
Antimicrobial Activity and Chemical Constitution of the Crude, Phenolic-Rich Extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization of Isolated CPRE
2.1.1. Total Anthocyanin, Total Phenolic and Total Flavonoid Contents
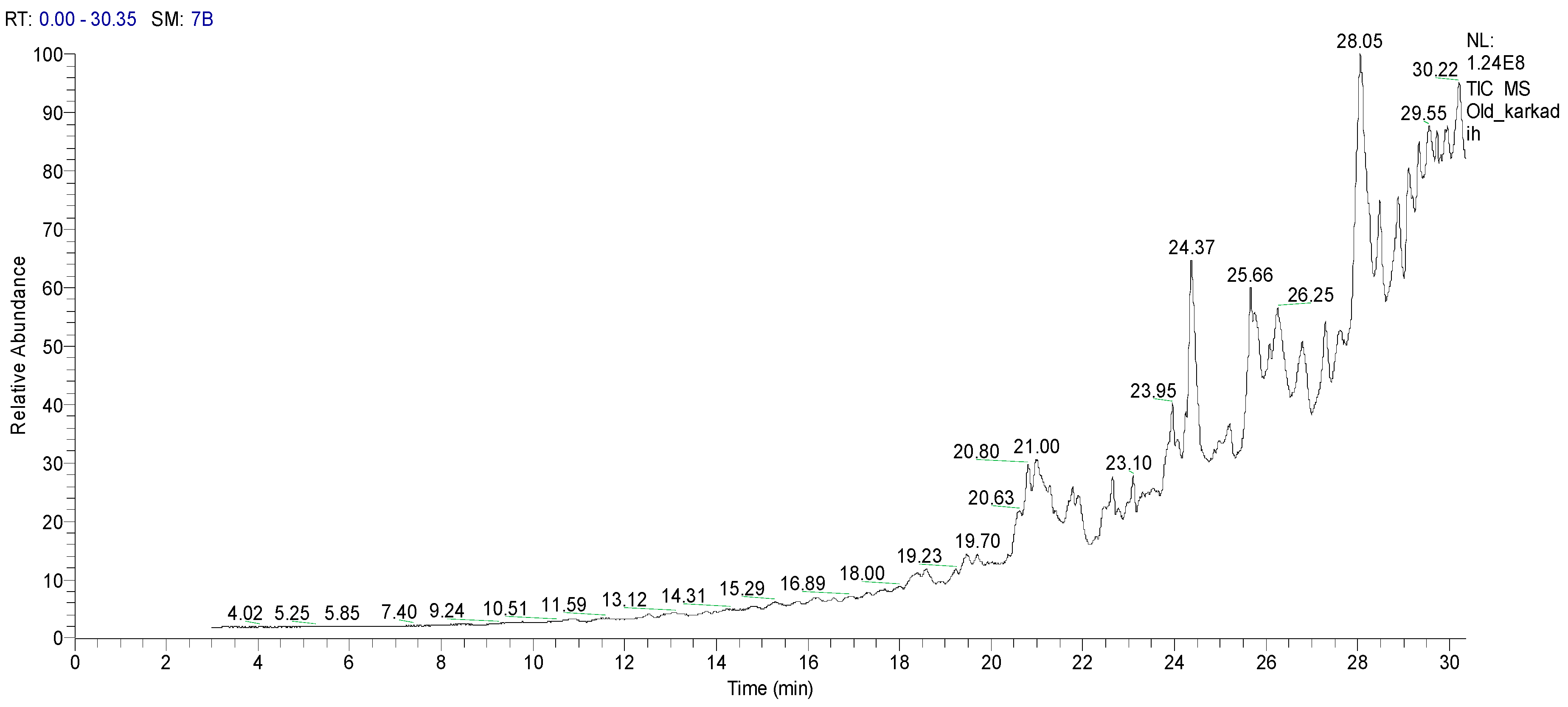
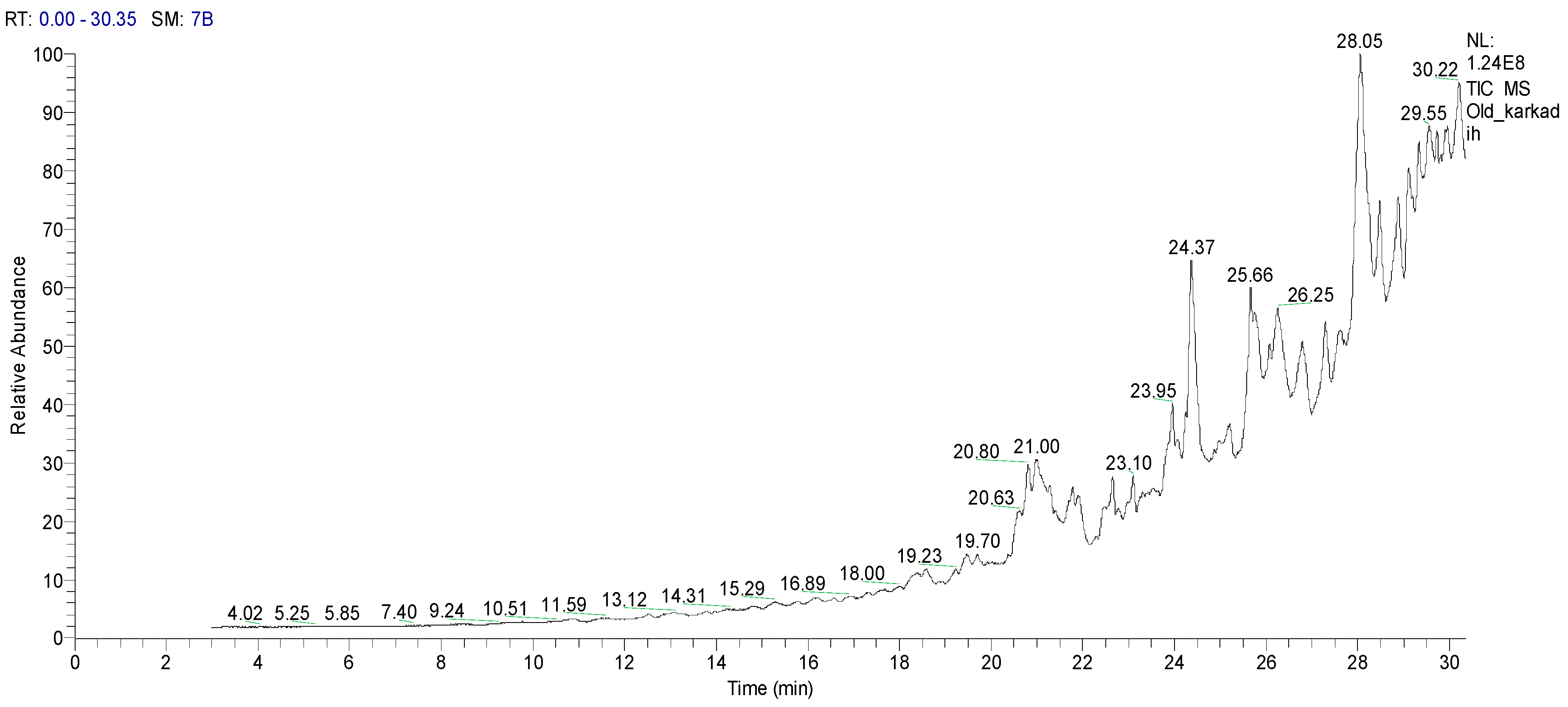
2.1.2. Gas Chromatography-Mass Spectrometry (GC–MS) Analysis of HCPRE
2.2. Antimicrobial Activity of Crude Phenolic Rich Extract (CPRE) (2000 µg/mL) against Pathogenic Bacteria
2.3. Minimum Inhibitory Concentrations (MICs) Values of HCPRE and B. oleracea Pigments against Bacteria
2.4. Antifungal Activity of HCPRE against Pathogenic Fungi and MIC Values
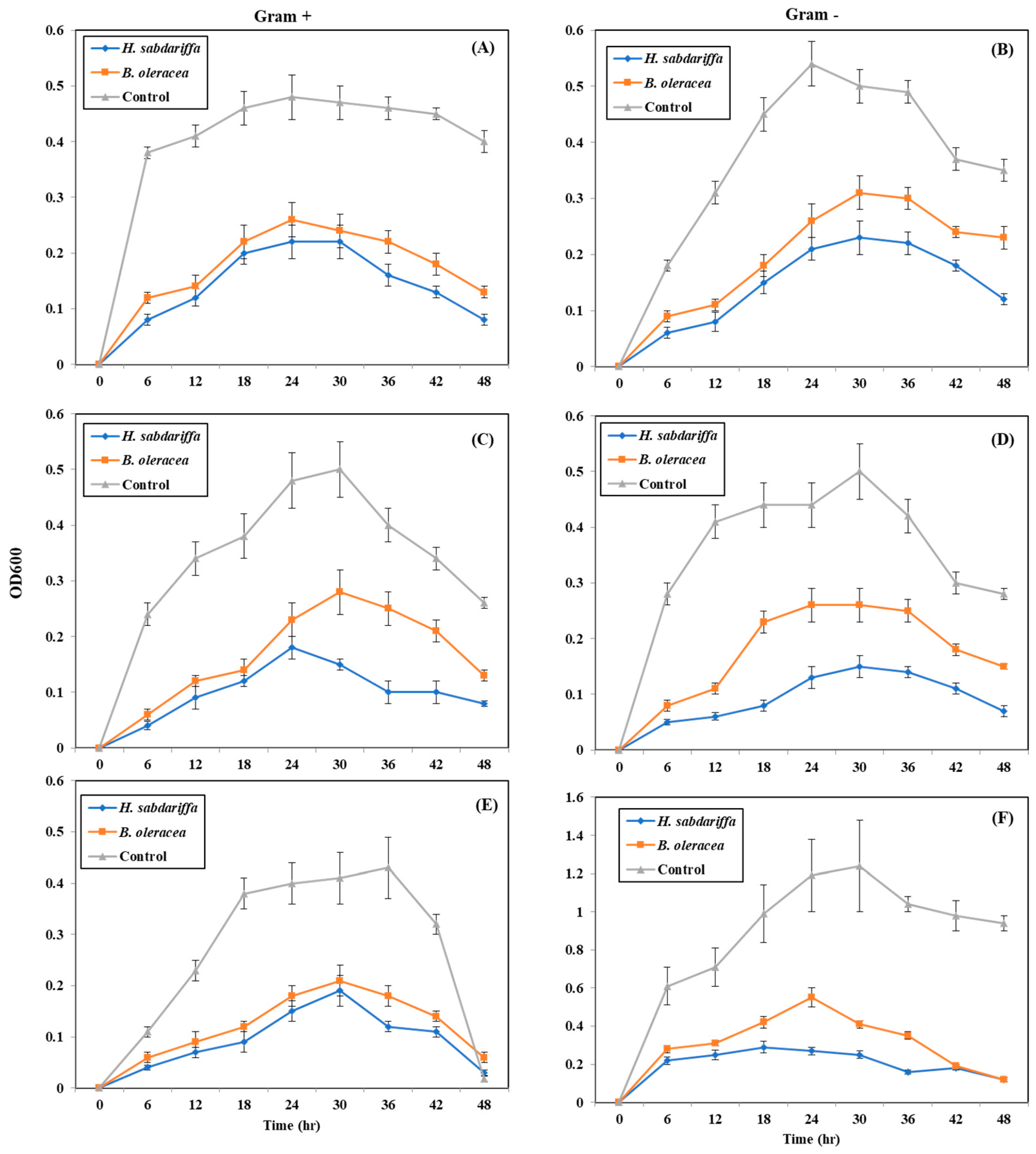
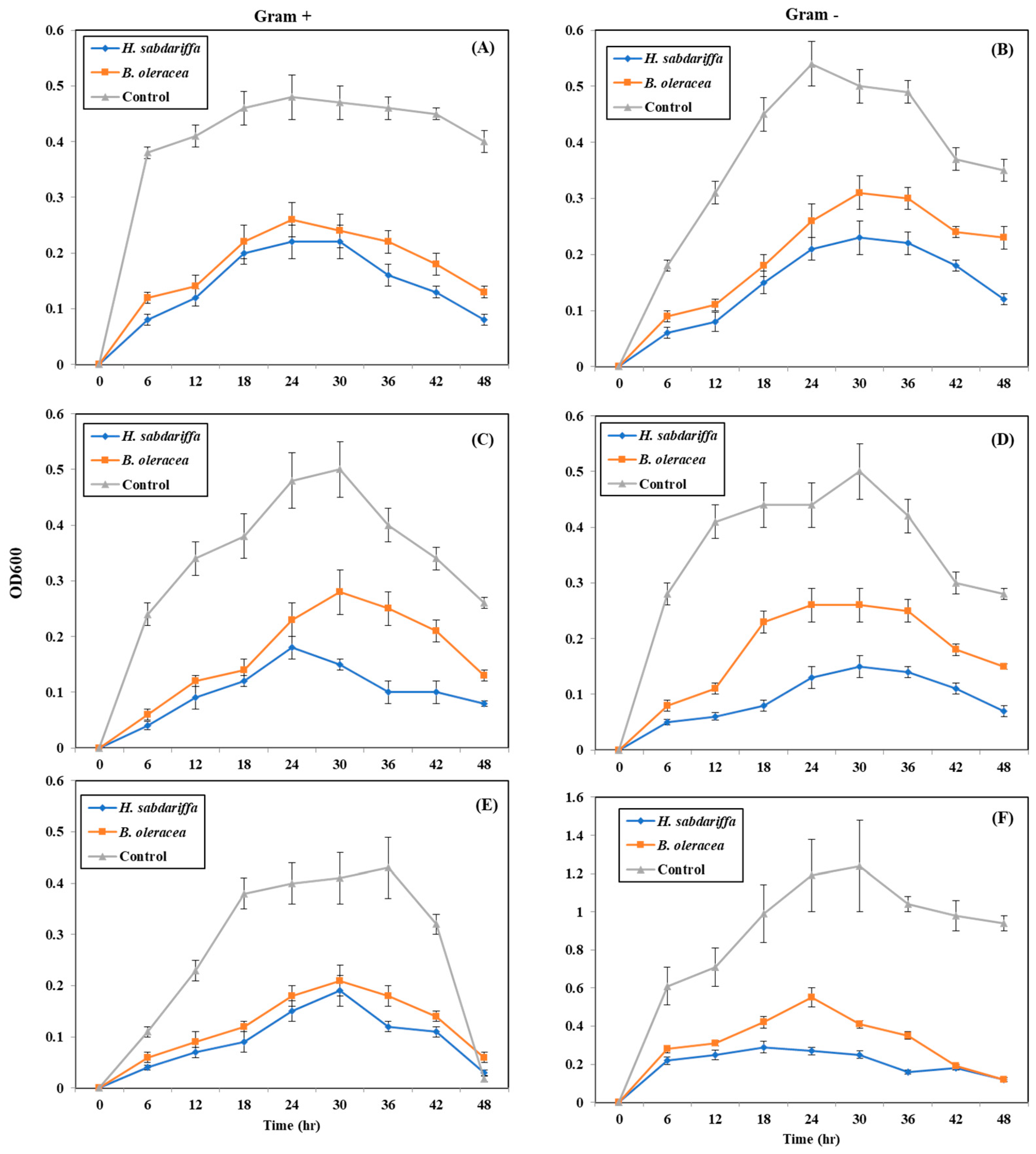
2.5. Quantitative Inhibition of Pathogenic Bacteria by Plant Pigments (Bacterial Growth Curve)
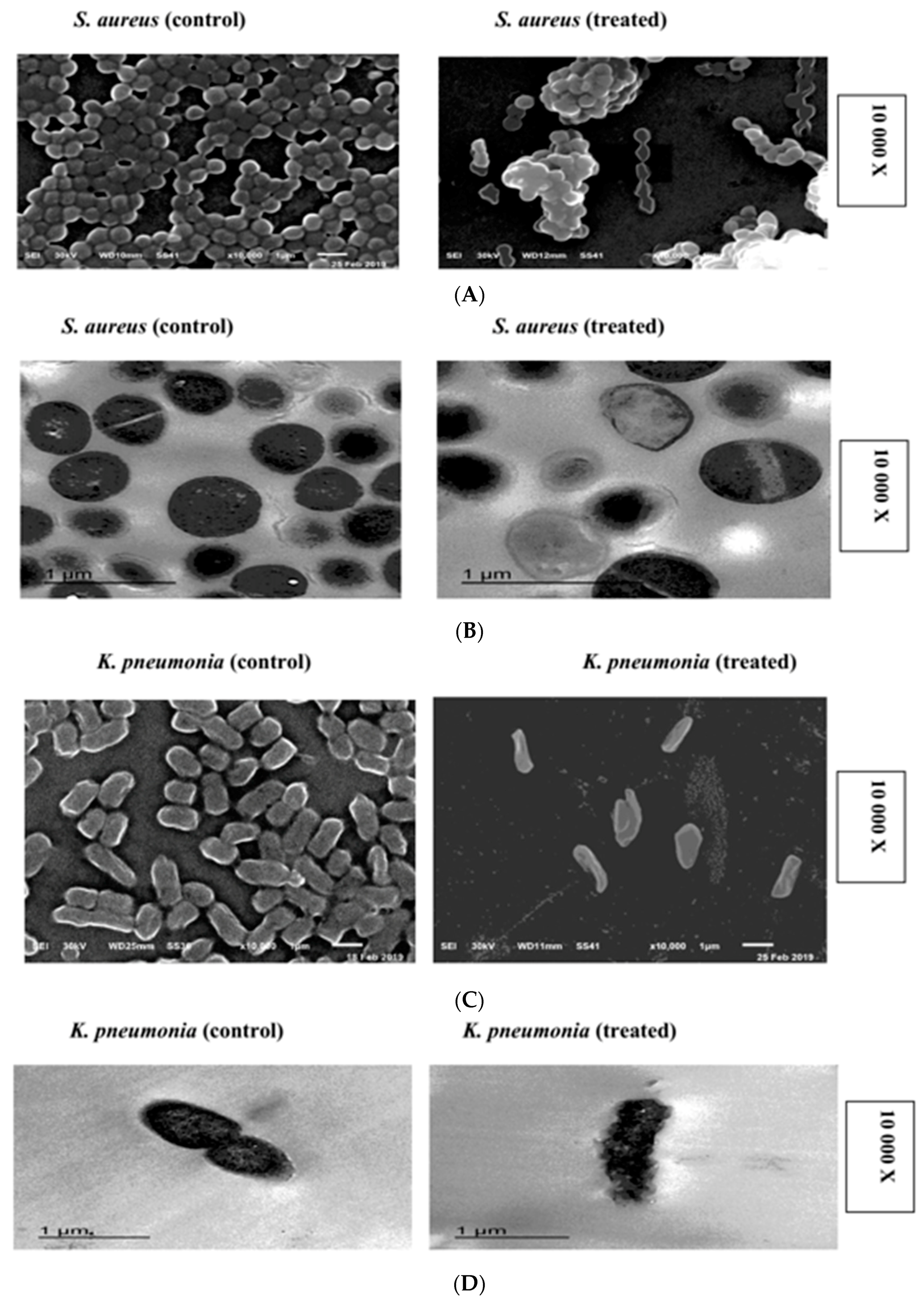
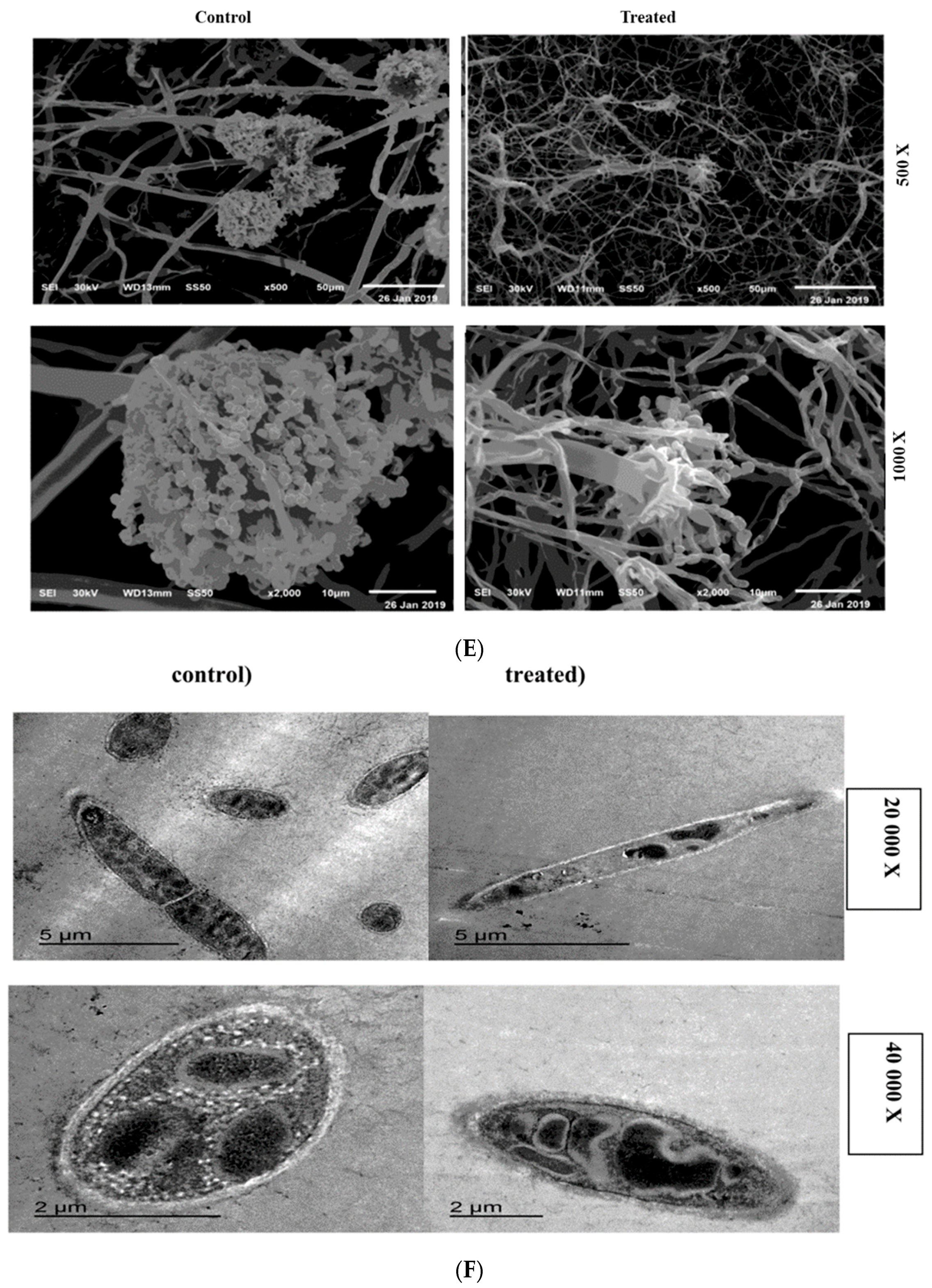
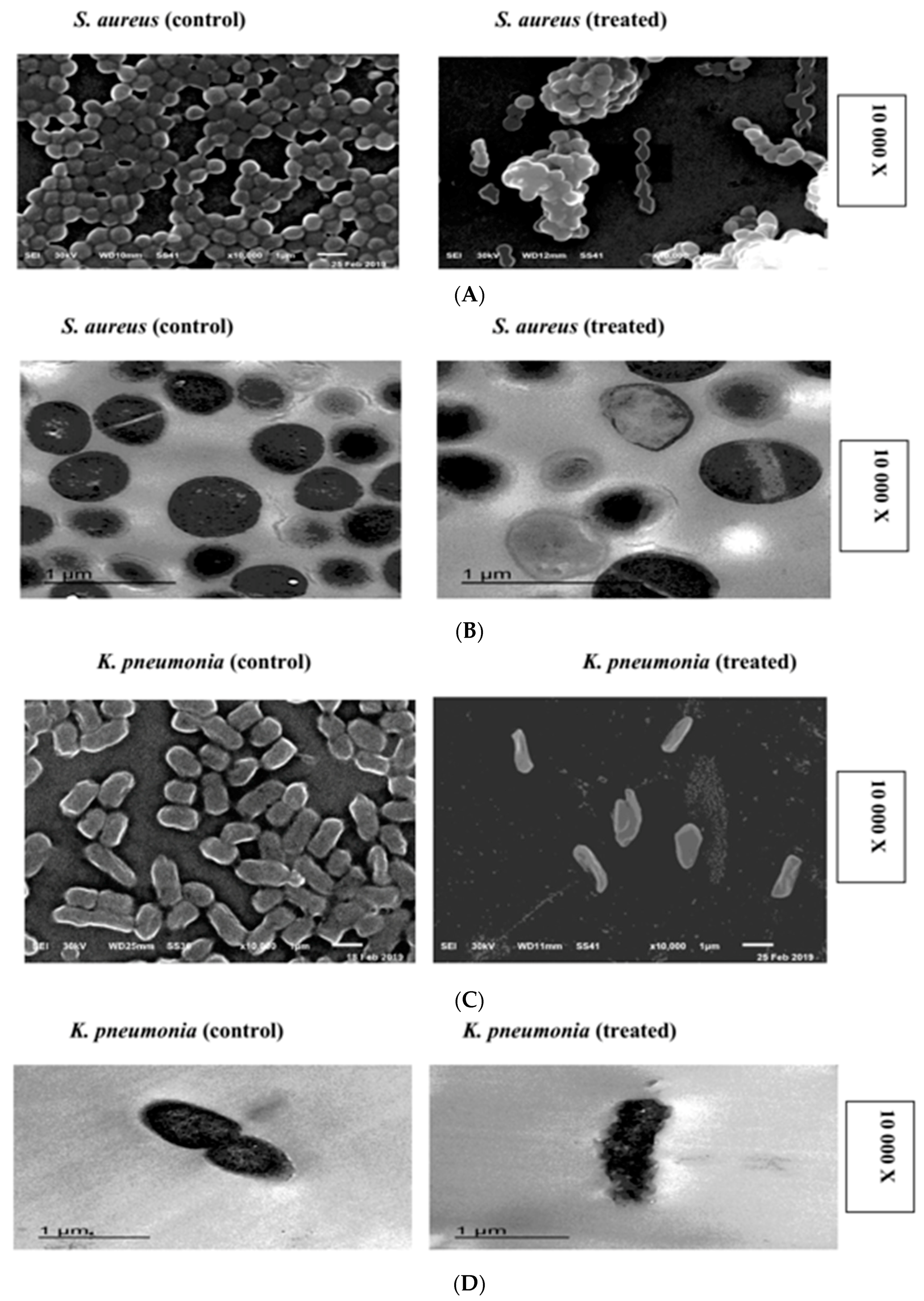
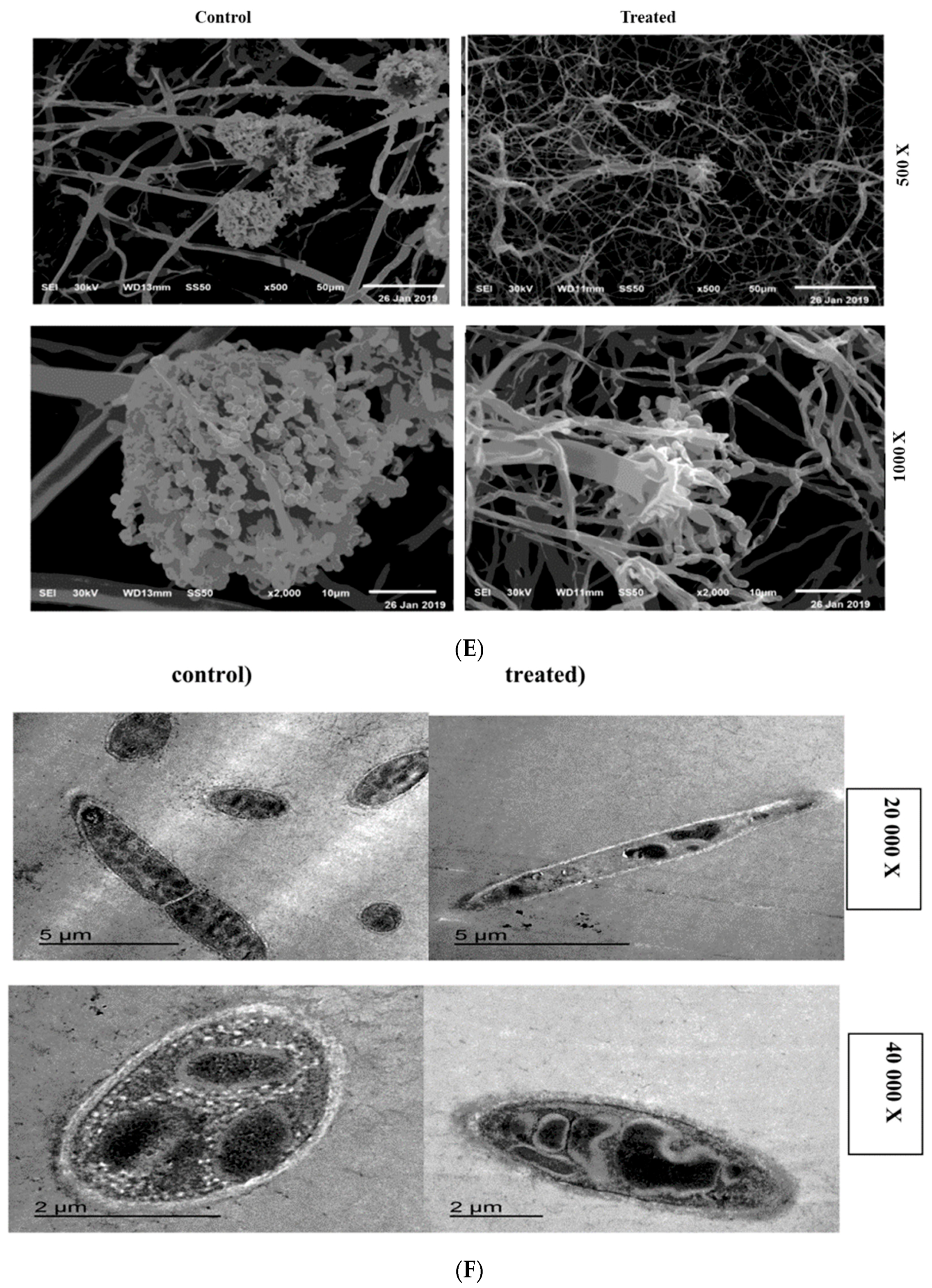
2.6. SEM and TEM Microscopy Analysis
3. Discussion
4. Materials and Methods
4.1. Crude Phenolic Rich Extract (CPRE) Preparation
4.2. Crude Phenolic Rich Extract Characterization
4.2.1. Determination of Anthocyanins
4.2.2. Determination of Total Phenolic Compounds (TPCs)
4.2.3. Total Flavonoids (TFs) Determination
4.2.4. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
4.3. Collection of Pathogenic Bacteria and Fungi
4.4. Antibacterial and Antifungal Activities of the CPREs
4.5. MIC Values of H. Sabdariffa CPRE
4.6. Quantitative Inhibition of Pathogenic Bacteria by CPRE (Bacterial Growth Curve)
4.7. Scanning and Transmission Electron Microscopy (SEM-TEM)
4.7.1. Scanning Electron Microscopy (SEM)
4.7.2. Transmission Electron Microscopy (TEM)
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.; Wertheim, H.F.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H. Antibiotic resistance—The need for global solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Mahgoub, S.A.; Osman, A.O. In vitro and in situ antimicrobial action and mechanism of glycinin and its basic subunit. Int. J. Food Microbiol. 2012, 154, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Goda, H.A.; Abdel-Hamid, M.; Badran, S.M.; Otte, J. Antibacterial peptides generated by Alcalase hydrolysis of goat whey. LWT-Food Sci. Technol. 2016, 65, 480–486. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A. Antimicrobial activity of native and esterified legume proteins against Gram-negative and Gram-positive bacteria. Food Chem. 2010, 120, 66–73. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.; Goda, H.A.; De Gobba, C.; Jenssen, H.; Osman, A. Antibacterial activity of papain hydrolysed camel whey and its fractions. Int. Dairy J. 2016, 61, 91–98. [Google Scholar] [CrossRef]
- Osman, A.O.; Mahgoub, S.A.; Sitohy, M.Z. Preservative action of 11S (glycinin) and 7S (β-conglycinin) soy globulin on bovine raw milk stored either at 4 or 25 °C. J. Dairy Res. 2013, 80, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, S.; Osman, A.; Sitohy, M. Inhibition of growth of pathogenic bacteria in raw milk by legume protein esters. J. Food Prot. 2011, 74, 1475–1481. [Google Scholar] [CrossRef]
- Sitohy, M.; Mahgoub, S.; Osman, A.; El-Masry, R.; Al-Gaby, A. Extent and mode of action of cationic legume proteins against Listeria monocytogenes and Salmonella Enteritidis. Probiotics Antimicrob. Proteins 2013, 5, 195–205. [Google Scholar] [CrossRef]
- Sitohy, M.; Mahgoub, S.; Osman, A. Controlling psychrotrophic bacteria in raw buffalo milk preserved at 4 C with esterified legume proteins. LWT-Food Sci. Technol. 2011, 44, 1697–1702. [Google Scholar] [CrossRef]
- Mahgoub, S.A.; Sitohy, M.Z.; Osman, A.O. Counteracting recontamination of pasteurized milk by methylated soybean protein. Food Bioprocess Technol. 2013, 6, 101–109. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Osman, A.; Enan, G.; El-Nemer, M.; Sitohy, M. Antibacterial activity of methylated egg white proteins against pathogenic Gram positive and Gram negative bacteria matching antibiotics. SpringerPlus 2016, 5, 983. [Google Scholar] [CrossRef] [PubMed]
- Sitohy, M.Z.; Osman, A.O. Enhancing milk preservation with esterified legume proteins. Probiotics Antimicrob. Proteins 2011, 3, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; El-Araby, G.M.; Taha, H. Potential use as a bio-preservative from lupin protein hydrolysate generated by alcalase in food system. J. Appl. Biol. Biotechnol. 2016, 4, 76–81. [Google Scholar]
- Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Osman, A.; Enan, G.; Abdel-Hameid, S.; Sitohy, M. Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein. Molecules 2019, 24, 1082. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, S.A.; Osman, A.O.; Sitohy, M.Z. Impeding Bacillus spore germination in vitro and in milk by soy glycinin during long cold storage. J. Gen. Appl. Microbiol. 2016, 62, 52–59. [Google Scholar] [CrossRef]
- Osman, A.; Daidamony, G.; Sitohy, M.; Khalifa, M.; Enan, G. Soybean glycinin basic subunit inhibits methicillin resistant-vancomycin intermediate Staphylococcus aureus (MRSA-VISA) in vitro. Int. J. Appl. Res. Nat. Prod. 2016, 9, 17–26. [Google Scholar]
- Delgado-Vargas, F.; Paredes-Lopez, O. Natural Colorants for Food and Nutraceutical Uses; CRC Press: Boca Raton, FL, USA; London, UK; New York, NY, USA; Washington, DC, USA, 2002. [Google Scholar]
- Delgado-Vargas, F.; Jiménez, A.; Paredes-López, O. Natural pigments: carotenoids, anthocyanins, and betalains—characteristics, biosynthesis, processing, and stability. Critical Rev. Food Sci. Nutr. 2000, 40, 173–289. [Google Scholar] [CrossRef]
- Lila, M.A. Anthocyanins and human health: an in vitro investigative approach. BioMed. Res. Int. 2004, 2004, 306–313. [Google Scholar] [CrossRef]
- Kumar, A.; Premoli, M.; Aria, F.; Bonini, S.A.; Maccarinelli, G.; Gianoncelli, A.; Memo, M.; Mastinu, A. Cannabimimetic plants: are they new cannabinoidergic modulators? Planta 2019, 249, 1681–1694. [Google Scholar] [CrossRef]
- Eder, R. Pigments. In Food Analysis by HPLC; Nollet, L.M., Ed.; Marcel Dekker: Monticello, NY, USA, 2000; pp. 825–880. [Google Scholar]
- Cissé, M.; Vaillant, F.; Pallet, D.; Dornier, M. Selecting ultrafiltration and nanofiltration membranes to concentrate anthocyanins from roselle extract (Hibiscus sabdariffa L.). Food Res. Int. 2011, 44, 2607–2614. [Google Scholar] [CrossRef]
- Diessana, A.; Parkouda, C.; Cissé, M.; Diawara, B.; Dicko, M.H. Optimization of aqueous extraction of anthocyanins from Hibiscus sabdariffa L. calyces for food application. Food Sci. Qual. Manage. 2015, 45, 23–31. [Google Scholar]
- Saed-Moucheshi, A.; Shekoofa, A.; Pessarakli, M. Reactive oxygen species (ROS) generation and detoxifying in plants. J. Plant Nutr. 2014, 37, 1573–1585. [Google Scholar] [CrossRef]
- Miguel, M.G. Anthocyanins: Antioxidant and/or anti-inflammatory activities. J. Appl. Pharm. Sci. 2011, 1, 7–15. [Google Scholar]
- Mastinu, A.; Kumar, A.; Maccarinelli, G.; Bonini, S.A.; Premoli, M.; Aria, F.; Gianoncelli, A.; Memo, M. Zeolite clinoptilolite: Therapeutic virtues of an ancient mineral. Molecules 2019, 24, 1517. [Google Scholar] [CrossRef] [PubMed]
- Elmanama, A.A.; Alyazji, A.A.; Abu-Gheneima, N. Antibacterial, antifungal and synergistic effect of Lawsonia inermis, Punica granatum and Hibiscus sabdariffa. Ann. Alquds Med. 2011, 7, 33–41. [Google Scholar]
- Fullerton, M.; Khatiwada, J.; Johnson, J.U.; Davis, S.; Williams, L.L. Determination of antimicrobial activity of sorrel (Hibiscus sabdariffa) on Esherichia coli O157, H7 isolated from food, veterinary, and clinical samples. J. Med. Food 2011, 14, 950–956. [Google Scholar] [CrossRef] [PubMed]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef]
- Ahmadiani, N.; Robbins, R.J.; Collins, T.M.; Giusti, M.M. Anthocyanins contents, profiles, and color characteristics of red cabbage extracts from different cultivars and maturity stages. J. Agric. Food Chem. 2014, 62, 7524–7531. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, J.; Zhang, Y.; Hu, X.; Liao, X.; Wang, Z. Extraction of anthocyanins from red cabbage using high pressure CO2. Bioresour. Technol. 2010, 101, 7151–7157. [Google Scholar] [CrossRef]
- Isabelle, M.; Lee, B.L.; Lim, M.T.; Koh, W.-P.; Huang, D.; Ong, C.N. Antioxidant activity and profiles of common vegetables in Singapore. Food Chem. 2010, 120, 993–1003. [Google Scholar] [CrossRef]
- Wiczkowski, W.; Szawara-Nowak, D.; Topolska, J. Red cabbage anthocyanins: Profile, isolation, identification, and antioxidant activity. Food Res. Int. 2013, 51, 303–309. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Kopec, A.; Piatkowska, E.; Borczak, B.; Leszczynska, T. The beneficial effects of Brassica vegetables on human health. Rocz. Panstw. Zakl. Hig. 2012, 63, 389–395. [Google Scholar] [PubMed]
- Osman, A.; Abd-Elaziz, S.; Salama, A.; Eita, A.A.; Sitohy, M. Health Protective Actions of Phycocyanin Obtained from an Egyptian Isolate of Spirulina platensis on Albino Rats. Eur. Asian J. BioSci. 2019, 13, 105–112. [Google Scholar]
- Sitohy, M.; Osman, A.; Ghany, A.; Salama, A. Antibacterial phycocyanin from Anabaena oryzae SOS13. Int J. Appl. Res. Nat. Prod. 2015, 8, 27–36. [Google Scholar]
- Salama, A.; Ghany, A.A.; Osman, A.; Sitohy, M. Maximising phycocyanin extraction from a newly identified Egyptian cyanobacteria strain: Anabaena oryzae SOS13. International Food Res. J. 2015, 22, 517. [Google Scholar]
- Khan, M.I. Plant betalains: Safety, antioxidant activity, clinical efficacy, and bioavailability. Compr. Rev Food Sci. Food Saf. 2016, 15, 316–330. [Google Scholar] [CrossRef]
- Wu, L.-C.; Hsu, H.-W.; Chen, Y.-C.; Chiu, C.-C.; Lin, Y.-I.; Ho, J.-A. Antioxidant and antiproliferative activities of red pitaya. Food Chem. 2006, 95, 319–327. [Google Scholar] [CrossRef]
- Adam Burrows, J.D. Palette of our palates: A brief history of food coloring and its regulation. Compr. Rev. Food Sci. Food Saf. 2009, 8, 394–408. [Google Scholar] [CrossRef]
- Chaitanya Lakshmi, G. Food coloring: The natural way. Res. J. Chem. Sci. 2014, 2231, 606X. [Google Scholar]
- Ouyang-Latimer, J.; Jafri, S.; VanTassel, A.; Jiang, Z.-D.; Gurleen, K.; Rodriguez, S.; Nandy, R.K.; Ramamurthy, T.; Chatterjee, S.; McKenzie, R. In vitro antimicrobial susceptibility of bacterial enteropathogens isolated from international travelers to Mexico, Guatemala, and India from 2006 to 2008. Antimicrob. Agents Chemother. 2011, 55, 874–878. [Google Scholar] [CrossRef] [Green Version]
- Iwu, M.W.; Duncan, A.R.; Okunji, C.O. New antimicrobials of plant origin. In Perspectives on New Crops and New Uses; ASHS Press: Alexandria, VA, USA, 1999; pp. 457–462. [Google Scholar]
- Giuliani, A.; Rinaldi, A.C. Beyond natural antimicrobial peptides: multimeric peptides and other peptidomimetic approaches. Cell. Mol. Life Sci. 2011, 68, 2255–2266. [Google Scholar] [CrossRef] [PubMed]
- Riaz, G.; Chopra, R. A review on phytochemistry and therapeutic uses of Hibiscus sabdariffa L. Biomed. Pharm. 2018, 102, 575–586. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.-Y.; Yin, M.-C. Antibacterial effects of roselle calyx extracts and protocatechuic acid in ground beef and apple juice. Foodborne Pathog. Dis. 2009, 6, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Arab, A.A.; Abu-Salem, F.M.; Abou-Arab, E.A. Physico-chemical properties of natural pigments (anthocyanin) extracted from Roselle calyces (Hibiscus subdariffa). J. Am. Sci. 2011, 7, 445–456. [Google Scholar]
- Vasudeva, N.; Sharma, S. Biologically Active Compounds from the Genus Hibiscus. Pharm. Biol. 2008, 46, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Khan, M.; Hailemariam, H. Nutritional and health importance of Hibiscus sabdariffa: a review and indication for research needs. J. Nutr. Health Food Eng. 2017, 6, 00212. [Google Scholar]
- Silva, S.; Costa, E.M.; Mendes, M.; Morais, R.; Calhau, C.; Pintado, M. Antimicrobial, antiadhesive and antibiofilm activity of an ethanolic, anthocyanin-rich blueberry extract purified by solid phase extraction. J. Appl. Microbiol. 2016, 121, 693–703. [Google Scholar] [CrossRef]
- Siva, R.; Palackan, M.G.; Maimoon, L.; Geetha, T.; Bhakta, D.; Balamurugan, P.; Rajanarayanan, S. Evaluation of antibacterial, antifungal, and antioxidant properties of some food dyes. Food Sci. Biotechnol. 2011, 20, 7–13. [Google Scholar] [CrossRef]
- Paśko, P.; Bartoń, H.; Zagrodzki, P.; Gorinstein, S.; Fołta, M.; Zachwieja, Z. Anthocyanins, total polyphenols and antioxidant activity in amaranth and quinoa seeds and sprouts during their growth. Food Chem. 2009, 115, 994–998. [Google Scholar] [CrossRef]
- Waterhouse, A.L. Determination of total phenolics. Curr. protoc. Food Anal. Chem. 2002, 6, I1.1.1–I1.1.8. [Google Scholar]
- Adebiyi, O.E.; Olayemi, F.O.; Ning-Hua, T.; Guang-Zhi, Z. In vitro antioxidant activity, total phenolic and flavonoid contents of ethanol extract of stem and leaf of Grewia carpinifolia. Beni-Suef Univer. J. Basic Appl. Sci. 2017, 6, 10–14. [Google Scholar] [CrossRef]
- Nanda, A.; Saravanan, M. Biosynthesis of silver nanoparticles from Staphylococcus aureus and its antimicrobial activity against MRSA and MRSE. Nanomed. Nanotechnol. Biol. Med. 2009, 5, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Shafi, S.; Osman, A.; Al-Mohammadi, A.-R.; Enan, G.; Kamal, N.; Sitohy, M. Biochemical, biological characteristics and antibacterial activity of glycoprotein extracted from the epidermal mucus of African catfish (Clarias gariepinus). Int. J. Biol. Macromol. 2019, 138, 773–780. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Total Anthocyanin Content (mg/100 g Dry Pigment) | Total Phenolic Content (mg GAE/100 g Dry Pigment) | Total Flavonoid Content (mg QE/100 g Dry Pigment) |
|---|---|---|---|
| H. sabdariffa | 4.2 | 2000 | 430 |
| B. oleracea | 2.7 | 150 | 50 |
| B. vulgaris | 3.8 | 400 | 120 |
| No | Classification | M. Formula | M. W. | Compound Name and Structure |
|---|---|---|---|---|
| 1 | Hydrocarbons (Alkan)-saturated compounds | C22H66 | 450 | CH3(CH2)3OCH3—Dotriacontan |
| 2 | Alcoholic componds | C17H36O | 256 |  |
| 3 | Triazine derivatives | C5H8ClN5 | 173 | 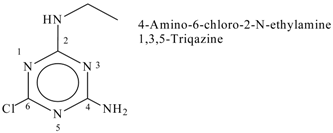 |
| 4 | Unsat. alcoholic compound | C19H38O | 280 | 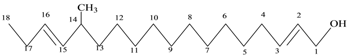 14-Methyl-2,15-octadecadien-1-OL |
| 5 | Unsaturated ester | C17H22O2 | 268 | 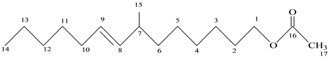 7-Methyl-2-tetradecan-1-OL acetate |
| 6 | Mercepto compound | C16H34S | 258 | 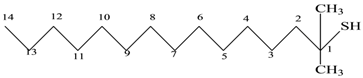 1,1-Dimethyl tetradecyl hydrosulfide or tert-headecanethiol (com.) |
| 7 | Alkenes | C19H38 | 266 | 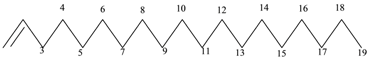 1-Nonadecene or Monadeca-1-ene |
| 8 | Primary alcohols | C37H76O | 536 |  1-Heptalriaontanol or Heptatricotanol |
| 9 | Unsaturated ester | C17H30O2 | 266 | 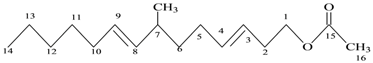 7-Methyl hexadeca-3,8-dienoate |
| 10 | Natural product (Cholesterol) | C28H48O | 400 | 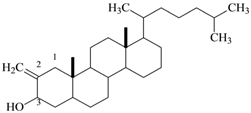 2-Methylene cholestan-3-oL |
| Microorganisms | Inhibition Zone (mm) | ||
|---|---|---|---|
| H. sabdariffa | B. oleracea | B. vulgaris | |
| Gram positive bacteria | |||
| S. aureus | 48 ± 8.0 | 38 ± 3.0 a | 24 ± 4.0 a |
| St. pyogenes | 40 ± 5.0 b | 32 ± 2.0 b | 10 ± 2.0 b |
| L. monocytogenes | 32 ± 4.0 c | 37 ± 3.0 a | 13 ± 3.0 b |
| Gram negative bacteria | |||
| E. coli | 46 ± 6.0 a | 28 ± 0.50 c | 7 ± 0.50 c |
| K. pneumonia | 48 ± 6.5 a | 30 ± 2.0 b | 18 ± 2.0 a |
| P. aeruginosa | 32 ± 3.0 c | 29 ± 0.50 c | 7 ± 0.50 c |
| Microorganisms | Inhibition Zone (mm) | |||||
|---|---|---|---|---|---|---|
| 0 | 25 | 50 | 100 | 200 | 250 | |
| H. sabdariffa pigment | ||||||
| Gram positive bacteria | ||||||
| S. aureus | −ve | −ve | 15 ± 3.0 c | 20 ± 2.0 b | 23 ± 3.0 b | 28 ± 4.0 a |
| St. pyogenes | -ve | −ve | 9 ± 0.5 c | 15 ± 1.0 b | 20 ± 2.0 a | 22 ± 2.0 a |
| L. monocytogenes | −ve | −ve | 11 ± 1.5 c | 22 ± 3.0 b | 23 ± 3.0 b | 26 ± 3.0 a |
| Gram negative bacteria | ||||||
| E. coli | −ve | 11 ± 1.5 c | 14 ± 2.0 c | 20 ± 2.0 b | 24 ± 4.0 b | 30 ± 5.0 a |
| K. pneumonia | −ve | −ve | 15 ± 3.0 c | 20 ± 2.0 b | 25 ± 5.0 a | 26 ± 3.0 a |
| P. aeruginosa | −ve | 10 ± 1.0 | 14 ± 2.0 | 23 ± 3.0 | 29 ± 6.0 | 30 ± 4.0 |
| B. oleracea pigment | ||||||
| Gram positive bacteria | ||||||
| S. aureus | −ve | 9 ± 1.0 c | 12 ± 2.0 c | 16 ± 2.0 b | 21±2.0 a | 25±3.0 a |
| St. pyogenes | −ve | −ve | −ve | 9 ± 0.5 b | 10±1.0 a | 11±1.0 a |
| L. monocytogenes | −ve | −ve | 9 ± 1.0 c | 12 ± 1.5 c | 23±3.0 b | 26±4.0 a |
| Gram negative bacteria | ||||||
| E. coli | −ve | −ve | −ve | 10 ± 1.0 b | 16 ± 2.0 a | 17 ± 2.0 a |
| K. pneumonia | −ve | −ve | −ve | −ve | 11 ± 1.0 b | 13 ± 1.5 a |
| P. aeruginosa | −ve | −ve | −ve | 11 ± 1.2 c | 15 ± 2.0 b | 19 ± 2.5 a |
| Microorganismis | Inhibition Zone (mm) | ||
|---|---|---|---|
| H. sabdariffa | B. oleracea | B. vulgaris | |
| A. ochraceus | 45 ± 5.0 a | 20 ± 2.0 b | −ve |
| F. oxysporum | 40 ± 4.0 b | 22 ± 3.0 a | −ve |
| P. expansum | 35 ± 3.0 c | −ve | −ve |
| P. citrinum | 36 ± 3.2 c | −ve | −ve |
| Microorganisms | Inhibition Zone (mm) | |||||
|---|---|---|---|---|---|---|
| 0 | 100 | 200 | 300 | 400 | 500 | |
| H. sabdariffa pigment | ||||||
| A. ochraceus | −ve | 23 ± 2.0 c | 30 ± 3.0 bc | 34 ± 3.0 b | 35 ± 3.0 b | 40 ± 5.0 a |
| F. oxysporum | −ve | 12 ± 1.0 d | 15 ± 1.5 c | 20 ± 2.0 b | 25 ± 2.0 a | 26 ± 2.0 a |
| P. expansum | −ve | 9 ± 0.5 c | 20 ± 2 b | 21 ± 2.0 b | 28 ± 2.5 a | 32 ± 3.0 a |
| P. citrinum | −ve | 13 ± 1.5 d | 25 ± 3.0 c | 32 ± 3.0 b | 33 ± 3.0 b | 38 ± 4.0 a |
| B. oleracea pigment | ||||||
| A. ochraceus | −ve | −ve | −ve | −ve | 13 ± 2.0 b | 15 ± 1.0 a |
| F. oxysporum | −ve | −ve | −ve | 9 ± 1.0 c | 11 ± 1.0 b | 18 ± 2.0 a |
| P. expansum | −ve | −ve | −ve | −ve | −ve | −ve |
| P. citrinum | −ve | −ve | −ve | −ve | −ve | −ve |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Sitohy, M.; Mosa, B.; Ismaiel, A.; Enan, G.; Osman, A. Antimicrobial Activity and Chemical Constitution of the Crude, Phenolic-Rich Extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris. Molecules 2019, 24, 4280. https://doi.org/10.3390/molecules24234280
Abdel-Shafi S, Al-Mohammadi A-R, Sitohy M, Mosa B, Ismaiel A, Enan G, Osman A. Antimicrobial Activity and Chemical Constitution of the Crude, Phenolic-Rich Extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris. Molecules. 2019; 24(23):4280. https://doi.org/10.3390/molecules24234280
Chicago/Turabian StyleAbdel-Shafi, Seham, Abdul-Raouf Al-Mohammadi, Mahmoud Sitohy, Basma Mosa, Ahmed Ismaiel, Gamal Enan, and Ali Osman. 2019. "Antimicrobial Activity and Chemical Constitution of the Crude, Phenolic-Rich Extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris" Molecules 24, no. 23: 4280. https://doi.org/10.3390/molecules24234280