Plasma Corticotropin-Releasing Factor Receptors and B7-2+ Extracellular Vesicles in Blood Correlate with Irritable Bowel Syndrome Disease Severity



Abstract
1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. Animals
2.3. Chemicals and Antibodies
2.4. Extracellular Vesicles Isolation
2.5. Western Blot Analysis
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
3.1. CRF Receptors Are Present in Human Plasma
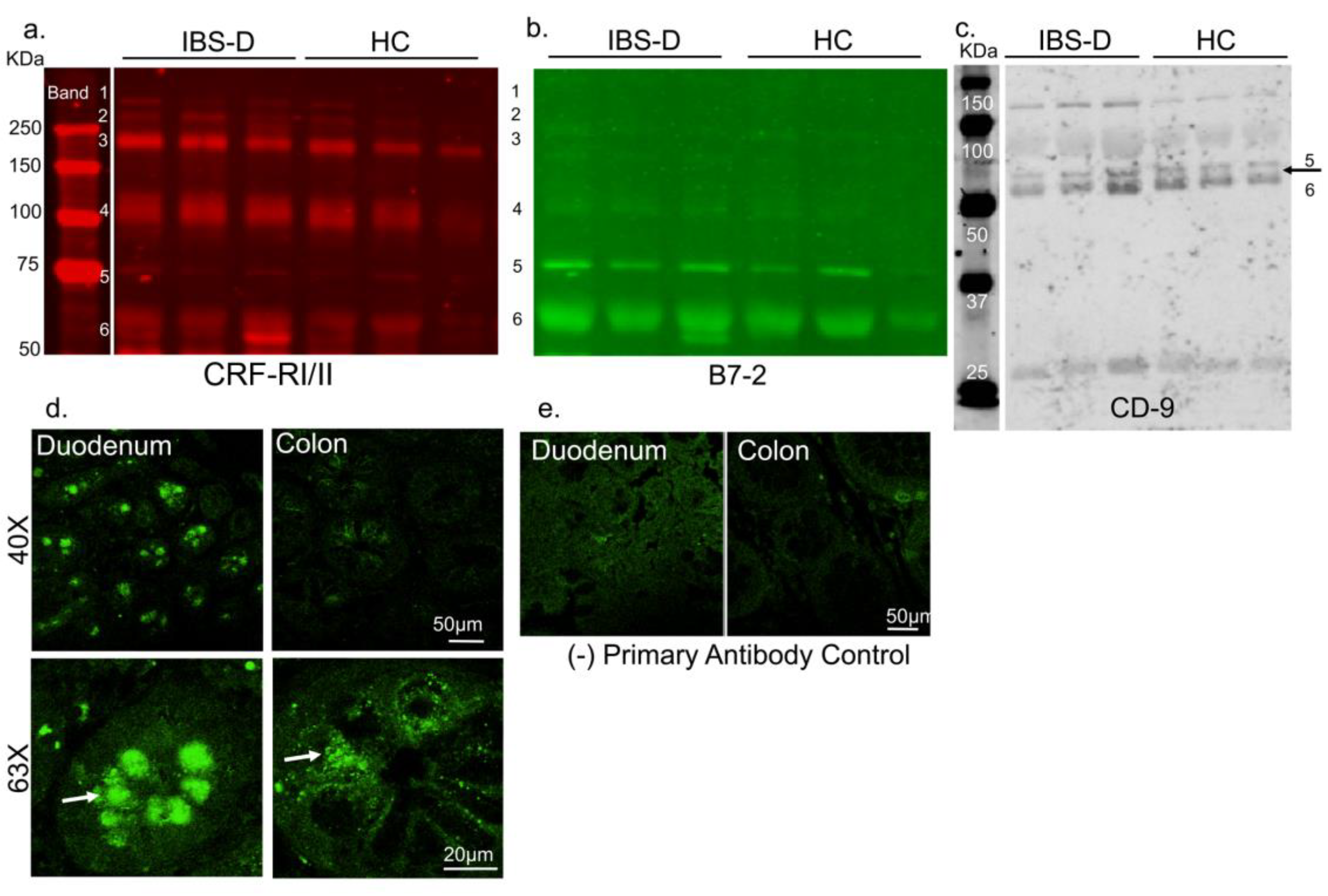
3.1.1. CRF Receptors Are Present in Human Gut
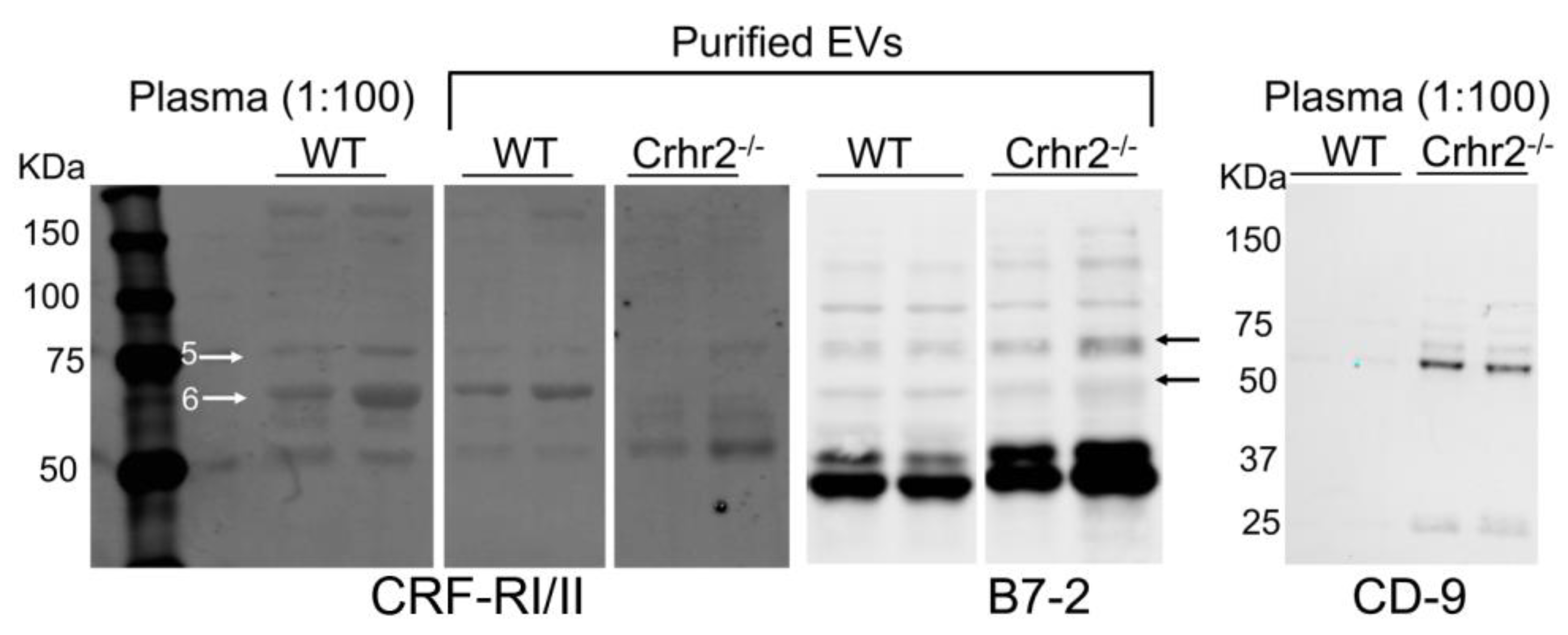
3.1.2. CRF Receptors Are Present in Mouse Plasma and Purified EVs
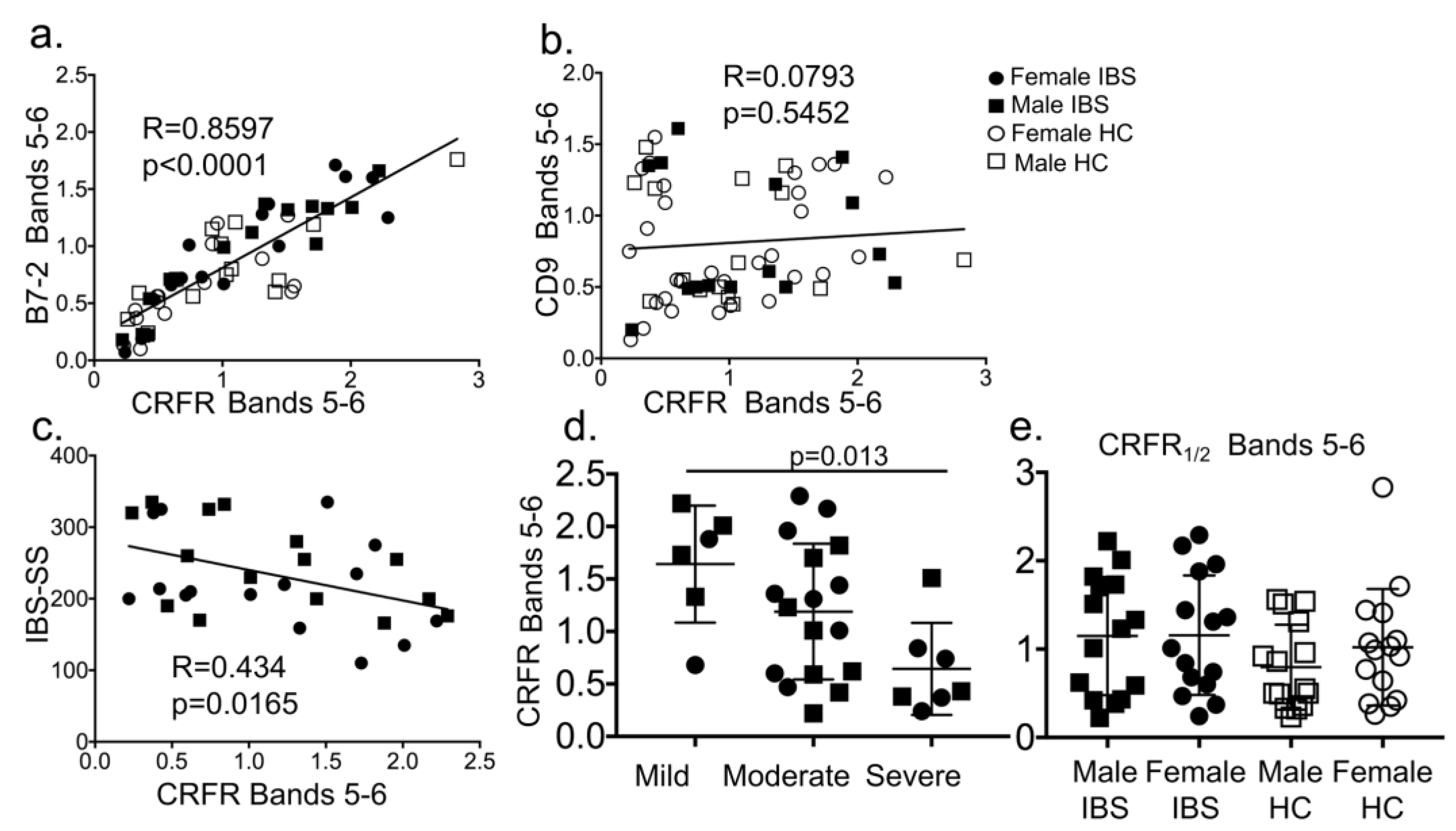
3.1.3. CRF Receptors are Present in Plasma of Patients with Irritable Bowel Syndrome (IBS) and Associate with EVs and Disease Severity Scores
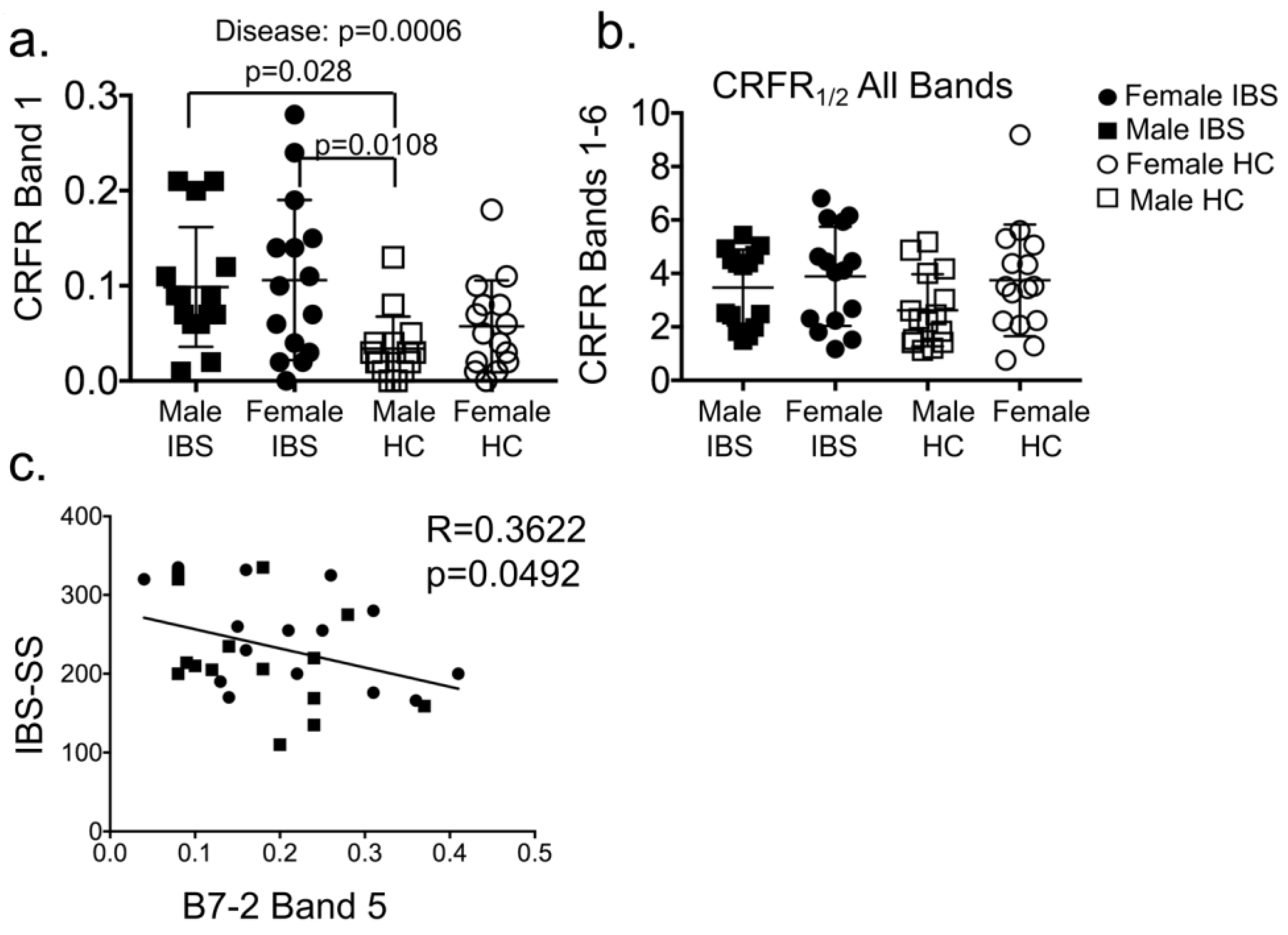
3.1.4. Plasma CRF Receptor Levels Associate with IBS Disease, but Not Sex
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mateescu, B.; Kowal, E.J.; van Balkom, B.W.; Bartel, S.; Bhattacharyya, S.N.; Buzas, E.I.; Buck, A.H.; de Candia, P.; Chow, F.W.; Das, S.; et al. Obstacles and opportunities in the functional analysis of extracellular vesicle rna—An isev position paper. J. Extracell. Vesicles 2017, 6, 1286095. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W.; Soekmadji, C.; Hill, A.F.; Wauben, M.H.; Buzas, E.I.; Di Vizio, D.; Falcon-Perez, J.M.; Gardiner, C.; Hochberg, F.; Kurochkin, I.V.; et al. Updating the misev minimal requirements for extracellular vesicle studies: Building bridges to reproducibility. J. Extracell. Vesicles 2017, 6, 1396823. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Shah, N.; Zanetti, B.R.; Maugeri, M.; Silvestre, R.N.; Fatima, F.; Neder, L.; Valadi, H. Extracellular vesicles and matrix remodeling enzymes: The emerging roles in extracellular matrix remodeling, progression of diseases and tissue repair. Cells 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Nager, A.R.; Goldstein, J.S.; Herranz-Perez, V.; Portran, D.; Ye, F.; Garcia-Verdugo, J.M.; Nachury, M.V. An actin network dispatches ciliary gpcrs into extracellular vesicles to modulate signaling. Cell 2017, 168, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Guescini, M.; Leo, G.; Genedani, S.; Carone, C.; Pederzoli, F.; Ciruela, F.; Guidolin, D.; Stocchi, V.; Mantuano, M.; Borroto-Escuela, D.O.; et al. Microvesicle and tunneling nanotube mediated intercellular transfer of g-protein coupled receptors in cell cultures. Exp. Cell Res. 2012, 318, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular vesicles: Unique intercellular delivery vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed]
- Segura, E.; Nicco, C.; Lombard, B.; Veron, P.; Raposo, G.; Batteux, F.; Amigorena, S.; Thery, C. Icam-1 on exosomes from mature dendritic cells is critical for efficient naive t-cell priming. Blood 2005, 106, 216–223. [Google Scholar] [CrossRef]
- Thery, C.; Duban, L.; Segura, E.; Veron, P.; Lantz, O.; Amigorena, S. Indirect activation of naive cd4+ t cells by dendritic cell-derived exosomes. Nat. Immunol. 2002, 3, 1156–1162. [Google Scholar] [CrossRef]
- Fitzgerald, W.; Freeman, M.L.; Lederman, M.M.; Vasilieva, E.; Romero, R.; Margolis, L. A system of cytokines encapsulated in extracellular vesicles. Sci. Rep. 2018, 8, 8973. [Google Scholar] [CrossRef]
- Mearin, F.; Lacy, B.E.; Chang, L.; Chey, W.D.; Lembo, A.J.; Simren, M.; Spiller, R. Bowel disorders. Gastroenterology 2016. [Google Scholar] [CrossRef]
- Hotta, M.; Shibasaki, T.; Arai, K.; Demura, H. Corticotropin-releasing factor receptor type 1 mediates emotional stress-induced inhibition of food intake and behavioral changes in rats. Brain Res. 1999, 823, 221–225. [Google Scholar] [CrossRef]
- Reyes, T.M.; Lewis, K.; Perrin, M.H.; Kunitake, K.S.; Vaughan, J.; Arias, C.A.; Hogenesch, J.B.; Gulyas, J.; Rivier, J.; Vale, W.W.; et al. Urocortin ii: A member of the corticotropin-releasing factor (crf) neuropeptide family that is selectively bound by type 2 crf receptors. Proc. Natl. Acad. Sci. USA 2001, 98, 2843–2848. [Google Scholar] [CrossRef] [PubMed]
- Paruthiyil, S.; Hagiwara, S.I.; Kundassery, K.; Bhargava, A. Sexually dimorphic metabolic responses mediated by crf2 receptor during nutritional stress in mice. Biol. Sex Differ. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Adams, M.R.; Clifton, M.S.; Liao, M.; Brooks, J.H.; Hasdemir, B.; Bhargava, A. Urocortin 1 modulates immunosignaling in a rat model of colitis via corticotropin-releasing factor receptor 2. AJP Gastrointest. Liver Physiol. 2011, 300, G884–G894. [Google Scholar] [CrossRef]
- Hasdemir, B.; Mhaske, P.; Paruthiyil, S.; Garnett, E.A.; Heyman, M.B.; Matloubian, M.; Bhargava, A. Sex- and corticotropin-releasing factor receptor 2-dependent actions of urocortin 1 during inflammation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R1244–R1257. [Google Scholar] [CrossRef] [PubMed]
- Kubat, E.; Mahajan, S.; Liao, M.; Ackerman, L.; Ohara, P.T.; Grady, E.F.; Bhargava, A. Corticotropin-releasing factor receptor 2 mediates sex-specific cellular stress responses. Mol. Med. 2013, 19, 212–222. [Google Scholar] [CrossRef] [PubMed]
- la Fleur, S.E.; Wick, E.C.; Idumalla, P.S.; Grady, E.F.; Bhargava, A. Role of peripheral corticotropin-releasing factor and urocortin ii in intestinal inflammation and motility in terminal ileum. Proc. Natl. Acad. Sci. USA 2005, 102, 7647–7652. [Google Scholar] [CrossRef]
- D'Costa, S.; Ayyadurai, S.; Gibson, A.J.; Mackey, E.; Rajput, M.; Sommerville, L.J.; Wilson, N.; Li, Y.; Kubat, E.; Kumar, A.; et al. Mast cell crf2 suppresses mast cell degranulation and limits the severity of anaphylaxis and stress-induced intestinal permeability. J. Allergy Clin. Immunol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, S.I.; Kaushal, E.; Paruthiyil, S.; Pasricha, P.J.; Hasdemir, B.; Bhargava, A. Gastric corticotropin-releasing factor influences mast cell infiltration in a rat model of functional dyspepsia. PLoS ONE 2018, 13, e0203704. [Google Scholar] [CrossRef]
- Im, E.; Rhee, S.H.; Park, Y.S.; Fiocchi, C.; TACHE, Y.; Pothoulakis, C. Corticotropin-releasing hormone family of peptides regulates intestinal angiogenesis. Gastroenterology 2010, 138, 2457–2467. [Google Scholar] [CrossRef] [PubMed]
- Gai, Z.; Li, K.; Sun, H.; She, X.; Cui, B.; Wang, R. Effects of chronic noise on mrna and protein expression of crf family molecules and its relationship with p-tau in the rat prefrontal cortex. J. Neurol. Sci. 2016, 368, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L.; Contarino, A.; Smith, G.W.; Chan, R.; Gold, L.H.; Sawchenko, P.E.; Koob, G.F.; Vale, W.W.; Lee, K.F. Mice deficient for corticotropin-releasing hormone receptor-2 display anxiety-like behaviour and are hypersensitive to stress. Nat. Genet. 2000, 24, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Coste, S.C.; Kesterson, R.A.; Heldwein, K.A.; Stevens, S.L.; Heard, A.D.; Hollis, J.H.; Murray, S.E.; Hill, J.K.; Pantely, G.A.; Hohimer, A.R.; et al. Abnormal adaptations to stress and impaired cardiovascular function in mice lacking corticotropin-releasing hormone receptor-2. Nat. Genet. 2000, 24, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Chatzaki, E.; Crowe, P.D.; Wang, L.; Million, M.; Tache, Y.; Grigoriadis, D.E. Crf receptor type 1 and 2 expression and anatomical distribution in the rat colon. J. Neurochem. 2004, 90, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.Q.; Wu, S.V.; Elliott, J.; Anton, P.A.; Chatzaki, E.; Million, M.; Tache, Y. Expression of corticotropin releasing factor receptor type 1 (crf1) in the human gastrointestinal tract and upregulation in the colonic mucosa in patients with ulcerative colitis. Peptides 2012, 38, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hagiwara, S.I.; Bhargava, A. Early-life adversity, epigenetics, and visceral hypersensitivity. Neurogastroenterol. Motil. 2017, 29. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Chang, J.; Long, N.; Beckwith, K.; Talhouarne, G.; Brooks, J.J.; Qu, M.H.; Ren, W.; Wood, J.D.; Cooper, S.; et al. Endogenous crf in rat large intestine mediates motor and secretory responses to stress. Neurogastroenterol. Motil. 2016, 28, 281–291. [Google Scholar] [CrossRef]
- Chatoo, M.; Li, Y.; Ma, Z.; Coote, J.; Du, J.; Chen, X. Involvement of corticotropin-releasing factor and receptors in immune cells in irritable bowel syndrome. Front. Endocrinol. (Lausanne) 2018, 9, 21. [Google Scholar] [CrossRef]
- Chang, J.; Hoy, J.J.; Idumalla, P.S.; Clifton, M.S.; Pecoraro, N.C.; Bhargava, A. Urocortin 2 expression in the rat gastrointestinal tract under basal conditions and in chemical colitis. Peptides 2007, 28, 1453–1460. [Google Scholar] [CrossRef]
- Reul, J.M.; Holsboer, F. Corticotropin-releasing factor receptors 1 and 2 in anxiety and depression. Curr. Opin. Pharmacol. 2002, 2, 23–33. [Google Scholar] [CrossRef]
- Karalis, K.; Sano, H.; Redwine, J.; Listwak, S.; Wilder, R.L.; Chrousos, G.P. Autocrine or paracrine inflammatory actions of corticotropin-releasing hormone in vivo. Science 1991, 254, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Drummen, G.P.; Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef] [PubMed]
- D'Souza-Schorey, C.; Clancy, J.W. Tumor-derived microvesicles: Shedding light on novel microenvironment modulators and prospective cancer biomarkers. Genes Dev. 2012, 26, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Severino, V.; Dumonceau, J.M.; Delhaye, M.; Moll, S.; Annessi-Ramseyer, I.; Robin, X.; Frossard, J.L.; Farina, A. Extracellular vesicles in bile as markers of malignant biliary stenoses. Gastroenterology 2017, 153, 495–504. [Google Scholar] [CrossRef]
- Longstreth, G.F.; Thompson, W.G.; Chey, W.D.; Houghton, L.A.; Mearin, F.; Spiller, R.C. Functional bowel disorders. Gastroenterology 2006, 130, 1480–1491. [Google Scholar] [CrossRef]
- Francis, C.Y.; Morris, J.; Whorwell, P.J. The irritable bowel severity scoring system: A simple method of monitoring irritable bowel syndrome and its progress. Aliment. Pharmacol. Ther. 1997, 11, 395–402. [Google Scholar] [CrossRef]
- Munakata, J.; Naliboff, B.; Harraf, F.; Kodner, A.; Lembo, T.; Chang, L.; Silverman, D.H.; Mayer, E.A. Repetitive sigmoid stimulation induces rectal hyperalgesia in patients with irritable bowel syndrome. Gastroenterology 1997, 112, 55–63. [Google Scholar] [CrossRef]
- Chugh, P.E.; Sin, S.H.; Ozgur, S.; Henry, D.H.; Menezes, P.; Griffith, J.; Eron, J.J.; Damania, B.; Dittmer, D.P. Systemically circulating viral and tumor-derived micrornas in kshv-associated malignancies. PLoS Pathog. 2013, 9, e1003484. [Google Scholar] [CrossRef]
- Umezu, T.; Ohyashiki, K.; Kuroda, M.; Ohyashiki, J.H. Leukemia cell to endothelial cell communication via exosomal mirnas. Oncogene 2013, 32, 2747–2755. [Google Scholar] [CrossRef]
- Ramirez, M.I.; Amorim, M.G.; Gadelha, C.; Milic, I.; Welsh, J.A.; Freitas, V.M.; Nawaz, M.; Akbar, N.; Couch, Y.; Makin, L.; et al. Technical challenges of working with extracellular vesicles. Nanoscale 2018, 10, 881–906. [Google Scholar] [CrossRef]
- Hasdemir, B.; Mahajan, S.; Oses-Prieto, J.; Chand, S.; Woolley, M.; Burlingame, A.; Grammatopoulos, D.K.; Bhargava, A. Actin cytoskeleton-dependent regulation of corticotropin-releasing factor receptor heteromers. Mol. Biol. Cell 2017, 28, 2386–2399. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006. [Google Scholar] [CrossRef]
- Pironti, G.; Strachan, R.T.; Abraham, D.; Mon-Wei Yu, S.; Chen, M.; Chen, W.; Hanada, K.; Mao, L.; Watson, L.J.; Rockman, H.A. Circulating exosomes induced by cardiac pressure overload contain functional angiotensin ii type 1 receptors. Circulation 2015, 131, 2120–2130. [Google Scholar] [CrossRef]
- De Souza, E.B. Corticotropin-releasing factor receptors: Physiology, pharmacology, biochemistry and role in central nervous system and immune disorders. Psychoneuroendocrinology 1995, 20, 789–819. [Google Scholar] [CrossRef]
- Domenis, R.; Cesselli, D.; Toffoletto, B.; Bourkoula, E.; Caponnetto, F.; Manini, I.; Beltrami, A.P.; Ius, T.; Skrap, M.; Di Loreto, C.; et al. Systemic t cells immunosuppression of glioma stem cell-derived exosomes is mediated by monocytic myeloid-derived suppressor cells. PLoS ONE 2017, 12, e0169932. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male IBS | Female IBS | Male HC | Female HC | |
| (n = 15) | (n = 15) | (n = 15) | (n = 15) | |
| Age (years) | 27.2 ± 1.613 | 27.07 ± 3.113 | 27.53 ± 1.756 | 27.47 ± 1.082 |
| IBS-SS | 221.2 ± 17.47 | 246.3 ± 15.92 | ||
| Usual Severity (n) | ||||
| Mild | 4 | 2 | ||
| Moderate | 8 | 9 | ||
| Severe | 3 | 4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hagiwara, S.-i.; Hasdemir, B.; Heyman, M.B.; Chang, L.; Bhargava, A. Plasma Corticotropin-Releasing Factor Receptors and B7-2+ Extracellular Vesicles in Blood Correlate with Irritable Bowel Syndrome Disease Severity. Cells 2019, 8, 101. https://doi.org/10.3390/cells8020101
Hagiwara S-i, Hasdemir B, Heyman MB, Chang L, Bhargava A. Plasma Corticotropin-Releasing Factor Receptors and B7-2+ Extracellular Vesicles in Blood Correlate with Irritable Bowel Syndrome Disease Severity. Cells. 2019; 8(2):101. https://doi.org/10.3390/cells8020101
Chicago/Turabian StyleHagiwara, Shin-ichiro, Burcu Hasdemir, Melvin B. Heyman, Lin Chang, and Aditi Bhargava. 2019. "Plasma Corticotropin-Releasing Factor Receptors and B7-2+ Extracellular Vesicles in Blood Correlate with Irritable Bowel Syndrome Disease Severity" Cells 8, no. 2: 101. https://doi.org/10.3390/cells8020101
APA StyleHagiwara, S.-i., Hasdemir, B., Heyman, M. B., Chang, L., & Bhargava, A. (2019). Plasma Corticotropin-Releasing Factor Receptors and B7-2+ Extracellular Vesicles in Blood Correlate with Irritable Bowel Syndrome Disease Severity. Cells, 8(2), 101. https://doi.org/10.3390/cells8020101