
Root and Shoot Biomass Contributions to Soil Carbon and Nitrogen Under Grazing Intensity and Crop Rotation in an Integrated Crop–Livestock System

 ,
, 
 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Site Description and Characterization of the Experimental Area
2.2. Experimental Design and Conduction of the Experiment
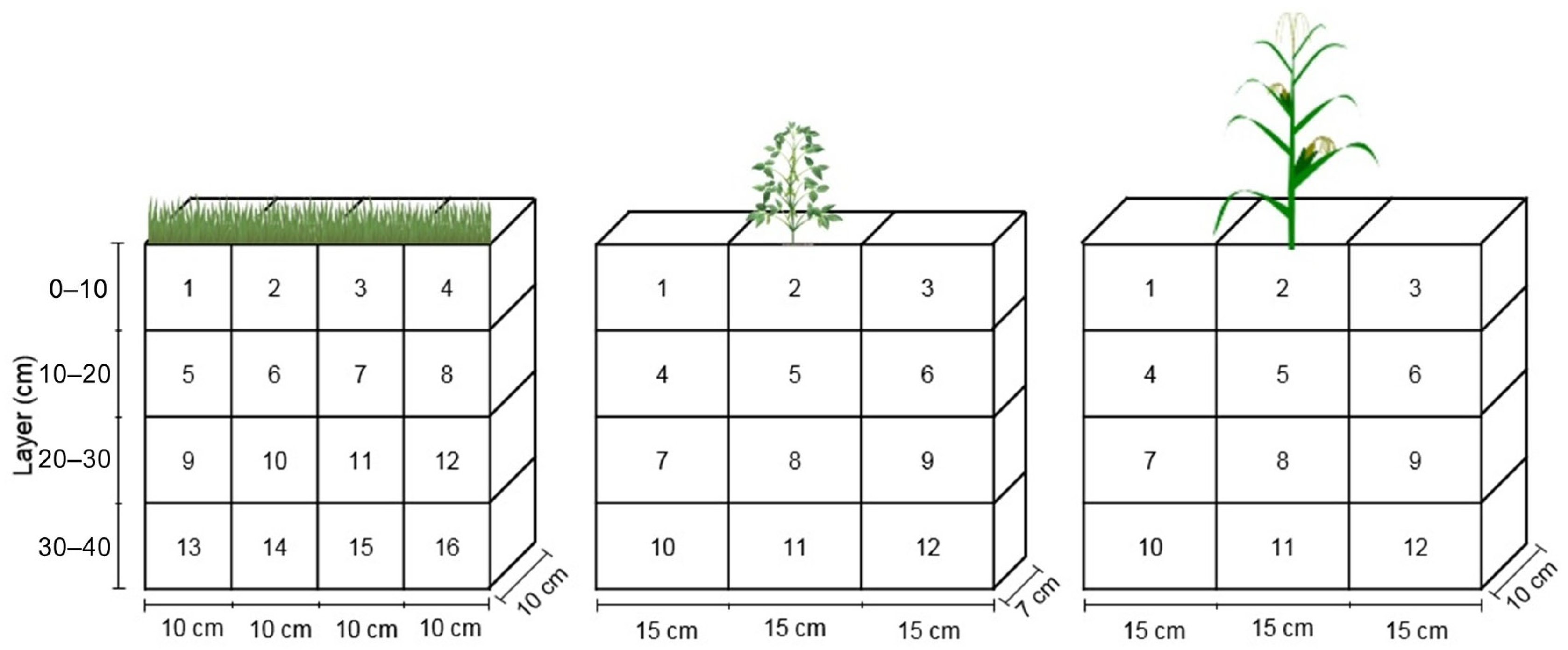
2.3. Root and Shoot Biomass Determination and C and N Inputs
2.4. Statistical Analysis
3. Results
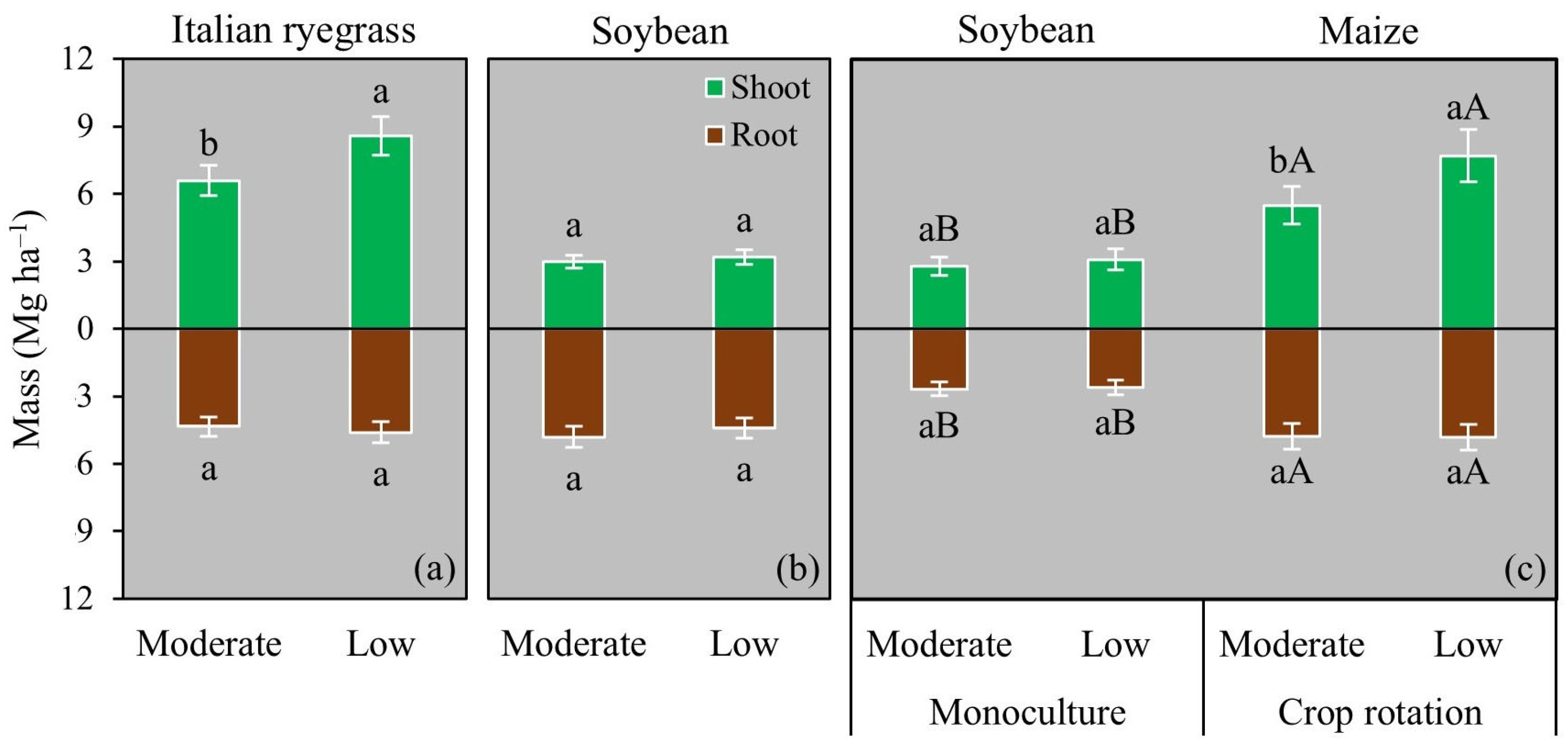
3.1. Root and Shoot Biomass
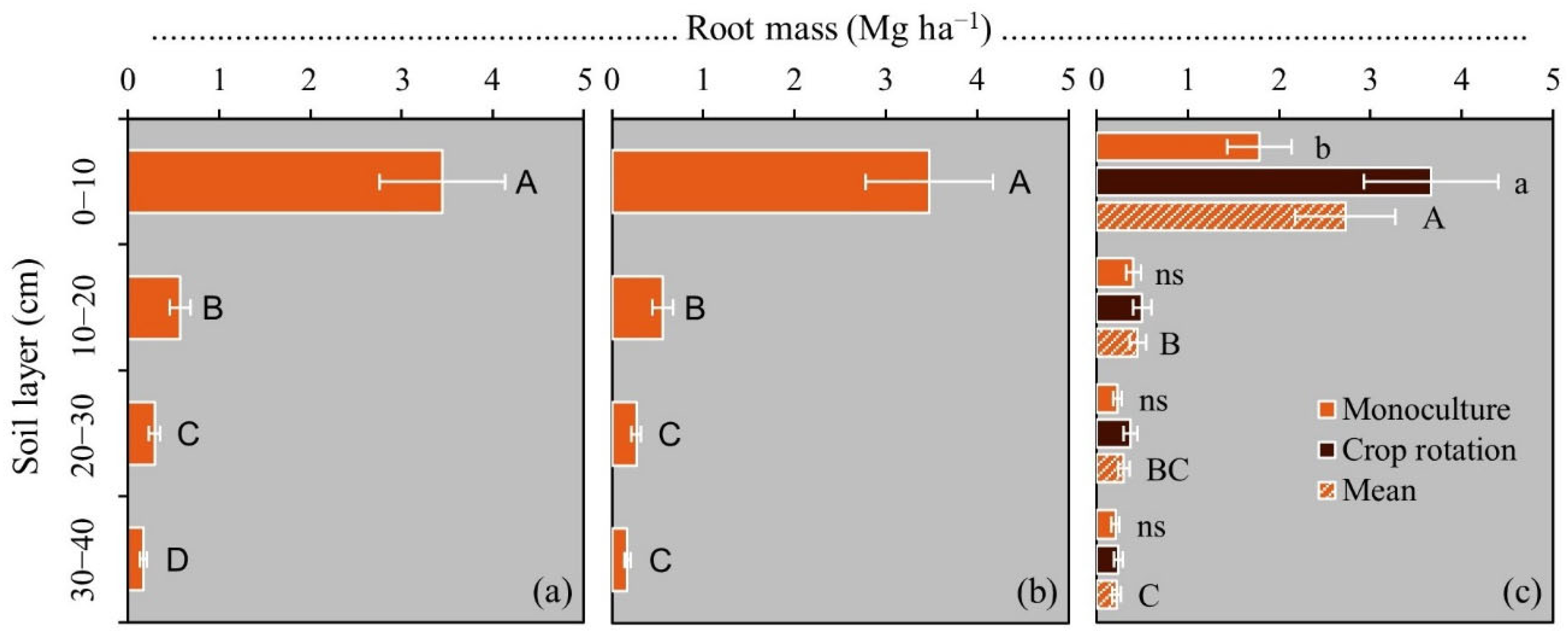
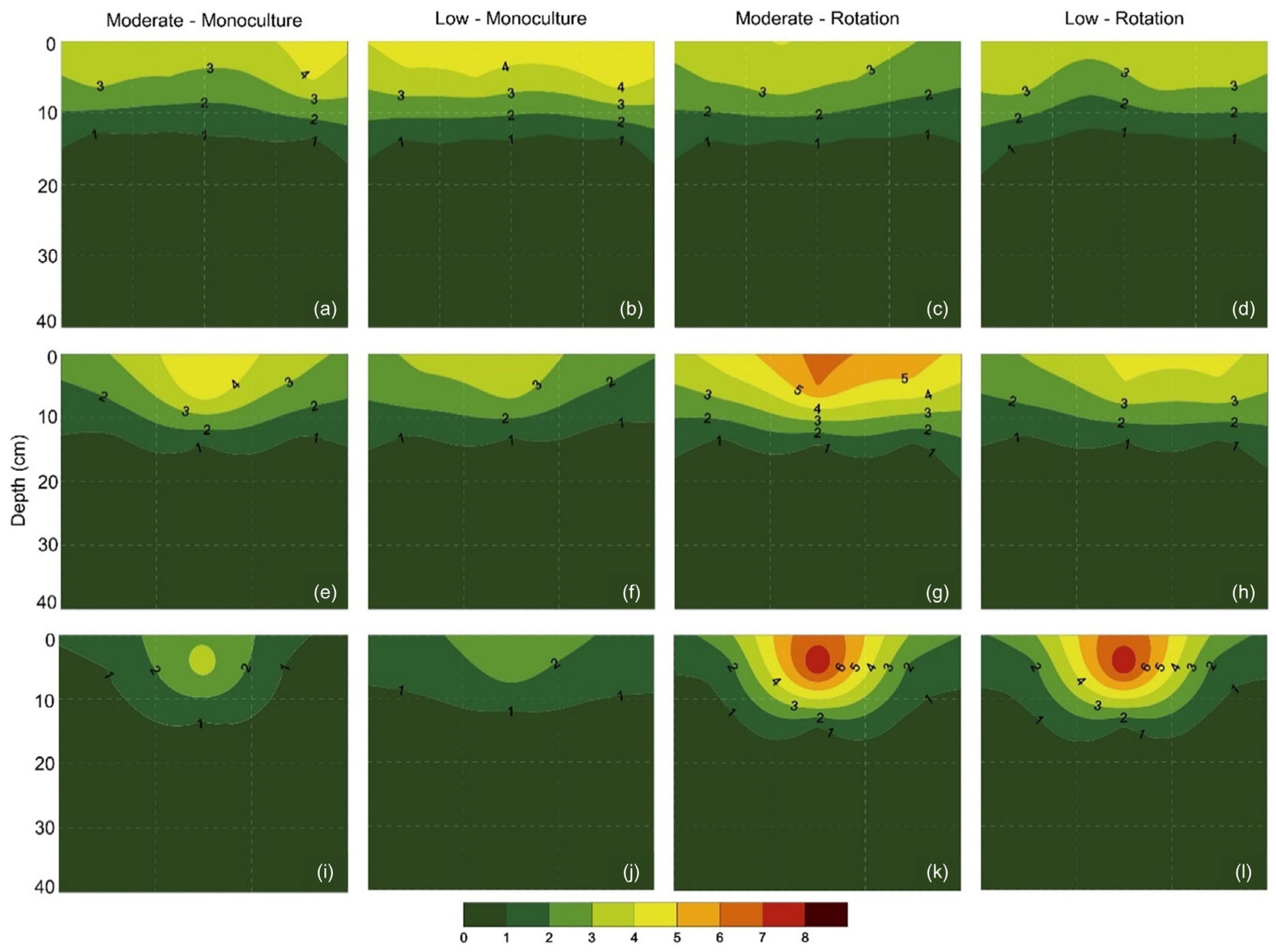
3.2. Root Distribution in Soil Profile
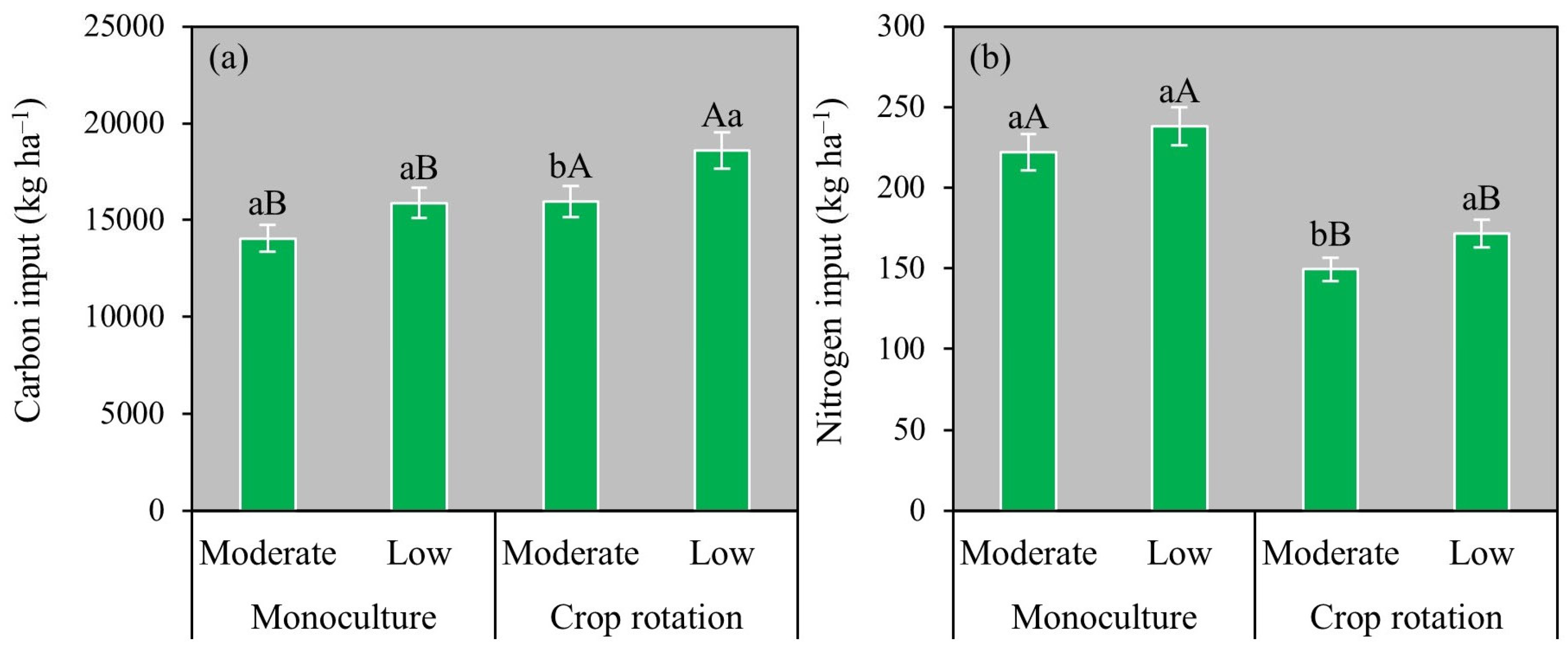
3.3. C and N Inputs in the Systems
4. Discussion
4.1. Importance of Root Biomass as a Carbon Source
4.2. Effects of Grazing Intensity and Crop Rotation on Biomass Production and Soil C Sequestration in ICLSs
4.3. Nitrogen Inputs and C/N Ratios in Different Cropping Systems
4.4. Root Distribution in the Soil Profile and Its Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moraes, A.; Carvalho, P.C.F.; Anghinoni, I.; Lustosa, S.B.C.; Costa, S.E.V.G.A.; Kunrath, T.R. Integrated crop-livestock systems in the Brazilian subtropics. Eur. J. Agron. 2014, 57, 4–9. [Google Scholar] [CrossRef]
- Salton, J.C.; Mercante, F.M.; Tomazi, M.; Zanatta, J.A.; Concenço, G.; Silva, W.M.; Retore, M. Integrated crop-livestock system in tropical Brazil: Toward a sustainable production system. Agric. Ecosyst. Environ. 2014, 190, 70–79. [Google Scholar] [CrossRef]
- Bieluczyk, W.; Piccolo, M.C.; Pereira, M.G.; Moraes, M.T.; Soltangheisi, A.; Bernardi, A.C.C.; Pezzopane, J.R.M.; Oliveira, P.P.A.; Moreira, M.Z.; Camargo, P.B.; et al. Integrated farming systems influence soil organic matter dynamics in southeastern Brazil. Geoderma 2020, 371, 114368. [Google Scholar] [CrossRef]
- Mori, L.D.; Simões, V.J.L.P.; Cargnelutti, C.S.; Duarte, L.P.; Leal, G.L.; Doberstein, A.P.S.; Kunrath, T.S.; Nunes, P.A.A.; Souza, E.D.; Bayer, C.; et al. Grazing intensity for enhanced resource use efficiency in integrated crop-livestock systems: Balancing soil carbon storage and food security. J. Environ. Manag. 2025, 373, 123541. [Google Scholar] [CrossRef]
- Bayer, C.; Lovato, T.; Dieckow, J.; Zanatta, J.A.; Mielniczuk, J. A method for estimating coefficients of soil organic matter dynamics based on long-term experiments. Soil Tillage Res. 2006, 91, 217–226. [Google Scholar] [CrossRef]
- Silva, F.D.; Amado, T.J.C.; Ferreira, A.O.; Assmann, J.M.; Anghinoni, I.; Carvalho, P.C.F. Soil carbon indices as affected by 10 years of integrated crop–livestock production with different pasture grazing intensities in Southern Brazil. Agric. Ecosyst. Environ. 2014, 190, 60–69. [Google Scholar] [CrossRef]
- Zhou, G.; Zhou, X.; He, Y.; Shao, J.; Hu, Z.; Liu, R.; Zhou, H.; Hosseinibai, S. Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: A meta-analysis. Glob. Change Biol. 2017, 23, 1167–1179. [Google Scholar] [CrossRef]
- Ribeiro, R.H.; Dieckow, J.; Piva, J.T.; Bratti, F. Roots and aboveground carbon and nitrogen inputs by black oats (Avena strigosa Schreb.) as affected by grazing and nitrogen in integrated crop-livestock system in subtropical Brazil. Plant Soil 2020, 451, 447–458. [Google Scholar] [CrossRef]
- Ribeiro, R.H.; Ibarr, M.A.; Besen, M.R.; Bayer, C.; Piva, J.T. Managing grazing intensity to reduce the global warming potential in integrated crop–livestock systems under no-till agriculture. Eur. J. Soil Sci. 2019, 70, 1–12. [Google Scholar] [CrossRef]
- Souza, E.D.; Costa, S.E.V.G.A.; Lima, C.V.S.; Anghinoni, I.; Meurer, E.J.; Carvalho, P.C.F. Carbono orgânico e fósforo microbiano em sistemas de integração agricultura-pecuária submetidos a diferentes intensidades de pastejo em plantio direto. Rev. Bras. Cienc. Solo 2008, 32, 1273–1282. [Google Scholar] [CrossRef]
- Yang, F.; Liao, D.; Wu, X.; Gao, R.; Fan, Y.; Raza, M.A.; Wang, X.; Young, T.; Liu, W.; Liu, J.; et al. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercropping systems. Field Crops Res. 2017, 203, 16–23. [Google Scholar] [CrossRef]
- Alves, L.A.; Denardin, L.G.O.; Martins, A.P.; Bayer, C.; Veloso, M.G.; Bremm, C.; Carvalho, P.C.F.; Machado, D.R.; Tiecher, T. The effect of crop rotation and sheep grazing management on plant production and soil C and N stocks in a long-term integrated crop-livestock system in Southern Brazil. Soil Tillage Res. 2020, 203, 104678. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Change Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–781. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Moraes, G.; Leonardo, J.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- FAO. World reference base for soil resources 2014. In World Soil Resources Reports; FAO: Rome, Italy, 2014; Volume 106. Available online: https://www.fao.org/3/i3794en/I3794en.pdf (accessed on 20 May 2025).
- Comissão de Fertilidade do Solo—RS/SC. Recomendações de Adubação e de Calagem para os Estados do Rio Grande do Sul e de Santa Catarina, 3rd ed.; SBCS-Núcleo Regional Sul/EMBRAPA-CNPT: Passo Fundo, Brazil, 1995; p. 224. [Google Scholar]
- Mott, G.O.; Lucas, H.L. The design conduct and interpretation of grazing trials on cultivated and improved pastures. In Proceedings of the 6th International Grassland Congress, State College, PA, USA, 17–23 August 1952; pp. 1380–1395. [Google Scholar]
- National Research Council. Nutrient Requirement of Sheep, 6th ed.; National Academy of Science: Washington, DC, USA, 1985; p. 99. [Google Scholar]
- Alves, L.A.; Denardin, L.G.O.; Martins, A.P.; Anghinoni, I.; Carvalho, P.C.F.; Tiecher, T. Soil acidification and P, K, Ca and Mg budget as affected by sheep grazing and crop rotation in a long-term integrated crop-livestock system in southern Brazil. Geoderma 2019, 351, 197–208. [Google Scholar] [CrossRef]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of development descriptions for soybeans, Glycine Max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Freschet, G.T.; Pagès, L.; Iversen, C.M.; Comas, L.H.; Rewald, B.; Roumet, C.; Klimešová, J.; Zadworny, M.; Poorter, H.; Postma, J.A.; et al. A starting guide to root ecology: Strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements. New Phytol. 2021, 232, 973–1122. [Google Scholar] [CrossRef]
- Mazzilli, S.R.; Kemanian, A.R.; Ernst, O.R.; Jackson, R.B.; Piñeiro, G. Priming of soil organic carbon decomposition induced by corn compared to soybean crops. Soil Biol. Biochem. 2014, 75, 273–281. [Google Scholar] [CrossRef]
- Redin, M.; Guénon, R.; Recous, S.; Schmatz, R.; Freitas, L.L.; Aita, C.; Giacomini, S.J. Carbon mineralization in soil of roots from twenty crop species, as affected by their chemical composition and botanical family. Plant Soil 2014, 378, 205–214. [Google Scholar] [CrossRef]
- Bai, Y.; Cotrufo, M.F. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 2022, 377, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Kätterer, T.; Bolinder, M.A.; Andrén, O.; Kirchmann, H.; Menichetti, L. Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric. Ecosyst. Environ. 2011, 141, 184–192. [Google Scholar] [CrossRef]
- Bolinder, M.A.; Kätterer, T.; Andrén, O.; Parent, L.E. Estimating carbon inputs to soil in forage-based crop rotations and modeling the effects on soil carbon dynamics in a Swedish long-term field experiment. Can. J. Soil Sci. 2012, 92, 821–833. [Google Scholar] [CrossRef]
- Li, X.; An, C.; Li, X.; Wu, Q.; Chen, X.; Guo, J.; Gao, T.; Wang, H.; Dong, Z. Impacts of grazing intensity on soil properties and carbon content in Xilamuren Grassland. J. Environ. Manag. 2025, 386, 125773. [Google Scholar] [CrossRef]
- Zhao, T.; Bisseling, T.; Yang, Y.; Zhao, M.; Zhang, B.; Han, G. Soil carbon mineralization decreased in desert steppe by light grazing but not fencing management. Catena 2024, 245, 108321. [Google Scholar] [CrossRef]
- Bonetti, J.A.; Paulino, H.B.; Souza, E.D.; Carneiro, M.A.C.; Silva, G.N. Influência do sistema integrado de produção agropecuária no solo e na produtividade de soja e braquiária. Pesq. Agropec. Trop. 2015, 45, 104–112. [Google Scholar] [CrossRef]
- Chakraborty, P.; Thotakuri, G.; Singh, N.; Dhaliwal, J.K.; Kumar, S. Crop-livestock integration influenced soil profile organic carbon and hydro-physical properties in converted grasslands to row crops. Soil Tillage Res. 2024, 240, 106093. [Google Scholar] [CrossRef]
- Augusto, J.G.; Paz, C.C.P.; Mendonça, G.G.; Moitinho, M.R.; Menegatto, L.S.; Bolonhezi, D.; Salles, M.S.V.; Simili, F.F. Integrated crop-livestock versus conventional systems: Effects on the chemical and physical characteristics of an Oxisol. Grass Forage Sci. 2024, 79, 254–264. [Google Scholar] [CrossRef]
- Sekaran, U.; Kumar, S.; Gonzalez-Hernandez, J.L. Integration of crop and livestock enhanced soil biochemical properties and microbial community structure. Geoderma 2021, 381, 114686. [Google Scholar] [CrossRef]
- Bonetti, J.A.; Anghinoni, I.; Moraes, M.T.; Fink, J.R. Resilience of soils with different texture, mineralogy and organic matter under long-term conservation systems. Soil Tillage Res. 2017, 174, 104–112. [Google Scholar] [CrossRef]
- Xia, H.Y.; Zhao, J.H.; Sun, J.H.; Bao, X.G.; Christie, P.; Zhang, F.S.; Li, L. Dynamics of root length and distribution and shoot biomass of maize as affected by intercropping with different companion crops and phosphorus application rates. Field Crop. Res. 2013, 150, 52–62. [Google Scholar] [CrossRef]
- Ordóñez, R.A.; Archontoulis, S.V.; Martinez-Feria, R.; Hatfield, J.L.; Wright, E.E.; Castellano, M.J. Root to shoot and carbon to nitrogen ratios of maize and soybean crops in the US Midwest. Eur. J. Agron. 2020, 120, 126130. [Google Scholar] [CrossRef]
- Manzoni, S.; Jackson, R.B.; Trofymow, J.A.; Porporato, A. The global stoichiometry of litter nitrogen mineralization. Science 2008, 321, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Veloso, M.G.; Cecagno, D.; Bayer, C. Legume cover crops under no-tillage favor organomineral association in microaggregates and soil C accumulation. Soil Tillage Res. 2019, 190, 139–146. [Google Scholar] [CrossRef]
- Li, H.; Mollier, A.; Ziadi, N.; Shi, Y.; Parent, L.É.; Morel, C. Soybean root traits after 24 years of different soil tillage and mineral phosphorus fertilization management. Soil Tillage Res. 2017, 165, 258–267. [Google Scholar] [CrossRef]
- Nichols, V.A.; Ordonez, R.A.; Wright, E.E.; Castellano, M.J.; Liebman, M.; Hatfield, J.L.; Helmers, M.; Archontoulis, S.V. Maize root distributions strongly associated with water tables in Iowa, USA. Plant Soil 2019, 444, 225–238. [Google Scholar] [CrossRef]
- Liedgens, M.; Soldati, A.; Stamp, P. Interactions of maize and Italian ryegrass in a living mulch system: (1) Shoot growth and rooting patterns. Plant Soil 2004, 262, 191–203. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, L.A.; Arnuti, F.; Oliveira, L.B.d.; Moraes, M.T.d.; Denardin, L.G.d.O.; Anghinoni, I.; Carvalho, P.C.d.F.; Tiecher, T. Root and Shoot Biomass Contributions to Soil Carbon and Nitrogen Under Grazing Intensity and Crop Rotation in an Integrated Crop–Livestock System. Grasses 2025, 4, 24. https://doi.org/10.3390/grasses4020024
Alves LA, Arnuti F, Oliveira LBd, Moraes MTd, Denardin LGdO, Anghinoni I, Carvalho PCdF, Tiecher T. Root and Shoot Biomass Contributions to Soil Carbon and Nitrogen Under Grazing Intensity and Crop Rotation in an Integrated Crop–Livestock System. Grasses. 2025; 4(2):24. https://doi.org/10.3390/grasses4020024
Chicago/Turabian StyleAlves, Lucas Aquino, Fernando Arnuti, Leandro Bittencourt de Oliveira, Moacir Tuzzin de Moraes, Luiz Gustavo de Oliveira Denardin, Ibanor Anghinoni, Paulo César de Faccio Carvalho, and Tales Tiecher. 2025. "Root and Shoot Biomass Contributions to Soil Carbon and Nitrogen Under Grazing Intensity and Crop Rotation in an Integrated Crop–Livestock System" Grasses 4, no. 2: 24. https://doi.org/10.3390/grasses4020024
APA StyleAlves, L. A., Arnuti, F., Oliveira, L. B. d., Moraes, M. T. d., Denardin, L. G. d. O., Anghinoni, I., Carvalho, P. C. d. F., & Tiecher, T. (2025). Root and Shoot Biomass Contributions to Soil Carbon and Nitrogen Under Grazing Intensity and Crop Rotation in an Integrated Crop–Livestock System. Grasses, 4(2), 24. https://doi.org/10.3390/grasses4020024