1. Introduction
Chronic bacterial infections exemplify how microenvironmental stress can transform microbial colonization into a lifelong therapeutic challenge, particularly in CF where redox imbalance and biofilm formation undermine antibiotic efficacy. CF is a life-shortening, monogenic disorder caused by mutations in the CFTR gene, leading to defective chloride transport, impaired mucociliary clearance, and the accumulation of dehydrated, viscous mucus in the airways. This pathological environment promotes chronic neutrophilic inflammation and persistent bacterial infections. Among the predominant pathogens colonizing the CF airway are
P. aeruginosa and
S. aureus, along with other opportunistic and emerging bacteria [
1]. These pathogens adapt to intensive and prolonged antibiotic exposure, forming resilient biofilms [
2] and developing multidrug resistance, which severely limits therapeutic options. Consequently, there is an urgent need for antimicrobials with novel modes of action, enhanced stability in pulmonary secretions and minimal propensity to select for resistance.
Redox dysregulation has emerged as a critical determinant of infection persistence and antibiotic performance in CF airways. Multiple studies have shown that bacterial infections in CF induce intracellular and extracellular shifts in redox homeostasis. In healthy individuals, the airway surface liquid maintains an oxidizing redox potential (+100 to +200 mV) and near-neutral pH (7.0–7.5), optimal for mucosal defense [
3]. In contrast, CF airway secretions are acidic (pH 5.0–6.5) and strongly reducing (−200 to −300 mV) due to hypoxia, neutrophilic inflammation, and the accumulation of reduced thiols such as glutathione and cysteine. Such a redox-imbalanced microenvironment alters the activity of both host-derived and therapeutic bioactive molecules, thereby promoting bacterial persistence [
4,
5]. In the context of CF lung infections, colistin (CST), a cyclic AMP, is among the few antibiotics that retain antibacterial efficacy under acidic and reducing microenvironments characteristic of the CF airway [
6]. In contrast, β-lactam antibiotics, including imipenem (IPM)and ceftazidime, demonstrate significantly reduced activity under these conditions, with both acidic pH and redox stress compromising their bactericidal functions [
7]. This underscores the importance of identifying novel antimicrobials that do not induce resistance and are not cytotoxic while maintaining their efficacy in fluctuating redox environments such as the CF lung.
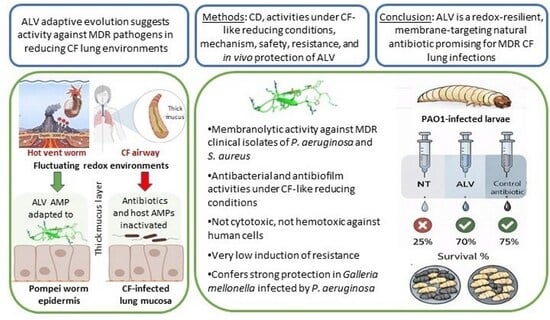
Interestingly, while arising in very different contexts, the chemical imbalance observed in the airways of individuals with CF provides a useful analogy to chemical fluctuations naturally occurring in hydrothermal vent ecosystems [
8]. In both systems, a steep gradient exists between oxidizing zones rich in oxygen and reducing zones dominated by reactive species such as sulfides. In CF lungs, thick mucus layers and persistent bacterial biofilms lead to the local depletion of oxygen, resulting in the formation of hypoxic or anoxic niches with redox potential (Eh) values dropping from +400 mV to below 0 mV [
9,
10]. While not directly comparable, this redox stratification provides a useful analogy to that described for
A. pompejana’s tube [
11].
A. pompejana, is a deep-sea worm endemic to hydrothermal vents along the East Pacific Rise. It inhabits a self-constructed tube situated at the interface between cold, oxygenated seawater and hot, anoxic, sulfur-rich vent fluids where the redox potential may shift by over 600 mV—from +450 mV to below −300 mV across less than 1 cm [
11,
12]. To thrive in chemically unstable environments such as hydrothermal vents, resident species have evolved specialized adaptive strategies to preserve protein structure and function [
13]. In
A. pompejana, one such adaptation is the production of ALV, a β-hairpin cationic AMP stabilized by two intramolecular disulfide bonds [
14]. In contrast, AMP analogs produced by temperate or polar annelid species contain either one single disulfide bond or none at all [
15]. Remarkably, ALV retains its bactericidal activity against hydrothermal
Pseudomonas strains related to
P. aeruginosa, the primary pathogen in CF, even under high salt concentrations and across a wide pH range [
15], conditions that typically inactivate many AMPs [
16] and antibiotics [
17,
18]. The salt-induced inactivation of host defense peptides represents a significant challenge in CF patients [
19,
20]. In this context, ALV’s capacity to rapidly and efficiently kill bacteria such as
Pseudomonas spp. in high-salt environments highlights its potential for drug development.
Based on the salt and pH adaptation of ALV to hydrothermal vents [
15,
21], we hypothesized that this extremophile AMP has also evolved to be redox-resilient. To test this, we evaluated the antibacterial and antibiofilm activities of ALV against the Gram-negative
P. aeruginosa and the Gram-positive
S. aureus, including MDR clinical isolates, under conditions mimicking the redox microenvironment of CF-infected airways (+6 mM DTT, corresponding to −300 mV). In parallel, we investigated the structural stability of ALV in the presence of DTT by CD spectroscopy to determine whether reducing conditions affect its secondary structure in tandem with the observed biological activities. By examining its functional spectrum, mechanism of action, and in vivo protective efficacy against infection, we aimed to assess the potential of ALV for the development of novel antimicrobials effective in biofilm-rich respiratory settings.
3. Discussion
Chronic infections caused by
P. aeruginosa and
S. aureus remain major clinical challenges in CF, largely due to antibiotic resistance, biofilm formation, and the altered physicochemical conditions of the CF lung [
1,
26]. The reducing and hypoxic microenvironment of CF airways not only promotes bacterial persistence but also compromises the efficacy of conventional antibiotics that rely on active bacterial metabolism or are sensitive to redox fluctuations [
3,
5]. In this context, the present study identifies the extremophile-derived peptide ALV, as a potent AMP capable of addressing these unmet therapeutic needs [
14,
27].
ALV exhibited strong, dose-dependent bactericidal activity against both P. aeruginosa and S. aureus pathogens, including multidrug-resistant clinical isolates. The identical MIC and MBC values across all tested strains confirm a primarily bactericidal mode of action. Permeabilization assays and scanning electron microscopy revealed extensive bacterial membrane damage consistent with a pore-forming mechanism, which likely explains both its rapid killing kinetics and the low propensity for resistance development. After 30 days of repeated exposure, ALV induced only a modest (eightfold) increase in MICs, comparable to CST—while conventional antibiotics such as IPM, ceftazidime, and methicillin triggered resistance increases exceeding 100- to 1000-fold.
The results of the lipid monolayer assays provide strong biophysical evidence that ALV displays a marked preference for bacterial membrane components over those found in human cells. Specifically, the higher insertion pressure thresholds and greater surface pressure variations in this positively charged AMP in the presence of bacterial lipids—such as cardiolipin, PG, LTA, and LPS—highlight its ability to interact more efficiently with the negatively charged and structurally distinct surfaces of bacterial membranes. In contrast, the much lower insertion into phosphatidylcholine (PC), a zwitterionic lipid predominantly found in eukaryotic membranes, suggests a reduced affinity for human cell membranes, which is consistent with the low cytotoxicity observed in parallel assays.
This lipid selectivity profile is consistent with the canonical mechanism of many AMPs [
28], which exploit the marked differences in charge and lipid composition between host and microbial membranes to achieve selective targeting. ALV displays strong preferential insertion into cardiolipin and PG, two anionic lipids highly enriched in bacterial membranes, indicating that these components likely constitute primary docking and disruption sites. In addition, ALV interacts with LPS and LTA, which are surface-exposed in Gram-negative and Gram-positive bacteria, respectively, suggesting that membrane destabilization may be initiated at the outer leaflet prior to deeper membrane insertion. This membrane-active mode of action is further supported by parallels with arenicin, a β-hairpin AMP from
Arenicola marina, which similarly inserts into bacterial membranes to form pores and induce rapid cell death [
29]. Collectively, these observations support a pore-like, receptor-independent mechanism for ALV, driven by preferential interactions with anionic bacterial lipids and surface components, which underlies its potent antimicrobial activity while limiting host toxicity and the resistance development.
We next investigated whether ALV retained its antimicrobial efficacy under salt or redox conditions mimicking the in vivo environment during infection, particularly within the CF lung, which is characterized by a highly reducing environment (modeled here with 6 mM DTT) and elevated salt concentrations (300 mM NaCl). The activity of ALV against both bacteria was clearly salt-independent. Remarkably, ALV also preserved both its bactericidal activities under redox conditions against P. aeruginosa, whereas IPM efficacy was substantially reduced and CST was only moderately impaired. The differential effect of DTT on IPM and CST suggests a redox-dependent modulation of the Gram-negative envelope rather than a direct interaction with the antibiotics themselves, as neither molecule contains disulfide bonds and is therefore unlikely to be chemically modified by DTT.
To better understand the antibacterial activities observed under reducing conditions, we investigated whether DTT affected the structural organization of ALV (which contains 2 disulfide bonds) using CD spectroscopy. Analyses consistent with previous NMR data [
14] confirmed a predominantly β-sheet/β-hairpin fold and showed that the secondary structure was largely preserved under reducing conditions. In the presence of DTT, ALV retained its anti-
P. aeruginosa activity but lost activity against
S. aureus, indicating that disulfide bonds are not strictly required for antimicrobial activity
per se, but are important for target specificity. Similar structure–function decoupling has been described for several AMPs, where partial structural integrity remains sufficient for activity against some membranes but not others [
30,
31].
In Gram-negative bacteria such as
P. aeruginosa, the outer membrane rich in LPS promotes strong electrostatic and amphipathic interactions with cationic peptides, which may not require a highly constrained tertiary structure. In this context, the preserved β-sheet scaffold of ALV which is a cationic AMP appears sufficient to maintain membrane-disruptive activity. By contrast, Gram-positive bacteria such as
S. aureus possess a thick peptidoglycan layer and lack an outer membrane, which may impose stricter conformational requirements for peptide activity. Reduction of disulfide bonds may alter the spatial arrangement of key residues and impair efficient interaction with this type of cell envelope. Similar dependence on disulfide-stabilized topology has been reported for defensins which are also cysteine-rich AMPs, where reduction preferentially decreases activity against Gram-positive bacteria [
31,
32]. An alternative explanation is that DTT-reduced ALV may also be reoxidized by reactive oxygen species or other oxidizing systems produced by Gram-negative bacteria (and not by Gram positive), restoring its active oxidized form [
33].
Overall, these results indicate that the β-sheet structure of ALV constitutes a robust structural framework sufficient for membrane interaction, while disulfide bonds fine-tune its three-dimensional conformation and optimize activity depending on bacterial envelope architecture. From an evolutionary perspective, the loss of anti-staphylococcal activity in reducing environments suggests that ALV may have evolved to function optimally within the physicochemical and microbial environment of its native habitat. Consistent with this, members of the genus Pseudomonas (and Gram-negative bacteria in general) are prevalent in the A. pompejana habitat, whereas bacteria of the genus Staphylococcus have not been described.
Because, in CF airways, biofilms contribute to long-term bacterial colonization, reduced antibiotic efficacy, and recurrent infections, we investigated the antibiofilm activity of ALV in the presence and absence of DTT. The inhibition of biofilm formation was consistent with the bactericidal activity observed for ALV under both reducing and non-reducing conditions. ALV efficiently inhibited biofilm formation in
P. aeruginosa strains, including clinical isolates, and this activity was maintained under reducing conditions. In contrast, the inhibition of
S. aureus biofilm formation by ALV was affected in a reducing environment. Interestingly, and specifically for
S. aureus strains, ALV was able to disrupt preformed biofilms of both the reference strain and the clinical isolate in a redox-independent manner. Even though the mechanism behind these data remains to be elucidated [
10], the disruptive effect of ALV on established biofilms highlights an additional therapeutic advantage, particularly in the context of chronic and persistent infections where mature biofilms are difficult to eradicate.
In this context, we investigated whether the robust in vitro properties of ALV could translate into measurable in vivo protection using the Galleria mellonella acute infection model. This invertebrate model represents an ethical and experimental preliminary step before progressing to mammalian studies, allowing early evaluation of antimicrobial efficacy while limiting the use of vertebrate animals. Under the experimental conditions tested, ALV treatment led to a dose-dependent improvement in larval survival following P. aeruginosa infection. Although this protective effect was more modest than that achieved with the antibiotic control meropenem, which consistently conferred strong protection and thereby validated the infection model, the observed survival benefit supports a biologically relevant in vivo activity of ALV. Importantly, the limited magnitude of protection in this acute infection setting may reflect the context-dependent nature of ALV’s antimicrobial efficacy and suggests that its full potential could be underestimated in models that do not recapitulate the chronic, biofilm-associated conditions characteristic of CF infections.
Natural antimicrobial molecules, particularly AMPs, represent a rich and largely untapped reservoir for therapeutic innovation [
34]. Few studies have demonstrated that AMPs or nature-inspired analogs can retain activity in hostile environments resembling CF sputum, where pH, ionic strength, and protease activity undermine the stability and efficacy of traditional antibiotics [
35]. Frog-skin–derived peptides, for instance, significantly reduced bacterial burden in murine lung infection models of
P. aeruginosa, demonstrating the translational feasibility of peptide-based therapies for respiratory infections. Overall, the accumulating body of evidence supports the potential of natural and bioinspired peptides [
27] as valuable complements to conventional antibiotics in the treatment of infections [
6].
Among these candidates, the present data identify ALV as particularly promising because of its excellent safety profile, which represents a major advantage over other AMPs such as CST, whose clinical use is limited by nephrotoxicity. No cytotoxicity was detected, even at ALV concentrations well above bactericidal levels, including on BEAS-2B human bronchial epithelial cells, which is particularly relevant for CF airways. These data strongly suggest that ALV is suitable for inhalation via nebulization as a potential treatment for CF pulmonary infections, where repeated direct exposure of the airway epithelium to the peptide is expected. In addition, ALV retains its antibacterial activity under reducing conditions, in the presence of salt, and after incubation in human serum, indicating good stability and preservation of function under physiologically relevant conditions. Its activity across a wide pH range, previously demonstrated, further supports its robustness in variable host environments. ALV also exhibits antibiofilm activity, a property that is uncommon among AMPs, and does not induce bacterial resistance. The promising results obtained in the Galleria mellonella infection model further highlight its therapeutic potential and support its continued preclinical development. Altogether, these findings reinforce the rationale for advancing toward in vivo evaluation in murine infection models. Future work will focus on better characterizing key pharmacological parameters, including the determination of the maximum tolerated dose and toxic dose, comprehensive toxicity profiling, ADME properties (absorption, distribution, metabolism, and excretion), pharmacokinetics, and biodistribution. Additional studies under more complex CF-associated conditions, particularly in the presence of proteases, neutrophil elastase, mucins, or even CF sputum, also represent important perspectives to further strengthen the therapeutic potential of ALV.
4. Materials and Methods
4.1. ALV Synthesis
ALV (Alvinellacin-NH2; sequence: RGC1YTRC2WKVGRNGRVC3MRVC4T-NH2; disulfide bonds C1–C4 and C2–C3) was synthesized by standard solid-phase peptide synthesis (SPPS) (GENEPEP, Hérault, France). Analytical characterization was performed by UPLC–MS using an Ultimate 3000 UPLC system (Thermo Fisher, Waltham, Franklin, USA) equipped with a BEH C18 column (Milford, MA, USA) (300 Å, 1.7 µm, 2.1 × 150 mm), diode array detection, and an LCQ Fleet ion-trap mass spectrometer. Samples were analyzed at 50 °C using a linear 0–70% CH3CN/0.1% TFA gradient over 20 min at 0.4 mL·min
−1. UV detection was carried out at 215, 254 and 280 nm. MS data were acquired in ESI+ mode (m/z 300–2000) (
Figure S1a). MALDI-TOF mass spectra of purified peptides were obtained on a Bruker Autoflex Speed instrument (Billerica, MA, USA) using CHCA or SA matrices (10 mg·mL
−1 in 0.1% TFA, H2O/CH3CN 1:1
v/
v). Reported m/z values correspond to monoisotopic ions unless specified otherwise (
Figure S1b).
4.2. CD Analysis of ALV w/o DTT
The concentration of synthetic ALV solubilized in 10 mM sodium phosphate buffer (pH 7.2) was determined by UV absorbance at 280 nm using a NanoDrop spectrophotometer (Wilmington, DE, USA), based on its theoretical extinction coefficient (ε = 7240 L·mol−1·cm−1). For CD measurements, the stock solution was diluted to 0.07 mg·mL−1 in 10 mM sodium phosphate buffer (pH 7.2) containing DTT (1 mM final concentration) or not. All the solutions used for CD measurements were prepared in a nitrogen glove box (O2 < 0.1 ppm) to minimize thiol oxydation. The use of 6 mM DTT precluded CD analysis due to strong absorbance below 200 nm. CD spectra were recorded on a Jasco J-815 spectropolarimeter ( Hachioji, Tokyo 193-0835, Japan) equipped with a PFD-425S Peltier temperature control unit, using a 0.1 cm path-length quartz cuvette (Hellma, Müllheim, Germany). Spectra were acquired between 185 and 260 nm at 25 °C with a bandwidth of 2 nm, a data pitch of 1 nm, and an integration time of 1 s. Each spectrum corresponded to the average of ten accumulations and was smoothed using a five-point algorithm. CD data were expressed as mean residue weight ellipticity.
CD spectra were deconvolved using BeStSeL webserver.1 [
24].
4.3. Reducing Redox Environment
A reducing redox environment was established with DTT at a final concentration of 6 mM. At this concentration, and under neutral pH (pH 7.0), the redox potential (Eh) is estimated to be approximately −330 mV. DTT was stored in the solid form in a glove box under nitrogen to prevent oxidation, and the solution was prepared freshly before each experiment to ensure maximal reducing capacity.
4.4. Bacterial Strains
Reference bacterial strains used in this study were obtained from the American Type Culture Collection (ATCC) and included
P. aeruginosa (PAO1, ATCC 27853) and
S. aureus (ATCC 29213). Clinical strains used were (i) the methicillin-resistant
S. aureus strain, MRSA 0.1, isolated from a blood culture of a patient hospitalized in geriatrics care, (ii) the
P. aeruginosa CF 9.19 isolated from the sputum of CF patient (National Reference Centre for Antibiotic Resistance, university hospital of Besançon, France) and (iii) the
P. aeruginosa PAL1.1 that was isolated from the airways of an intensive care unit (ICU) patient (Lille CHU hospital, France), with ventilator-associated pneumonia. A detailed characterization of the antibiotic susceptibility profiles of PAO1, PAL1.1 and CF9.19 is provided in
Table S1.
4.5. Antibacterial Activity Assays
The antibacterial activity was tested against both reference and clinical strains. For P. aeruginosa, reference antibiotics used were: (i) CST sulfate (European Pharmacopoeia), a last-resort polymyxin, widely used clinically to treat severe multidrug-resistant P. aeruginosa infections, including healthcare-associated pneumonia (ii) IPM, a broad-spectrum carbapenem, is prescribed as a first-line therapy for serious P. aeruginosa infections and iii) ceftazidime, a third-generation cephalosporin, used to manage P. aeruginosa infections, particularly during acute pulmonary exacerbations in CF patients or in immunocompromised individuals. For S. aureus, methicillin was used as a phenotypic marker distinguishing MSSA from MRSA strains, guiding the use of appropriate agents such as vancomycin or linezolid in resistant cases. Mupirocin was also tested, as it is used as a topical antibiotic and routinely applied for nasal decolonization of S. aureus carriers, especially preoperatively or in patients at heightened risk of nosocomial infection.
MIC and MBC were determined using the broth microdilution method in 96-well plates as previously described [
15]. All assays were performed in triplicate. MIC and MBC were also measured in the presence of NaCl (at 150 and 300 mM final concentration) or after incubation of ALV with 50% (v:v) of human serum (from Sigma Aldrich, city, state, country) at 37 °C during 4 h
4.6. Time-Kill Assay
Kinetic activity of bactericidal activity was assessed against
P. aeruginosa and
S. aureus [
36]. Cultures in exponential phase (10
6 CFU/mL) in Mueller–Hinton broth (MHB; Difco) were treated with serial concentrations of ALV (0.25–16 μM) in 96-well plates (CytoOne
®, STARLAB). OD
600 was recorded every hour over 20 h at 37 °C with shaking (140 rpm) using a Tecan Sunrise microplate reader.
Dose-dependent killing was also measured. Bacteria (10
6 CFU/mL) were incubated overnight with increasing ALV concentrations (4, 8, 12, 16 μM). Viability was assessed by CFU counting after plating on MHB agar and 48 h incubation at 37 °C [
37]. The impact of DTT was assessed by comparing +DTT and −DTT conditions using the same approach. Statistical analyses were conducted with GraphPad Prism 10, using a significance threshold of α = 0.05. Comparisons between control (no treatment) and treated conditions (peptide or antibiotics) were performed using the Mann–Whitney U test.
4.7. Scanning Electron Microscopy (SEM)
Bacteria were grown to the exponential phase, diluted by 1:100 in MHB, and incubated for 2 h at 37 °C. A total of 50 μL of bacterial suspension was incubated with 5 μL of peptides (50 μM) or deionized water (control) for 4 h at 20 °C. Samples were fixed with 2.5% glutaraldehyde, centrifuged onto poly-lysine-coated slides (SHANDON 3, 1000 rpm, 8 min), sputter-coated with 5 nm platinum (Quorum Q150T ES), and imaged with a Merlin VP Compact Zeiss SEM.
4.8. Bacterial Membrane Permeabilization Assay
The effect of ALV on bacterial membrane integrity was evaluated through measurement of the release of intracellular ATP in culture media, as previously described [
38]. Log-phase bacterial suspensions of
P. aeruginosa (PA01) or
S. aureus (ATCC 29213) were prepared in MH media from overnight liquid cultures as explained above. Bacteria were diluted at a density of 10 × 10
7 bacteria per mL of MH containing ATP detection reagent based on luciferase (Yelen, France) into 96-well white plates for luminescence (Greiner BioOne) and exposed to concentrations of ALV corresponding to ¼ of MIC to 8 times MIC (1/2 dilutions). After 1, 5, 10, 20, 40, and 80 min of incubation at 37 °C, the luminescence of the wells was measured using a microplate reader (Biotek, Synergy Mx). Untreated conditions correspond to bacteria treated with vehicle alone. Lysis buffer provided in the ATP detection kit was used as a positive control, giving 100% bacterial membrane permeabilization. Experiments were conducted in triplicate (n = 3).
4.9. Antibiofilm Assays
Antibiofilm activity [
39] was evaluated against the reference and clinical isolates of
P. aeruginosa and
S. aureus. Biofilm-forming ability was classified according to the crystal violet assay using the Optical Density (OD) of the untreated control biofilm for each strain. The cut-off value (ODc) was calculated as: ODc = meanOD negative control +3× Standard Deviation. Strains were then classified as weak, moderate or strong biofilm producers according to [
40]. Based on this classification, PAO1,
S. aureus ATCC 29213 and MRSA 0,1 were moderate biofilm formers, whereas PAL1.1 and CF9.19 were strong biofilm formers.
Overnight cultures in LB broth were diluted to an OD580 of 0.08. Then, 200 µL of suspension was added to sterile 96-well polystyrene microplates and incubated statically at 37 °C for 24 h to allow biofilm formation. ALV was applied either concurrently with inoculation (preventative assay: growth inhibition of the biofilm) or after the 24-h biofilm formation period (therapeutic assay: disruption of the pre-formed biofilm), at various concentrations starting at 0.25, 0.5, 1, 2, 4, 8, 16 and 32 µM. The subinhibitory concentrations were ≤4 µM for P. aeruginosa and ≤8 µM for S. aureus.
Plates were further incubated for 24 h. Following treatment, wells were gently rinsed three times with phosphate-buffered saline (PBS) to remove planktonic cells. Biofilms were fixed in methanol for 15 min, stained with 0.1% crystal violet for 15 min, and then rinsed thoroughly. The biofilm-associated dye was dissolved in 95% ethanol, and absorbance was measured at 570 nm. Planktonic growth controls were performed in parallel under the same conditions, with bacterial growth monitored at OD600.
4.10. Induction of Resistance
P. aeruginosa PAO1 or S. aureus ATCC 29213 cultures were exposed daily for 30 days to ALV or reference antibiotics. MICs were determined by standard broth microdilution on day 1. Each day, the culture from the highest antibiotic concentration allowing growth was diluted 1:100 in MHB to initiate the next passage. Wells without growth were plated on MHA with high ALV concentrations (80 and 160 µM) to detect resistant clones. Experiments were conducted at pH 7.
4.11. Toxicity Assay Using Human Cells
Cytotoxicity of ALV was assessed as previously described [
41] using human kidney (A498; ATCC
® HTB-44™), lung (BEAS-2B; ECACC), intestinal (Caco-2; ATCC
® HTB-37™), skin (HaCaT; Creative Bioarray), and liver cells (HepG2; ATCC
® HB-8065™). Cells were maintained in DMEM supplemented with 10% FBS, 1% L-glutamine, and 1% penicillin–streptomycin, and incubated at 37 °C with 5% CO
2. After detachment with trypsin–EDTA and counting, cells were seeded in 96-well plates at ~10,000 cells/well. Once confluent (48 h), media were replaced, and cells were exposed to increasing ALV concentrations. After 48 h incubation, viability was quantified by resazurin (30 µg/mL in PBS++), with fluorescence measured at 530/590 nm. Data were normalized to untreated controls; 0.1% Triton X-100 served as a cytotoxicity control (100% toxicity). Experiments were performed in triplicate (n = 3).
4.12. Hemotoxicity Evaluation
Hemolytic activity of ALV was evaluated using human red blood cells (hRBCs; Divbio Science Europe) as previously reported (DOI: 10.1038/s41522-022-00320-0). After three PBS washes and centrifugation (800× g, 5 min), hRBCs were resuspended at 8% in DMEM without phenol red supplemented with 10% FBS. A total of 150 µL of hRBCs was added to 96-well plates containing 150 µL of ALV at increasing concentrations. Following 1 h incubation at 37 °C, samples were centrifuged (800× g, 5 min) and 100 µL of supernatant was transferred to fresh plates. Hemolysis was quantified at 450 nm. Triton X-100 (0.1% v/v) was used as a positive control (100% hemolysis). All experiments were performed in triplicate (n = 3).
4.13. Peptide–Lipid Interaction Assay
The interaction of ALV with lipids was assessed using a lipid monolayer assay as previously described [
42]. Prokaryotic and eukaryotic lipids (cardiolipin, POPC, POPE, POPG; Avanti Polar Lipids) as well as LTA (
S. aureus) and LPS (
P. aeruginosa) (Invitrogen) were tested. Lipids were prepared at 1 mg/mL in chloroform:methanol (2:1) and stored under nitrogen at −20 °C. Monolayers were formed at the air–water interface (PBS) by spreading lipids to the desired initial surface pressure. After 5–10 min stabilization, 8 µL of ALV were injected into the subphase to reach a final concentration of 1 µM, previously identified as optimal for insertion. Surface pressure changes induced by peptide insertion were continuously monitored until equilibrium using an automated microtensiometer (µTROUGH SX, Kibron). The critical insertion pressure (CIP) was determined by measuring peptide-induced pressure increases at initial surface pressures between 10 and 30 mN/m and extrapolating the initial pressure corresponding to zero insertion. ALV insertion was also evaluated at 30 ± 0.5 mN/m, a value representative of the outer leaflet of biological membranes. Experiments were performed at 20 ± 1 °C and analyzed using Firmware 2.5 (Kibron). The accuracy of surface pressure measurements under our conditions was ±0.25 mN/m.
4.14. In Vivo Evaluation of the ALV Activity Using the Galleria mellonella Larval Model
Final instar
Galleria mellonella larvae (four weeks old; 220–300 mg) were used for all
in vivo experiments [
43,
44]. Larvae originated from a genetically homogeneous colony maintained for more than 20 years and were supplied by the Microbial Genetics and Environment team at the Micalis Institute (INRAE). Injections were performed using a microinfusion system consisting of a 27-gauge microperfuser (Venofix
® 27G, B. Braun) mounted on a 1 mL syringe (BD Plastipak™ Tuberculin, ref. 00655) and driven by a syringe pump (KD Scientific) to ensure accurate and reproducible injection volumes. Bacterial suspensions or control solutions (10 µL) were injected into the hemocoel
via the last left proleg. For each experiment, a group of non-infected larvae injected with phosphate-buffered saline (PBS) was included as an injection control to monitor procedure-related mortality and to verify the absence of toxicity associated with the injected volumes. Ten larvae were used per condition unless otherwise specified
To assess the virulence of P. aeruginosa strains PAO1, PAL1.1, and CF9.19, groups of ten larvae were infected with increasing bacterial inocula ranging from 101 to 107 CFU/mL. Larvae were incubated at 37 °C, and survival was monitored daily for three days post-infection. Dose–response survival data were analyzed using logistic regression to model the relationship between bacterial dose and larval survival. A total of 15 independent experiments, each comprising ten larvae per condition, were included in the dose–response analysis.
Prior to infection experiments, the toxicity of ALV was assessed in Galleria mellonella. Larvae (n = 10 per condition) were injected with increasing concentrations of ALV (5, 10, 20, 50 and 100 mg/kg) and monitored over 72 h. No mortality or signs of melanization were observed at any tested dose. The highest concentration tested corresponded to the maximal injectable dose (10 µL of a 10 mM stock solution).
ALV efficacy was evaluated using the P. aeruginosa reference strain PAO1. Based on the analysis of the dose–response survival profile, an infective dose of 1 × 105 CFU/mL was selected, corresponding to a condition yielding less than 25% larval survival at 24 h. This experimental setting ensured a robust infection while allowing the detection of potential treatment effects. ALV was administered 1 h post-infection at doses of 10 mg/kg or 100 mg/kg by injection into the contralateral proleg to avoid interference with the infection site. Meropenem (40 mg/kg) was used as a positive control for protection. Untreated infected larvae and PBS-injected larvae served as negative and injection controls, respectively. Larval survival was assessed at 24 h post-infection. Experiments were performed in three independent replicates, with 10 larvae in one experiment and 6 larvae in each of the two others.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}