Effects of Light-Emitting Diodes on the Accumulation of Glucosinolates and Phenolic Compounds in Sprouting Canola (Brassica napus L.)

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Samples
2.2. Extraction of Glucosinolates from Brassica napus and HPLC Analysis
2.3. Extraction of Phenylpropanoids from Brassica napus and HPLC Analysis
2.4. Statistical Analysis
3. Results
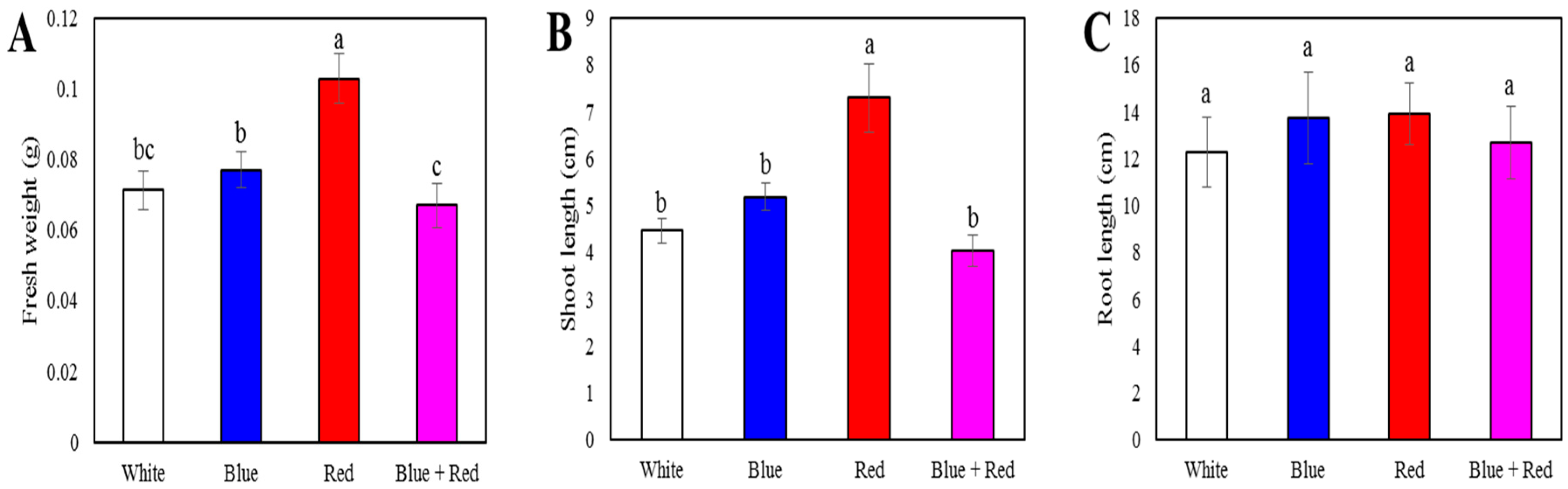
3.1. Sprout Length and Fresh Weight
3.2. Accumulation of Glucosinolates in Sprouts
3.3. Accumulation of Phenolics in Sprouts
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marton, M.; Mandoki, Z.; Csapo-Kiss, Z.; Csapo, J. The role of sprouts in human nutrition. A review. Acta Univ. Sapientiae 2010, 3, 81–117. [Google Scholar]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef]
- Fernandez-Orozco, R.; Piskula, M.K.; Zielinski, H.; Kozlowska, H.; Frias, J.; Vidal-Valverde, C. Germination as a process to improve the antioxidant capacity of Lupinus angustifolius L. var. Zapaton. Eur. Food Res. Technol. 2006, 223, 495. [Google Scholar] [CrossRef]
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: Metabolism and excretion in humans. Cancer Epidemiol. Biomark. Prev. 2001, 10, 501–508. [Google Scholar]
- Baenas, N.; Ferreres, F.; García-Viguera, C.; Moreno, D.A. Radish sprouts—Characterization and elicitation of novel varieties rich in anthocyanins. Food Res. Int. 2015, 69, 305–312. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, Y.B.; Li, X.; Choi, S.R.; Park, S.; Park, J.S.; Lim, Y.P.; Park, S.U. Accumulation of phenylpropanoids by white, blue, and red light irradiation and their organ-specific distribution in Chinese cabbage (Brassica rapa ssp. pekinensis). J. Agric. Food Chem. 2015, 63, 6772–6778. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Park, N.I.; Park, W.T.; Kim, Y.B.; Kim, J.K.; Lee, J.; Lee, S.-H.; Yang, T.-J.; Park, S.U. Carotenoids accumulation and expression of carotenogenesis genes during seedling and leaf development in Chinese cabbage (‘Brassica rapa subsp. pekinensis’). Plant Omics 2012, 5, 143–148. [Google Scholar]
- Kristal, A.R.; Lampe, J.W. Brassica vegetables and prostate cancer risk: A review of the epidemiological evidence. Nutr. Cancer 2002, 42, 1–9. [Google Scholar] [CrossRef] [PubMed]
- USDA. Oilseeds: World Markets and Trade; United States Department of Agriculture, Foreing Agricultural Service: Washington, DC, USA, 2018; pp. 1–40.
- Kumar, A.; Premi, O.; Thomas, L. Rapeseed-Mustard Cultivation in India—An Overview; National Research Centre on Canolaseed-Mustard: Bharatpur, India, 2008. [Google Scholar]
- Szydłowska-Czerniak, A.; Trokowski, K.; Karlovits, G.R.; Szłyk, E. Determination of antioxidant capacity, phenolic acids, and fatty acid composition of rapeseed varieties. J. Agric. Food Chem. 2010, 58, 7502–7509. [Google Scholar] [CrossRef]
- Khattab, R.; Eskin, M.; Aliani, M.; Thiyam, U. Determination of sinapic acid derivatives in canola extracts using high-performance liquid chromatography. J. Am. Oil Chem. Soc. 2010, 87, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.; Morra, M. Hydrolysis products of glucosinolates in Brassica napus tissues as inhibitors of seed germination. Plant Soil 1996, 181, 307–316. [Google Scholar] [CrossRef]
- Vierheilig, H.; Bennett, R.; Kiddle, G.; Kaldorf, M.; Ludwig-Müller, J. Differences in glucosinolate patterns and arbuscular mycorrhizal status of glucosinolate-containing plant species. New Phytol. 2000, 146, 343–352. [Google Scholar] [CrossRef]
- Yasumoto, S.; Matsuzaki, M.; Hirokane, H.; Okada, K. Glucosinolate content in rapeseed in relation to suppression of subsequent crop. Plant Prod. Sci. 2010, 13, 150–155. [Google Scholar] [CrossRef]
- Ludwig-Müller, J.; Schubert, B.; Pieper, K.; Ihmig, S.; Hilgenberg, W. Glucosinolate content in susceptible and resistant Chinese cabbage varieties during development of clubroot disease. Phytochemistry 1997, 44, 407–414. [Google Scholar] [CrossRef]
- Vaughn, S.F. Glucosilates as natural pesticides. In Biologically Active Natural Products: Agrochemicals; CRC Press: Boca Raton, FL, USA, 1999; pp. 81–91. [Google Scholar]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Heber, D. Vegetables, fruits and phytoestrogens in the prevention of diseases. J. Postgrad. Med. 2004, 50, 145–149. [Google Scholar] [PubMed]
- Higdon, J.V.; Delage, B.; Williams, D.E.; Dashwood, R.H. Cruciferous vegetables and human cancer risk: Epidemiologic evidence and mechanistic basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. New Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Ullah, M.F.; Khan, M.W. Food as medicine: Potential therapeutic tendencies of plant derived polyphenolic compounds. Asian Pac. J. Cancer Prev. 2008, 9, 187–196. [Google Scholar]
- Critchfield, J.W.; Butera, S.T.; Folks, T.M. Inhibition of HIV activation in latently infected cells by flavonoid compounds. AIDS Res. Hum. Retroviruses 1996, 12, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant activity and phenolic compounds of 112 traditional Chinese medicinal plants associated with anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Fang, S.-C.; Hsu, C.-L.; Yen, G.-C. Anti-inflammatory effects of phenolic compounds isolated from the fruits of Artocarpus heterophyllus. J. Agric. Food Chem. 2008, 56, 4463–4468. [Google Scholar] [CrossRef] [PubMed]
- Ferrazzano, G.F.; Amato, I.; Ingenito, A.; De Natale, A.; Pollio, A. Anti-cariogenic effects of polyphenols from plant stimulant beverages (cocoa, coffee, tea). Fitoterapia 2009, 80, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.; Samman, S. Flavonoids—Chemistry, metabolism, cardioprotective effects, and dietary sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Fukuda, N.; Kobayashi, M.; Ubukawa, M.; Takayanagi, K.; Sase, S. Effects of light quality, intensity and duration from different artificial light sources on the growth of petunia (Petunia × hybrida Vilm.). J. Jpn. Soc. Hortic. Sci. 2002, 71, 509–516. [Google Scholar] [CrossRef]
- Yeh, N.; Chung, J.-P. High-brightness LEDs—Energy efficient lighting sources and their potential in indoor plant cultivation. Renew. Sust. Energ. Rev. 2009, 13, 2175–2180. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.-N.; Yoshihara, T. Blue light-emitting diode light irradiation of seedlings improves seedling quality and growth after transplanting in red leaf lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- International Standards Organization (ISO). Part 1: Method Using High Performance Liquid Chromatography. In Rapeseed: Determination of Glucosinolates Content, (ISO 9167-1); ISO: Geneva, Switzerland, 1992; pp. 1–9. [Google Scholar]
- Al-Dhabi, N.A.; Arasu, M.V.; Kim, S.J.; RomijUddin, M.; Park, W.T.; Lee, S.Y.; Park, S.U. Methyl Jasmonate-and Light-Induced Glucosinolate and Anthocyanin Biosynthesis in Radish Seedlings. Nat. Prod. Commun. 2015, 10, 1211–1214. [Google Scholar]
- Kim, S.-J.; Park, W.T.; Uddin, M.R.; Kim, Y.B.; Nam, S.Y.; Jho, K.H.; Park, S.U. Glucosinolate biosynthesis in hairy root cultures of broccoli (Brassica oleracea var. italica). Nat. Prod. Commun. 2013, 8, 217–220. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Park, Y.J.; Morgan, A.; Valan Arasu, M.; Al-Dhabi, N.A.; Park, S.U. Influence of indole-3-acetic acid and gibberellic acid on phenylpropanoid accumulation in common buckwheat (Fagopyrum esculentum Moench) sprouts. Molecules 2017, 22, 374. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Zobayed, S.M.; Kozai, T.; Goto, E. Medicinally important secondary metabolites and growth of Hypericum perforatum L. plants as affected by light quality and intensity. Environ. Control Biol. 2007, 45, 113–120. [Google Scholar] [CrossRef]
- McNellis, T.W.; Deng, X.-W. Light control of seedling morphogenetic pattern. Plant Cell 1995, 7, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, Y.; Kitaya, Y.; Kozai, T.; Kimura, T. Effects of red and far-red light on the growth and morphology of potato plantlets in vitro: Using light emitting diode as a light source for micropropagation. Acta Hortic. 1994, 393, 189–194. [Google Scholar] [CrossRef]
- Wu, M.-C.; Hou, C.-Y.; Jiang, C.-M.; Wang, Y.-T.; Wang, C.-Y.; Chen, H.-H.; Chang, H.-M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Thwe, A.A.; Kim, Y.B.; Li, X.; Seo, J.M.; Kim, S.-J.; Suzuki, T.; Chung, S.-O.; Park, S.U. Effects of light-emitting diodes on expression of phenylpropanoid biosynthetic genes and accumulation of phenylpropanoids in Fagopyrum tataricum sprouts. J. Agric. Food Chem. 2014, 62, 4839–4845. [Google Scholar] [CrossRef]
- Velasco, P.; Francisco, M.; Moreno, D.A.; Ferreres, F.; García-Viguera, C.; Cartea, M.E. Phytochemical fingerprinting of vegetable Brassica oleracea and Brassica napus by simultaneous identification of glucosinolates and phenolics. Phytochem. Anal. 2011, 22, 144–152. [Google Scholar] [CrossRef]
- Jiang, J.; Shao, Y.; Li, A.; Lu, C.; Zhang, Y.; Wang, Y. Phenolic composition analysis and gene expression in developing seeds of yellow-and black-seeded Brassica napus. J. Integr. Plant Biol. 2013, 55, 537–551. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Lee, J.G.; Yoo, J.H.; Kim, J.K.; Yu, C.Y. The Influence of Light-Emitting Diodes (LEDs) on the Growth, Antioxidant Activities, and Metabolites in Adventitious Root of Panax ginseng CA Meyer. In Light Emitting Diodes for Agriculture, 1st ed.; Springer: Singapore, 2017; pp. 259–272. [Google Scholar]
- Yeo, H.J.; Park, C.H.; Lee, K.B.; Kim, J.K.; Park, J.S.; Lee, J.W.; Park, S.U. Metabolic Analysis of Vigna unguiculata Sprouts Exposed to Different Light-Emitting Diodes. Nat. Prod. Commun. 2018, 13, 1349–1354. [Google Scholar]
- Szopa, A.; Starzec, A.; Ekiert, H. The importance of monochromatic lights in the production of phenolic acids and flavonoids in shoot cultures of Aronia melanocarpa, Aronia arbutifolia and Aronia × prunifolia. J. Photochem. Photobiol. B Biol. 2018, 179, 91–97. [Google Scholar] [CrossRef]
- Chen, C.-C.; Agrawal, D.C.; Lee, M.-R.; Lee, R.-J.; Kuo, C.-L.; Wu, C.-R.; Tsay, H.-S.; Chang, H.-C. Influence of LED light spectra on in vitro somatic embryogenesis and LC–MS analysis of chlorogenic acid and rutin in Peucedanum japonicum Thunb.: A medicinal herb. Bot. Stud. 2016, 57, 9. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Abbasi, B.H.; Younas, M.; Ahmad, W.; Zahir, A.; Hano, C. LED-enhanced biosynthesis of biologically active ingredients in callus cultures of Ocimum basilicum. J. Photochem. Photobiol. B: Biol. 2019, 190, 172–178. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, G.; Kato, M.; Yamawaki, K.; Takagi, T.; Kiriiwa, Y.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Nesumi, H. Regulation of carotenoid accumulation and the expression of carotenoid metabolic genes in citrus juice sacs in vitro. J. Exp. Bot. 2011, 63, 871–886. [Google Scholar] [CrossRef] [PubMed]
- Tuan, P.A.; Park, C.H.; Park, W.T.; Kim, Y.B.; Kim, Y.J.; Chung, S.O.; Kim, J.K.; Park, S.U. Expression levels of carotenoid biosynthetic genes and carotenoid production in the callus of Scutellaria baicalensis exposed to white, blue, and red light-emitting diodes. Appl. Biol. Chem. 2017, 60, 591–596. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hort. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef]
- Cioć, M.; Szewczyk, A.; Żupnik, M.; Kalisz, A.; Pawłowska, B. LED lighting affects plant growth, morphogenesis and phytochemical contents of Myrtus communis L. in vitro. Plant Cell Tissue Organ Cult. 2018, 132, 433–447. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Yamawaki, K.; Kiriiwa, Y.; Yahata, M.; Ikoma, Y.; Matsumoto, H. Effect of blue and red LED light irradiation on β-cryptoxanthin accumulation in the flavedo of citrus fruits. J. Agric. Food Chem. 2011, 60, 197–201. [Google Scholar] [CrossRef]
- Tuan, P.A.; Thwe, A.A.; Kim, Y.B.; Kim, J.K.; Kim, S.-J.; Lee, S.; Chung, S.-O.; Park, S.U. Effects of white, blue, and red light-emitting diodes on carotenoid biosynthetic gene expression levels and carotenoid accumulation in sprouts of tartary buckwheat (Fagopyrum tataricum Gaertn.). J. Agric. Food Chem. 2013, 61, 12356–12361. [Google Scholar] [CrossRef]
- Lee, S.-W.; Seo, J.M.; Lee, M.-K.; Chun, J.-H.; Antonisamy, P.; Arasu, M.V.; Suzuki, T.; Al-Dhabi, N.A.; Kim, S.-J. Influence of different LED lamps on the production of phenolic compounds in common and Tartary buckwheat sprouts. Ind. Crops Prod. 2014, 54, 320–326. [Google Scholar] [CrossRef]
- Nhut, D.T.; Huy, N.P.; Tai, N.T.; Nam, N.B.; Luan, V.Q.; Hien, V.T.; Tung, H.T.; Vinh, B.T.; Luan, T.C. Light-emitting diodes and their potential in callus growth, plantlet development and saponin accumulation during somatic embryogenesis of Panax vietnamensis Ha et Grushv. Biotechnol. Biotechnol. Equip. 2015, 29, 299–308. [Google Scholar] [CrossRef]
- Kim, Y.B.; Chun, J.-H.; Kim, H.R.; Kim, S.-J.; Lim, Y.P.; Park, S.U. Variation of glucosinolate accumulation and gene expression of transcription factors at different stages of Chinese cabbage seedlings under light and dark conditions. Nat. Prod. Commun. 2014, 9, 533–537. [Google Scholar] [PubMed]


{kind=link}
| Glucosinolate | White | Blue | Red | Blue + Red |
|---|---|---|---|---|
| Glucoiberin | 0.15 ± 0.01 a 1 | 0.14 ± 0.03 a | 0.12 ± 0.01 a | 0.13 ± 0.01 a |
| Progoitrin | 9.38 ± 0.12 b | 10.68 ± 0.16 a | 10.01 ± 1.07 ab | 7.70 ± 0.17 c |
| Glucoraphanin | 0.21 ± 0.01 b | 0.25 ± 0.01 a | 0.22 ± 0.03 b | 0.20 ± 0.00 b |
| Sinigrin | 1.04 ± 0.04 b | 0.82 ± 0.00 c | 1.60 ± 0.14 a | 1.00 ± 0.02 b |
| Glucoalyssin | 0.07 ± 0.01 a | 0.07 ± 0.00 a | 0.05 ± 0.01 b | 0.05 ± 0.00 b |
| Gluconapoleiferin | 0.36 ± 0.01 ab | 0.39 ± 0.03 a | 0.34 ± 0.05 ab | 0.32 ± 0.02 b |
| Gluconapin | 1.56 ± 0.04 a | 1.58 ± 0.02 a | 1.43 ± 0.13 b | 1.18 ± 0.01 c |
| 4-Hydroxy glucobrassicin | 1.89 ± 0.07 a | 1.78 ± 0.12 a | 1.58 ± 0.06 b | 1.91 ± 0.05 a |
| Glucobrassicanapin | 0.14 ± 0.00 a | 0.13 ± 0.00 a | 0.12 ± 0.01 a | 0.11 ± 0.00 b |
| Glucoerucin | 0.03 ± 0.00 ab | 0.04 ± 0.00 a | 0.03 ± 0.00 b | 0.05 ± 0.01 a |
| Glucoraphasatin | 0.03 ± 0.00 a | 0.04 ± 0.00 a | 0.04 ± 0.00 a | 0.04 ± 0.00 a |
| Glucobrassicin | 1.26 ± 0.03 b | 1.07 ± 0.04 c | 1.36 ± 0.04 a | 1.29 ± 0.02 b |
| 4-Methoxy glucobrassicin | 0.88 ± 0.03 c | 0.96 ± 0.05 b | 1.05 ± 0.02 a | 0.91 ± 0.02 c |
| Glucoberteroin | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.04 ± 0.01 a | 0.03 ± 0.00 a |
| Gluconasturtiin | 0.16 ± 0.00 ab | 0.15 ± 0.01 ab | 0.17 ± 0.02 a | 0.14 ± 0.00 b |
| Neoglucobrassicin | 2.12 ± 0.10 b | 2.55 ± 0.01 a | 2.54 ± 0.17 a | 2.07 ± 0.04 b |
| Total | 19.32 ± 0.13 a | 20.69 ± 0.05 a | 20.65 ± 1.70 a | 17.08 ± 0.28 b |
| Class | Compound | White | Blue | Red | Blue + Red |
|---|---|---|---|---|---|
| Phenolic acid | Caffeic acid | 0.41 ± 0.01 c | 0.55 ± 0.01 a | 0.40 ± 0.02 c | 0.46 ± 0.02 b |
| Chlorogenic acid | 0.14 ± 0.01 a | 0.15 ± 0.01 a | 0.07 ± 0.00 b | 0.15 ± 0.04 a | |
| Sinapic acid | 0.14 ± 0.00 a | 0.08 ± 0.01 b | 0.10 ± 0.01 b | 0.09 ± 0.01 b | |
| Catechin | (-)-Epicatechin | 1.58 ± 0.07 b | 1.77 ± 0.05 a | 1.26 ± 0.04 c | 1.47 ± 0.11 b |
| (+)-Catechin | 0.44 ± 0.00 b 1 | 0.65 ± 0.01 a | 0.49 ± 0.02 b | 0.44 ± 0.07 b | |
| Organic acid | Benzoic acid | ND 2 | 0.09 ± 0.01 | ND | ND |
| Flavonoid | Rutin | 0.30 ± 0.00 a | 0.23 ± 0.00 d | 0.28 ± 0.01 b | 0.25 ± 0.01 c |
| Quercetin | 0.29 ± 0.00 a | 0.29 ± 0.00 a | 0.28 ± 0.00 b | 0.28 ± 0.00 b | |
| Total | 3.29 ± 0.04 b | 3.81 ± 0.08 a | 2.87 ± 0.05 d | 3.14 ± 0.04 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.H.; Kim, N.S.; Park, J.S.; Lee, S.Y.; Lee, J.-W.; Park, S.U. Effects of Light-Emitting Diodes on the Accumulation of Glucosinolates and Phenolic Compounds in Sprouting Canola (Brassica napus L.). Foods 2019, 8, 76. https://doi.org/10.3390/foods8020076
Park CH, Kim NS, Park JS, Lee SY, Lee J-W, Park SU. Effects of Light-Emitting Diodes on the Accumulation of Glucosinolates and Phenolic Compounds in Sprouting Canola (Brassica napus L.). Foods. 2019; 8(2):76. https://doi.org/10.3390/foods8020076
Chicago/Turabian StylePark, Chang Ha, Nam Su Kim, Jong Seok Park, Sook Young Lee, Jong-Won Lee, and Sang Un Park. 2019. "Effects of Light-Emitting Diodes on the Accumulation of Glucosinolates and Phenolic Compounds in Sprouting Canola (Brassica napus L.)" Foods 8, no. 2: 76. https://doi.org/10.3390/foods8020076
APA StylePark, C. H., Kim, N. S., Park, J. S., Lee, S. Y., Lee, J.-W., & Park, S. U. (2019). Effects of Light-Emitting Diodes on the Accumulation of Glucosinolates and Phenolic Compounds in Sprouting Canola (Brassica napus L.). Foods, 8(2), 76. https://doi.org/10.3390/foods8020076