. [Google Scholar] [CrossRef]
- Legrand, P. XDSME: XDS Made Easier. 2017. Available online: https://github.com/legrandp/xdsme (accessed on 20 October 2020).
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Echols, N.; Headd, J.J.; Hung, L.W.; Jain, S.; Kapral, G.J.; Grosse Kunstleve, R.W.; et al. The Phenix software for automated determination of macromolecular structures. Methods 2011, 55, 94–106. [Google Scholar] [CrossRef] [PubMed]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [PubMed]
- Raman, S.; Vernon, R.; Thompson, J.; Tyka, M.; Sadreyev, R.; Pei, J.; Kim, D.; Kellogg, E.; DiMaio, F.; Lange, O.; et al. Structure prediction for CASP8 with all-atom refinement using Rosetta. Proteins 2009, 77 (Suppl. 9), 89–99. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. Sect. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Petoukhov, M.V.; Franke, D.; Shkumatov, A.V.; Tria, G.; Kikhney, A.G.; Gajda, M.; Gorba, C.; Mertens, H.D.; Konarev, P.V.; Svergun, D.I. New developments in the ATSAS program package for small-angle scattering data analysis. J. Appl. Crystallogr. 2012, 45, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Franke, D.; Petoukhov, M.V.; Konarev, P.V.; Panjkovich, A.; Tuukkanen, A.; Mertens, H.D.T.; Kikhney, A.G.; Hajizadeh, N.R.; Franklin, J.M.; Jeffries, C.M.; et al. ATSAS 2.8: A comprehensive data analysis suite for small-angle scattering from macromolecular solutions. J. Appl. Crystallogr. 2017, 50, 1212–1225. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CrRPI1 | |
|---|---|
| Wavelength (Å) | 0.979 |
| Resolution range (Å) | 40.22–1.40 (1.45–1.40) |
| Space group | P 21 |
| Unit cell | 45.15 63.95 80.91 90 96.13 90 |
| Total reflections | 613,155 (55,349) |
| Unique reflections | 89,012 (8611) |
| Multiplicity | 6.9 (6.4) |
| Completeness (%) | 99.6 (96.5) |
| Mean I/sigma(I) | 20.58 (1.24) |
| Wilson B-factor (Å2) | 21.45 |
| R-merge | 0.0519 (1.272) |
| R-meas | 0.05611 (1.382) |
| R-pim | 0.02114 (0.5327) |
| CC1/2 | 0.999 (0.382) |
| CC* | 1 (0.744) |
| Reflections used in refinement | 88,983 (8601) |
| Reflections used for R-free | 2000 (194) |
| R-work | 0.1594 (0.3457) |
| R-free | 0.1774 (0.3813) |
| CC (work) | 0.972 (0.730) |
| CC (free) | 0.967 (0.836) |
| Number of non-hydrogen atoms | 4206 |
| macromolecules | 3648 |
| ligands | 47 |
| solvent | 511 |
| Protein residues | 474 |
| RMS (bonds) (Å) | 0.011 |
| RMS (angles) (deg) | 1.18 |
| Ramachandran favored (%) | 99.79 |
| Ramachandran allowed (%) | 0.21 |
| Ramachandran outliers (%) | 0.00 |
| Rotamer outliers (%) | 1.00 |
| Clashscore | 2.69 |
| Average B-factor (Å2) | 25.46 |
| macromolecules | 23.78 |
| ligands | 47.04 |
| Solvent | 35.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Moigne, T.; Crozet, P.; Lemaire, S.D.; Henri, J. High-Resolution Crystal Structure of Chloroplastic Ribose-5-Phosphate Isomerase from Chlamydomonas reinhardtii—An Enzyme Involved in the Photosynthetic Calvin-Benson Cycle. Int. J. Mol. Sci. 2020, 21, 7787. https://doi.org/10.3390/ijms21207787
Le Moigne T, Crozet P, Lemaire SD, Henri J. High-Resolution Crystal Structure of Chloroplastic Ribose-5-Phosphate Isomerase from Chlamydomonas reinhardtii—An Enzyme Involved in the Photosynthetic Calvin-Benson Cycle. International Journal of Molecular Sciences. 2020; 21(20):7787. https://doi.org/10.3390/ijms21207787
Chicago/Turabian StyleLe Moigne, Théo, Pierre Crozet, Stéphane D. Lemaire, and Julien Henri. 2020. "High-Resolution Crystal Structure of Chloroplastic Ribose-5-Phosphate Isomerase from Chlamydomonas reinhardtii—An Enzyme Involved in the Photosynthetic Calvin-Benson Cycle" International Journal of Molecular Sciences 21, no. 20: 7787. https://doi.org/10.3390/ijms21207787
APA StyleLe Moigne, T., Crozet, P., Lemaire, S. D., & Henri, J. (2020). High-Resolution Crystal Structure of Chloroplastic Ribose-5-Phosphate Isomerase from Chlamydomonas reinhardtii—An Enzyme Involved in the Photosynthetic Calvin-Benson Cycle. International Journal of Molecular Sciences, 21(20), 7787. https://doi.org/10.3390/ijms21207787