

Seed Morphology of Three Neotropical Orchid Species of the Lycaste Genus

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Seed Material
2.2. Seed Capsule Assessment
2.3. Seed and Embryo Morphology
2.4. Data Analysis
3. Results
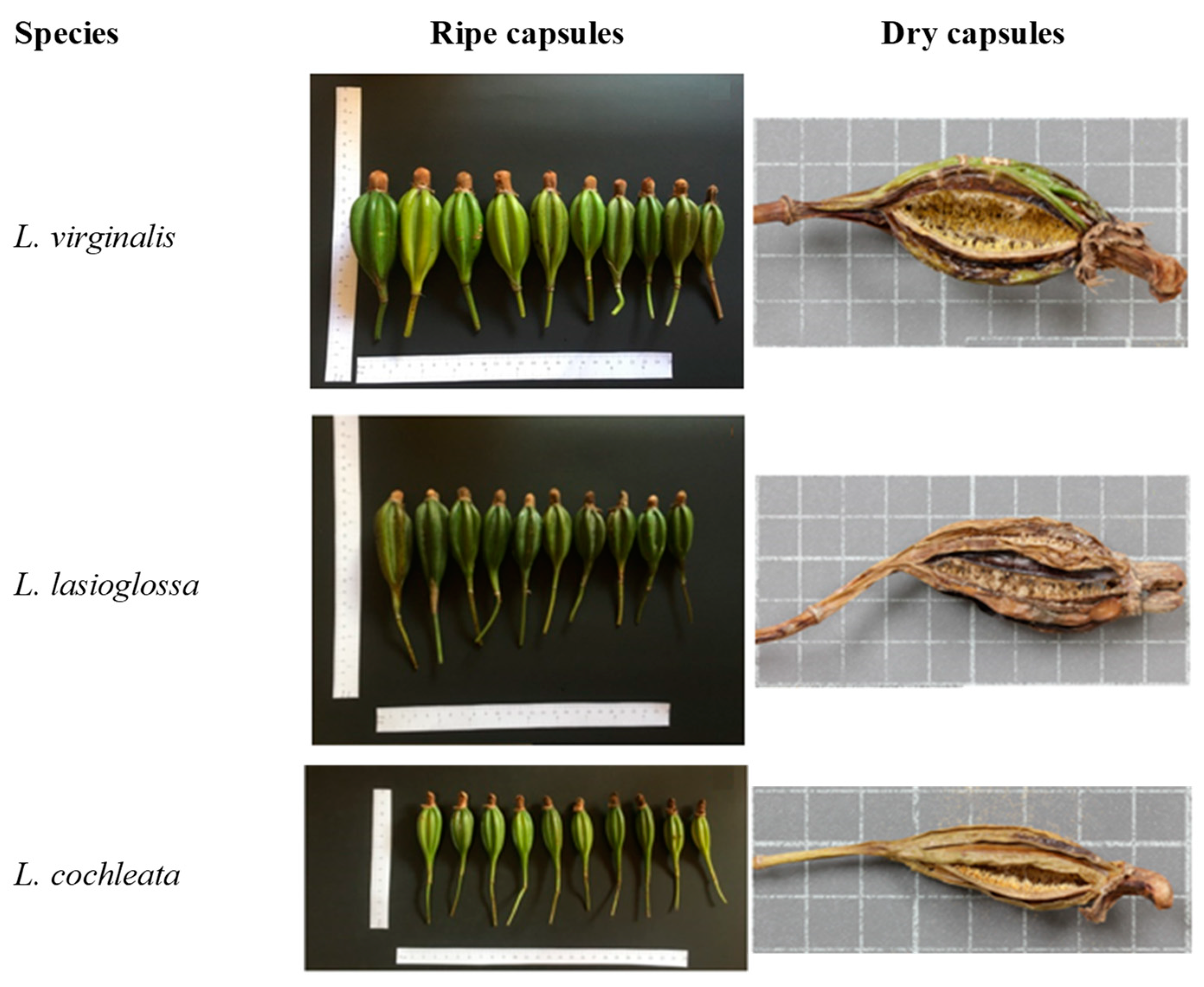
3.1. Seed Capsule Morphology
3.2. Seed Morphology
4. Discussion
4.1. Seed Capsule Morphology
4.2. Seed Morphology
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Didier, L.; Tamay, C. Orquídeas: Importancia y uso en México. Bioagrociencias 2013, 6, 7. [Google Scholar]
- Meisel, J.E.; Kaufmann, R.S.; Pupulin, F. Orchids of Tropical America: An Introduction and Guide; Cornell University Press: Ithaca, NY, USA; Comstock Publishing Associates: New York, NY, USA, 2014. [Google Scholar]
- Cavero, M.; Collantes, B.; Patroni, C. Orquídeas del Perú; Centro de datos para la Conservación del Perú: Lima, Perú, 1991. [Google Scholar]
- Lawson, C.; Wraith, J.; Pickering, C. Regulating wild collected orchids? The CBD, Nagoya Protocol and CITES overlaps. Environ. Plan. Law J. 2019, 36, 339–361. [Google Scholar]
- Martija-Ochoa, M. El Gran Libro de las Orquídeas; Parkstone International: New York, NY, USA, 2019. [Google Scholar]
- Sailo, N.; Rai, D.; De, L. Physiology of temperate and tropical orchids-an overview. Int. J. Sci. 2014, 3, 3–8. [Google Scholar]
- Archila Morales, F. Listado actualizado de orquídeas de Guatemala. Guatemalensis 2022, 25, 129–419. [Google Scholar]
- Gaskett, A.C.; Gallagher, R.V. Orchid diversity: Spatial and climatic patterns from herbarium records. Ecol. Evoution 2018, 22, 11235–11245. [Google Scholar] [CrossRef] [PubMed]
- Archila Morales, F.; Chiron, G.R. El género Lycaste en Lindley en Guatemala. Guatemalensis 2011, 14, 1–55. [Google Scholar]
- World Flora Online, Lycaste Lindl. Available online: http://www.worldfloraonline.org/taxon/wfo-4000022472;jsessionid=994875A46326DB4656AA9765C0EF5F5C (accessed on 16 July 2021).
- KEW, Plants of the World Online: Lycaste Lindl. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:30000072-2 (accessed on 29 June 2023).
- Ames, O.; Correll, D.S. Orchids of Guatemala; Chicago Natural History Museum: Chicago, IL, USA, 1953; Volume 26, Number 2. [Google Scholar]
- CONAP. Lista de Especies Amenazadas de Guatemala (LEA); Departamento de Vida Silvestre del Consejo Nacional de Áreas Protegidas de Guatemala: Guatemala City, Guatemala, 2009. [Google Scholar]
- CITES Species Lists. 2021. Available online: https://www.speciesplus.net/species (accessed on 14 July 2021).
- Diantina, S.; Clavijo McCormick, A.; Pritchard, H.W.; Millner, J.; Nadarajan, J.; Mastur, M.; McGill, C. Orchid seed micro-morphometry: Importance to species biology, ecology, and conservation. II Int. Symp. Trop. Subtrop. Ornam. 2021, 1334, 153–162. [Google Scholar] [CrossRef]
- Verma, J.; Sharma, K.; Thakur, K.; Sembi, J.K.; Vij, S.P. Study on seed morphometry of some threatened Western Himalayan orchids. Turk. J. Bot. 2014, 38, 234–251. [Google Scholar] [CrossRef]
- Barthlott, W.; Große-Veldmann, B.; Korotkova, N. Orchid seed diversity. Englera 2014, 32, 1–245. [Google Scholar]
- Tsutsumi, C.; Yukawa, T.; Lee, N.S.; Lee, C.S.; Kato, M. Phylogeny and comparative seed morphology of epiphytic and terrestrial species of Liparis (Orchidaceae) in Japan. J. Plant Res. 2007, 120, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Leck, M.A.; Parker, V.T.; Simpson, R. Seedling Ecology and Evolution; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Weather Atlas, Monthly Weather Forecast and Climate Cobán, Guatemala. 2021. Available online: https://www.weather-atlas.com/en/guatemala/coban-climate (accessed on 28 December 2021).
- Pantone, I. PANTONE® Colours. 2017. Available online: http://www.pantone-colours.com/ (accessed on 28 December 2021).
- ISTA. International Rules for Seed Testing; The International Seed Testing Association: Bassersdorf, Switzerland, 2022. [Google Scholar]
- Diantina, S.; McGill, C.; Millner, J.; Nadarajan, J.; WPritchard, H.; Clavijo McCormick, A. Comparative seed morphology of tropical and temperate orchid species with different growth habits. Plants 2020, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Arditti, J.; Ghani, A. Numerical and physical properties of orchid seeds and their biological implications. New Phytologist. 2000, 146, 569. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows; IBM Corp: Armonk, NY, USA, 2022. [Google Scholar]
- Rasmussen, F.N.; Johansen, B. Carpology of orchids. Selbyana 2006, 44–53. [Google Scholar]
- Ames, O.; Correll, D.S. Orchids of Guatemala; Chicago Natural History Museum: Chicago, IL, USA, 1952; Volume 26, Number 1. [Google Scholar]
- Vafaee, Y.; Mohammadi, G.; Nazari, F.; Fatahi, M.; Kaki, A.; Gholami, S.; Ghorbani, A.; Khadivi, A. Phenotypic characterization and seed-micromorphology diversity of the threatened terrestrial orchids: Implications for conservation. S. Afr. J. Bot. 2021, 137, 386–398. [Google Scholar] [CrossRef]
- Hariyanto, S.; Pratiwi, I.A.; Utami, E.S.W. Seed morphometry of native indonesian orchids in the genus Dendrobium. Scientifica 2020, 2020, 3986369. [Google Scholar] [CrossRef]
- Prasongsom, S.; Thammasiri, K.; Pritchard, H.W. Seed micromorphology and ex vitro germination of Dendrobium orchids. I Int. Symp. Trop. Subtrop. Ornam. 2016, 1167, 339–344. [Google Scholar] [CrossRef]
- Ortúñez, E.; Gamarra, R. Seed morphology, life form and distribution in three Bromheadia species (Epidendroideae, Orchidaceae). Diversity 2023, 15, 195. [Google Scholar] [CrossRef]
- Diantina, S.; Kartikaningrum, S.; McCormick, A.C.; Millner, J.; McGill, C.; Pritchard, H.W.; Nadarajan, J. Comparative in vitro seed germination and seedling development in tropical and temperate epiphytic and temperate terrestrial orchids. Plant Cell Tissue Organ Cult. (PCTOC) 2020, 143, 619–633. [Google Scholar] [CrossRef]
- Diantina, S.; McGill, C.; Millner, J.; Nadarajan, J.; Pritchard, H.W.; Colville, L.; Clavijo McCormick, A. Seed viability and fatty acid profiles of five orchid species before and after ageing. Plant Biol. 2022, 24, 168–175. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Capsule Measurement (cm) | |||
|---|---|---|---|---|
| Length | Ø Top | Ø Central | Ø Bottom | |
| L. cochleata | 7.90 ± 0.74 b | 1.0 ± 0.00 b | 2.1 ± 2.13 n.s. | 1.0 ± 0.00 c |
| L. lasioglossa | 9.80 ± 1.03 a | 2.1 ± 0.32 a | 2.9 ± 0.32 n.s. | 1.6 ± 0.52 b |
| L. virginalis | 10.3 ± 1.42 a | 2.2 ± 0.42 a | 3.0 ± 0.47 n.s. | 2.0 ± 0.00 a |
| Seed Dimensions | ||||
|---|---|---|---|---|
| Species | Length (mm) | Width (mm) | Volume (mm3) | |
| L. cochleata | 0.21 ± 0.02 c | 0.11 ± 0.01 a | 0.00063 ± 0.0002 a | |
| L. lasioglossa | 0.23 ± 0.03 b | 0.09 ± 0.01 b | 0.00051 ± 0.0001 b | |
| L. virginalis | 0.26 ± 0.02 a | 0.09 ± 0.01 b | 0.00060 ± 0.0002 a | |
| Embryo Dimensions | Air-space % in the Seed | |||
| Species | Length (mm) | Width (mm) | Volume (mm3) | |
| L. cochleata | 0.14 ± 0.01 a | 0.082 ± 0.01 a | 0.00051 ± 0.00011 a | 17.4 ± 8.0 c |
| L. lasioglossa | 0.12 ± 0.01 b | 0.066 ± 0.01 c | 0.00029 ± 0.0007 c | 42.3 ± 8.1 a |
| L. virginalis | 0.15 ± 0.01 a | 0.073 ± 0.01 b | 0.00041 ± 0.00009 b | 29.7 ± 9.5 b |
| Species | SL/SW | EL/EW | SV/EV |
|---|---|---|---|
| L. cochleata | 1.98 ± 0.26 c | 1.78 ± 0.24 b | 1.22 ± 0.13 c |
| L. lasioglossa | 0.57 ± 0.37 b | 1.86 ± 0.19 b | 1.77 ± 0.25 a |
| L. virginalis | 2.75 ± 0.38 a | 2.04 ± 0.26 a | 1.45 ± 0.21 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfaro Pinto, A.; McGill, C.; Nadarajan, J.; Archila Morales, F.; Clavijo McCormick, A. Seed Morphology of Three Neotropical Orchid Species of the Lycaste Genus. Seeds 2023, 2, 331-339. https://doi.org/10.3390/seeds2030025
Alfaro Pinto A, McGill C, Nadarajan J, Archila Morales F, Clavijo McCormick A. Seed Morphology of Three Neotropical Orchid Species of the Lycaste Genus. Seeds. 2023; 2(3):331-339. https://doi.org/10.3390/seeds2030025
Chicago/Turabian StyleAlfaro Pinto, Alejandra, Craig McGill, Jayanthi Nadarajan, Fredy Archila Morales, and Andrea Clavijo McCormick. 2023. "Seed Morphology of Three Neotropical Orchid Species of the Lycaste Genus" Seeds 2, no. 3: 331-339. https://doi.org/10.3390/seeds2030025
APA StyleAlfaro Pinto, A., McGill, C., Nadarajan, J., Archila Morales, F., & Clavijo McCormick, A. (2023). Seed Morphology of Three Neotropical Orchid Species of the Lycaste Genus. Seeds, 2(3), 331-339. https://doi.org/10.3390/seeds2030025