Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation


 and
and 
Abstract
1. Introduction
2. Results
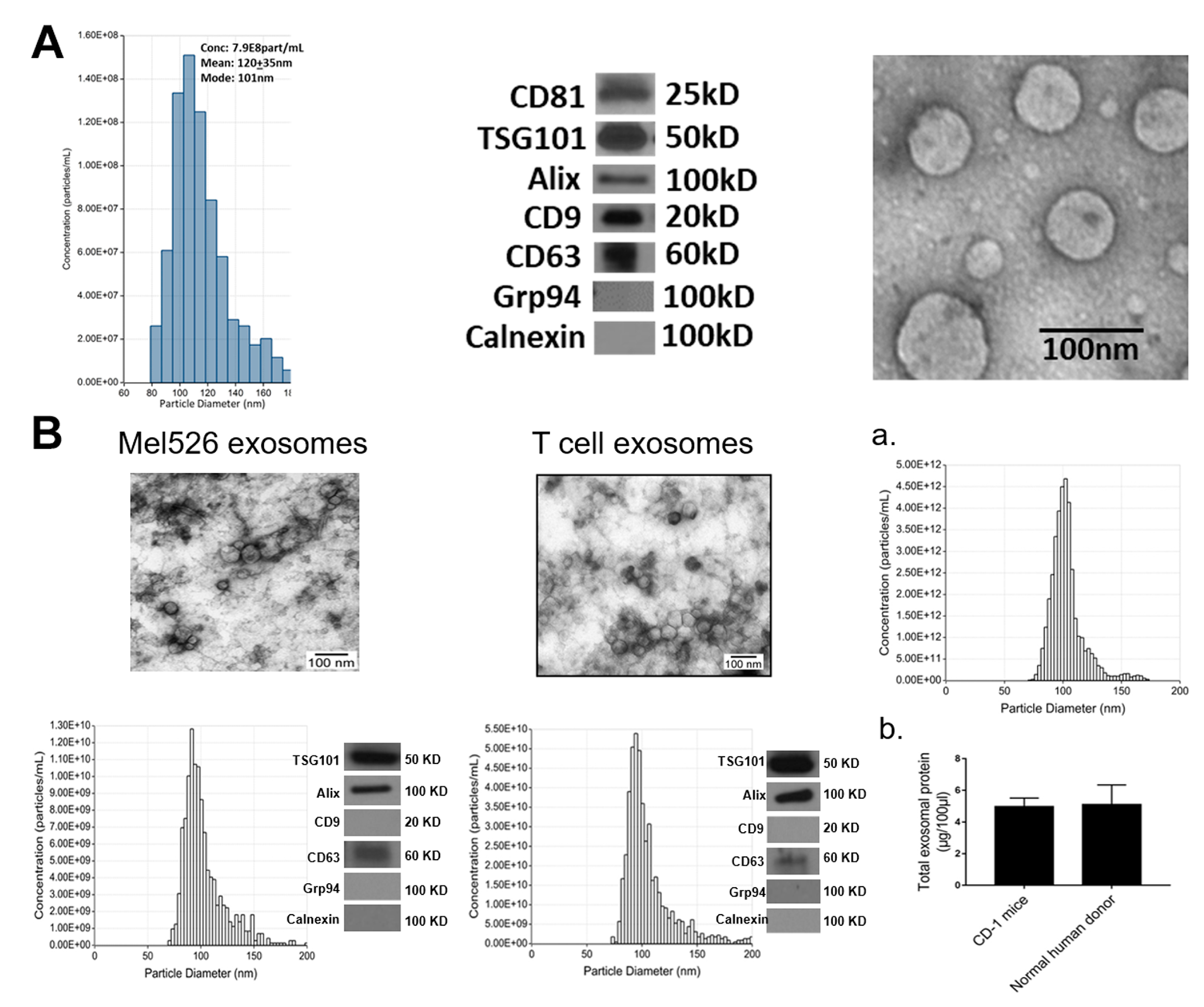
2.1. Exosome Characteristics
2.2. Clearance of Exosomes from Blood
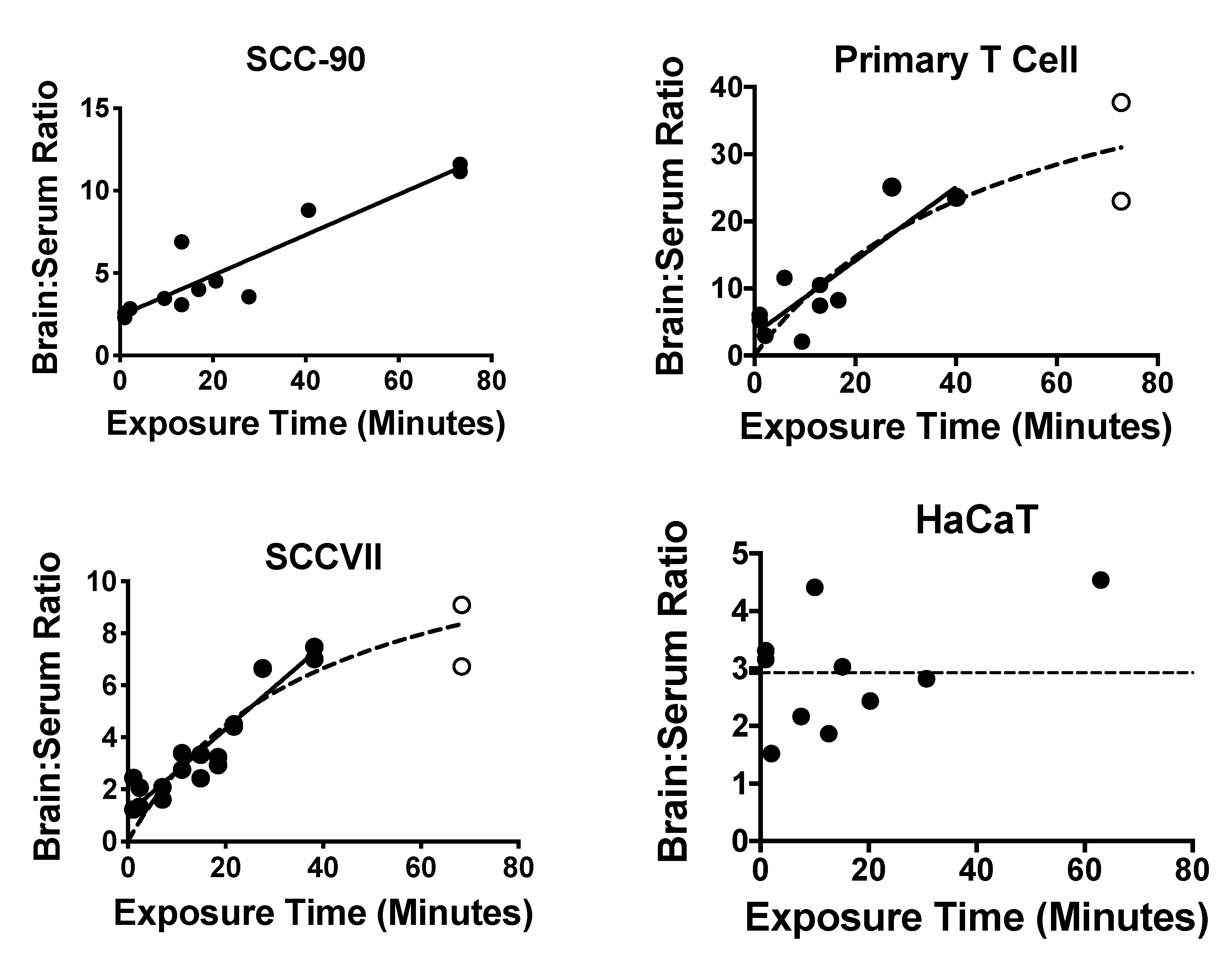
2.3. Exosome Uptake by Whole Brain
2.4. Capillary Depletion
2.5. Exosome Uptake by Different Brain Regions
2.6. Effects of LPS on Exosome Uptake
2.7. Effects of WGA and M6P on Exosome Uptake
2.8. Brain-to-Blood Efflux of Exosomes
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Culture of Primary Human T Cells
4.3. Isolation of Exosomes by Mini-Size Exclusion Chromatography
4.4. BCA Protein Assay
4.5. Transmission Electron Microscopy
4.6. Tunable Resistive Pulse Sensing (TRPS)
4.7. Western Blot Analysis of Exosomes
4.8. On-Bead Flow Cytometry for Exosomes
4.9. Radioactive Labeling and Purifying
4.10. Circulating Levels of Exosomes in Mice
4.11. Intravenous (IV) Time Curve and Brain Uptake
4.12. Lipopolysaccharide (LPS) Preparation and Treatment
4.13. Effects of Mannose 6-Phosphate and Wheatgerm Agglutinin on Tissue Uptake of Exosomes
4.14. Capillary Depletion with Washout
4.15. Intracerebroventricular (ICV) Injection of Exosomes
4.16. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BBB | Blood–brain barrier |
| CNS | Central nervous system |
| EVs | Extracellular vesicles |
| LPS | Lipopolysaccharide |
| MVs | Microvesicles |
| MVBs | Multivesicular bodies |
| t1/2 | Half-time clearance |
| WGA | Wheatgerm agglutinin |
| %Inj/mL | Percent of the injected dose present in one ml of serum |
References
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [Google Scholar] [CrossRef] [PubMed]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol 2017, 27, 172–188. [Google Scholar] [CrossRef]
- Shi, M.; Kovac, A.; Korff, A.; Cook, D.G.; Ginghina, C.; Bullock, K.M.; Yang, L.; Zheng, D.; Aro, P.; Stewart, T.; et al. CNS tau efflux via exosomes is likey decreased in Parkinson disease but not in Alzheimer disease. Alzhemier Dement. 2016, 12, 1125–1131. [Google Scholar] [CrossRef]
- Shi, M.; Liu, C.; Cook, T.J.; Bullock, K.M.; Zhao, Y.; Ginghina, C.; Li, Y.; Aro, P.; Dator, R.; He, C.; et al. Plasma exosomal alpha-synuclein is likely CNS-derived and increased in Parkinson’s disease. Acta Neuropathol. 2014, 128, 639–650. [Google Scholar] [CrossRef]
- Dickens, A.M.; Tovar-y-Romo, L.B.; Yoo, S.-W.; Trout, A.L.; Bae, M.; Kanmogne, M.; Megra, B.; Williams, D.W.; Witwer, K.W.; Gacias, M.; et al. Astrocyte-shed extracellular vesicles regulate the peripheral leukocyte response to inflammatory brain lesions. Sci. Signal. 2012, 10, 473. [Google Scholar] [CrossRef]
- Ridder, K.; Keller, S.; Dams, M.; Rupp, A.-K.; Schlaudraff, J.; Del Turco, D.; Starmann, J.; Macas, J.; Karpova, D.; Devraj, K.; et al. Extracellular vesicle-mediated transfer of genetic information between the hematopoietic system and the brain response in inflammation. PLoS Biol. 2014, 12, e1001874. [Google Scholar] [CrossRef]
- Matsumoto, J.; Stewart, T.; Sheng, L.; Li, N.; Bullock, N.; Song, N.; Shi, M.; Banks, W.A.; Zhang, J. Transmission of alpha-synuclein-containing erythrocyte-derived extracellular vesicles across the blood-brain barrier via adsorptive mediated transcytosis: Another mechanism for initiation and progression of Parkinson’s disease. Acta Neurol. Commun. 2017, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Whiteside, T.L. Potential roles of tumor-derived exosomes in angiogenesis. Expert Opin. Targets 2018, 22, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, N.; Yerneni, S.S.; Razzo, B.M.; Whiteside, T.L. Exosomes from HNSCC promote angiogenesis through reprogramming of endothelial cells. Mol. Cancer Res. 2018, 16, 1798–1808. [Google Scholar] [CrossRef]
- Whiteside, T.L. Exosomes carrying immunoinhibitory proteins and their role in cancer. Clin. Exp. Immunol. 2017, 189, 259–267. [Google Scholar] [CrossRef]
- Banks, W.A. From blood-brain barrier to blood-brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2016. [Google Scholar] [CrossRef]
- Erickson, M.A.; Banks, W.A. Neuroimmune axes of the blood-brain barriers and blood-brain interfaces: Bases for physiological regulation, disease states, and pharmacological interventions. Pharmacol. Rev. 2018, 70, 278–314. [Google Scholar] [CrossRef]
- Turowski, P.; Adamson, P.; Greenwood, J. Pharmacological targeting of ICAM-1 signaling in brain endothelial cells: Potential for treating neuroinflammation. Cell. Mol. Neurobiol. 2005, 25, 153–170. [Google Scholar] [CrossRef]
- Wolburg, H.; Wolburg-Buchholz, K.; Engelhardt, B. Diapedesis of mononuclear cells across cerebral venules during experimental autoimmune encephalomyelitis leaves tight junctions intact. Acta Neuropathol. 2005, 109, 181–190. [Google Scholar] [CrossRef]
- Banks, W.A.; Freed, E.O.; Wolf, K.M.; Robinson, S.M.; Franko, M.; Kumar, V.B. Transport of human immunodeficiency virus type 1 pseudoviruses across the blood-brain barrier: Role of envelope proteins and adsorptive endocytosis. J. Virol. 2001, 75, 4681–4691. [Google Scholar] [CrossRef]
- Matsumoto, J.; Stewart, T.; Banks, W.A.; Zhang, J. The transport mechanism of extracellular vesicles at the blood-brain barrier. Curr. Pharm. Des. 2017, 23, 6206–6214. [Google Scholar] [CrossRef] [PubMed]
- Wurdinger, T.; Gatson, N.N.; Balaj, L.; Kaur, B.; Breakefield, X.O.; Pegtel, D.M. Extracellular vesicles and their convergence with viral pathways. Adv. Virol. 2012, 2012, 767694. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Liu, L.; Ma, F.; Wong, C.W.; Guo, X.E.; Chacko, J.V.; Farhoodi, H.P.; Zhang, S.X.; Zimak, J.; Segaliny, A.; et al. Elucidation of exosome migration across the blood-brain barrier model in vitro. Cell. Mol. Bioeng. 2016, 9, 509–529. [Google Scholar] [CrossRef] [PubMed]
- Broadwell, R.D.; Balin, B.J.; Salcman, M. Transcytotic pathway for blood-borne protein through the blood-brain barrier. Proc. Natl. Acad. Sci. USA 1988, 85, 632–636. [Google Scholar] [CrossRef]
- Villegas, J.C.; Broadwell, R.D. Transcytosis of protein through the mammalian cerebral epithelium and endothelium: II. Adsorptive transcytosis of WGA-HRP and the blood-brain and brain-blood barriers. J. Neurocytol. 1993, 22, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Raub, T.J.; Audus, K.L. Adsorptive endocytosis and membrane recycling by cultured primary bovine brain microvessel endothelial cell monolayers. J. Cell Sci. 1990, 97, 127–138. [Google Scholar]
- Herve, F.; Ghinea, N.; Scherrmann, J.M. CNS delivery via adsorptive transcytosis. Aaps. J. 2008, 10, 455–472. [Google Scholar] [CrossRef]
- Vorbrodt, A.W.; Trowbridge, R.S. Ultrastructural study of transcellular transport of native and cationized albumin in cultured sheep brain microvascular endothelium. J. Neurocytol. 1991, 20, 998–1006. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J.; Brennan, J.M.; Vallance, K.L. Adsorptive endocytosis of HIV-1gp120 by blood-brain barrier is enhanced by lipopolysaccharide. Exp. Neurol. 1999, 156, 165–171. [Google Scholar] [CrossRef]
- Banks, W.A.; Niehoff, M.L.; Ponzio, N.M.; Erickson, M.A.; Zalcman, S.S. Pharmacokinetics and modeling of immune cell trafficking: Quantifying differential influences of target tissues versus lymphocytes in SJL and lippolysaccaride-treated mice. J. Neuroinflam. 2012, 9, 231. [Google Scholar] [CrossRef]
- Banks, W.A.; Kastin, A.J.; Akerstrom, V. HIV-1 protein gp120 crosses the blood-brain barrier: Role of adsorptive endocytosis. Life Sci. 1997, 61, L119–L125. [Google Scholar] [CrossRef]
- Banks, W.A.; Broadwell, R.D. Blood to brain and brain to blood passage of native horseradish peroxidase, wheat germ agglutinin and albumin: Pharmacokinetic and morphological assessments. J. Neurochem. 1994, 62, 2404–2419. [Google Scholar] [CrossRef] [PubMed]
- Dohgu, S.; Ryerse, J.S.; Robinson, S.M.; Banks, W.A. Human immunodeficiency virus-1 uses the mannose-6-phosphate receptor to cross the blood-brain barrier. PLoS ONE 2012, 7, e41623. [Google Scholar] [CrossRef] [PubMed]
- Minami, T.; Okazaki, J.; Kawabata, A.; Kawaki, H.; Okazaki, Y.; Tohno, Y. Roles of nitric oxide and prostaglandins in the increased permeability of the blood-brain barrier caused by lipopolysaccharide. Environ. Toxicol Pharm. 1998, 5, 35–41. [Google Scholar] [CrossRef]
- Pan, W.; Yu, C.; Hsuchou, H.; Kastin, A.J. The role of cerebrovascular NFkappaB in LPS-induced inflammation: Differntial regulation of efflux transporter and transporting cytokine receptors. Cell Physiol. Biochem. 2010, 25, 623–630. [Google Scholar] [CrossRef]
- Ludwig, N.; Razzo, B.M.; Yerneni, S.S.; Whiteside, T.L. Optimization of cell culture conditions for exosome isolation using mini-size exclusion chromatography (mini-SEC). Exp. Cell Res. 2019, 378, 149–157. [Google Scholar] [CrossRef]
- Ludwig, N.; Hong, C.-S.; Ludwig, S.; Azambuja, J.H.; Sharma, P.; Theodoraki, M.-N.; Whiteside, T.L. Isolation and analysis of tumor-derived exosomes. Curr. Protoc. Immunol. 2019, 127, e91. [Google Scholar] [CrossRef]
- Yuan, D.; Zhao, Y.; Banks, W.A.; Bullock, K.M.; Haney, M.; Batrakova, E.; Kabanov, A.V. Macrophage exosomes as natural nanocarriers for protein delivery to inflamed brain. Biomaterials 2017, 142, 1–12. [Google Scholar] [CrossRef]
- Kuroda, H.; Tachikawa, M.; Yagi, Y.; Umetsu, M.; Nurdin, A.; Miyauchi, E.; Watanabe, M.; Uchida, Y.; Terasaki, T. Cluster of Differentiation 46 is the Major Receptor in Human Blood-Brain Barrier Endothelial Cells for Uptake of Exosomes Derived from Brain-Metastatic Melanoma Cells (SK-Mel-28). Mol. Pharm. 2018, 16, 292–304. [Google Scholar] [CrossRef]
- Malm, T.; Loppi, S.; Kanninen, K.M. Exosomes in Alzheimer’s disease. Neurochem. Int. 2016, 97, 193–199. [Google Scholar] [CrossRef]
- Charoenviriyakul, C.; Takahashi, Y.; Morishita, M.; Matsumoto, A.; Nishikawa, M.; Takakura, Y. Cell type-specific and common characteristics of exosomes derived from mouse cell lines: Yield, physicochemical properties, and pharmacokinetics. Eur. J. Pharm. Sci. 2017, 96, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Takahashi, Y.; Nishikawa, M.; Kato, K.; Morishita, M.; Yamashita, T.; Matsumoto, A.; Charoenviriyakul, C.; Takakura, Y. Macrophage-dependent clearance of systemically administered B16BL6-derived exosomes from the blood circulation in mice. J. Extracell Vesicles 2015, 4, 26238. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Takahashi, Y.; Chang, H.-Y.; Wu, Y.-W.; Yamamoto, A.; Ishihama, Y.; Takakura, Y. Blood concentrations of small extracelluar vesicles are determined by a balance between abundant secretion and rapid clearance. J. Extracell Vesicles 2020, 9, 1. [Google Scholar] [CrossRef]
- Saunders, N.R.; Dziegielewska, K.M.; Mollgard, K.; Habgood, M.D. Markers for blood-brain barrier integrity: How appropriate is Evans blue in the twenty-first century and what are the alternatives? Front. Neurosci. 2015, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Moinuddin, A.; Morley, J.E. Regional transport of TNF-alpha across the blood-brain barrier in young ICR and young and aged SAMP8 mice. Neurobiol. Aging 2001, 22, 671–676. [Google Scholar] [CrossRef]
- Davson, H.; Welch, K.; Segal, M.B. Some special aspects of the blood-brain barrier. In The Physiology and Pathophysiology of the Cerebrospinal Fluid; Churchill Livingstone: Edinburgh, UK, 1987; pp. 247–374. [Google Scholar]
- Banks, W.A.; Dohgu, S.; Nakaoke, R.; Lynch, J.L.; Fleegal-DeMotta, M.A.; Erickson, M.A.; Vo, T.Q. Nitric oxide isoenzymes regulate LPS-enhanced insulin transport across the blood-brain barrier. Endocrinology 2008, 149, 1514–1523. [Google Scholar] [CrossRef] [PubMed]
- Dohgu, S.; Banks, W.A. Lipopolysaccharide-enhanced transcellular transport of HIV-1 across the blood-brain barrier is medited by the p38 mitogen-activated protein kinase pathway. Exp. Neurol. 2008, 210, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Osburg, B.; Peiser, C.; Domling, D.; Schomburg, L.; Ko, Y.T.; Voight, K.; Bickel, U. Effect of endotoxin on expression of TNF receptors and transport of TNF-alpha at the blood-brain barrier of the rat. Am. J. Physiol. 2002, 283, E899–E908. [Google Scholar]
- Vorbrodt, A.W. Glycoconjugates and anionic sites in the blood-brain barrier. In Glycobiology and the Brain; Nicolini, M., Zatta, P.F., Eds.; Pergamon Press: Oxford, UK, 1994; pp. 37–62. [Google Scholar]
- Kim, J.J.; Olson, L.J.; Dahms, N.M. Carbohydrate recognition by the mannose-6-phosphate receptors. Curr. Opin. Struct. Biol. 2009, 19, 534–542. [Google Scholar] [CrossRef]
- Schuler, P.J.; Schilling, B.; Harasymczuk, M.; Hoffmann, T.K.; Johnson, J.; Lang, S.; Whiteside, T.L. Phenotypic and functional characteristics of CD4+ CD39+ FOXP3+, and CD4+ CD39+ FOXP3neg T-cell subsets in cancer patients. Eur. J. Immunol. 2012, 42, 1876–1885. [Google Scholar] [CrossRef]
- Hong, C.S.; Funk, S.; Muller, L.; Boyiadzis, M.; Whiteside, T.L. Isolation of biologically active and morphologically intact exosomes from plasma of patients with cancer. J. Extracell Vesicles 2016, 5, 29289. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Diergaarde, B.; Ferrone, S.; Kirkwood, J.M.; Whiteside, T.L. Melanoma cell-derived exosomes in plasma of melanoma patients suppress functions of immune effector cells. Sci. Rep. 2020, 10, 92. [Google Scholar] [CrossRef]
- Sharma, P.; Ludwig, S.; Muller, L.; Hong, C.S.; Kirkwood, J.M.; Ferrone, S.; Whiteside, T.L. Immunoaffinity-based isolation of melanoma cell-derived exosomes from plasma of patients with melanoma. J. Extracell Vesicles 2018, 7, 1435138. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.S.; Sharma, P.; Yerneni, S.S.; Simms, P.; Jackson, E.K.; Whiteside, T.L.; Boyiadzis, M. Circulating exosomes carrying an immunosuppresive cargo interfere with cellular immunotherapty in acute myeloid leukemia. Sci. Rep. 2017, 71, 14684. [Google Scholar] [CrossRef] [PubMed]
- Blasberg, R.G.; Fenstermacher, J.D.; Patlak, C.S. Transport of alpha-aminoisobutyric acid across brain capillary and cellular membranes. J. Cereb. Blood Flow Metab. 1983, 3, 8–32. [Google Scholar] [CrossRef] [PubMed]
- Patlak, C.S.; Blasberg, R.G.; Fenstermacher, J.D. Graphical evaluation of blood-to-brain transfer constants from multiple-time uptake data. J. Cereb. Blood Flow Metab. 1983, 3, 1–7. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Species | Tissue | Non-Cancerous/Cancer |
|---|---|---|---|
| J774A.1 | Mouse | Macrophage | Non-Cancerous |
| NIH-3T3 | Mouse | Fibroblast | Non-Cancerous |
| Primary T Cell | Human | T Cell | Non-Cancerous |
| HaCaT | Human | Keratinocyte | Non-Cancerous |
| SCCVII | Mouse | Oral Squamous | Cancer |
| MEL526 | Human | Melanoma | Cancer |
| MDA-MB-231 | Human | Breast | Cancer |
| PCI-30 | Human | Head & Neck | Cancer |
| SCC-90 | Human | Head & Neck | Cancer |
| Kasumi | Human | Leukemia | Cancer |
| Exosome | Y0 (Vd) | Plateau (Vd) | k (t1/2) | R2 (n) |
|---|---|---|---|---|
| Primary T CellNC | 1.30 (5.01) | 0.989 (10.2) | 0.227(3.05) | 0.610 (12) |
| SCC-90C | 1.63 (2.34) | 1.32 (4.79) | 0.095 (7.29) | 0.538 (13) |
| SCCVIIC | 1.72 (1.91) | 1.22 (6.03) | 0.182 (3.80) | 0.899 (20) |
| MDA-MB-231C | 1.77 (1.70) | 1.39 (4.07) | 0.339 (2.04) | 0.542 (12) |
| PCI-30C | 1.81 (1.41) | 1.41 (3.89) | 0.459 (1.51) | 0.464 (13) |
| %Inj/mL: Mean +/−SE (Vd) | n | |||
| KasumiC | 34.8 +/− 2.68 (2.87) | 13 | ||
| HaCaTNC | 43.2 +/− 1.48 (2.31) | 11 | ||
| NIH-3T3NC | 45.2 +/− 3.05 (2.21) | 12 | ||
| J774A.1NC | 33.6 +/− 2.19 (2.98) | 13 | ||
| MEL526C | 27.3 +/− 2.02 (3.66) | 12 | ||
| Exosome | Ki | Vi | n * | r2 * | p< * | Pattern | %Par |
|---|---|---|---|---|---|---|---|
| J774A.1NC | 0.083 ± 0.017 | 1.72 | 12 | 0.710 | 0.001 | Linear | 92 ± 0.6 |
| NIH-3T3NC | 0.103 ± 0.040 | 1.11 | 10 | 0.455 | 0.05 | Plateau | 58 ± 3.7 |
| Primary T CellNC | 0.547 ± 0.116 | 3.19 | 10 | 0.736 | 0.005 | Plateau | 85 ± 2.9 |
| HaCaTNC | Flat | 58 ± 8.1 | |||||
| SCC-90C | 0.123 ± 0.015 | 2.40 | 12 | 0.866 | 0.001 | Linear | 77 ± 2.0 |
| PCI-30C | 0.057 ± 0.011 | 4.11 | 13 | 0.721 | 0.001 | Linear | 71 ± 2.9 |
| SCCVIIC | 0.162 ± 0.015 | 1.07 | 18 | 0.884 | 0.001 | Plateau | 93 ± 3.0 |
| MDA-MB-231C | 0.098 ± 0.034 | 3.66 | 10 | 0.514 | 0.05 | Plateau | 64 ± 4.5 |
| MEL526C | 0.091 ± 0.035 | 1.7 | 7 | 0.579 | 0.05 | Plateau | 72 ± 3.9 |
| KasumiC | 0.044 ± 0.013 | 3.3 | 11 | 0.568 | 0.01 | Linear | 79 ± 4.6 |
| LPS No Increase | LPS Increase | |
|---|---|---|
| WGA No Increase | SCCVII * | Primary T Cell |
| MEL526 | MDA-MB-231 | |
| PCI-30 | ||
| WGA Increase | J774A.1 | HaCaT |
| NIH-3T3 | SCC-90 Kasumi |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banks, W.A.; Sharma, P.; Bullock, K.M.; Hansen, K.M.; Ludwig, N.; Whiteside, T.L. Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation. Int. J. Mol. Sci. 2020, 21, 4407. https://doi.org/10.3390/ijms21124407
Banks WA, Sharma P, Bullock KM, Hansen KM, Ludwig N, Whiteside TL. Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation. International Journal of Molecular Sciences. 2020; 21(12):4407. https://doi.org/10.3390/ijms21124407
Chicago/Turabian StyleBanks, William A., Priyanka Sharma, Kristin M. Bullock, Kim M. Hansen, Nils Ludwig, and Theresa L. Whiteside. 2020. "Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation" International Journal of Molecular Sciences 21, no. 12: 4407. https://doi.org/10.3390/ijms21124407
APA StyleBanks, W. A., Sharma, P., Bullock, K. M., Hansen, K. M., Ludwig, N., & Whiteside, T. L. (2020). Transport of Extracellular Vesicles across the Blood-Brain Barrier: Brain Pharmacokinetics and Effects of Inflammation. International Journal of Molecular Sciences, 21(12), 4407. https://doi.org/10.3390/ijms21124407