Mitochondrial Function in Modulating Human Granulosa Cell Steroidogenesis and Female Fertility

 ,
,  , , and
, , and 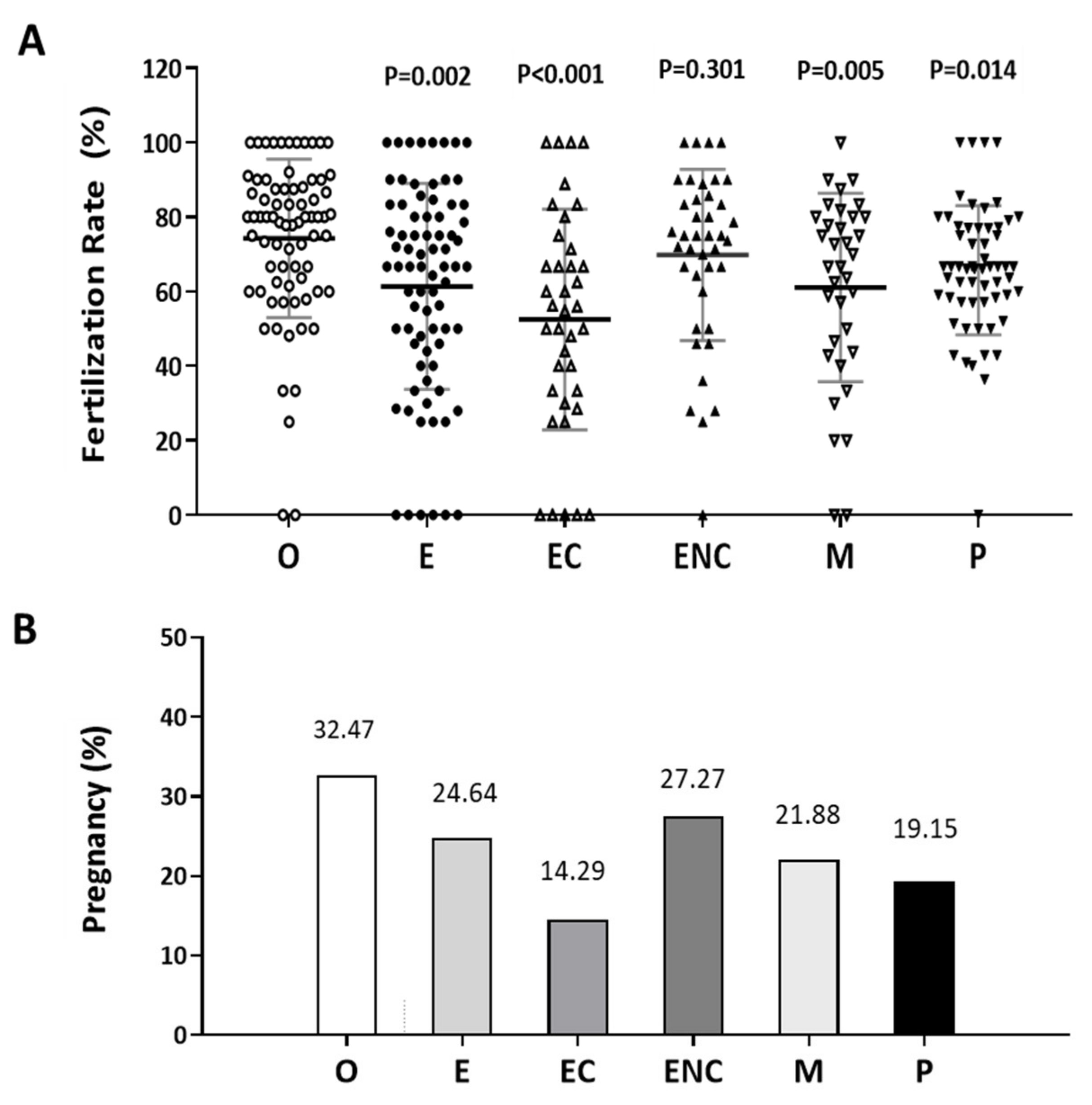{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. A Decreased Fertilization Rate and Pregnancy Outcome were Found in the Patient Group with Ovarian Endometriotic Cyst
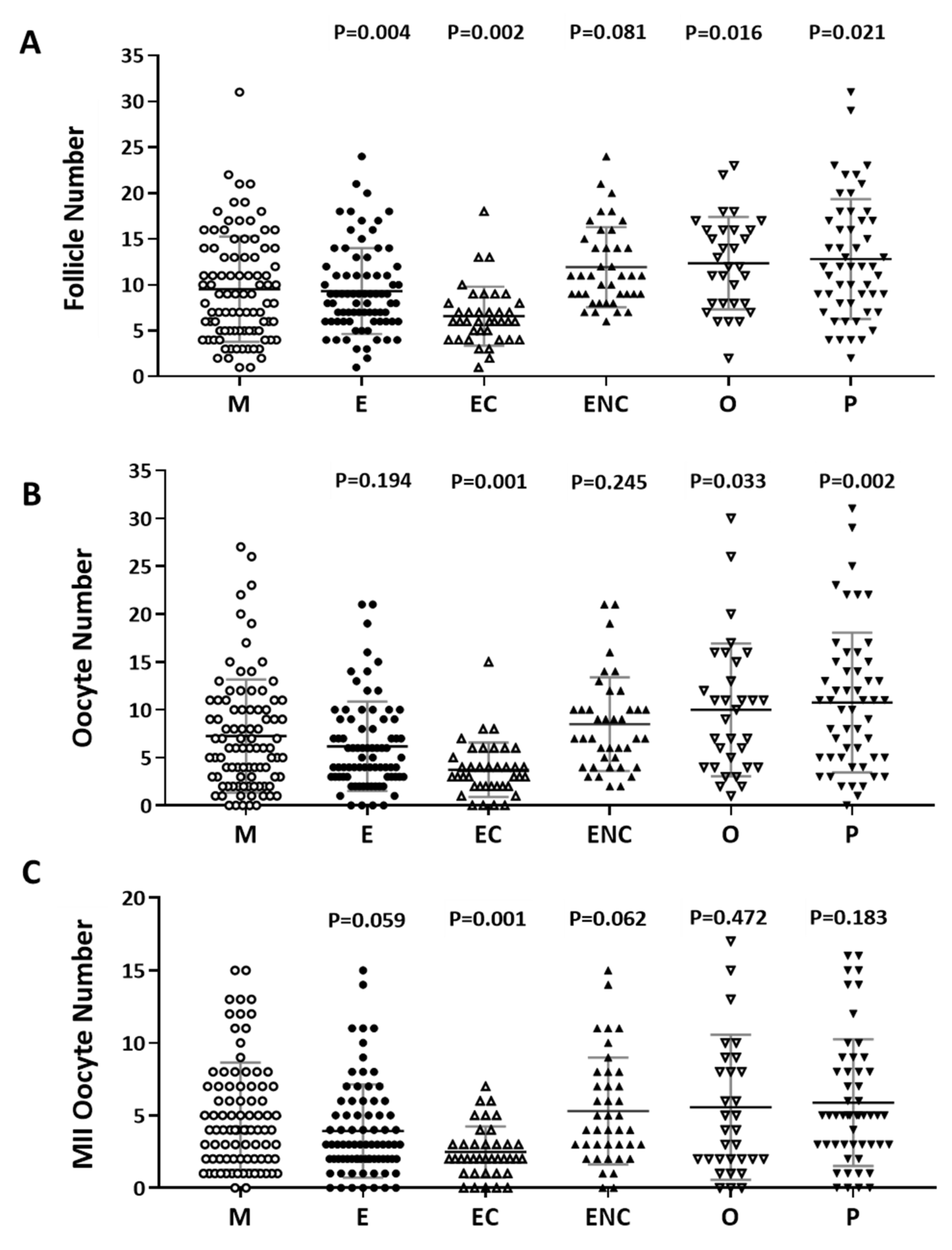
2.2. Decreased IVF Outcomes were Found in the Various Infertile Groups
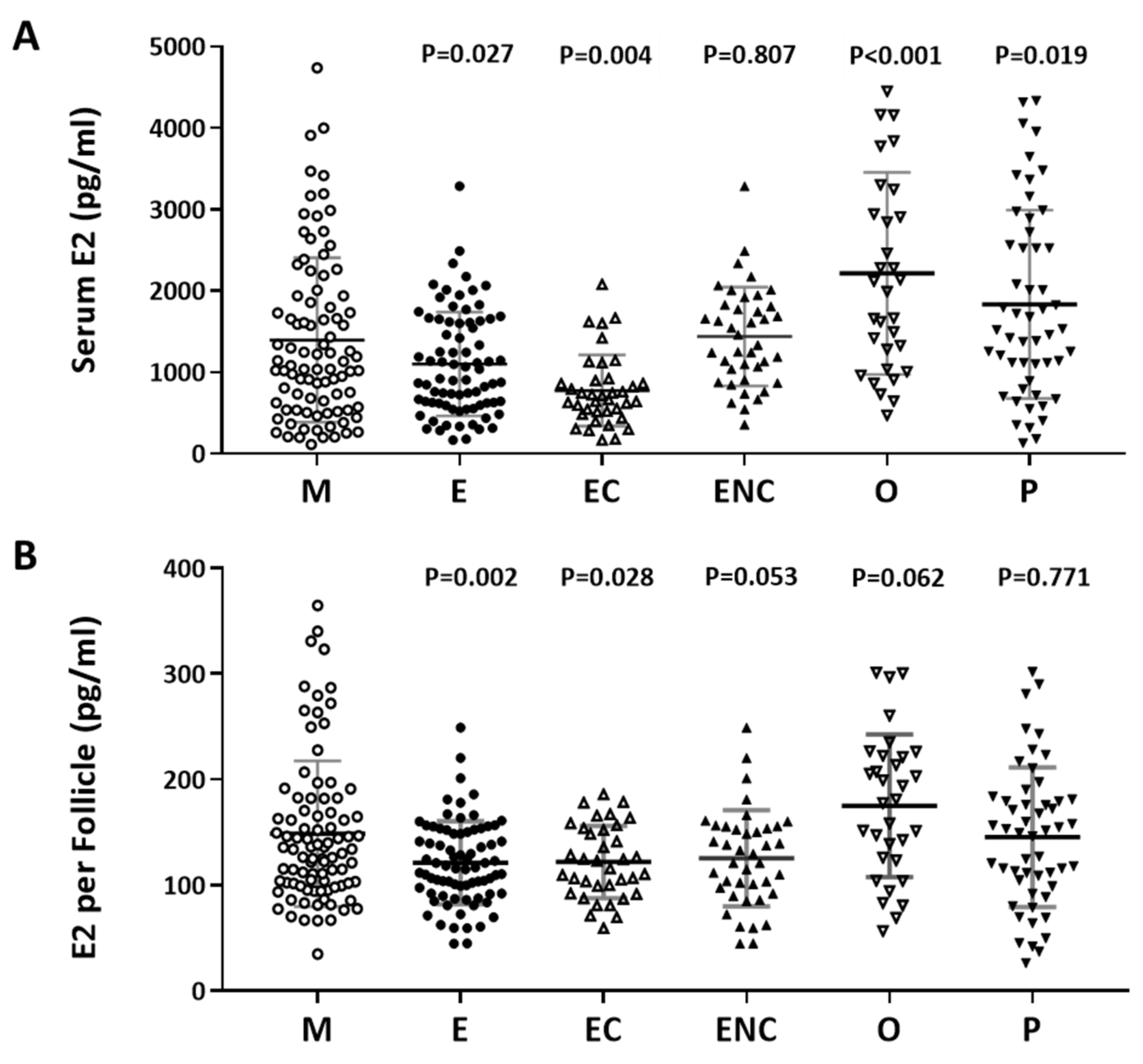
2.3. A Positive Correlation was Found between Serum Estradiol and Cycle Follicle Outcomes
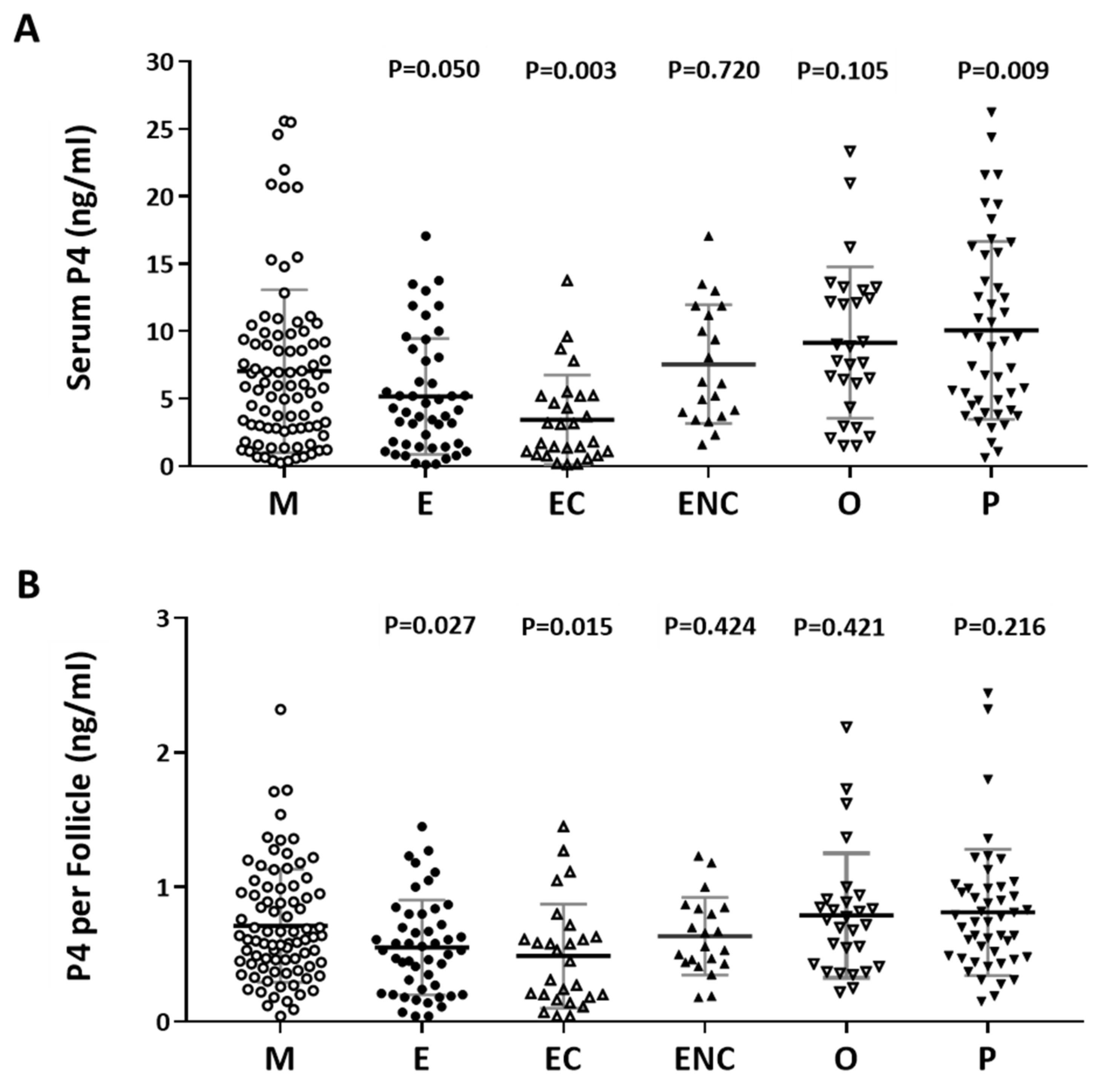
2.4. Serum Progesterone Was Positively Correlated with Follicle Cycle Outcomes
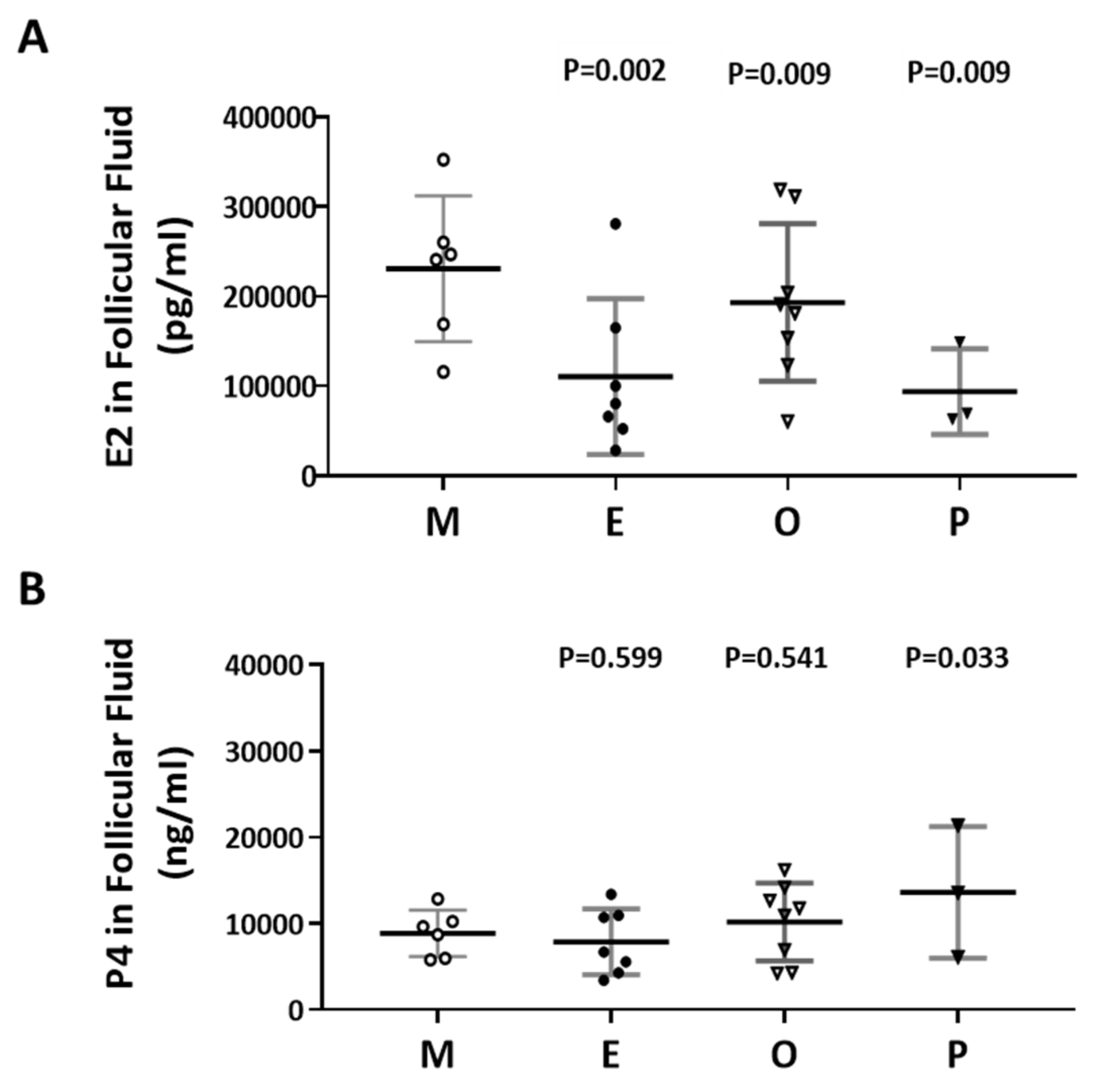
2.5. Decreased E2 Content in Follicular Fluids were Found in the Patient Group with Ovarian Endometrioma
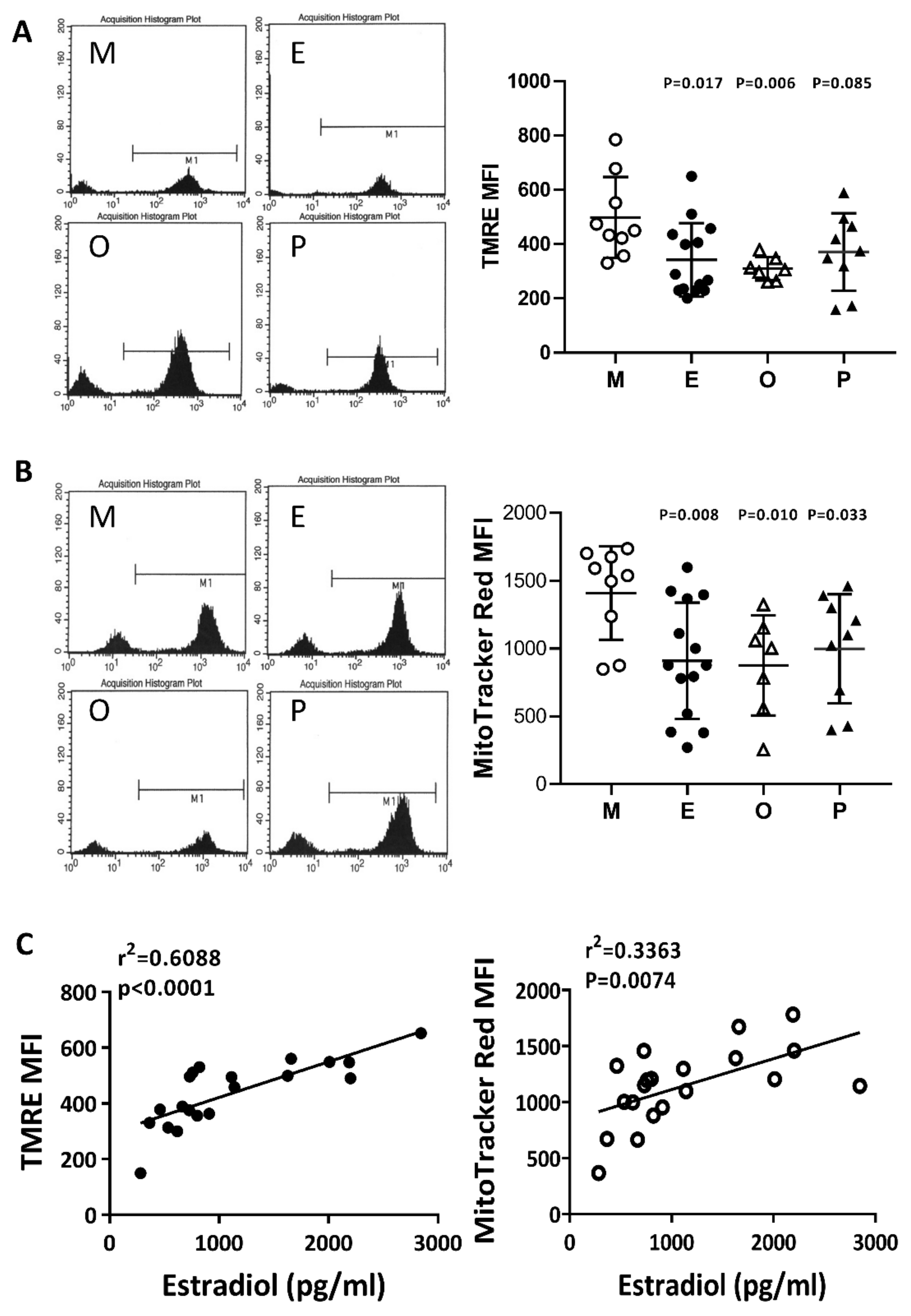
2.6. Impaired Mitochondrial Function in Cumulus Cells was Found in the Various Infertile Groups
2.7. Reduced Mitochondrial Mass in Cumulus Cells was Found in the Various Infertile Groups
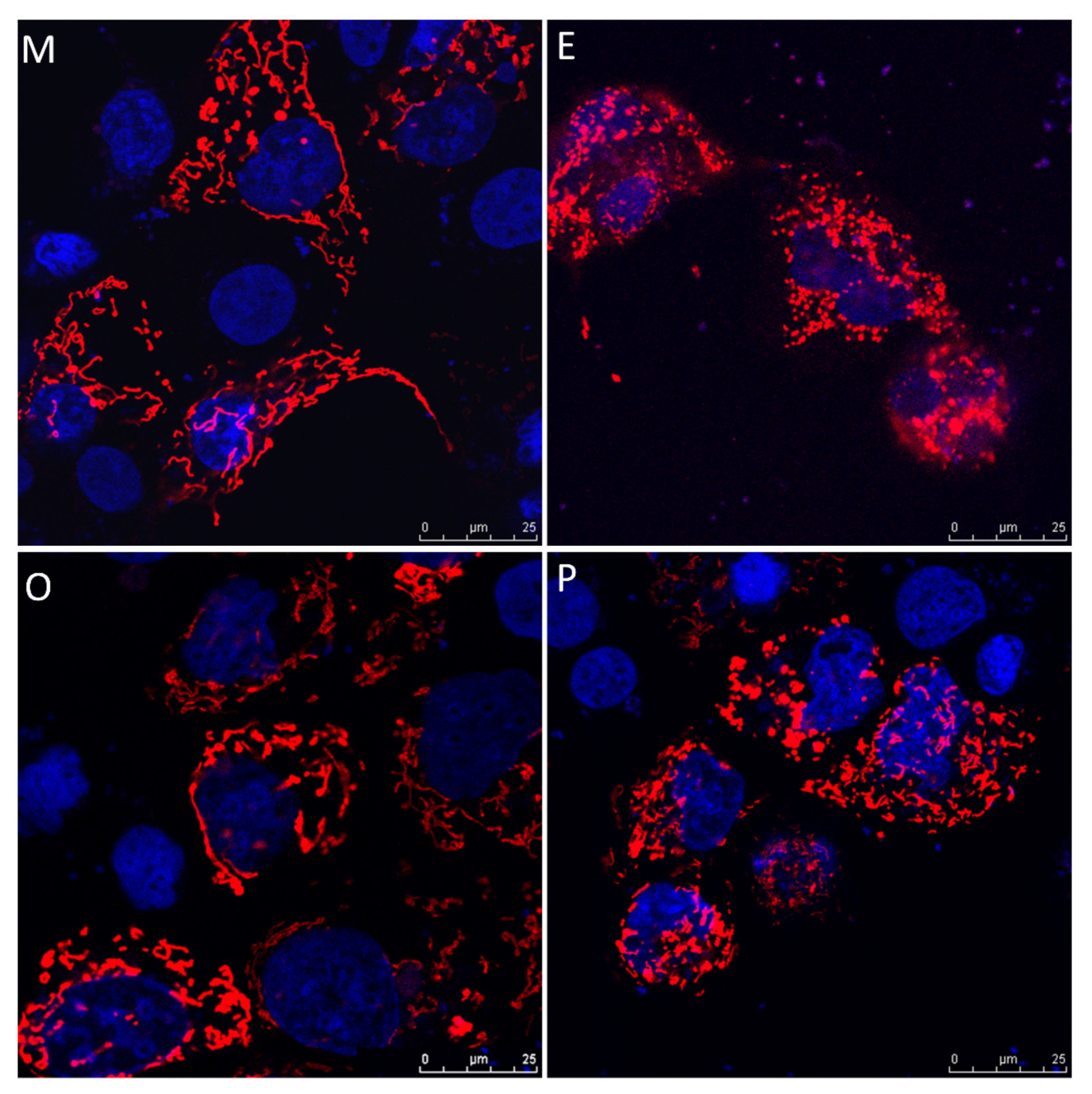
2.8. Induced Mitochondrial Fragmentation and Constrained Cumulus Cell Expansion were Found in the Endometriosis and PCOS Group
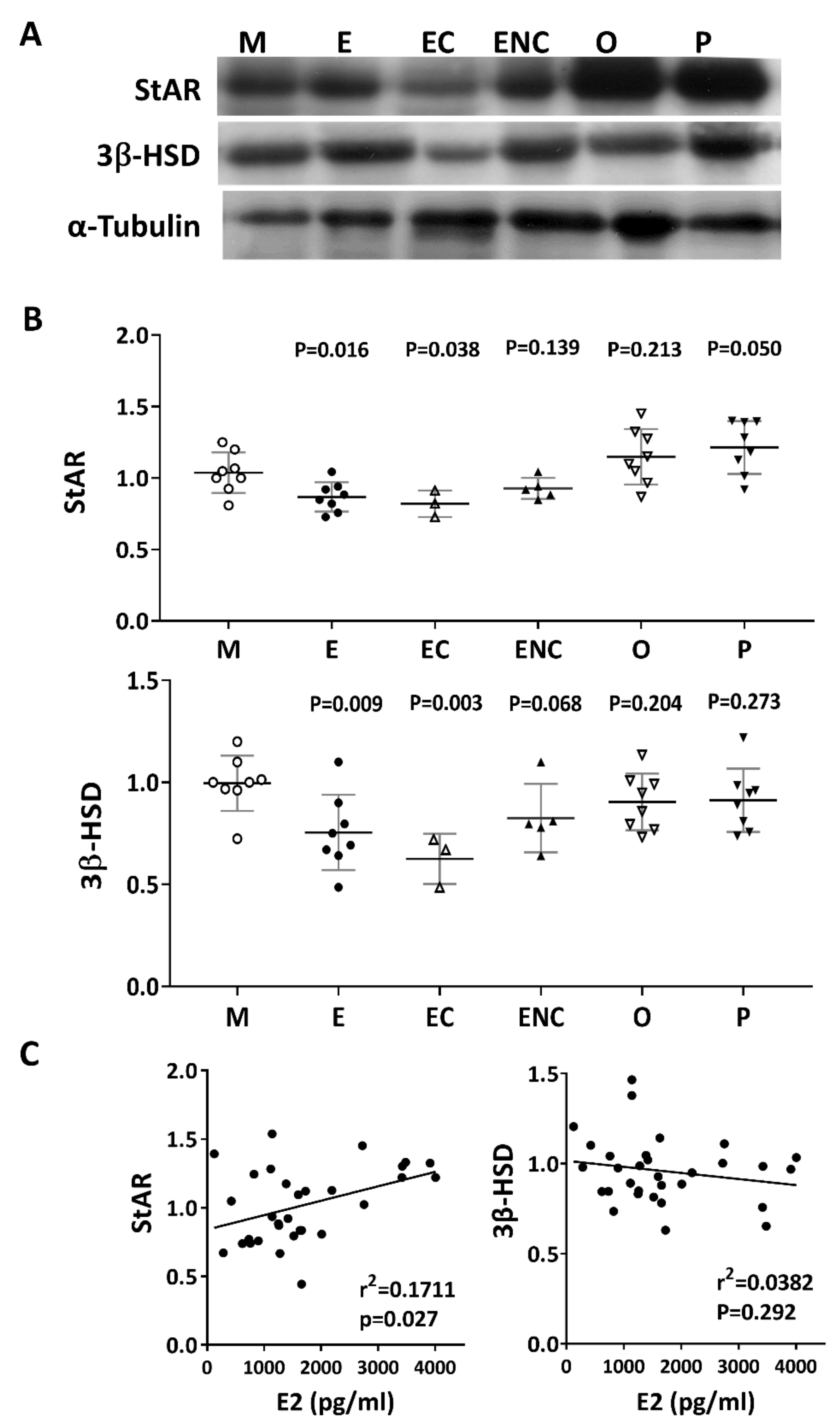
2.9. Lower Levels of Steroidogenic Enzymes were Observed in the Various Infertile Groups
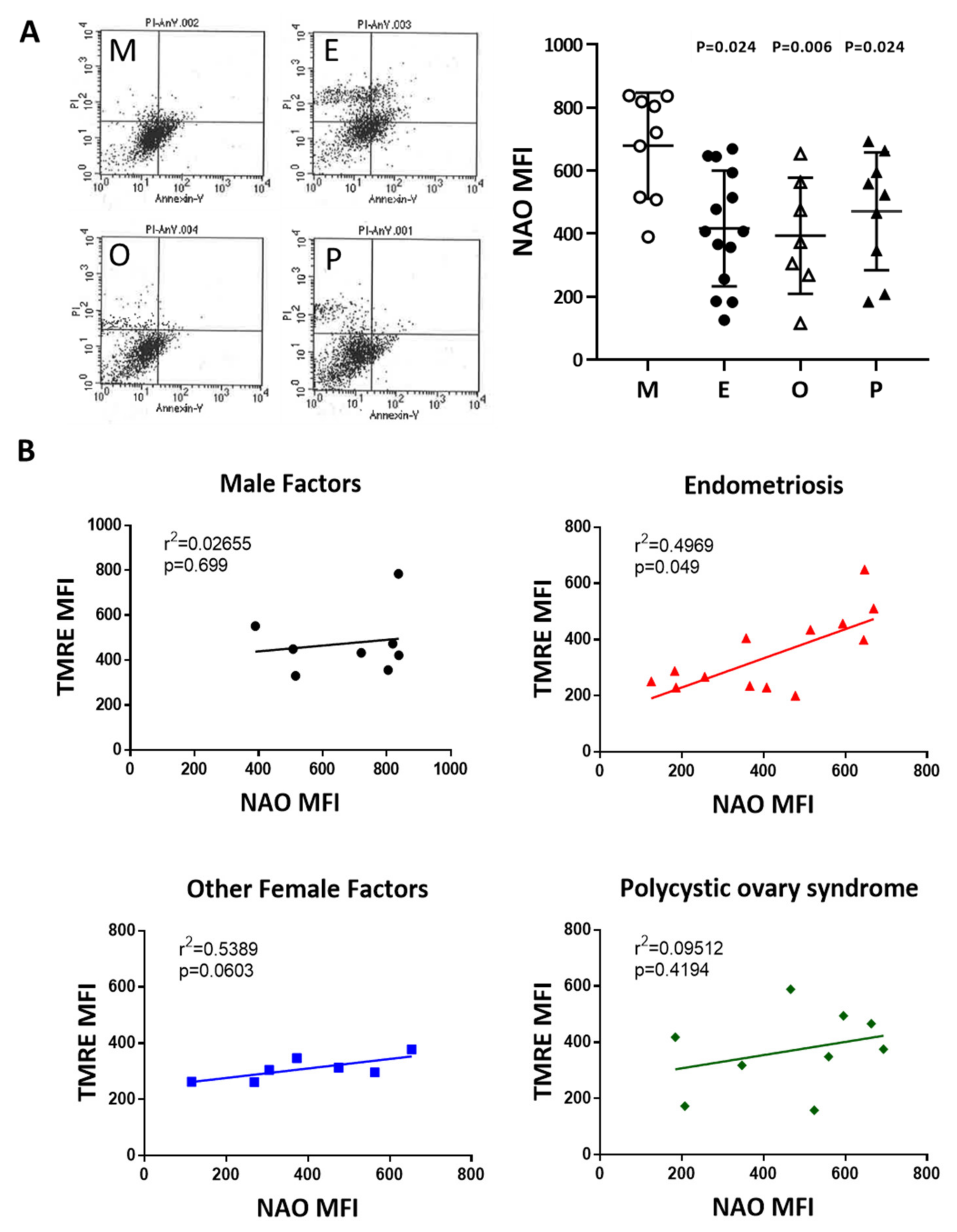
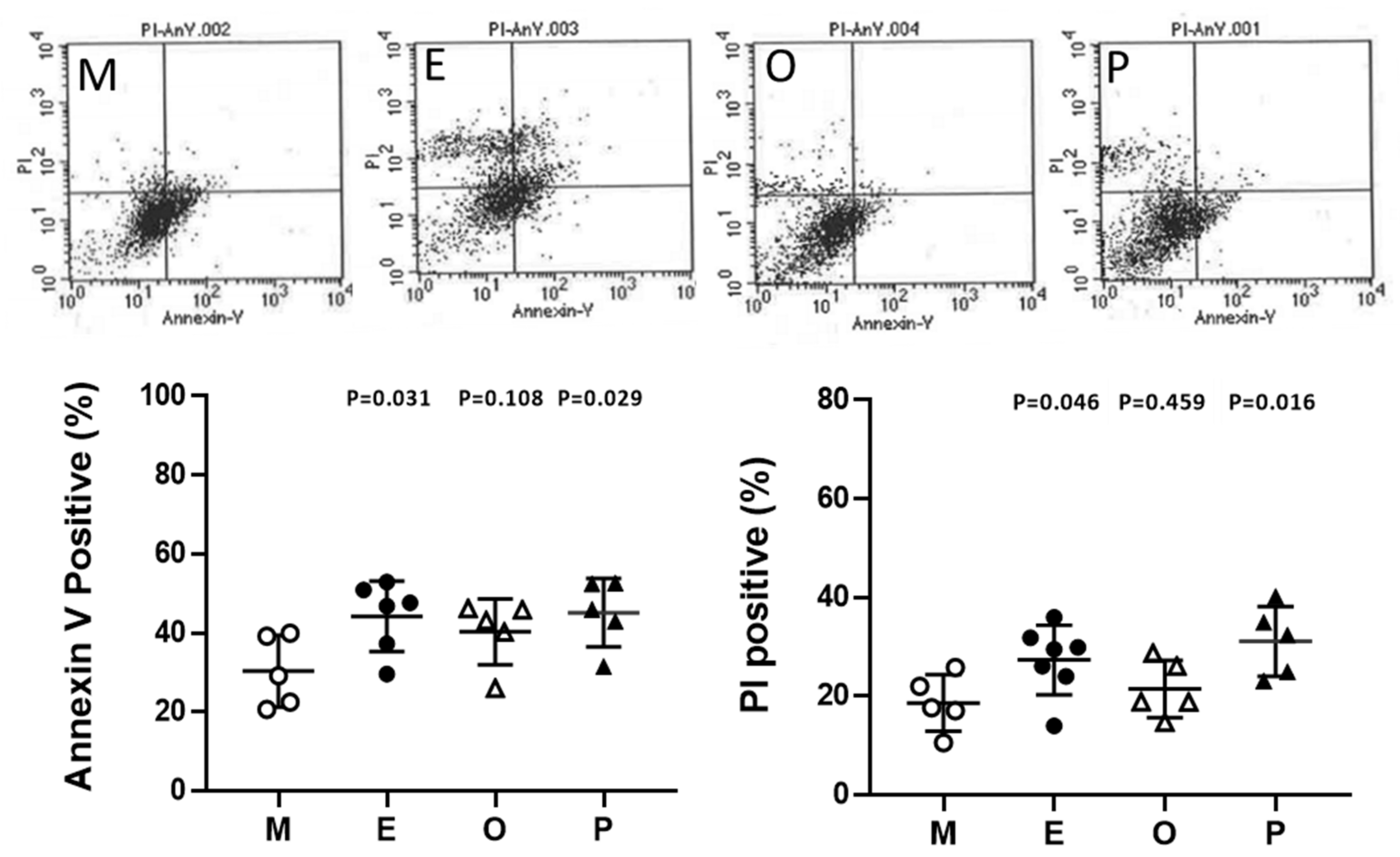
2.10. Induced Cumulus Cell Apoptosis was Found in the Patient Group with Ovarian Endometrioma and PCOS
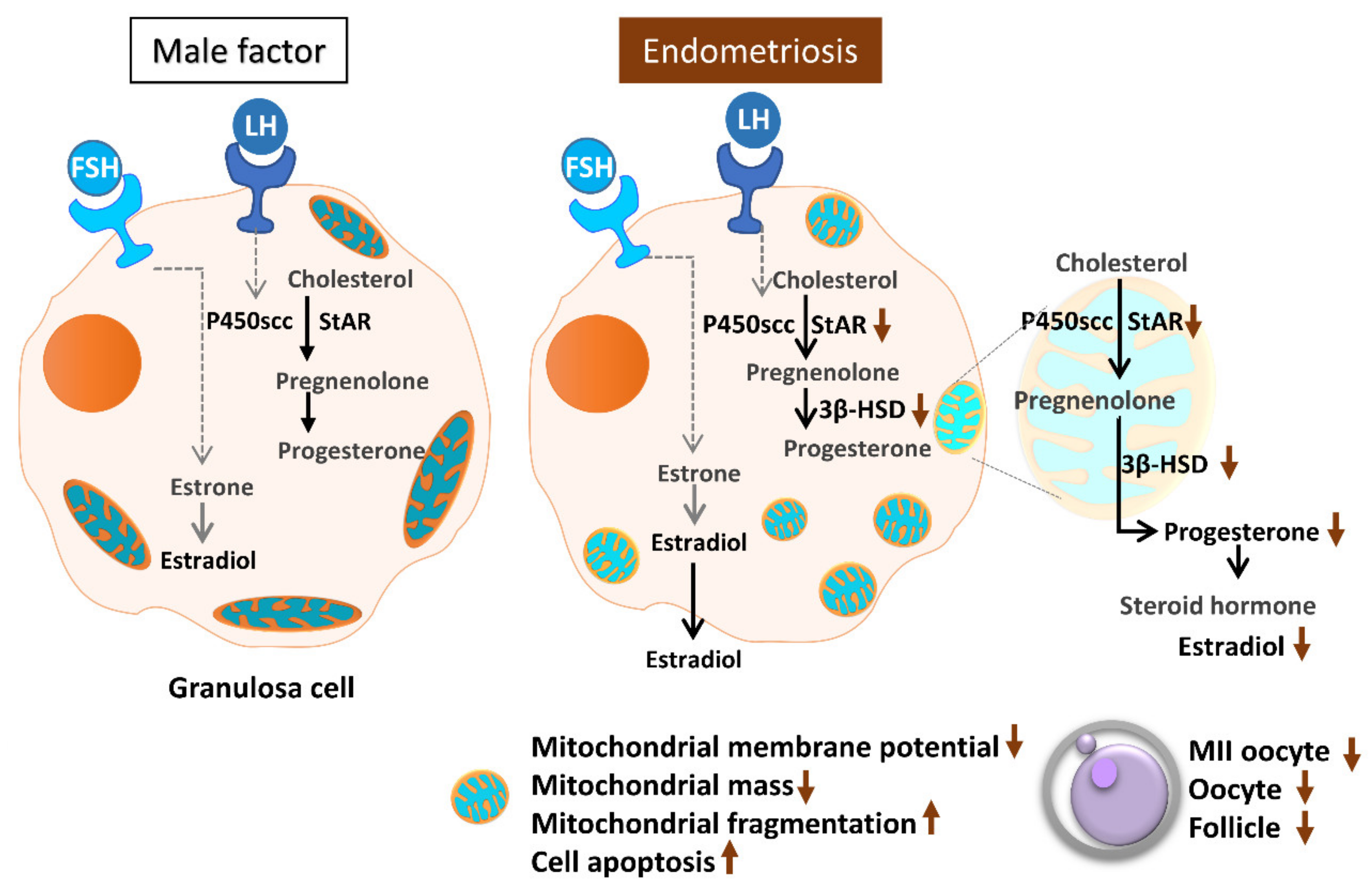
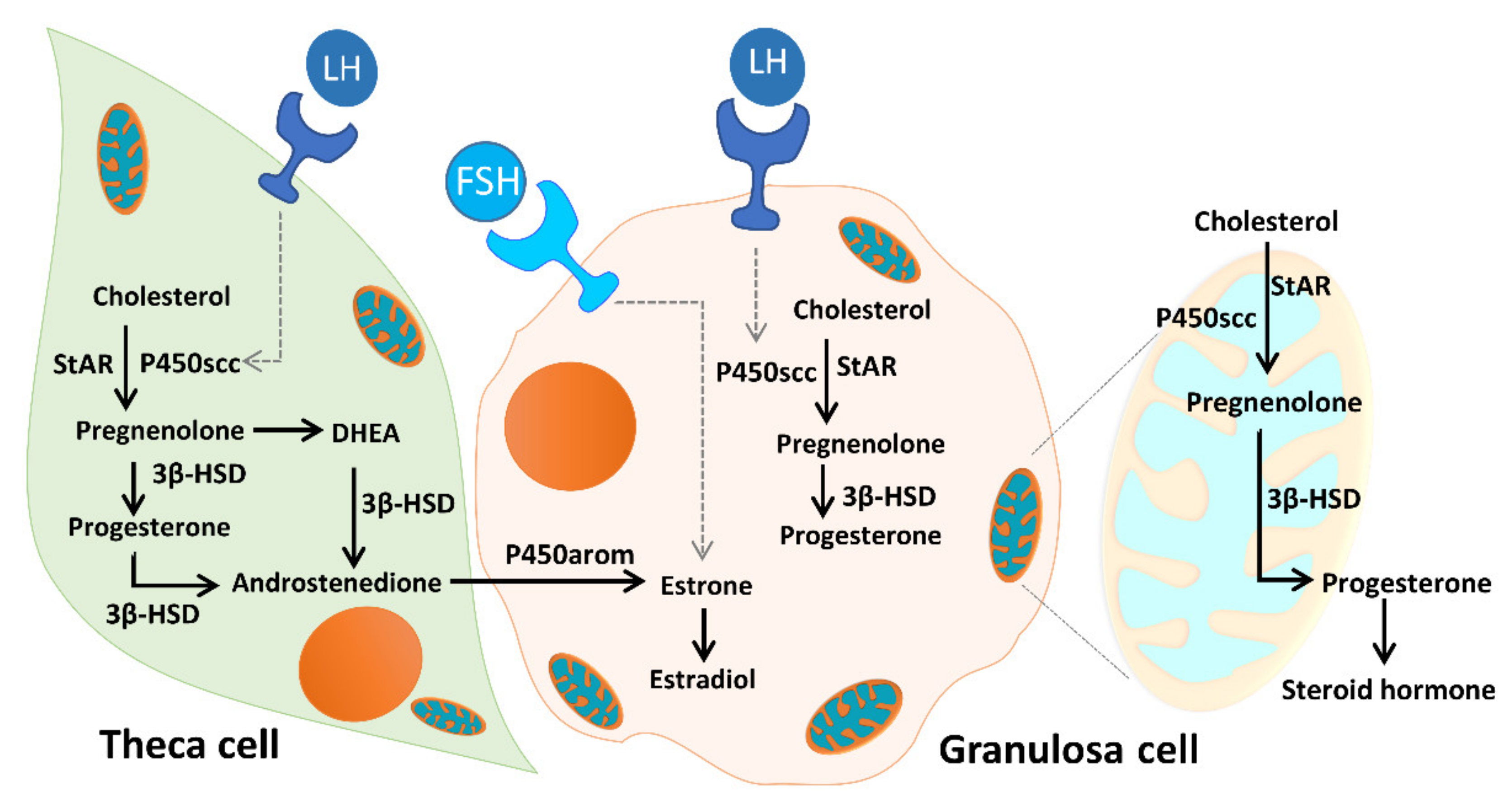
3. Discussion
4. Materials and Methods
4.1. Sample Collections
4.2. Assessment of Hormones
4.3. Measurement of Mitochondrial Membrane Potential and Mitochondrial Mass
4.4. Assessment of Mitochondrial Morphology
4.5. Protein Immunoblotting
4.6. Analysis of Apoptosis
4.7. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Practice Committee of American Society for Reproductive Medicine. Definitions of infertility and recurrent pregnancy loss: A committee opinion. Fertil. Steril. 2013, 99, 63. [Google Scholar] [CrossRef] [PubMed]
- Practice Committee of the American Society for Reproductive Medicine. Testing and interpreting measures of ovarian reserve: A committee opinion. Fertil. Steril. 2015, 103, e9–e17. [Google Scholar] [CrossRef] [PubMed]
- Committee on Gynecologic Practice. Committee opinion no. 618: Ovarian reserve testing. Obstet. Gynecol. 2015, 125, 268–273. [Google Scholar]
- Smith, S.; Pfeifer, S.M.; Collins, J.A. Diagnosis and management of female infertility. JAMA 2003, 290, 1767–1770. [Google Scholar] [CrossRef]
- Chattopadhyay, A.B.; Rath, S.K. Understanding male factor in infertility. Med. J. Armed Forces India 1999, 55, 181–182. [Google Scholar] [CrossRef]
- Niederberger, C. WHO manual for the standardized investigation, diagnosis and management of the infertile male. Urology 2001, 57, 208. [Google Scholar] [CrossRef]
- Farquhar, C. Endometriosis. BMJ 2007, 334, 249–253. [Google Scholar] [CrossRef]
- Brosens, I.; Benagiano, G. Endometriosis, a modern syndrome. Indian J. Med. Res. 2011, 133, 581–593. [Google Scholar]
- Lagana, A.S.; Garzon, S.; Gotte, M.; Vigano, P.; Franchi, M.; Ghezzi, F.; Martin, D.C. The pathogenesis of endometriosis: Molecular and cell biology insights. Int. J. Mol. Sci 2019, 20, 5615. [Google Scholar] [CrossRef]
- Vercellini, P.; Vigano, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef]
- Brosens, I.A.; Puttemans, P.J.; Deprest, J. The endoscopic localization of endometrial implants in the ovarian chocolate cyst. Fertil. Steril. 1994, 61, 1034–1038. [Google Scholar] [CrossRef]
- Zhang, J.; Bao, Y.; Zhou, X.; Zheng, L. Polycystic ovary syndrome and mitochondrial dysfunction. Reprod. Biol. Endocrinol. 2019, 17, 67. [Google Scholar] [CrossRef] [PubMed]
- Teede, H.J.; Misso, M.L.; Costello, M.F.; Dokras, A.; Laven, J.; Moran, L.; Piltonen, T.; Norman, R.J.; International, P.N. Recommendations from the international evidence-based guideline for the assessment and management of polycystic ovary syndrome. Fertil. Steril. 2018, 110, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Uyar, A.; Torrealday, S.; Seli, E. Cumulus and granulosa cell markers of oocyte and embryo quality. Fertil. Steril. 2013, 99, 979–997. [Google Scholar] [CrossRef]
- Wyndham, N.; Marin Figueira, P.G.; Patrizio, P. A persistent misperception: Assisted reproductive technology can reverse the aged biological clock. Fertil. Steril. 2012, 97, 1044–1047. [Google Scholar] [CrossRef]
- Cakmak, H.; Franciosi, F.; Zamah, A.M.; Cedars, M.I.; Conti, M. Dynamic secretion during meiotic reentry integrates the function of the oocyte and cumulus cells. Proc. Natl. Acad. Sci. USA 2016, 113, 2424–2429. [Google Scholar] [CrossRef]
- Hillier, S.G.; Whitelaw, P.F.; Smyth, C.D. Follicular oestrogen synthesis: The two-cell, two-gonadotrophin model revisited. Mol. Cell. Endocrinol. 1994, 100, 51–54. [Google Scholar] [CrossRef]
- Ting, A.Y.; Xu, J.; Stouffer, R.L. Differential effects of estrogen and progesterone on development of primate secondary follicles in a steroid-depleted milieu in vitro. Hum. Reprod. 2015, 30, 1907–1917. [Google Scholar] [CrossRef]
- Hamel, M.; Dufort, I.; Robert, C.; Gravel, C.; Leveille, M.C.; Leader, A.; Sirard, M.A. Identification of differentially expressed markers in human follicular cells associated with competent oocytes. Hum. Reprod. 2008, 23, 1118–1127. [Google Scholar] [CrossRef]
- Wathlet, S.; Adriaenssens, T.; Segers, I.; Verheyen, G.; Janssens, R.; Coucke, W.; Devroey, P.; Smitz, J. New candidate genes to predict pregnancy outcome in single embryo transfer cycles when using cumulus cell gene expression. Fertil. Steril. 2012, 98, 432–439.e4. [Google Scholar] [CrossRef]
- Feuerstein, P.; Cadoret, V.; Dalbies-Tran, R.; Guerif, F.; Bidault, R.; Royere, D. Gene expression in human cumulus cells: One approach to oocyte competence. Hum. Reprod. 2007, 22, 3069–3077. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Disorders in the initial steps of steroid hormone synthesis. J. Steroid. Biochem. Mol. Biol. 2017, 165, 18–37. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Steroid hormone synthesis in mitochondria. Mol. Cell. Endocrinol. 2013, 379, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.A.; Shankara, T.; Janus, P.; Buck, S.; Diemer, T.; Hales, K.H.; Hales, D.B. Energized, polarized, and actively respiring mitochondria are required for acute Leydig cell steroidogenesis. Endocrinology 2006, 147, 3924–3935. [Google Scholar] [CrossRef]
- Artemenko, I.P.; Zhao, D.; Hales, D.B.; Hales, K.H.; Jefcoate, C.R. Mitochondrial processing of newly synthesized steroidogenic acute regulatory protein (StAR), but not total StAR, mediates cholesterol transfer to cytochrome P450 side chain cleavage enzyme in adrenal cells. J. Biol. Chem. 2001, 276, 46583–46596. [Google Scholar] [CrossRef]
- Au, H.K.; Lin, S.H.; Huang, S.Y.; Yeh, T.S.; Tzeng, C.R.; Hsieh, R.H. Deleted mitochondrial DNA in human luteinized granulosa cells. Ann. N. Y. Acad. Sci. 2005, 1042, 136–141. [Google Scholar] [CrossRef]
- Von Mengden, L.; Klamt, F.; Smitz, J. Redox biology of human cumulus cells: Basic concepts, impact on oocyte quality, and potential clinical use. Antioxid. Redox Signal. 2020, 32, 522–535. [Google Scholar] [CrossRef]
- Karuputhula, N.B.; Chattopadhyay, R.; Chakravarty, B.; Chaudhury, K. Oxidative status in granulosa cells of infertile women undergoing IVF. Syst. Biol. Reprod. Med. 2013, 59, 91–98. [Google Scholar] [CrossRef]
- Hsu, A.L.; Townsend, P.M.; Oehninger, S.; Castora, F.J. Endometriosis may be associated with mitochondrial dysfunction in cumulus cells from subjects undergoing in vitro fertilization-intracytoplasmic sperm injection, as reflected by decreased adenosine triphosphate production. Fertil. Steril. 2015, 103, 347–352.e1. [Google Scholar] [CrossRef]
- Hoshino, Y. Updating the markers for oocyte quality evaluation: Intracellular temperature as a new index. Reprod. Med. Biol. 2018, 17, 434–441. [Google Scholar] [CrossRef]
- Tanghe, S.; Van Soom, A.; Nauwynck, H.; Coryn, M.; de Kruif, A. Minireview: Functions of the cumulus oophorus during oocyte maturation, ovulation, and fertilization. Mol. Reprod. Dev. 2002, 61, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wells, D. The human oocyte and cumulus cells relationship: New insights from the cumulus cell transcriptome. Mol. Hum. Reprod. 2010, 16, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Boucret, L.; Chao de la Barca, J.M.; Moriniere, C.; Desquiret, V.; Ferre-L’Hotellier, V.; Descamps, P.; Marcaillou, C.; Reynier, P.; Procaccio, V.; May-Panloup, P. Relationship between diminished ovarian reserve and mitochondrial biogenesis in cumulus cells. Hum. Reprod. 2015, 30, 1653–1664. [Google Scholar] [CrossRef] [PubMed]
- Senapati, S.; Sammel, M.D.; Morse, C.; Barnhart, K.T. Impact of endometriosis on in vitro fertilization outcomes: An evaluation of the Society for Assisted Reproductive Technologies Database. Fertil. Steril. 2016, 106, 164–171.e1. [Google Scholar] [CrossRef]
- Harb, H.M.; Gallos, I.D.; Chu, J.; Harb, M.; Coomarasamy, A. The effect of endometriosis on in vitro fertilization outcome: A systematic review and meta-analysis. BJOG 2013, 120, 1308–1320. [Google Scholar] [CrossRef]
- Coccia, M.E.; Rizzello, F.; Mariani, G.; Bulletti, C.; Palagiano, A.; Scarselli, G. Impact of endometriosis on in vitro fertilization and embryo transfer cycles in young women: A stage-dependent interference. Acta Obstet. Gynecol. Scand. 2011, 90, 1232–1238. [Google Scholar] [CrossRef]
- Suzuki, T.; Izumi, S.; Matsubayashi, H.; Awaji, H.; Yoshikata, K.; Makino, T. Impact of ovarian endometrioma on oocytes and pregnancy outcome in in vitro fertilization. Fertil. Steril. 2005, 83, 908–913. [Google Scholar] [CrossRef]
- Opoien, H.K.; Fedorcsak, P.; Omland, A.K.; Abyholm, T.; Bjercke, S.; Ertzeid, G.; Oldereid, N.; Mellembakken, J.R.; Tanbo, T. In vitro fertilization is a successful treatment in endometriosis-associated infertility. Fertil. Steril. 2012, 97, 912–918. [Google Scholar] [CrossRef]
- Zamah, A.M.; Hassis, M.E.; Albertolle, M.E.; Williams, K.E. Proteomic analysis of human follicular fluid from fertile women. Clin. Proteomics 2015, 12, 5. [Google Scholar] [CrossRef]
- Rodgers, R.J.; Irving-Rodgers, H.F. Formation of the ovarian follicular antrum and follicular fluid. Biol. Reprod. 2010, 82, 1021–1029. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Vanni, V.S.; Bartiromo, L.; Papaleo, E.; Zilberberg, E.; Candiani, M.; Orvieto, R.; Vigano, P. Is the oocyte quality affected by endometriosis? A review of the literature. J. Ovarian Res. 2017, 10, 43. [Google Scholar] [CrossRef]
- Ishihara, Y.; Takemoto, T.; Ishida, A.; Yamazaki, T. Protective actions of 17beta-estradiol and progesterone on oxidative neuronal injury induced by organometallic compounds. Oxid. Med. Cell. Longev. 2015, 2015, 343706. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.D.; Zamah, A.M.; Shen, S.; McCulloch, C.; Cedars, M.I.; Rosen, M.P. Follicular fluid steroid hormone levels are associated with fertilization outcome after intracytoplasmic sperm injection. Fertil. Steril. 2010, 94, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Carpintero, N.L.; Suarez, O.A.; Mangas, C.C.; Varea, C.G.; Rioja, R.G. Follicular steroid hormones as markers of oocyte quality and oocyte development potential. J. Hum. Reprod. Sci. 2014, 7, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.W., Jr.; Acosta, A.; Andrews, M.C.; Garcia, J.E.; Jones, G.S.; Mantzavinos, T.; McDowell, J.; Sandow, B.; Veeck, L.; Whibley, T.; et al. The importance of the follicular phase to success and failure in in vitro fertilization. Fertil. Steril. 1983, 40, 317–321. [Google Scholar] [CrossRef]
- Fisher, S.; Grin, A.; Paltoo, A.; Shapiro, H.M. Falling estradiol levels as a result of intentional reduction in gonadotrophin dose are not associated with poor IVF outcomes, whereas spontaneously falling estradiol levels result in low clinical pregnancy rates. Hum. Reprod. 2005, 20, 84–88. [Google Scholar] [CrossRef]
- Segawa, T.; Teramoto, S.; Omi, K.; Miyauchi, O.; Watanabe, Y.; Osada, H. Changes in estrone and estradiol levels during follicle development: A retrospective large-scale study. Reprod. Biol. Endocrinol. 2015, 13, 54. [Google Scholar] [CrossRef]
- Nebert, D.W.; Wikvall, K.; Miller, W.L. Human cytochromes P450 in health and disease. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120431. [Google Scholar] [CrossRef]
- Prasad, M.; Thomas, J.L.; Whittal, R.M.; Bose, H.S. Mitochondrial 3beta-hydroxysteroid dehydrogenase enzyme activity requires reversible pH-dependent conformational change at the intermembrane space. J. Biol. Chem. 2012, 287, 9534–9546. [Google Scholar] [CrossRef]
- Castillo, A.F.; Orlando, U.; Helfenberger, K.E.; Poderoso, C.; Podesta, E.J. The role of mitochondrial fusion and StAR phosphorylation in the regulation of StAR activity and steroidogenesis. Mol. Cell. Endocrinol. 2015, 408, 73–79. [Google Scholar] [CrossRef]
- Regan, S.L.P.; Knight, P.G.; Yovich, J.L.; Leung, Y.; Arfuso, F.; Dharmarajan, A. Granulosa cell apoptosis in the ovarian follicle-a changing view. Front. Endocrinol. 2018, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Schreier, S.; Sawaisorn, P.; Udomsangpetch, R.; Triampo, W. Advances in rare cell isolation: An optimization and evaluation study. J. Transl. Med. 2017, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Vigone, G.; Merico, V.; Redi, C.A.; Mazzini, G.; Garagna, S.; Zuccotti, M. FSH and LH receptors are differentially expressed in cumulus cells surrounding developmentally competent and incompetent mouse fully grown antral oocytes. Reprod. Fertil. Dev. 2015, 27, 497–503. [Google Scholar] [CrossRef] [PubMed]












© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sreerangaraja Urs, D.B.; Wu, W.-H.; Komrskova, K.; Postlerova, P.; Lin, Y.-F.; Tzeng, C.-R.; Kao, S.-H. Mitochondrial Function in Modulating Human Granulosa Cell Steroidogenesis and Female Fertility. Int. J. Mol. Sci. 2020, 21, 3592. https://doi.org/10.3390/ijms21103592
Sreerangaraja Urs DB, Wu W-H, Komrskova K, Postlerova P, Lin Y-F, Tzeng C-R, Kao S-H. Mitochondrial Function in Modulating Human Granulosa Cell Steroidogenesis and Female Fertility. International Journal of Molecular Sciences. 2020; 21(10):3592. https://doi.org/10.3390/ijms21103592
Chicago/Turabian StyleSreerangaraja Urs, Dilip Bhargava, Wen-Han Wu, Katerina Komrskova, Pavla Postlerova, Yung-Feng Lin, Chii-Ruey Tzeng, and Shu-Huei Kao. 2020. "Mitochondrial Function in Modulating Human Granulosa Cell Steroidogenesis and Female Fertility" International Journal of Molecular Sciences 21, no. 10: 3592. https://doi.org/10.3390/ijms21103592
APA StyleSreerangaraja Urs, D. B., Wu, W.-H., Komrskova, K., Postlerova, P., Lin, Y.-F., Tzeng, C.-R., & Kao, S.-H. (2020). Mitochondrial Function in Modulating Human Granulosa Cell Steroidogenesis and Female Fertility. International Journal of Molecular Sciences, 21(10), 3592. https://doi.org/10.3390/ijms21103592