
Detection of Hepatitis E Virus Infections in Wild Boars in Southwest Germany Using a Stepwise Laboratory Diagnostic Approach


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collections
2.2. Serological Analyses
2.3. RNA Extraction and RT-qPCR
2.4. HEV RNA Genotyping
2.5. Statistical Analyses
3. Results
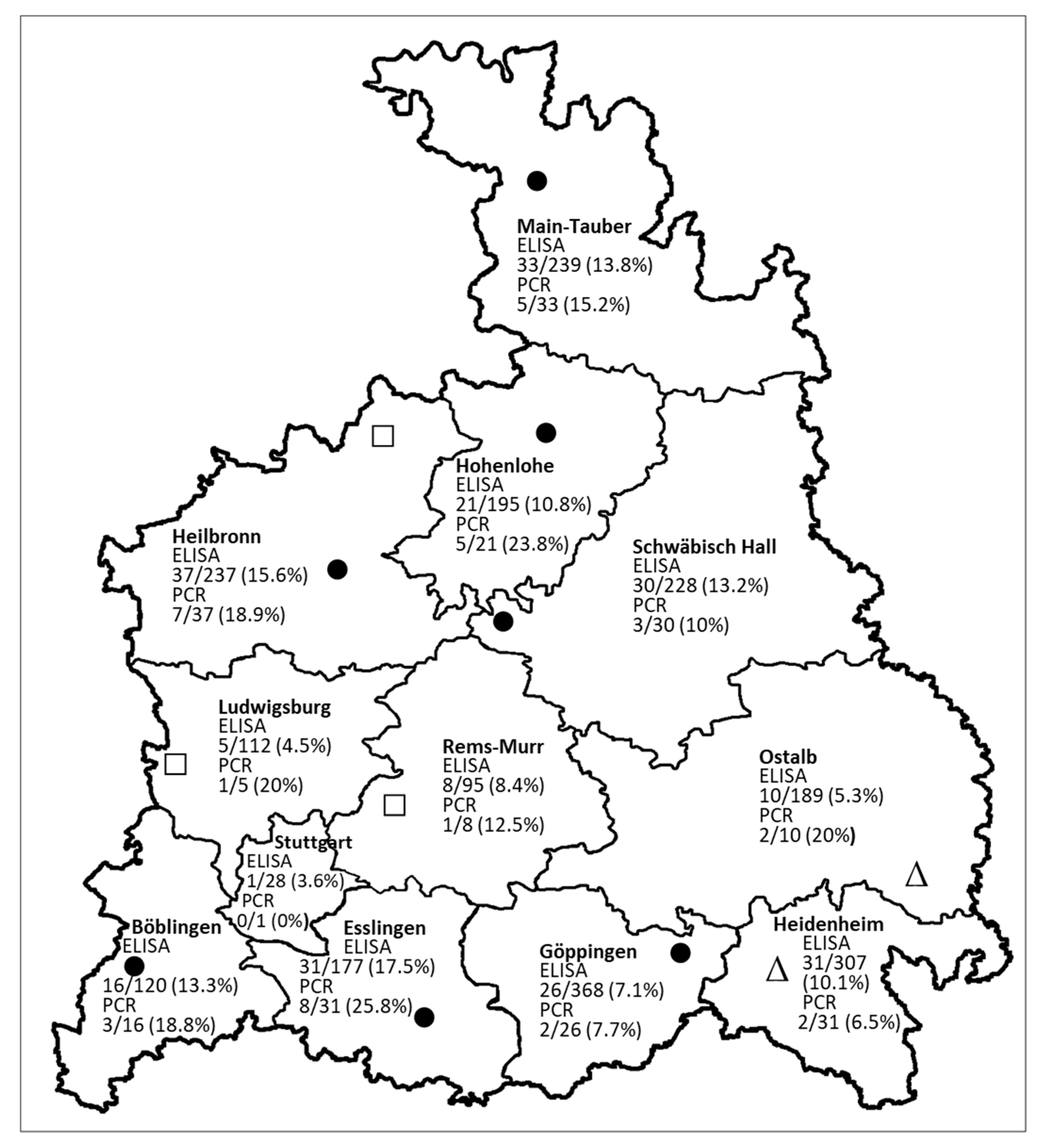
3.1. Serological and RT-qPCR Studies
3.2. Genotyping
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clemente-Casares, P.; Ramos-Romero, C.; Ramirez-Gonzalez, E.; Mas, A. Hepatitis E Virus in Industrialized Countries: The Silent Threat. Biomed. Res. Int. 2016, 2016, 9838041. [Google Scholar] [CrossRef]
- Pérez-Gracia, M.T.; Garcia, M.; Suay, B.; Mateos-Lindemann, M.L. Current knowledge on hepatitis E. J. Clin. Transl. Hepatol. 2015, 3, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Treagus, S.; Wright, C.; Baker-Austin, C.; Longdon, B.; Lowther, J. The foodborne transmission of hepatitis E virus to humans. Food Environ. Virol. 2021, 13, 127–145. [Google Scholar] [CrossRef]
- Faber, M.; Askar, M.; Stark, K. Case-control study on risk factors for acute hepatitis E in Germany, 2012 to 2014. Eurosurveillance 2018, 23, 469. [Google Scholar] [CrossRef]
- Pischke, S.; Behrendt, P.; Bock, C.T.; Jilg, W.; Manns, M.P.; Wedemeyer, H. Hepatitis E in Germany—An under-reported infectious disease. Deutsches Ärzteblatt Int. 2014, 111, 577–583. [Google Scholar]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Escamez, P.S.F.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; Nørrung, B.; et al. Scientific Opinion on the public health risks associated with hepatitis E virus (HEV) as a food-borne pathogen. EFSA J. 2017, 15, e04886. [Google Scholar] [CrossRef]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic Hepatitis E Virus: Classification, animal reservoirs and transmission routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef]
- Sridhar, S.; Teng, J.L.L.; Chiu, T.-H.; Lau, S.K.P.; Woo, P.C.Y. Hepatitis E Virus genotypes and evolution: Emergence of camel hepatitis E variants. Int. J. Mol. Sci. 2017, 18, 869. [Google Scholar] [CrossRef] [Green Version]
- Brost, S.; Wenzel, J.J.; Ganten, T.M.; Filser, M.; Flechtenmacher, C.; Boehm, S.; Astanief, A.; Jilg, W.; Zeier, M.; Schnitzler, P. Sporadic cases of acute autochthonous hepatitis E virus infection in Southwest Germany. J. Clin. Virol. 2010, 47, 89–92. [Google Scholar] [CrossRef]
- Preiss, J.C.; Plentz, A.; Engelmann, E.; Schneider, T.; Jilg, W.; Zeitz, M.; Duchmann, R. Autochthonous hepatitis E virus infection in Germany with sequence similarities to other European isolates. Infection 2006, 34, 173–175. [Google Scholar] [CrossRef]
- Veitt, R.; Reichardt, M.; Wenzel, J.; Jilg, W. Autochthonous hepatitis E-virus infection as cause of acute hepatitis in Germany—A case report. Z. Gastroenterol. 2011, 49, 42–46. [Google Scholar] [CrossRef]
- Wichmann, O.; Schimanski, S.; Koch, J.; Kohler, M.; Rothe, C.; Plentz, A.; Jilg, W.; Stark, K. Phylogenetic and case-control study on hepatitis E virus infection in Germany. J. Infect. Dis. 2008, 198, 1732–1741. [Google Scholar] [CrossRef] [Green Version]
- Kozyra, I.; Jabłonski, A.; Bigoraj, E.; Rzezutka, A. Wild Boar as a sylvatic reservoir of hepatitis E virus in Poland: A cross-sectional population study. Viruses 2020, 12, 1113. [Google Scholar] [CrossRef]
- Ferri, G.; Vergara, A. Hepatitis E virus in the food of animal origin: A review. Foodborne Pathog. Dis. 2021, 18, 368–377. [Google Scholar] [CrossRef]
- Kaci, S.; Nöckler, K.; Johne, R. Detection of hepatitis E virus in archived German wild boar serum samples. Vet. Microbiol. 2008, 128, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Li, C.; Hagedorn, C.H. Phylogenetic analysis of global hepatitis E virus sequences: Genetic diversity, subtypes and zoonosis. Rev. Med. Virol. 2006, 16, 5–36. [Google Scholar] [CrossRef]
- Schielke, A.; Sachs, K.; Lierz, M.; Appel, B.; Jansen, A.; Johne, R. Detection of hepatitis E virus in wild boars of rural and urban regions in Germany and whole genome characterization of an endemic strain. Virol. J. 2009, 6, 58. [Google Scholar] [CrossRef] [Green Version]
- Anheyer-Behmenburg, H.E.; Szabo, K.; Schotte, U.; Binder, A.; Klein, G.; Johne, R. Hepatitis E virus in wild boars and spillover infection in red and roe deer, Germany, 2013–2015. Emerg. Infect. Dis. 2017, 23, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Li, T.C.; Chijiwa, K.; Sera, N.; Ishibashi, T.; Etoh, Y.; Shinohara, Y.; Kurata, Y.; Ishida, M.; Sakamoto, S.; Takeda, N.; et al. Hepatitis E virus transmission from wild boar meat. Emerg. Infect. Dis. 2005, 11, 1958–1960. [Google Scholar] [CrossRef]
- Schielke, A.; Ibrahim, V.; Czogiel, I.; Faber, M.S.; Schrader, C.; Dremsek, P.; Ulrich, R.G.; Johne, R. Hepatitis E virus antibody prevalence in hunters from a district in Central Germany, 2015: A cross-sectional study providing evidence for the benefit of protective gloves during disemboweling of wild boars. BMC Infect. Dis. 2015, 15, 440. [Google Scholar] [CrossRef] [Green Version]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of hepatitis E virus transmission to humans. J. Infect. Dis. 2020, 202, 825–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallerla, S.R.; Schembecker, S.; Meyer, C.G.; Linh, L.T.K.; Johne, R.; Wedemeyer, H.; Bock, C.-T.; Kremsner, P.G.; Velavan, T.P. Hepatitis E virus genome detection in commercial pork livers and pork meat products in Germany. J. Viral Hepat. 2021, 28, 196–204. [Google Scholar] [CrossRef]
- Said, B.; Ijaz, S.; Chand, M.A.; Kafatos, G.; Tedder, R.; Morgan, D. Hepatitis E virus in England and Wales: Indigenous infection is associated with the consumption of processed pork products. Epidemiol. Infect. 2014, 142, 1467–1475. [Google Scholar] [CrossRef]
- Adlhoch, C.; Wolf, A.; Meisel, H.; Kaiser, M.; Ellerbrok, H.; Pauli, G. High HEV presence in four different wild boar populations in East and West Germany. Vet. Microbiol. 2009, 139, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, H.C.; Wichmann, O.; Duizer, E. Transmission routes and risk factors for autochthonous hepatitis E virus infection in Europe: A systematic review. Epidemiol. Infect. 2010, 138, 145–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutjes, S.A.; Lodder-Verschoor, F.; Lodder, W.J.; van der Giessen, J.; Reesink, H.; Bouwknegt, M.; Husman, A.M.D.R. Seroprevalence and molecular detection of hepatitis E virus in wild boar and red deer in The Netherlands. J. Virol. Methods 2010, 168, 197–206. [Google Scholar] [CrossRef]
- Strakova, P.; Kubankova, M.; Vasickova, P.; Juricova, Z.; Rudolf, I.; Hubalek, Z. Hepatitis E virus in archived sera from wild boars (Sus scrofa), Czech Republic. Transbound. Emerg. Dis. 2018, 65, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, A.; Chaussade, H.; Rigaud, E.; Rodriguez, J.; Berthault, C.; Boué, F.; Tognon, M.; Touzé, A.; Garcia-Bonnet, N.; Choutet, P.; et al. High hepatitis E virus seroprevalence in forestry workers and in wild boars in France. J. Clin. Microbiol. 2012, 50, 2888–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burri, C.; Vial, F.; Ryser-Degiorgis, M.P.; Schwermer, H.; Darling, K.; Reist, M.; Wu, N.; Beerli, O.; Schöning, J.; Cavassini, M.; et al. Seroprevalence of hepatitis E virus in domestic pigs and wild boars in Switzerland. Zoonoses Public Health 2014, 61, 537–544. [Google Scholar] [CrossRef] [Green Version]
- BMJV (Bundesministerium der Justiz und für Verbraucherschutz Deutschland). Verordnung zur Durchführung eines Monitorings auf das Virus der Klassischen und der Afrikanischen Schweinepest bei Wild und Hausschweinen (Schweinepest-Monitoring-Verordnung (SchwPestMonV); BMJV: Berlin, Germany, 2013; p. 2518. [Google Scholar]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Trojnar, E.; Contzen, M.; Moor, D.; Carl, A.; Burkhardt, S.; Kilwinski, J.; Berghof-Jäger, K.; Mormann, S.; Schotte, U.; Kontek, A.; et al. Interlaboratory Validation of a Detection Method for Hepatitis E Virus RNA in pig Liver. Microorganisms 2020, 8, 1460. [Google Scholar] [CrossRef] [PubMed]
- Vina-Rodriguez, A.; Schlosser, J.; Becher, D.; Kaden, V.; Groschup, M.H.; Eiden, M. Hepatitis E virus genotype 3 diversity: Phylogenetic analysis and presence of subtype 3b in wild boar in Europe. Viruses 2015, 7, 2704–2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, A.G.; Sullivan, K.M.; Soe, M.M. OpenEpi: Open Source Epidemiologic Statistics for Public Health, version 3.01, updated 6 April 2013. 2013. Available online: www.OpenEpi.com (accessed on 13 October 2021).
- United Nations Environment Programme and International Livestock Research Institute Preventing the Next Pandemic: Zoonotic Diseases and how to Break the Chain of Transmission. Nairobi, Kenya, 2000. Available online: https://www.unep.org/resources/report/preventing-future-zoonotic-disease-outbreaks-protecting-environment-animals-and (accessed on 2 July 2021).
- Modiyinji, A.F.; Sanding, G.M.A.M.; Atsama, M.A.; Monamele, C.G.; Nola, M.; Njouom, R. Serological and molecular investigation of hepatitis E virus in pigs reservoirs from Cameroon reveals elevated seroprevalence and presence of genotype 3. PLoS ONE 2020, 15, e0229073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Deus, N.; Peralta, B.; Pina, S.; Allepuz, A.; Mateu, E.; Vidal, D.; Ruiz-Fons, F.; Martin, M.; Gortazar, C.; Segales, J. Epidemiological study of hepatitis E virus infection in European wild boars (Sus scrofa) in Spain. Vet. Microbiol. 2008, 129, 163–170. [Google Scholar] [CrossRef]
- Halbur, P.G.; Kasorndorkbua, C.; Gilbert, C.; Guenette, D.; Potters, M.B.; Purcell, R.H.; Emerson, S.U.; Toth, T.E.; Meng, X.J. Comparative pathogenesis of infection of pigs with hepatitis E viruses recovered from a pig and a human. J. Clin. Microbiol. 2001, 39, 918–923. [Google Scholar] [CrossRef] [Green Version]
- Seminati, C.; Mateu, E.; Peralta, B.; de Deus, N.; Martin, M. Distribution of hepatitis E virus infection and its prevalence in pigs on comercial farms in Spain. Vet. J. 2008, 175, 130–132. [Google Scholar] [CrossRef]
- Denzin, N.; Borgwardt, J. Occurance and geographical distribution of antibodies to hepatitis E virus in wild boars of Saxony-Anhalt, Germany. Berl. Muench. Tieraerztl. Wschr. 2013, 126, 230–235. [Google Scholar] [CrossRef]
- Spancerniene, U.; Buitkuviene, J.; Grigas, J.; Pampariene, I.; Salomskas, A.; Cepuliene, R.; Žymantienė, J.; Stankevicius, A. Seroprevalence of hepatitis E virus in Lithuanian domestic pigs and wildlife. Acta Vet. Brno 2016, 85, 319–327. [Google Scholar] [CrossRef]
- Tsachev, I.; Baymakova, M.; Marutsov, P.; Gospodinova, K.; Kundurzhiev, T.; Petrov, V.; Pepovich, R. Seroprevalence of hepatitis E virus infection among wild boars in Western Bulgaria. Vector Borne Zoonotic Dis. 2021, 21, 441–445. [Google Scholar] [CrossRef]
- Larska, M.; Krzysiak, M.K.; Jabłonski, A.; Kezsik, J.; Bednarski, M.; Rola, J. Hepatitis E Virus antibody prevalence in wildlife in Poland. Zoonoses Public Health 2015, 62, 105–110. [Google Scholar] [CrossRef]
- Dorn-In, S.; Schwaiger, K.; Twarużek, M.; Grajewski, J.; Gottschalk, C.; Gareis, M. Hepatitis E Virus in wild boar in Northwest Poland: Sensitivity of methods of detection. Foodborne Pathog. Dis. 2017, 14, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Boadella, M.; Vicente, J.; Ruiz-Fons, F.; Martín, M.; Segales, J.; Gortázar, C. Seroprevalence evolution of selected pathogens in Iberian wild boar. Transbound. Emerg. Dis. 2021, 59, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, S.; Filipello, V.; Pavoni, E.; Carta, V.; Bolzoni, L.; Corradi, M.; Gilioli, S.; Losio, M.N. Geographical restriction of Hepatitis E virus circulation in wild boars (Sus scrofa) in Emilia-Romagna region, Northern Italy. Ital. J. Food Saf. 2020, 9, 8463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santis, P.; Di Pasquale, S.; La Rosa, G.; De Medici, D.; Micarelli, G.; Bilei, S.; Suffredini, E. High levels of Hepatitis E virus in wild boar hunted for domestic consumption in Central Italy. Eur. J. Public Health 2018, 28. [Google Scholar] [CrossRef]
- Rivero-Juarez, A.; Risalde, M.A.; Frias, M.; García-Bocanegra, I.; Lopez-Lopez, P.; Cano-Terriza, D.; Camacho, A.; Jimenez-Ruiz, S.; Gomez-Villamandos, J.C.; Rivero, A. Prevalence of hepatitis E virus infection in wild boars from Spain: A possible seasonal pattern? BMC Vet. Res. 2018, 14, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halbur, P.G. Attempt to Transmit Swine Hepatitis E Virus (HEV) by Consumption of Fresh and Frozen Pork Loin or Liver. Research Report Swine Health. 2005. Available online: https://porkcheckoff.org/wp-content/uploads/2021/02/04-087-HALBUR-ISU.pdf (accessed on 19 August 2021).
- Mazzei, M.; Nardini, R.; Verin, R.; Forzan, M.; Poli, A.; Tolari, F. Serologic and molecular survey for hepatitis E virus in wild boar (Sus scrofa) in Central Italy. New Microbes New Infect. 2015, 7, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Pisano, M.B.; Winter, M.; Raimondo, N.; Martínez-Wassaf, M.G.; Abate, S.D.; Réa, V.E. New pieces in the transmission cycle of the hepatitis E virus in South America: First viral detection in wild boars from Argentina. Trans. R Soc. Trop. Med. Hyg. 2019, 113, 497–499. [Google Scholar] [CrossRef] [Green Version]
- Bigoraj, E.; ·Paszkiewicz, W.; Rzeżutka, A. Porcine blood and liver as sporadic sources of Hepatitis E Virus (HEV) in the production chain of offal-derived foodstuffs in Poland. Food Environ. Virol. 2021, 13, 347–356. [Google Scholar] [CrossRef]
- Aprea, G.; Scattolini, S.; D’Angelantonio, D.; Chiaverini, A.; Di Lollo, V.; Olivieri, S.; Marcacci, M.; Mangone, I.; Salucci, S.; Antoci, S.; et al. Whole genome sequencing characterization of HEV3-e and HEV3-f subtypes among the wild boar population in the Abruzzo region, Italy: First report. Microorganisms 2020, 8, 1393. [Google Scholar] [CrossRef]


{kind=link}
| Year | HEV ELISA | HEV RT-qPCR | |||||
|---|---|---|---|---|---|---|---|
| Negative | Questionable | Positive | Total | Negative | Positive | Total | |
| 2016 | 135 | 0 | 22 | 157 | 20 | 2 | 22 |
| (86%) | (0%) | (14%) | (6.8%) | (90.9%) | (9.1%) | (8.8%) | |
| 2017 | 476 | 1 | 57 | 534 | 50 | 7 | 57 |
| (89.1%) | (0.2%) | (10.7%) | (23.3%) | (87.7%) | (12.3%) | (22.9%) | |
| 2018 | 630 | 5 | 100 | 735 | 81 | 19 | 100 |
| (85.7%) | (0.7%) | (13.6%) | (32%) | (81%) | (19%) | (40.2%) | |
| 2019 | 577 | 0 | 52 | 629 | 45 | 7 | 52 |
| (91.7%) | (0%) | (8.3%) | (27.4%) | (86.5%) | (13.5%) | (20.9%) | |
| 2020 | 219 | 3 | 18 | 240 | 14 | 4 | 18 |
| (91.3%) | (1.2%) | (7.5%) | (10.5%) | (77.8%) | (22.2%) | (7.2%) | |
| Total | 2037 | 9 | 249 | 2295 | 210 | 39 | 249 |
| (88.8%) | (0.4%) | (10.8%) | (100%) | (84.3%) | (15.7%) | (100%) | |
| HEV RT-qPCR | ||||
|---|---|---|---|---|
| Positive 1 | Negative | Total | ||
| HEV ELISA | Positive | 39 (15.7%) | 210 (84.3%) | 249 (39.2%) |
| Negative | 5 (1.3%) | 381 (98.7%) | 386 (60.8%) | |
| Total | 44 (6.9%) | 591 (93.1%) | 635 (100%) | |
| Age (Years) | HEV RT-qPCR | ||
|---|---|---|---|
| Positive | Negative | Total | |
| <1 | 26 (24.5%) | 80 (75.5%) | 106 (44.9%) |
| >1 | 11 (8.5%) | 119 (91.5%) | 130 (55.1%) |
| Total | 37 (15.7%) | 199 (84.3%) | 236 (100%) 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polley, B.; Contzen, M.; Wiedmann, S.; Schneider-Bühl, L.; Sting, R. Detection of Hepatitis E Virus Infections in Wild Boars in Southwest Germany Using a Stepwise Laboratory Diagnostic Approach. Zoonotic Dis. 2022, 2, 9-18. https://doi.org/10.3390/zoonoticdis2010002
Polley B, Contzen M, Wiedmann S, Schneider-Bühl L, Sting R. Detection of Hepatitis E Virus Infections in Wild Boars in Southwest Germany Using a Stepwise Laboratory Diagnostic Approach. Zoonotic Diseases. 2022; 2(1):9-18. https://doi.org/10.3390/zoonoticdis2010002
Chicago/Turabian StylePolley, Birgitta, Matthias Contzen, Sandra Wiedmann, Lisa Schneider-Bühl, and Reinhard Sting. 2022. "Detection of Hepatitis E Virus Infections in Wild Boars in Southwest Germany Using a Stepwise Laboratory Diagnostic Approach" Zoonotic Diseases 2, no. 1: 9-18. https://doi.org/10.3390/zoonoticdis2010002