
p38- and ERK-MAPK Signalling Modulate Developmental Neurotoxicity of Nickel and Vanadium in the Caenorhabditis elegans Model

 , , and
, , and 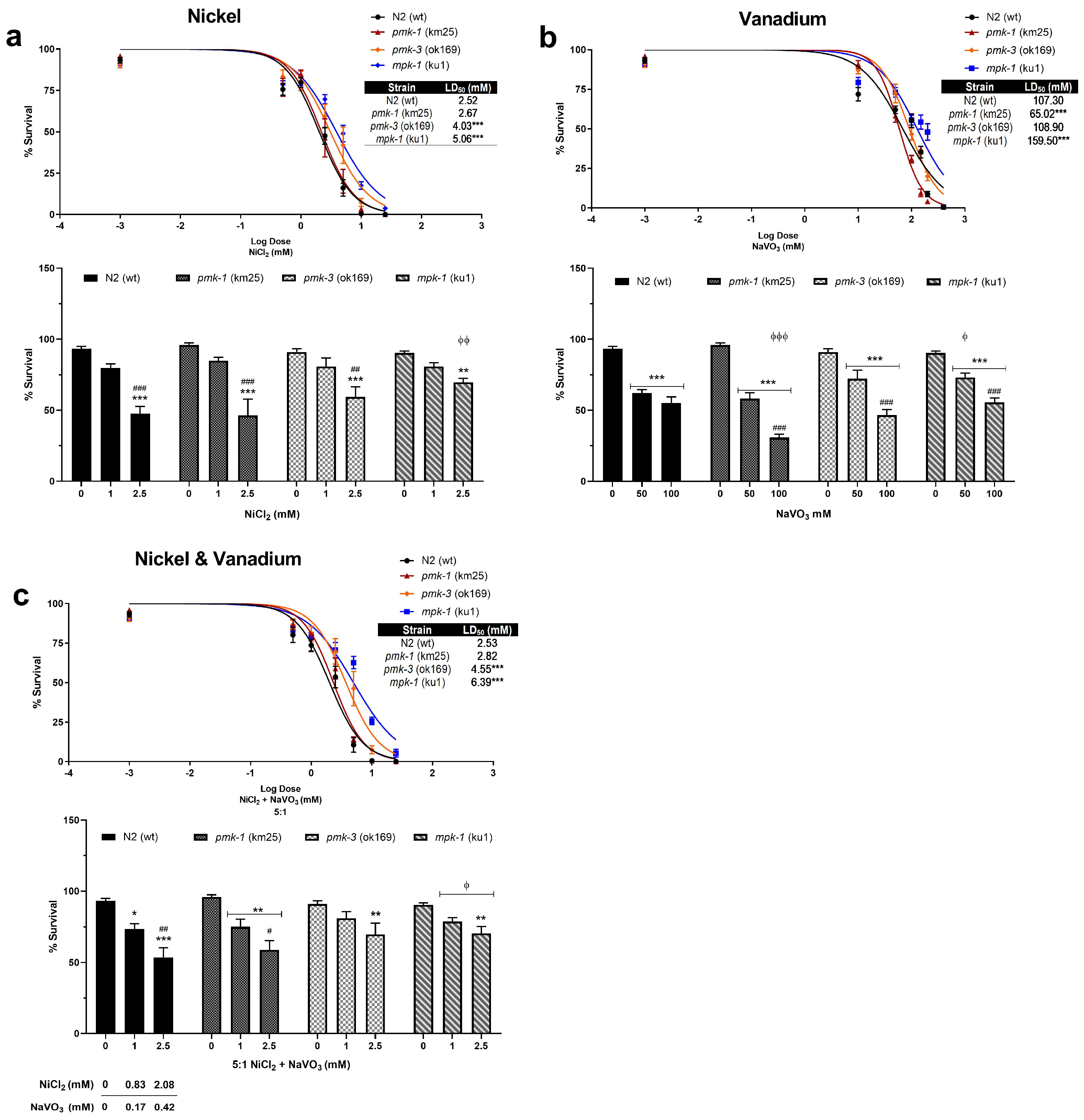{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
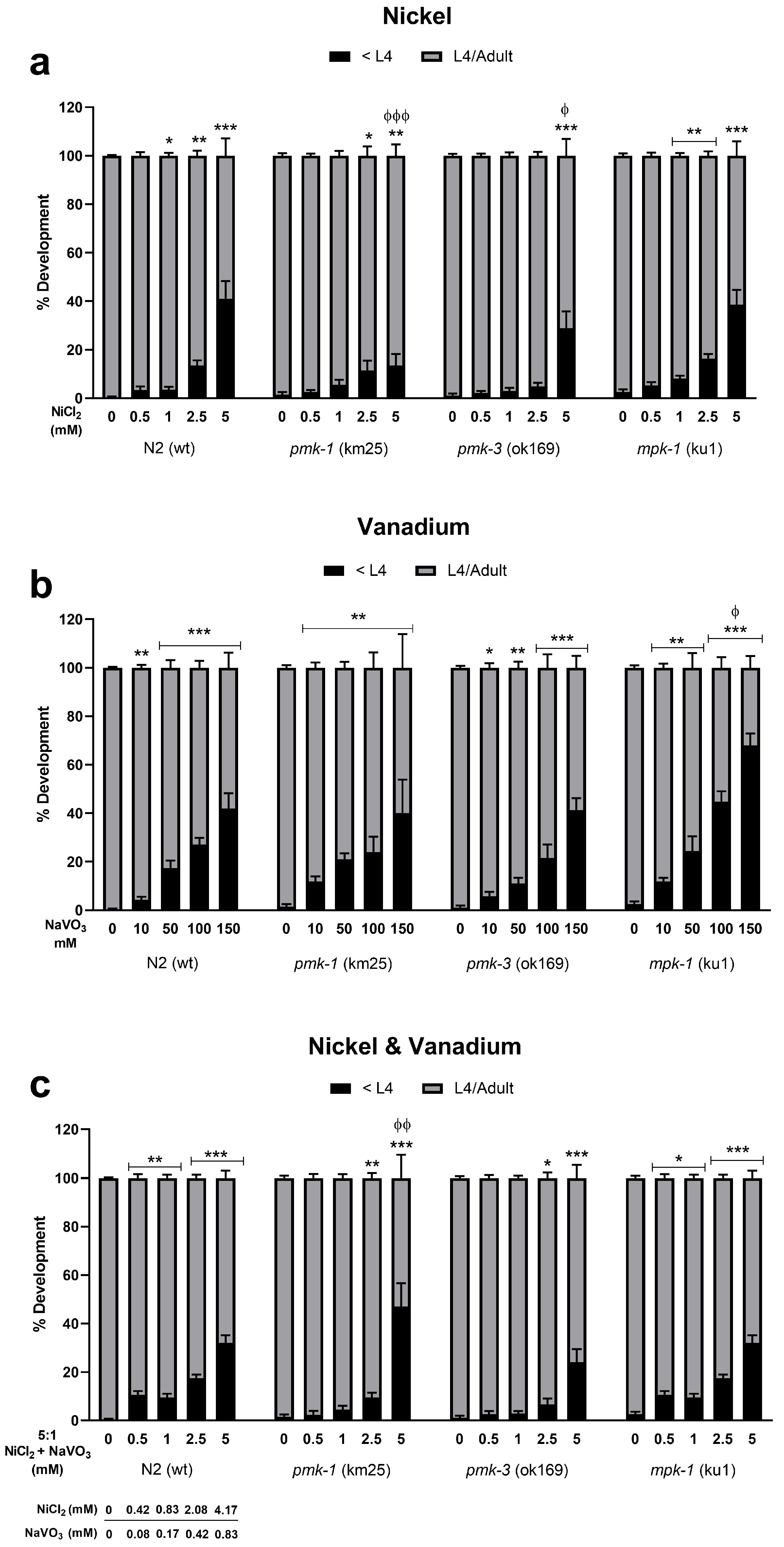
2.1. Toxicity and Developmental Delays after Ni and V Treatment
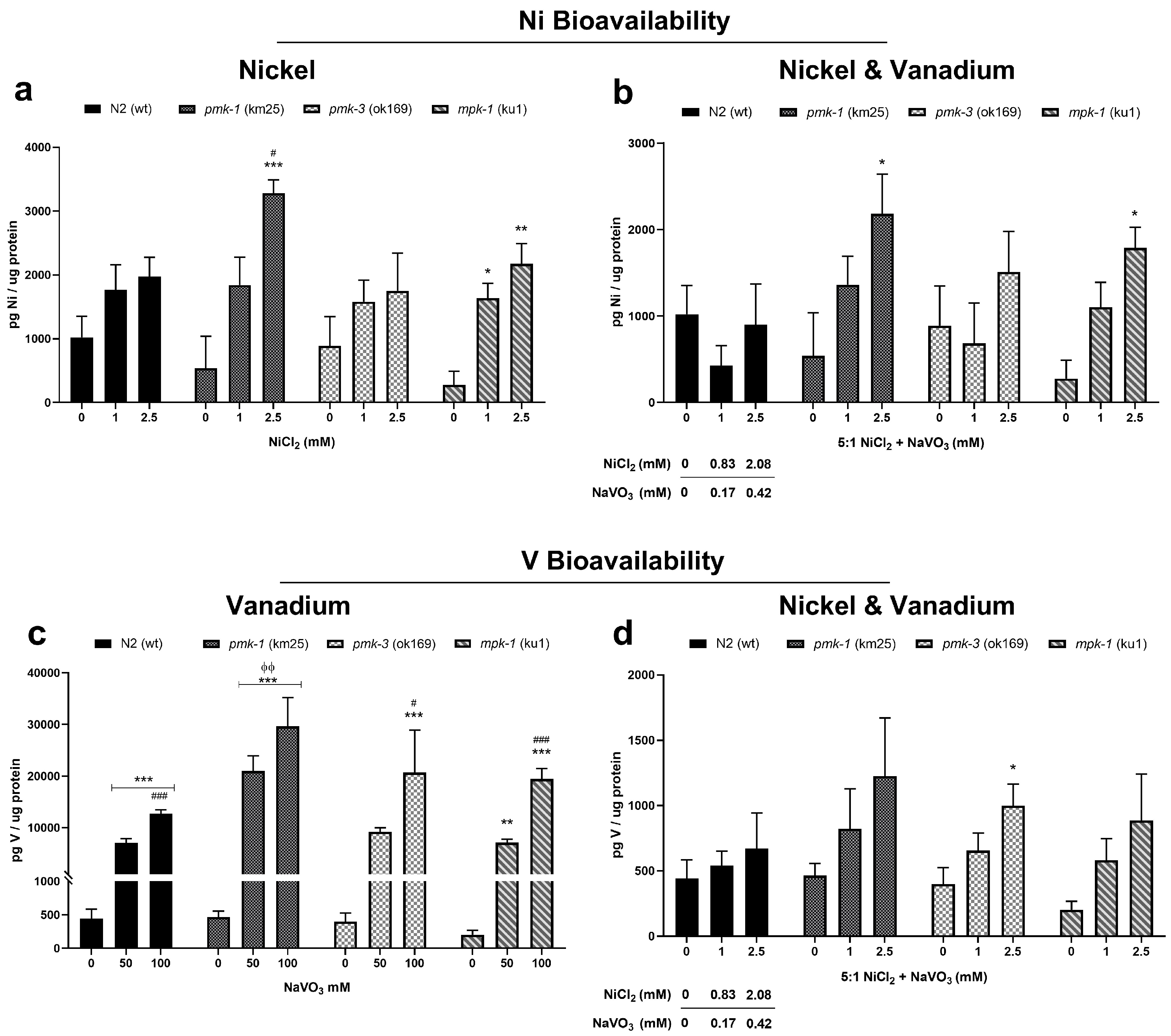
2.2. Metal Bioavailability after Exposures
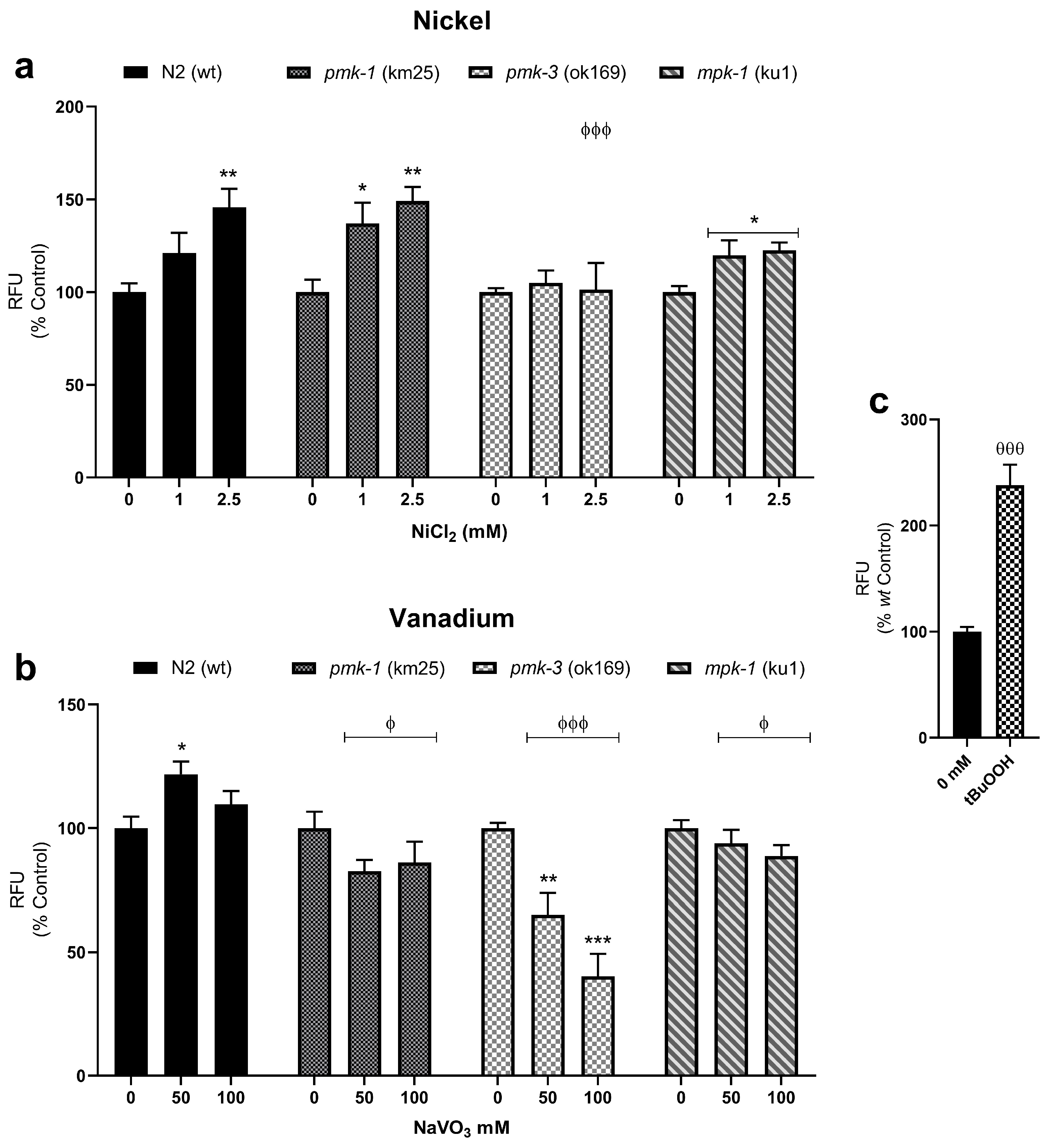
2.3. Changes to Mitochondrial-Derived ROS following Metal Exposures
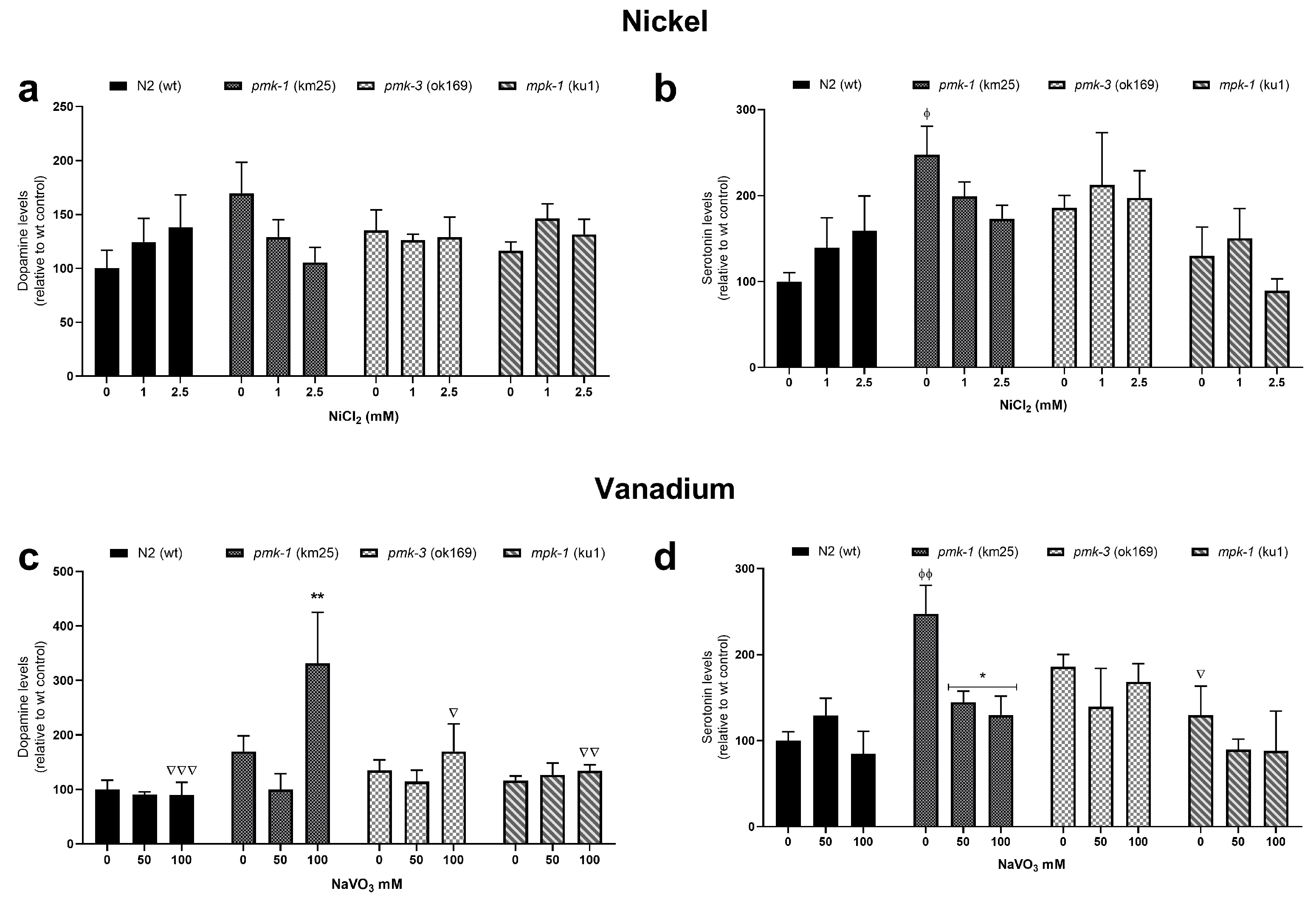
2.4. Dopamine and Serotonin Levels after Metal Treatment
3. Discussion
4. Materials and Methods
4.1. C. elegans Strains and Maintenance
4.2. Preparation of Stock Solutions
4.3. Survival and Developmental Assays
4.4. Metal Bioavailability
4.5. MitoTracker Dyes and Fluorescence Quantification
4.6. Neurotransmitter Measurements
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cicero, C.E.; Mostile, G.; Vasta, R.; Rapisarda, V.; Signorelli, S.S.; Ferrante, M.; Zappia, M.; Nicoletti, A. Metals and neurodegenerative diseases. A systematic review. Environ. Res. 2017, 159, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Amadi, C.N.; Orish, C.N.; Frazzoli, C.; Orisakwe, O.E. Association of autism with toxic metals: A systematic review of case-control studies. Pharmacol. Biochem. Behav. 2022, 212, 173313. [Google Scholar] [CrossRef] [PubMed]
- Dickson, U.J.; Udoessien, E.I. Physicochemical studies of Nigeria’s crude oil blends. Pet. Coal 2012, 54, 243–252. [Google Scholar]
- Fatoba, P.; Ogunkunle, C.; Folarin, O.; Oladele, F. Heavy metal pollution and ecological geochemistry of soil impacted by activities of oil industry in the Niger Delta, Nigeria. Environ. Earth Sci. 2016, 75, 297. [Google Scholar] [CrossRef]
- Adekola, J.; Fischbacher-Smith, M.; Fischbacher-Smith, D.; Adekola, O. Health risks from environmental degradation in the Niger Delta, Nigeria. Environ. Plan. C Politics Space 2017, 35, 334–354. [Google Scholar] [CrossRef]
- Fayiga, A.O.; Ipinmoroti, M.O.; Chirenje, T. Environmental pollution in Africa. Environ. Dev. Sustain. 2018, 20, 41–73. [Google Scholar] [CrossRef]
- ExxonMobil. Crude Oils by Region. Available online: https://corporate.exxonmobil.com/Crude-oils/Crude-trading/Crude-oils-by-region#Africa (accessed on 27 January 2020).
- TOTSA. Crude Assays. Available online: https://www.totsa.com/pub/crude/index2.php?expand=1&iback=1&rub=11&image=africa (accessed on 27 January 2020).
- Ijomone, O.M.; Okori, S.O.; Ijomone, O.K.; Ebokaiwe, A.P. Sub-acute nickel exposure impairs behavior, alters neuronal microarchitecture, and induces oxidative stress in rats’ brain. Drug Chem. Toxicol. 2018, 41, 377–384. [Google Scholar] [CrossRef]
- Ijomone, O.M.; Olatunji, S.Y.; Owolabi, J.O.; Naicker, T.; Aschner, M. Nickel-induced neurodegeneration in the hippocampus, striatum and cortex; an ultrastructural insight, and the role of caspase-3 and α-synuclein. J. Trace Elem. Med. Biol. 2018, 50, 16–23. [Google Scholar] [CrossRef]
- Fatola, O.I.; Olaolorun, F.A.; Olopade, F.E.; Olopade, J.O. Trends in vanadium neurotoxicity. Brain Res. Bull. 2019, 145, 75–80. [Google Scholar] [CrossRef]
- Mustapha, O.; Oke, B.; Offen, N.; Sirén, A.-l.; Olopade, J. Neurobehavioral and cytotoxic effects of vanadium during oligodendrocyte maturation: A protective role for erythropoietin. Environ. Toxicol. Pharmacol. 2014, 38, 98–111. [Google Scholar] [CrossRef]
- Folarin, O.R.; Snyder, A.M.; Peters, D.G.; Olopade, F.; Connor, J.R.; Olopade, J.O. Brain metal distribution and neuro-inflammatory profiles after chronic vanadium administration and withdrawal in mice. Front. Neuroanat. 2017, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Kahloula, K.; Adli, D.E.H.; Slimani, M.; Terras, H.; Achour, S. Effet de l’exposition chronique au nickel sur les fonctions neurocomportementales chez les rats Wistar pendant la période de développement. Effect of nickel chronic exposure on the neurobehavioral functions in Wistar rats during the development period. Toxicol. Anal. Clin. 2014, 26, 186–192. [Google Scholar]
- Sun, L.; Wang, K.; Li, Y.; Fan, Q.; Zheng, W.; Li, H. Vanadium exposure-induced striatal learning and memory alterations in rats. Neurotoxicology 2017, 62, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Ijomone, O.M.; Miah, M.R.; Akingbade, G.T.; Bucinca, H.; Aschner, M. Nickel-Induced Developmental Neurotoxicity in C. elegans Includes Cholinergic, Dopaminergic and GABAergic Degeneration, Altered Behaviour, and Increased SKN-1 Activity. Neurotox. Res. 2020, 37, 1010–1028. [Google Scholar] [CrossRef] [PubMed]
- Berman, K.; McKay, J.; Avery, L.; Cobb, M. Isolation and characterization of pmk-(1–3): Three p38 homologs in Caenorhabditis elegans. Mol. Cell Biol. Res. Commun. 2001, 4, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Samet, J.M.; Graves, L.M.; Quay, J.; Dailey, L.A.; Devlin, R.B.; Ghio, A.J.; Wu, W.; Bromberg, P.A.; Reed, W. Activation of MAPKs in human bronchial epithelial cells exposed to metals. Am. J. Physiol. 1998, 275, L551–L558. [Google Scholar] [CrossRef]
- Cicenas, J.; Zalyte, E.; Rimkus, A.; Dapkus, D.; Noreika, R.; Urbonavicius, S. JNK, p38, ERK, and SGK1 Inhibitors in Cancer. Cancers 2018, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Hendricks, G.; Mylonakis, E. Expanding the nematode model system: The molecular basis of inflammation and infection recovery in C. elegans. Virulence 2017, 8, 244. [Google Scholar] [CrossRef][Green Version]
- Kaminska, B.; Gozdz, A.; Zawadzka, M.; Ellert-Miklaszewska, A.; Lipko, M. MAPK signal transduction underlying brain inflammation and gliosis as therapeutic target. Anat. Rec. Adv. Integr. Anat. Evol. Biol. Adv. Integr. Anat. Evol. Biol. 2009, 292, 1902–1913. [Google Scholar] [CrossRef]
- Moens, U.; Kostenko, S.; Sveinbjørnsson, B. The role of mitogen-activated protein kinase-activated protein kinases (MAPKAPKs) in inflammation. Genes 2013, 4, 101–133. [Google Scholar] [CrossRef]
- Kang, M.; Lee, Y.-S. The impact of RASopathy-associated mutations on CNS development in mice and humans. Mol. Brain 2019, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Layden, M.J.; Johnston, H.; Amiel, A.R.; Havrilak, J.; Steinworth, B.; Chock, T.; Röttinger, E.; Martindale, M.Q. MAPK signaling is necessary for neurogenesis in Nematostella vectensis. BMC Biol. 2016, 14, 61. [Google Scholar] [CrossRef] [PubMed]
- Queirós, L.; Pereira, J.L.; Gonçalves, F.J.M.; Pacheco, M.; Aschner, M.; Pereira, P. Caenorhabditis elegans as a tool for environmental risk assessment: Emerging and promising applications for a "nobelized worm". Crit. Rev. Toxicol. 2019, 49, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.J.; Jarrell, T.A.; Brittin, C.A.; Wang, Y.; Bloniarz, A.E.; Yakovlev, M.A.; Nguyen, K.C.; Tang, L.T.-H.; Bayer, E.A.; Duerr, J.S. Whole-animal connectomes of both Caenorhabditis elegans sexes. Nature 2019, 571, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Lackner, M.R.; Kim, S.K. Genetic analysis of the Caenorhabditis eleganss MAP kinase gene mpk-1. Genetics 1998, 150, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Birdsey, J.M.; Wendolowski, M.A.; Dobbin, K.K.; Williams, P.L. Differential toxicities of nickel salts to the nematode Caenorhabditis elegans. Bull. Environ. Contam. Toxicol. 2016, 97, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Rudel, D.; Douglas, C.D.; Huffnagle, I.M.; Besser, J.M.; Ingersoll, C.G. Assaying environmental nickel toxicity using model nematodes. PLoS ONE 2013, 8, e77079. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Williams, P.L.; Xue, K.S.; Wang, J.-S.; Tang, L. Detoxification mechanisms of nickel sulfate in nematode Caenorhabditis elegans. Chemosphere 2020, 260, 127627. [Google Scholar] [CrossRef]
- Ijomone, O.M. Neurotoxicity of nickel. In Neurotoxicity of Metals: Old Issues and New Developments; Aschner, M., Costa, L.G., Eds.; Advances in Neurotoxicology; Academic Press; Elsevier: Cambridge, MA, USA, 2021; Volume 5, pp. 263–284. [Google Scholar]
- Anyachor, C.P.; Dooka, D.B.; Orish, C.N.; Amadi, C.N.; Bocca, B.; Ruggieri, F.; Senofonte, M.; Frazzoli, C.; Orisakwe, O.E. Mechanistic considerations and biomarkers level in nickel-induced neurodegenerative diseases: An updated systematic review. IBRO Neurosci. Rep. 2022, 13, 136–146. [Google Scholar] [CrossRef]
- Cseh, L.; Ingerman, L.; Keith, S.; Taylor, J. Toxicological Profile for Vanadium; Agency for Toxic Substances and Disease Registry (ATSDR), United States Department of Health and Human Services (HHS): Atlanta, GA, USA, 2012.
- Ghosh, S.K.; Saha, R.; Saha, B. Toxicity of inorganic vanadium compounds. Res. Chem. Intermed. 2015, 41, 4873–4897. [Google Scholar] [CrossRef]
- Chiarelli, R.; Martino, C.; Roccheri, M.C.; Geraci, F. Vanadium Toxicity Monitored by Fertilization Outcomes and Metal Related Proteolytic Activities in Paracentrotus lividus Embryos. Toxics 2022, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Treviño, S.; Díaz, A.; Sánchez-Lara, E.; Sanchez-Gaytan, B.L.; Perez-Aguilar, J.M.; González-Vergara, E. Vanadium in biological action: Chemical, pharmacological aspects, and metabolic implications in diabetes mellitus. Biol. Trace Elem. Res. 2019, 188, 68–98. [Google Scholar] [CrossRef] [PubMed]
- Furimsky, E. On exclusivity of vanadium and nickel porphyrins in crude oil. Energy Fuels 2016, 30, 9978–9980. [Google Scholar] [CrossRef]
- López, L.; Mónaco, S.L. Vanadium, nickel and sulfur in crude oils and source rocks and their relationship with biomarkers: Implications for the origin of crude oils in Venezuelan basins. Org. Geochem. 2017, 104, 53–68. [Google Scholar] [CrossRef]
- Yakubov, M.; Milordov, D.; Yakubova, S.; Borisov, D.; Ivanov, V.; Sinyashin, K. Concentrations of vanadium and nickel and their ratio in heavy oil asphaltenes. Pet. Chem. 2016, 56, 16–20. [Google Scholar] [CrossRef]
- Anyanwu, B.O.; Ezejiofor, A.N.; Igweze, Z.N.; Orisakwe, O.E. Heavy metal mixture exposure and effects in developing nations: An update. Toxics 2018, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Fiati Kenston, S.S.; Su, H.; Li, Z.; Kong, L.; Wang, Y.; Song, X.; Gu, Y.; Barber, T.; Aldinger, J.; Hua, Q. The systemic toxicity of heavy metal mixtures in rats. Toxicol. Res. 2018, 7, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, Y. Nickel sulfate induces numerous defects in Caenorhabditis elegans that can also be transferred to progeny. Environ. Pollut. 2008, 151, 585–592. [Google Scholar] [CrossRef]
- Wilk, A.; Szypulska-Koziarska, D.; Wiszniewska, B. The toxicity of vanadium on gastrointestinal, urinary and reproductive system, and its influence on fertility and fetuses malformations. Adv. Hyg. Exp. Med./Postep. Hig. I Med. Dosw. 2017, 71, 850–859. [Google Scholar] [CrossRef]
- Tuck, S. The control of cell growth and body size in Caenorhabditis elegans. Exp. Cell Res. 2014, 321, 71–76. [Google Scholar] [CrossRef]
- Soete, G.; Betist, M.C.; Korswagen, H.C. Regulation of Caenorhabditis elegans body size and male tail development by the novel gene lon-8. BMC Dev. Biol. 2007, 7, 20. [Google Scholar] [CrossRef]
- Lee, H.-W.; Jose, C.C.; Cuddapah, S. Epithelial-mesenchymal transition: Insights into nickel-induced lung diseases. Semin. Cancer Biol. 2021, 76, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Liu, H.; Jian, Z.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Nickel induces inflammatory activation via NF-κB, MAPKs, IRF3 and NLRP3 inflammasome signaling pathways in macrophages. Aging 2019, 11, 11659. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Chang, Q.; Wang, X.; Son, Y.; Zhang, Z.; Chen, G.; Luo, J.; Bi, Y.; Chen, F.; Shi, X. Reactive oxygen species-activated Akt/ASK1/p38 signaling pathway in nickel compound-induced apoptosis in BEAS 2B cells. Chem. Res. Toxicol. 2010, 23, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Capasso, L.; Camatini, M.; Gualtieri, M. Nickel oxide nanoparticles induce inflammation and genotoxic effect in lung epithelial cells. Toxicol. Lett. 2014, 226, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Ito, Y.; Kosaka, N.; Nukada, Y.; Sakaguchi, H.; Suzuki, H.; Nishiyama, N. Role of MAPK signaling pathway in the activation of dendritic type cell line, THP-1, induced by DNCB and NiSO4. J. Toxicol. Sci. 2008, 33, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Wang, Y.; Zhou, Q.; Bowman, L.; Mao, G.; Zou, B.; Xu, J.; Liu, Y.; Liu, K.; Zhao, J. Inhibition of nickel nanoparticles-induced toxicity by epigallocatechin-3-gallate in JB6 cells may be through down-regulation of the MAPK signaling pathways. PLoS ONE 2016, 11, e0150954. [Google Scholar] [CrossRef]
- Kezhou, C.; Chong, R.; Zengliang, Y. Nickel-induced apoptosis and relevant signal transduction pathways in Caenorhabditis elegans. Toxicol. Ind. Health 2010, 26, 249–256. [Google Scholar] [CrossRef]
- Ingram, J.L.; Rice, A.B.; Santos, J.; Van Houten, B.; Bonner, J.C. Vanadium-induced HB-EGF expression in human lung fibroblasts is oxidant dependent and requires MAP kinases. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2003, 284, L774–L782. [Google Scholar] [CrossRef]
- Chien, P.-S.; Mak, O.-T.; Huang, H.-J. Induction of COX-2 protein expression by vanadate in A549 human lung carcinoma cell line through EGF receptor and p38 MAPK-mediated pathway. Biochem. Biophys. Res. Commun. 2006, 339, 562–568. [Google Scholar] [CrossRef]
- Wang, J.; Huang, X.; Zhang, K.; Mao, X.; Ding, X.; Zeng, Q.; Bai, S.; Xuan, Y.; Peng, H. Vanadate oxidative and apoptotic effects are mediated by the MAPK-Nrf2 pathway in layer oviduct magnum epithelial cells. Metallomics 2017, 9, 1562–1575. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Sun, Y.; Lin, H.; Qian, Y.; Li, Z.; Leonard, S.S.; Huang, C.; Shi, X. Activation of JNK by vanadate induces a Fas-associated death domain (FADD)-dependent death of cerebellar granule progenitors in vitro. J. Biol. Chem. 2003, 278, 4542–4551. [Google Scholar] [CrossRef] [PubMed]
- Zwolak, I. Protective effects of dietary antioxidants against vanadium-induced toxicity: A review. Oxidative Med. Cell. Longev. 2020, 2020, 1490316. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ye, L.; Liu, H.; Xia, Q.; Zhang, Y.; Yang, X.; Wang, K. Vanadium compounds induced mitochondria permeability transition pore (PTP) opening related to oxidative stress. J. Inorg. Biochem. 2010, 104, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Lamtai, M.; Chaibat, J.; Ouakki, S.; Zghari, O.; Mesfioui, A.; El Hessni, A.; Rifi, E.-H.; Marmouzi, I.; Essamri, A.; Ouichou, A. Effect of chronic administration of nickel on affective and cognitive behavior in male and female rats: Possible implication of oxidative stress pathway. Brain Sci. 2018, 8, 141. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Martínez, M.I.; Muñoz-Fambuena, I.; Cauli, O. Neurotransmitters and behavioral alterations induced by nickel exposure. Endocr. Metab. Immune Disord.-Drug Targets (Former. Curr. Drug Targets-Immune Endocr. Metab. Disord.) 2020, 20, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Ijomone, O.M.; Miah, M.R.; Peres, T.V.; Nwoha, P.U.; Aschner, M. Null allele mutants of trt-1, the catalytic subunit of telomerase in Caenorhabditis elegans, are less sensitive to Mn-induced toxicity and DAergic degeneration. Neurotoxicology 2016, 57, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Bornhorst, J.; Ebert, F.; Meyer, S.; Ziemann, V.; Xiong, C.; Guttenberger, N.; Raab, A.; Baesler, J.; Aschner, M.; Feldmann, J. Toxicity of three types of arsenolipids: Species-specific effects in Caenorhabditis elegans. Metallomics 2020, 12, 794–798. [Google Scholar] [CrossRef]
- Nicolai, M.M.; Weishaupt, A.-K.; Baesler, J.; Brinkmann, V.; Wellenberg, A.; Winkelbeiner, N.; Gremme, A.; Aschner, M.; Fritz, G.; Schwerdtle, T. Effects of manganese on genomic integrity in the multicellular model organism Caenorhabditis elegans. Int. J. Mol. Sci. 2021, 22, 10905. [Google Scholar] [CrossRef]
- Neumann, C.; Baesler, J.; Steffen, G.; Nicolai, M.M.; Zubel, T.; Aschner, M.; Bürkle, A.; Mangerich, A.; Schwerdtle, T.; Bornhorst, J. The role of poly (ADP-ribose) polymerases in manganese exposed Caenorhabditis elegans. J. Trace Elem. Med. Biol. 2020, 57, 21–27. [Google Scholar] [CrossRef]
- Schumacher, F.; Chakraborty, S.; Kleuser, B.; Gulbins, E.; Schwerdtle, T.; Aschner, M.; Bornhorst, J. Highly sensitive isotope-dilution liquid-chromatography–electrospray ionization–tandem-mass spectrometry approach to study the drug-mediated modulation of dopamine and serotonin levels in Caenorhabditis elegans. Talanta 2015, 144, 71–79. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ijomone, O.M.; Weishaupt, A.-K.; Michaelis, V.; Ijomone, O.K.; Bornhorst, J. p38- and ERK-MAPK Signalling Modulate Developmental Neurotoxicity of Nickel and Vanadium in the Caenorhabditis elegans Model. Kinases Phosphatases 2024, 2, 28-42. https://doi.org/10.3390/kinasesphosphatases2010003
Ijomone OM, Weishaupt A-K, Michaelis V, Ijomone OK, Bornhorst J. p38- and ERK-MAPK Signalling Modulate Developmental Neurotoxicity of Nickel and Vanadium in the Caenorhabditis elegans Model. Kinases and Phosphatases. 2024; 2(1):28-42. https://doi.org/10.3390/kinasesphosphatases2010003
Chicago/Turabian StyleIjomone, Omamuyovwi M., Ann-Kathrin Weishaupt, Vivien Michaelis, Olayemi K. Ijomone, and Julia Bornhorst. 2024. "p38- and ERK-MAPK Signalling Modulate Developmental Neurotoxicity of Nickel and Vanadium in the Caenorhabditis elegans Model" Kinases and Phosphatases 2, no. 1: 28-42. https://doi.org/10.3390/kinasesphosphatases2010003
APA StyleIjomone, O. M., Weishaupt, A.-K., Michaelis, V., Ijomone, O. K., & Bornhorst, J. (2024). p38- and ERK-MAPK Signalling Modulate Developmental Neurotoxicity of Nickel and Vanadium in the Caenorhabditis elegans Model. Kinases and Phosphatases, 2(1), 28-42. https://doi.org/10.3390/kinasesphosphatases2010003