
Anti-Amyloidogenic and Cyclooxygenase Inhibitory Activity of Guettarda speciosa



Abstract
1. Introduction
2. Results
2.1. Acute Oral Toxicity
2.2. COX-1 and COX-2 Assay
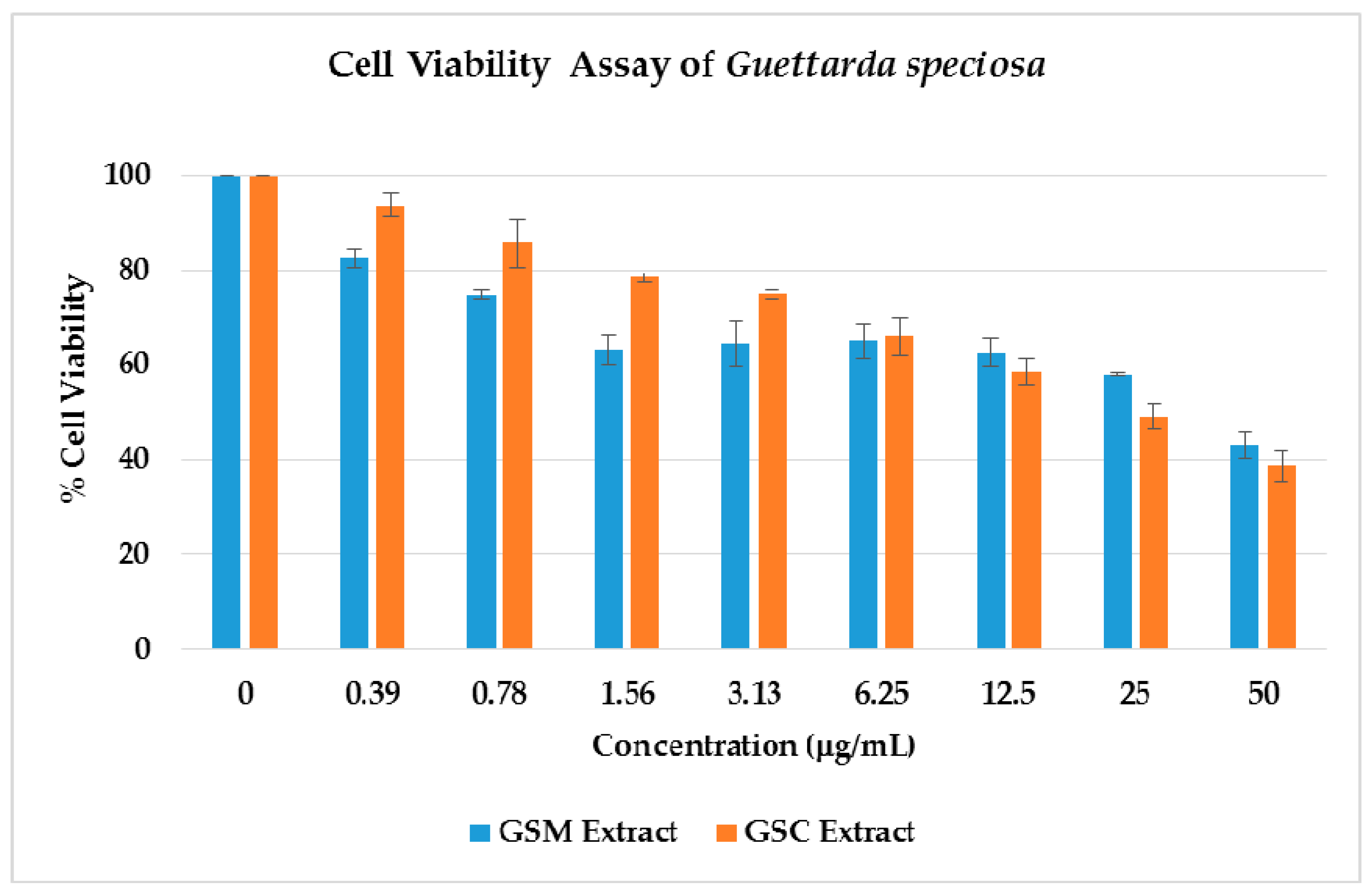
2.3. Cell Viability
2.4. Thioflavin T (ThT) Assay
2.5. Metabolite Profiling
3. Discussion.
4. Materials and Methods
4.1. PlantMaterials
4.2. Extraction and Fractionation of Extracts
4.3. Animal Study
4.4. Acute Oral Toxicity (OECD 425 Guidelines)
4.5. COX-1 and COX-2 Assay
4.6. Cell Viability Assay
4.7. Thioflavin T (ThT) Fluorescense Assay
4.8. Untargeted LC-MS Metabolite Profiling
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alejandro, G.J.D. The Current Status of the Philippine Rubiaceae. Phil. J. Syst. Biol. 2007, 1, 47–60. [Google Scholar] [CrossRef]
- Puff, C.; Chayamarit, K.; Chamchumroon, V. Rubiaceae of Thailand: A Pictorial Guide to Indigenous and Cultivated Genera; Forest Herbarium, National Park, Wildlife and Plant Conservation Department: Prachachon, Bangkok, 2005. [Google Scholar]
- Capasso, A.; Balderrama, L.; Sivila, S.C.; De Tommasi, N.; Sorrentino, L.; Pizza, C. Phytochemical and pharmacological studies of Guettarda acreana. Planta Med. 1998, 64, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Gandhimathi, R.; Saravana, K.A.; Senthil Kumar, K.K.; Kusuma, P.K.; Uma, M.J. Pharmacological studies of anti-diarrhoeal activity of Guettarda speciosa (L.) in experimental animals. J. Pharm. Sci. Res. 2009, 2, 61–67. [Google Scholar]
- Saravana Kumar, A.; Gandhimathi, R. Effect of Guettarda speciosa extracts on biogenic amines concentrations in rat brain after induction of seizure. Int. J. Pharm. Pharm. Sci. 2009, 1, 237–243. [Google Scholar]
- Mohotti, S.; Rajendran, S.; Muhammad, T.; Strömstedt, A.; Adhikari, A.; Burman, R.; deSilva, E.; Göransson, U.; Hettiarachchi, C.; Gunasekera, S. Screening for bioactive metabolites in Sri Lankan medicinal plants by microfractionation and targeted isolation of antimicrobial flavonoids from Derris scandens. J. Ethnopharm. 2020, 246, 112158. [Google Scholar] [CrossRef]
- Kaou, A.M.; Mahiou-Leddet, V.; Hutter, S.; Aïnouddine, S.; Hassani, S.; Yahaya, I.; Azas, N.; Ollivier, E. Antimalarial activity of crude extracts from nine African medicinal plants. J. Ethnopharm. 2008, 116, 74–83. [Google Scholar] [CrossRef]
- Le, H.T.; Cho, Y.C.; Cho, S. Methanol extract of Guettarda speciosa Linn. inhibits the production of inflammatory mediators through the inactivation of Syk and JNK in macrophages. Int. J. Mol. Med. 2018, 41, 1783–1791. [Google Scholar] [CrossRef]
- Cai, W.-H.; Matsunami, K.; Otsuka, H.; Shinzato, T.; Takeda, Y. A glycerol α-d-glucuronide and a megastigmane glycoside from the leaves of Guettarda speciosa L. J. Nat. Med. 2011, 65, 364–369. [Google Scholar] [CrossRef]
- Inouye, H.; Takeda, Y.; Nishimura, H.; Kanomi, A.; Okuda, T.; Puff, C. Chemotaxonomic studies of Rubiaceous plants containing iridoid glycosides. Phytochem. 1988, 27, 2591–2598. [Google Scholar] [CrossRef]
- Mongrand, S.; Badoc, A.; Patouille, B.; Lacomblez, C.; Chavent, M.; Bessoule, J. Chemotaxonomy of the Rubiaceae family based on leaf fatty acid composition. Phytochem. 2005, 66, 549–559. [Google Scholar] [CrossRef]
- Bagyinszky, E.; Giau, V.V.; Shim, K.; Suk, K.; An, S.S.A.; Kim, S. Role of inflammatory molecules in the Alzheimer’s disease progression and diagnosis. J. Neurol. Sci. 2017, 376, 242–254. [Google Scholar] [CrossRef] [PubMed]
- Giau, V.V.; An, S.S.A. Epitope mapping immunoassay analysis of the interaction between β-amyloid and fibrinogen. Int. J. Mol. Sci. 2019, 20, 496. [Google Scholar] [CrossRef] [PubMed]
- Olivar, J.E.; Sy, K.; Villanueva, C.; Alejandro, G.J.; Tan, M.A. Alkaloids as chemotaxonomic markers from the Philippine endemic Uncaria perrottetii and Uncaria lanosa f. philippinensis. J. King Saud. Univ. Sci. 2018, 30, 283–285. [Google Scholar] [CrossRef]
- Tan, M.A.; Callanta, R.B.; Apurillo, C.C.; delaCruz, T.E.; Alejandro, G.J.; Ysrael, M.C. Anti-inflammatory and antimicrobial constituents from the leaves of Villaria odorata. Acta. Manil. 2014, 62, 47–52. [Google Scholar]
- Wu, G.-J.; Chen, W.-F.; Hung, H.-C.; Jean, Y.-H.; Sung, C.-S.; Chakraborty, C.; Lee, H.-P.; Chen, N.-F.; Wen, Z.-H. Effects of propofol on proliferation and anti-apoptosis of neuroblastoma SH-SY5Y cell line: New insights into neuroprotection. Brain Res. 2011, 1384, 42–50. [Google Scholar] [CrossRef]
- Chen, C.-Y. TCM database in Taiwan: The world’s largest traditional Chinese medicine database for drug screening in silico. Plos ONE 2011, 6, 15939. [Google Scholar] [CrossRef]
- He, M.; Grkovic, T.; Evans, J.R.; Thornburg, C.C.; Akee, R.K.; Thompson, J.R.; Whitt, J.A.; Harris, M.J.; Loyal, J.A.; Britt, J.R.; et al. The NCI library of traditional Chinese medicinal plant extracts- Preliminary assessment of the NCI-60 activity and chemical profiling of selected species. Fitoterapia 2019, 137, 104285. [Google Scholar] [CrossRef]
- Aquino, R.; De Simone, F.; Senatore, F.; Pizza, C. Iridoids and secoiridoids from Guettarda platypoda. Pharm. Res.Commun. 1988, 20, 105–108. [Google Scholar] [CrossRef]
- De Oliveira, P.R.; Testa, G.; Medina, R.P.; De Oliveira, C.M.; Kato, L.; Da Silva, C.C.; Santin, S.M. Cytotoxic activity of Guettarda pohliana Müll. Arg. (Rubiaceae). Nat. Prod. Res. 2013, 27, 1677–1681. [Google Scholar] [CrossRef]
- Sousa, M.P.; Matos, M.E.; Machado, M.I.; Filho, R.B.; Vencato, I.; Mascarenhas, Y.P. Triterpenoids from Guettarda angelica. Phytochem. 1984, 23, 2589–2592. [Google Scholar] [CrossRef]
- Bhattacharyya, J.; De Almeida, M. Isolation of the constituents of the root-bark of Guettarda platypoda. J. Nat. Prod. 1985, 48, 148–149. [Google Scholar] [CrossRef]
- Matos, M.E.; Sousa, M.P.; Machado, M.L.; Filho, R.B. Quinovic acid glycosides from Guettarda angelica. Phytochem. 1986, 25, 1419–1422. [Google Scholar] [CrossRef]
- Ferrari, F.; Messana, I.; Botta, B. Constituents of Guettarda platypoda. J. Nat. Prod. 1986, 49, 1150–1151. [Google Scholar] [CrossRef]
- Luijendijk, T.J.; van der Meijden, E.; Verpoorte, R. Involvement of strictosidine as a defensive chemical in Catharanthus roseus. J. Chem. Ecol. 1996, 22, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Han, X.-L.; Li, J.-D.; Yang, C.; Li, Z.-Y. Sweroside eradicated leukemia cells and attenuated pathogenic processes in mice by inducing apoptosis. Biomed. Pharm. 2017, 95, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; West, B.J.; Jarakae Jensen, C. UPLC-TOF-MS characterization and identification of bioactive iridoids in Cornus mas fruit. J. Anal. Methods Chem 2013. [Google Scholar] [CrossRef]
- Sun, H.; Li, L.; Zhang, A.; Zhang, N.; Lu, H.; Sun, W.; Wang, X. Protective effects of sweroside on human MG-63 cells and rat osteoblast. Fitoterapia 2013, 84, 174–179. [Google Scholar] [CrossRef]
- Wang, R.; Dong, Z.; Lan, X.; Liao, Z.; Chen, M. Sweroside alleviated LPS-induced inflammation via SIRT1 mediating NF-κB and FOXO1 signaling pathways in RAW 264.7 cells. Molecules 2019, 24, 872. [Google Scholar] [CrossRef]
- Xu, Y.-D.; Sun, M.-F.; Zhu, Y.-L.; Chu, M.; Shi, Y.-W.; Lin, S.-L.; Yang, X.-S.; Shen, Y.-Q. Neuroprotective effects of loganin on MPTP-induced Parkinson’s disease mice: Neurochemistry, Glial reaction and autophagy studies. J. Cell Biochem. 2017, 118, 3495–3510. [Google Scholar] [CrossRef]
- Khan, M.; Garg, A.; Srivastava, S.K.; Darokar, M.P. Acytotoxic agent from Strychnosnux-vomica and biological evaluation of its modified analogues. Med. Chem. Res. 2012, 21, 2975–2980. [Google Scholar] [CrossRef]
- Pina, E.M.; Araujo, F.W.; Souza, I.A.; Bastos, I.V.; Silva, T.G.; Nascimento, S.C.; Militao, G.C.; Soares, L.A.; Xavier, H.S.; Melo, S.J. Pharmacological screening and acute toxicity of bark roots of Guettarda platypoda. Rev. Bras. Farm. 2012, 22, 1315–1322. [Google Scholar] [CrossRef]
- Gupta, M.B.; Nath, R.; Srivastava, N.; Shanker, K.; Kishor, K.; Bhargava, K.P. Anti-inflammatory and antipyretic activities of β-sitosterol. Planta Med. 1980, 39, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Villasenor, I.M.; Angelada, J.; Canlas, A.P.; Echegoyen, D. Bioactivity studies on β-sitosterol and its glucoside. Phytother. Res. 2002, 16, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Bajko, E.; Kalinowska, M.; Borowski, P.; Siergiejczyk, L.; Lewandowski, W. 5-O-Caffeoylquinicacid: A spectroscopic study and biological screening for antimicrobial activity. LWT Food Sci. Technol. 2016, 65, 471–479. [Google Scholar] [CrossRef]
- Toyama, D.O.; Ferreira, M.; Romoff, P.; Favero, O.A.; Gaeta, H.H.; Toyama, M.H. Effect of chlorogenic acid (5-caffeoylquinic acid) isolated from Baccharis oxyodonta on the structure and pharmacological activities of secretory phospholipase A2 from Crotalus durissus terrificus. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, H.; Kataoka, S.; Okamoto, S.; Hanafi, M.; Shibata, K. Antimicrobial triterpenes from Ilex integra and the mechanism of antifungal action. Phytother. Res. 1999, 13, 151–156. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Ho, D.V.; Vo, H.Q.; Le, A.T.; Nguyen, H.M.; Kodama, T.; Ito, T.; Morita, H.; Raal, A. Antibacterial activities of chemical constituents from the aerial parts of Hedyoti spilulifera. Pharm. Biol. 2017, 55, 787–791. [Google Scholar] [CrossRef]
- Tabassum, N.; Lee, J.H.; Yim, S.H.; Batkhuu, G.J.; Jung, D.W.; Williams, D.R. Isolation of 4,5-O-dicaffeoylquinic acid as a pigmentation inhibitor occurring in Artemisi acapillaris Thunberg and its validation in vivo. Evid. Based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef]
- Martins, D.; Nunez, C.V. Secondary metabolites from Rubiaceae species. Molecules 2015, 20, 13422–13495. [Google Scholar] [CrossRef]
- Chadha, N.; Silakari, O. Indoles as therapeutics of interest in medicinal chemistry: Bird’s eye view. Eur. J. Med. Chem. 2017, 134, 159–184. [Google Scholar] [CrossRef]
- Viljoen, A.; Mncwangi, N.; Vermaak, I. Anti-inflammatory iridoids of botanical origin. Curr. Med. Chem. 2012, 19, 2104–2127. [Google Scholar] [CrossRef] [PubMed]
- Perrone, M.G.; Scilimati, A.; Simone, L.; Vitale, P. Selective COX-1 inhibition: A therapeutic target to be reconsidered. Curr. Med. Chem. 2010, 17, 3769–3805. [Google Scholar] [CrossRef] [PubMed]
- Bjarnason, I.; Scarpignato, C.; Holmgren, E.; Olszewski, M.; Rainsford, K.D.; Lanas, A. Mechanism of damage to the gastrointestinal tract from nonsteroidal anti-inflammatory drugs. Gastroenterology 2018, 154, 500–514. [Google Scholar] [CrossRef] [PubMed]
- Hošek, J.; Leláková, V.; Bobál, P.; Pížová, H.; Gazdová, M.; Malaník, M.; Jakubczyk, K.; Vesely, O.; Landa, P.; Temml, V.; et al. Prenylated stilbenoids affect inflammation by inhibiting the NF-κB/AP-1 signaling pathway and cyclooxygenases and lipoxygenase. J. Nat. Prod. 2019, 82, 1839–1848. [Google Scholar] [CrossRef]
- Xia, C.-L.; Tang, G.-H.; Guo, Y.-Q.; Xu, Y.-K.; Huang, Z.-S.; Yin, S. Mulberry Diels-Alder-type adducts from Morus alba as multi-targeted agents for Alzheimer’s disease. Phytochem. 2019, 157, 82–91. [Google Scholar] [CrossRef]
- Alghazwi, M.; Smid, S.; Musgrave, I.; Zhang, W. In vitro studies of the neuroprotective activities of astaxanthin and fucoxanthin against amyloid beta (Aβ1–42) toxicity and aggregation. Neurochem. Int. 2019, 124, 215–224. [Google Scholar] [CrossRef]
- Tran, A.V.; Shim, K.H.; Vo Thi, T.T.; Kook, J.K.; An, S.S.A.; Lee, S.W. Targetted and controlled drug delivery by multifunctional mesoporous silica nanoparticles with internal fluorescent conjugates and external polydopamine and graphene oxide layers. Acta. Biomater. 2018, 74, 397–413. [Google Scholar] [CrossRef]
Sample Availability: Samples of the plant extracts are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aβ1-42 Aggregation Inhibition (%) a | |
|---|---|
| Phenol Red (50 μM) b | 69.85 ± 0.29 |
| GSM Extract (50 μg/mL) | 54.71 ± 2.92 |
| GSM Extract (5 μg/mL) | 17.20 ± 0.85 |
| GSC Extract (50 μg/mL) | 65.78 ± 3.12 * |
| GSC Extract (5 μg/mL) | 24.63 ± 1.19 |
| RT | Exact Mass | Elemental Composition | Error (ppm) | Putative Identity | Biological Activity | |
|---|---|---|---|---|---|---|
| Calculated | Observed | |||||
| 1.27 | 530.22644 | 530.22500 | C27H34N2O9 | −2.63 | Strictosidine (1) | Antimicrobial [25] |
| 2.26 | 358.12638 | 358.12690 | C16H22O9 | 1.45 | Sweroside (2) | Cytotoxic [26]; Antigenotoxic [27]; Antiosteoporotic [28]; Anti-inflammatory [29] |
| 2.35 | 390.15259 | 390.15900 | C17H26O10 | 2.35 | Loganin (3) | Antigenotoxic [27]; Neuroprotective [30]; Cytotoxic [31] |
| 2.42 | 414.38617 | 414.38650 | C29H50O | 0.72 | β-Sitosterol (4) | Anti-inflammatory [32,33]; Antipyretic [33] Anthelminthic, Antimutagenic, Analgesic [34] |
| 3.25 | 528.21100 | 528.21050 | C27H32N2O9 | −0.94 | Sickingine (5) | |
| 3.37 | 354.09509 | 354.09430 | C16H18O9 | −2.25 | 5-Caffeoylquinic acid (6) | Antimicrobial [35] Anti-inflammatory [36] |
| 4.93 | 488.35019 | 488.35210 | C30H48O5 | 3.88 | Rotundic acid (7) | Antimicrobial [37,38] |
| 15.37 | 516.45500 | 516.45460 | C25H24O12 | −0.77 | 4,5-Dicaffeoylquinic acid (8) | Antipigmentation [39] |
| 17.35 | 486.33453 | 486.33290 | C30H46O5 | −3.28 | Quinovic acid (9) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, M.A.; Lagamayo, M.W.D.; Alejandro, G.J.D.; An, S.S.A. Anti-Amyloidogenic and Cyclooxygenase Inhibitory Activity of Guettarda speciosa. Molecules 2019, 24, 4112. https://doi.org/10.3390/molecules24224112
Tan MA, Lagamayo MWD, Alejandro GJD, An SSA. Anti-Amyloidogenic and Cyclooxygenase Inhibitory Activity of Guettarda speciosa. Molecules. 2019; 24(22):4112. https://doi.org/10.3390/molecules24224112
Chicago/Turabian StyleTan, Mario A., Mark Wilson D. Lagamayo, Grecebio Jonathan D. Alejandro, and Seong Soo A. An. 2019. "Anti-Amyloidogenic and Cyclooxygenase Inhibitory Activity of Guettarda speciosa" Molecules 24, no. 22: 4112. https://doi.org/10.3390/molecules24224112
APA StyleTan, M. A., Lagamayo, M. W. D., Alejandro, G. J. D., & An, S. S. A. (2019). Anti-Amyloidogenic and Cyclooxygenase Inhibitory Activity of Guettarda speciosa. Molecules, 24(22), 4112. https://doi.org/10.3390/molecules24224112