
Environmental Regulation of Weed Seed Dormancy and Germination †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Seed Dormancy: A Survival Strategy
3. Seed Germination: The Initiation of a New Generation
4. The Seed Bank: A Rewarding Investment
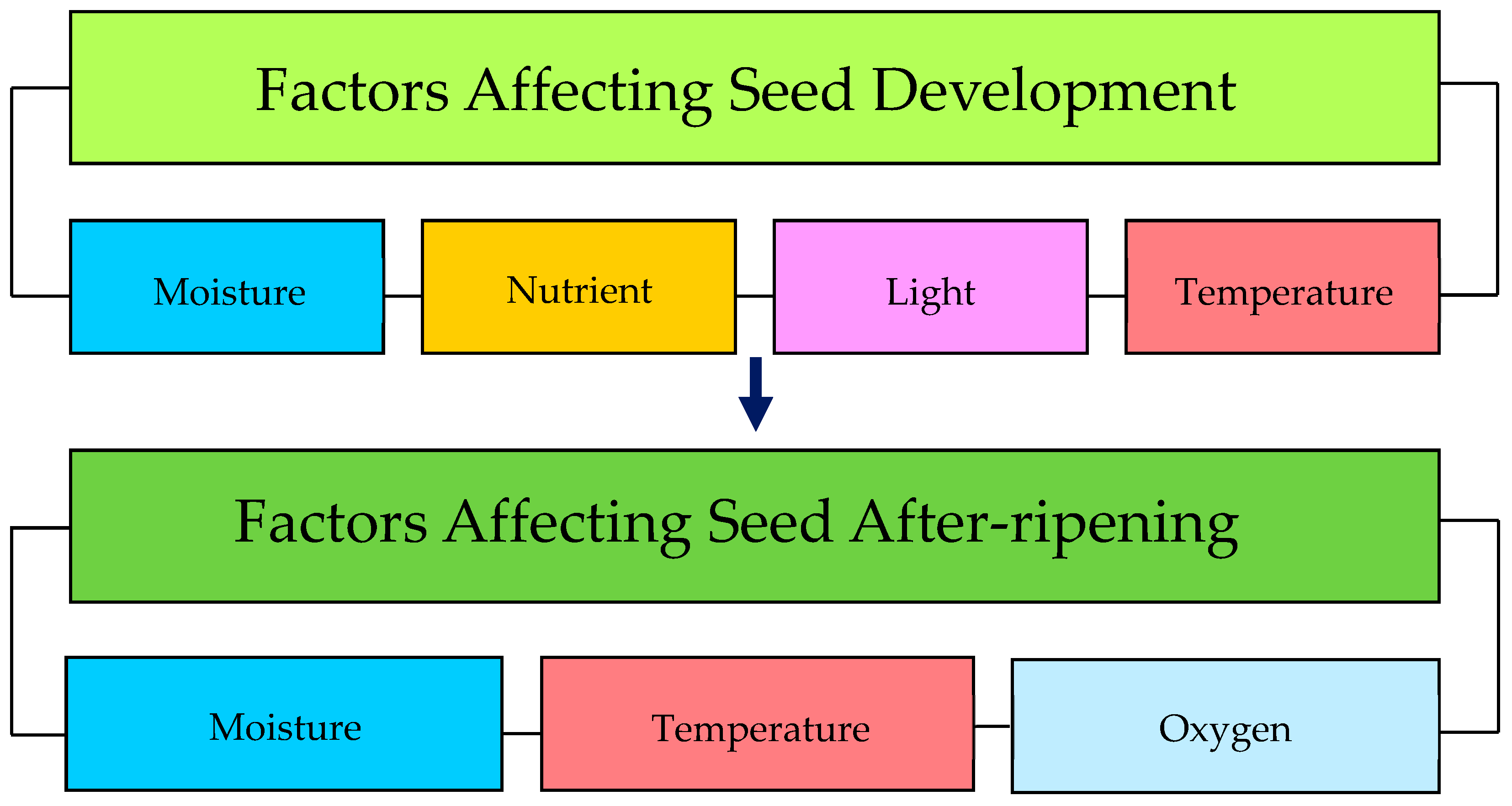
5. The Effects of Pre-Dispersal Factors on Seed Dormancy: Maturation Conditions
6. The Effects of Post-Dispersal Factors on Seed Dormancy: After-Ripening Conditions
7. Scotch Thistle: A Model Plant Species to Study Seed Dormancy
8. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gutterman, Y. Environmental factors and survival strategies of annual plant species in the Negev Desert, Israel. Plant Species Biol. 2000, 15, 113–125. [Google Scholar] [CrossRef]
- Gutterman, Y. Maternal effects on seeds during development. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 59–84. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Pre- and post-dispersal factors regulate germination patterns and structural characteristics of Scotch thistle (Onopordum acanthium) cypselas. New Phytol. 2003, 159, 263–278. [Google Scholar] [CrossRef] [PubMed]
- LeFait, B.M.; Qaderi, M.M. Maternal environmental effects of temperature and exogenous gibberellic acid on seed and seedling traits of four populations of evening primrose (Oenothera biennis). Seeds 2022, 1, 110–125. [Google Scholar] [CrossRef]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Reid, D.M. Combined effects of temperature and carbon dioxide on plant growth and subsequent seed germinability of Silene noctiflora. Int. J. Plant Sci. 2008, 169, 1200–1209. [Google Scholar] [CrossRef]
- Née, G.; Xiang, Y.; Soppe, W.J.J. The release of dormancy, a wake-up call for seeds to germinate. Curr. Opin. Plant Biol. 2017, 35, 8–14. [Google Scholar] [CrossRef]
- Gray, D.; Steckel, J.R.A.; Dearman, J.; Brocklehurst, P.A. Some effects of temperature during seed development on carrot (Daucus carota) seed growth and quality. Ann. Appl. Biol. 1988, 112, 367–376. [Google Scholar] [CrossRef]
- Wulff, R.D. Environmental maternal effects on seed quality and germination. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 491–505. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Klupczyńska, E.A.; Pawłowski, T.A. Regulation of seed dormancy and germination mechanisms in changing environment. Int. J. Mol. Sci. 2021, 22, 1357. [Google Scholar] [CrossRef]
- Nautiyal, P.C.; Sivasubramaniam, K.; Dadlani, M. Seed dormancy and regulation of germination. In Seed Science and Technology: Biology, Production, Quality; Dadlani, M., Yadava, D.K., Eds.; Springer: Gateway East, Singapore, 2023; pp. 39–66. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Isolation and structural characterization of a water-soluble germination inhibitor from Scotch thistle (Onopordum acanthium) cypselas. J. Chem. Ecol. 2003, 29, 2425–2438. [Google Scholar] [CrossRef]
- Cavers, P.B.; Qaderi, M.M.; Manku, R.; Downs, M.P. Intermittent germination: Causes and ecological implications. In Seed Biology: Advances and Applications; Black, M., Bradford, K.J., Vázquez-Ramos, J., Eds.; CAB International: Wallingford, UK, 2000; pp. 363–374. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Smith, R.G.; Birthisel, S.K.; Bosworth, S.C.; Brown, B.; Davis, T.M.; Gallandt, E.R.; Hazelrigg, A.; Venturini, E.; Warren, N.D. Environmental correlates with germinable weed seedbanks on organic farms across northern New England. Weed Sci. 2018, 66, 78–93. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Physiology and Biochemistry of Seeds. 2. Viability, Dormancy, and Environmental Control; Springer: Berlin/Heidelberg, Germany, 1982. [Google Scholar]
- Egley, G.H.; Duke, S.O. Physiology of weed seed dormancy and germination. In Weed Physiology. Vol. I. Reproduction and Ecophysiology; Duke, S.O., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 27–64. [Google Scholar]
- Simpson, G.M. Seed Dormancy in Grasses; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Qaderi, M.M.; Presti, A.; Cavers, P.B. Dry storage effects on germinability of Scotch thistle (Onopordum acanthium) cypselas. Acta Oecol. 2005, 27, 67–74. [Google Scholar] [CrossRef]
- Zhang, R.; Baskin, J.M.; Baskin, C.C.; Mo, Q.; Chen, L.; Hu, X.; Wang, Y. Effect of population, collection year, after-ripening and incubation condition on seed germination of Stipa bungeana. Sci. Rep. 2017, 7, 13893. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Bentsink, L.; Koornneef, M. Seed dormancy and germination. Arab. Book 2008, 6, e0119. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; de Casas, R.R.; NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Sun, L.; Baskin, C.C.; Baskin, J.M.; Cao, M.; Yang, J. Seed dormancy in space and time: Global distribution, paleoclimatic and present climatic drivers, and evolutionary adaptations. New Phytol. 2022, 234, 1770–1781. [Google Scholar] [CrossRef]
- Salisbury, E.J. Weeds and Aliens; Collins: London, UK, 1961. [Google Scholar]
- Nonogaki, H. Seed dormancy and germination—emerging mechanisms and new hypotheses. Front. Plant Sci. 2014, 5, 233. [Google Scholar] [CrossRef]
- Foley, M.E. Weeds, seeds, and buds–opportunities and systems for dormancy investigations. Weed Sci. 2002, 50, 267–272. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Bradford, K.J. Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Graeber, K.; Nakabayashi, K.; Miatton, E.; Leubner-Metzger, G.; Soppe, W.J.J. Molecular mechanisms of seed dormancy. Plant Cell Environ. 2012, 35, 1769–1786. [Google Scholar] [CrossRef]
- Werker, E. Seed Anatomy; Gebrüder Borntraeger: Berlin/Stuttgart, Germany, 1997. [Google Scholar]
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977. [Google Scholar]
- Nikolaeva, M.G. Factors affecting the seed dormancy pattern. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 51–74. [Google Scholar]
- Hilhorst, H.W.M. A critical update on seed dormancy. I. Primary dormancy. Seed Sci. Res. 1995, 5, 61–73. [Google Scholar] [CrossRef]
- Vleeshouwers, L.M.; Boumeester, H.J.; Karssen, C.M. Redefining seed dormancy: An attempt to integrate physiology and ecology. J. Ecol. 1995, 83, 1031–1037. [Google Scholar] [CrossRef]
- Harper, J.L. The ecological significance of dormancy and its importance in weed control. Proc. IVth Int. Cong. Crop Prot. 1957, 4, 415–420. [Google Scholar]
- Rees, M. Seed dormancy. In Plant Ecology, 2nd ed.; Crawley, M.J., Ed.; Blackwell Science Ltd.: Oxford, UK, 1997; pp. 214–238. [Google Scholar]
- Doucet, C.; Cavers, P.B. Induced dormancy and colour polymorphism in seeds of the bull thistle Cirsium vulgare (Savi) Ten. Seed Sci. Res. 1997, 7, 399–407. [Google Scholar] [CrossRef]
- Bradbeer, J.W. Seed Dormancy and Germination; Chapman & Hall: New York, NY, USA, 1988. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. The great diversity in kinds of seed dormancy: A revision of the Nikolaeva-Baskin classification system for primary seed dormancy. Seed Sci. Res. 2021, 31, 249–277. [Google Scholar] [CrossRef]
- Kitajima, K.; Fenner, M. Ecology of seedling regeneration. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 331–359. [Google Scholar]
- de Casas, R.R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global biogeography of seed dormancy is determined by seasonal and seed size: A case study in the legumes. New Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Cavers, P.B.; Hamill, A.S.; Downs, M.P.; Bernards, M.A. Maturation temperature regulates germinability and chemical constituents of Scotch thistle (Onopordum acanthium) cypselas. Can. J. Bot. 2006, 84, 28–38. [Google Scholar] [CrossRef]
- Gardarin, A.; Dürr, C.; Colbach, N. Prediction of germination rates of weed species: Relationships between germination speed parameters and species traits. Ecol. Modell. 2011, 222, 626–636. [Google Scholar] [CrossRef]
- Casal, J.J.; Sánchez, R.A. Phytochromes and seed germination. Seed Sci. Res. 1998, 8, 317–329. [Google Scholar] [CrossRef]
- Malavert, C.; Batlla, D.; Benech-Arnold, R.L. Light sensitivity changes during dormancy induction in Polygonum aviculare L. seeds: Development of a predictive model of annual changes in seed-bank light sensitivity in relation to soil temperature. Weed Res. 2021, 61, 115–125. [Google Scholar] [CrossRef]
- Bączek-Kwinta, R. An interplay of light and smoke compounds in photoblastic seeds. Plants 2022, 11, 1773. [Google Scholar] [CrossRef] [PubMed]
- Pons, T.L. Seed responses to light. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI International: Wallingford, UK, 2000; pp. 237–260. [Google Scholar]
- Hilhorst, H.W.M.; Toorop, P.E. Review on dormancy, germinability and germination in crop and weed seeds. Adv. Agron. 1997, 61, 111–165. [Google Scholar]
- Taylorson, R.B. Environmental and chemical manipulation of weed seed dormancy. Rev. Weed Sci. 1987, 3, 135–154. [Google Scholar]
- Werker, E. Seed dormancy as explained by the anatomy of embryo envelopes. Isr. J. Bot. 1980, 29, 22–44. [Google Scholar]
- Kelly, K.M.; Van Staden, J.; Bell, W.E. Seed coat structure and dormancy. Plant Growth Regul. 1992, 11, 201–209. [Google Scholar] [CrossRef]
- Pereira, M.P.; Corrêa, F.F.; Polo, M.; de Castro, E.M.; Cardoso, A.A.; Pereira, F.J. Seed germination of Schinus molle L. (Anacardiaceae) as related to its anatomy and dormancy alleviation. Seed Sci. Res. 2016, 26, 351–361. [Google Scholar] [CrossRef]
- Karssen, C.M. Seasonal patterns of dormancy in weed seeds. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 243–270. [Google Scholar]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental control of dormancy in weed seed banks in soil. Field Crops Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Just, M.; Cross, A.T.; Lewandrowski, W.; Turner, S.R.; Merritt, D.J.; Dixon, K. Seed dormancy alleviation by warm stratification progressively widens the germination window in Mediterranean climate Rutaceae. Aust. J. Bot. 2023, 71, 55–66. [Google Scholar] [CrossRef]
- Pearson, C.F.; Ison, R.L. Agronomy of Grassland Systems, 2nd ed.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Taiz, L.; Møller, I.M.; Murphy, A.; Zeiger, E. Plant Physiology and Development, 7th ed.; Sinauer: Sunderland, MA, USA, 2023. [Google Scholar]
- Hilhorst, H.W.M.; Karssen, C.M. Seed dormancy and germination: The role of abscisic acid and gibberellins and the importance of hormone mutants. Plant Growth Regul. 1992, 11, 225–238. [Google Scholar] [CrossRef]
- Footitt, S.; Walley, P.G.; Lynn, J.R.; Hambidge, A.J.; Penfield, S.; Finch-Savage, W.E. Trait analysis reveals DOG1 determines initial depth of seed dormancy, but not changes during dormancy cycling that result in seedling emergence timing. New Phytol. 2020, 225, 2035–2047. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.-Q.; Luan, S.; Li, J.; He, Z.-H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef]
- Shu, K.; Liu, X.-D.; Xie, Q.; He, Z.-H. Two faces of one seed: Hormonal regulation of dormancy and germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- MacGregor, D.R.; Zhang, N.; Iwasaki, M.; Chen, M.; Dave, A.; Lopez-Molina, L.; Penfield, S. ICE1 and ZOU determine the depth of primary seed dormancy in Arabidopsis independently of their role in endosperm development. Plant J. 2019, 98, 277–290. [Google Scholar] [CrossRef]
- Radosevich, S.; Holt, J.; Ghersa, C. Weed Ecology: Implications for Management, 2nd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Balyan, R.S.; Bhan, V.M. Germination of horse purslane (Trianthema portulacastrum) in relation to temperature, storage conditions, and seedling depths. Weed Sci. 1986, 34, 513–515. [Google Scholar] [CrossRef]
- Thompson, K.; Grime, J.P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. J. Ecol. 1979, 67, 893–921. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Leubner-Metzger, G. Seed dormancy and weed emergence: From simulating environmental change to understanding trait plasticity, adaptive evolution, and population fitness. J. Exp. Bot. 2021, 72, 4181–4185. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Footitt, S. Seed dormancy cycling and the regulation of dormancy mechanisms to time germination in variable field environments. J. Exp. Bot. 2017, 68, 843–856. [Google Scholar] [CrossRef]
- Wang, G.; Baskin, C.C.; Baskin, J.M.; Yang, X.; Liu, G.; Zhang, X.; Ye, X.; Huang, Z. Timing of seed germination in two alpine herbs on the southeastern Tibetan plateau: The role of seed dormancy and annual dormancy cycling in soil. Plant Soil 2017, 421, 465–476. [Google Scholar] [CrossRef]
- Zhang, K.; Ji, Y.; Fu, G.; Yao, L.; Liu, H.; Tao, J.; Walck, J.L. Dormancy cycles in Aquilegia oxysepala Trautv. et Mey. (Ranunculaceae), a species with non-deep simple morphophysiological dormancy. Plant Soil 2021, 464, 223–235. [Google Scholar] [CrossRef]
- Soltani, E.; Baskin, C.C.; Baskin, J.M.; Soltani, A.; Galeshi, S.; Ghaderi-far, F.; Zeinali, E. A quantitative analysis of seed dormancy and germination in the winter annual weed Sinapis arvensis (Brassicaceae). Botany 2016, 94, 289–300. [Google Scholar] [CrossRef]
- Garcia, Q.S.; Barreto, L.C.; Bicalho, E.M. Environmental factors driving seed dormancy and germination in tropical ecosystems: A perspective from campo rupestre species. Environ. Exp. Bot. 2020, 178, 104164. [Google Scholar] [CrossRef]
- Mašková, T.; Poschlod, P. Soil seed bank persistence across time and burial depth in calcareous grassland habitats. Front. Plant Sci. 2022, 12, 790867. [Google Scholar] [CrossRef]
- Telewski, F.W.; Zeevaart, J.A.D. The 120-yr period for Dr. Beal’s seed viability experiment. Am. J. Bot. 2002, 89, 1285–1288. [Google Scholar] [CrossRef] [PubMed]
- Fenner, M. Ecology of seed banks. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 507–528. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B. Interpopulation and interyear variation in germination in Scotch thistle, Onopordum acanthium L., grown in a common garden: Genetics vs environment. Plant Ecol. 2002, 162, 1–8. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Seed bank dynamics of Onopordum acanthium: Emergence patterns and chemical attributes. J. Ecol. 2002, 90, 672–683. [Google Scholar] [CrossRef]
- Loades, E.; Pérez, M.; Turečková, V.; Tarkowská, D.; Strnad, M.; Seville, A.; Nakabayashi, K.; Leubner- Metzger, G. Distinct hormonal and morphological control of dormancy and germination in Chenopodium album dimorphic seeds. Front. Plant Sci. 2023, 14, 1156794. [Google Scholar] [CrossRef]
- Evert, R.F.; Eichhorn, S.E. Raven Biology of Plants, 8th ed.; W.H. Freeman & Company: New York, NY, USA, 2013. [Google Scholar]
- Boesewinkel, F.D.; Bouman, F. The seed: Structure and function. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 1–24. [Google Scholar]
- Yadegari, R.; Goldberg, R.B. Embryogenesis in dicotyledonous plants. In Cellular and Molecular Biology of Plant Seed Development; Larkins, B.A., Vasil, I.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 3–52. [Google Scholar]
- Perez-Grau, L. Plant embryogenesis–the cellular design of a plant. In Plant Reproduction; O’Neill, S.D., Roberts, J.A., Eds.; Sheffield Academic Press: Sheffield, UK, 2002; pp. 154–192. [Google Scholar]
- Bryant, J.A. Seed Physiology; Edward Arnold (Publishers) Ltd.: London, UK, 1985. [Google Scholar]
- Demason, D.A. Endosperm structure and development. In Cellular and Molecular Biology of Plant Seed Development; Larkins, B.A., Vasil, I.K., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 73–115. [Google Scholar]
- Brown, R.C.; Lemmon, B.E.; Nguyen, H. Endosperm development. In Plant Reproduction; O’Neill, S.D., Roberts, J.A., Eds.; Sheffield Academic Press: Sheffield, UK, 2002; pp. 193–220. [Google Scholar]
- Penfield, S. Seed dormancy and germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef]
- Tran, V.N.; Cavanagh, A.K. Structural aspects of dormancy. In Seed Physiology. 2. Germination and Reserve Mobilization; Murray, D.R., Ed.; Academic Press Australia: North Ryde, NSW, Australia, 1984; pp. 1–44. [Google Scholar]
- Zhang, Y.; Zhang, K.; Ji, Y.; Tao, J. Physical dormancy and soil seed bank dynamics in seeds of Melilotus albus (Fabaceae). Flora 2020, 266, 151600. [Google Scholar] [CrossRef]
- Roach, D.A.; Wulff, R.D. Maternal effects in plants. Ann. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Fenner, M. Environmental influences on seed size and composition. Hortic. Rev. 1992, 13, 183–213. [Google Scholar]
- Murdoch, A.J.; Ellis, R.H. Dormancy, viability and longevity. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CAB International: Wallingford, UK, 2000; pp. 183–214. [Google Scholar]
- Huang, Z.; Footitt, S.; Tang, A.; Finch-Savage, W.E. Predicted global warming scenarios impact on the mother plant to alter seed dormancy and germination behaviour in Arabidopsis. Plant Cell Environ. 2018, 41, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Fitter, A.H.; Hay, R.K.M. Environmental Physiology of Plants, 3rd ed.; Academic Press: New York, NY, USA, 2002. [Google Scholar]
- Lyndon, R.F. The environmental control of reproductive development. In Fruit and Seed Production: Aspects of Development, Environmental Physiology and Ecology; Marshall, C., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 9–32. [Google Scholar]
- Duffus, C.M. Control of grain growth and development. In Fruit and Seed Production: Aspects of Development, Environmental Physiology and Ecology; Marshall, C., Grace, J., Eds.; Cambridge University Press: Cambridge, UK, 1992; pp. 125–149. [Google Scholar]
- Boyce, K.G. Effect of temperature and dormancy on germination of tall fescue. Crop Sci. 1976, 16, 15–18. [Google Scholar] [CrossRef]
- Van der Vegte, F.W. Population differentiation and germination ecology in Stellaria media (L.) Vill. Oecologia 1978, 37, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Sawhney, R.; Naylor, J.M. Dormancy studies in seed of Avena fatua. 12. Influence of temperature on germination behavior of nondormant families. Can. J. Bot. 1980, 58, 578–581. [Google Scholar] [CrossRef]
- Somody, C.N.; Nalewaja, J.D.; Miller, S.E. The response of wild oat (Avena fatua) and Avena sterilis accessions to photoperiod and temperature. Weed Sci. 1984, 32, 206–213. [Google Scholar] [CrossRef]
- Probert, R.J.; Smith, R.D.; Birch, P. Germination responses to light and alternating temperatures in European populations of Dactylis glomerata L. II. The genetic and environmental components of germination. New Phytol. 1985, 99, 317–322. [Google Scholar] [CrossRef]
- Karssen, C.M. The light promoted germination of the seeds of Chenopodium album L. III. Effect of the photoperiod during growth and development of the plants on the dormancy of the produced seeds. Acta Bot. Neerl. 1970, 19, 81–94. [Google Scholar] [CrossRef]
- McCullough, J.M.; Shropshire, W. Physiological predetermination of responses in Arabidopsis thaliana (L.) Heynh. Plant Cell Physiol. 1970, 11, 139–148. [Google Scholar] [CrossRef]
- Cresswell, E.G.; Grime, J.P. Induction of a light requirement during seed development and its ecological consequences. Nature 1981, 291, 583–585. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Fenner, M.; Edwards, P.J. Changes in dormancy level in Sorghum halepense seeds induced by water stress during seed development. Funct. Ecol. 1992, 6, 596–605. [Google Scholar] [CrossRef]
- Abo Gamar, M.I.; Qaderi, M.M. Interactive effects of temperature, carbon dioxide and watering regime on seed germinability oftwo genotypes of Arabidopsis thaliana. Seed Sci. Res. 2019, 29, 12–20. [Google Scholar] [CrossRef]
- Hill, H.J.; West, S.H.; Hinson, K. Effect of water stress during seed fill on impermeable seed expression in soybean. Crop Sci. 1986, 26, 807–812. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Year-to-year variation in the germination of freshly-harvested seeds of Arenaria patula var. robusta from the same site. J. Tenn. Acad. Sci. 1975, 50, 106–108. [Google Scholar]
- Fawcett, R.S.; Slife, F.W. Effects of field applications of nitrate on weed seed germination and dormancy. Weed Sci. 1978, 26, 594–596. [Google Scholar] [CrossRef]
- Coffelt, T.A.; Hallock, D.L. Soil fertility responses of Virginia-type peanut cultivars. Agron. J. 1986, 78, 131–137. [Google Scholar] [CrossRef]
- Mayer, A.M.; Poljakoff-Mayber, A. The Germination of Seeds, 4th ed.; Pergamon Press: Oxford, UK, 1989. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B.; Hamill, A.S.; Bernards, M.A. Effects of collection time and after-ripening on chemical constituents and germinability of Scotch thistle (Onopordum acanthium) cypselas. Botany 2012, 90, 755–762. [Google Scholar] [CrossRef]
- Lewak, S.; Rudnicki, R.M. After-ripening in cold-requiring seeds. In The Physiology and Biochemistry of Seed Development, Dormancy and Germination; Khan, A.A., Ed.; Elsevier Biomedical Press: Amsterdam, The Netherlands, 1982; pp. 193–217. [Google Scholar]
- Thomson, J.R. An Introduction to Seed Technology; Leonard Hill: Glasgow, UK, 1979. [Google Scholar]
- Adkins, S.W.; Simpson, G.M. The physiological basis of seed dormancy in Avena fatua. IX. Characterization of two dormancy states. Physiol. Plant. 1988, 73, 15–20. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Germination ecophysiology of seeds of the winter annual Chaerophyllum tainturiei: A new type of morphophysiological dormancy. J. Ecol. 1990, 78, 993–1004. [Google Scholar] [CrossRef]
- Allen, P.S.; Meyer, S.E.; Beckstead, J. Patterns of seed after-ripening in Bromus tectorum L. J. Exp. Bot. 1995, 46, 1737–1744. [Google Scholar] [CrossRef]
- Chaharsoghi, A.T.; Jacobs, B. Manipulating dormancy of Capeweed (Arctotheca calendula L.) seed. Seed Sci. Res. 1998, 8, 139–146. [Google Scholar] [CrossRef]
- Liava, V.; Ntatsi, G.; Karkanis, A. Seed germination of three milk thistle (Silybum marianum (L.) Gaertn.) populations of Greek origin: Temperature, duration, and storage conditions effects. Plants 2023, 12, 1025. [Google Scholar] [CrossRef]
- Westra, R.N.; Loomis, N.E. Seed dormancy in Uniola paniculata. Am. J. Bot. 1966, 53, 407–411. [Google Scholar] [CrossRef]
- Bradbeer, J.W. Studies in seed dormancy. IV. The role of endogenous inhibitors and gibberellin in the dormancy and germination of Corylus avellana L. seeds. Planta 1968, 78, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Beardsell, D.; Mullett, J. Seed germination of Eucalyptus pauciflora Sieb. ex Spreng. from low and high altitude populations in Victoria. Aust. J. Bot. 1984, 32, 475–480. [Google Scholar] [CrossRef]
- Dong, H.; Liu, T.; Liu, Z.; Song, Z. Fate of the soil seed bank of giant ragweed and its significance in preventing and controlling its invasion in grasslands. Ecol. Evol. 2020, 20, 4854–4866. [Google Scholar] [CrossRef]
- Rojas-Aréchiga, M.; García-Morales, E. Dormancy and viability of Ferocactus peninsulae (Cactaceae) seeds. Plant Species Biol. 2022, 37, 173–181. [Google Scholar] [CrossRef]
- Froud-Williams, R.J.; Hilton, J.R.; Dixon, J. Evidence for an endogenous cycle of dormancy in dry stored seeds of Poa trivialis. New Phytol. 1986, 102, 123–131. [Google Scholar] [CrossRef]
- Christensen, M.; Meyer, S.E.; Allen, P.S. A hydrothermal time model of seed after-ripening in Bromus tectorum L. Seed Sci. Res. 1996, 6, 1–9. [Google Scholar] [CrossRef]
- Bauer, M.C.; Meyer, S.E.; Allen, P.S. A simulation model to predict seed dormancy loss in the field for Bromus tectorum L. J. Exp. Bot. 1998, 49, 1235–1244. [Google Scholar]
- Meyer, S.E.; Debaene-Gill, S.B.; Allen, P.S. Using hydrothermal time concepts to model seed germination response to temperature, dormancy loss, and priming effects in Elymus elymoides. Seed Sci. Res. 2000, 10, 213–223. [Google Scholar] [CrossRef]
- Bradford, K.J. Water relations in seed germination. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 351–396. [Google Scholar]
- Narasimhareddy, S.B.; Swamy, P.M. Abscisic acid-like inhibitors and cytokinins during after-ripening of dormant peanut seeds (Arachis hypogaea). Physiol. Plant. 1979, 46, 191–193. [Google Scholar] [CrossRef]
- Rao, M.R.K.; Rao, I.M. Dormancy regulation in groundnut seeds (Arachis hypogaea L.) C. Var. TMV-3. Curr. Sci. 1979, 48, 1000–1001. [Google Scholar]
- Kigel, J. Seed germination in arid and semiarid regions. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 645–699. [Google Scholar]
- Esashi, Y.; Zhang, M.; Segawa, K.; Furihata, T.; Nakaya, M.; Maeda, Y. Possible involvement of volatile compounds in the after-ripening of cocklebur seeds. Physiol. Plant. 1994, 90, 577–583. [Google Scholar] [CrossRef]
- Weidner, S.; Paprocka, J.; Lukaszewicz, D. Changes in free, esterified and glycosidic phenolic acids in cereal grains during the after-ripening. Seed Sci. Technol. 1996, 24, 107–114. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B. Variation in germination response within Scotch thistle, Onopordum acanthium L., populations matured under greenhouse and field conditions. Écoscience 2000, 7, 57–65. [Google Scholar] [CrossRef]
- Moore, R.J.; Frankton, C. The Thistles of Canada; Monograph No. 10; Canada Department of Agriculture: Ottawa, ON, Canada, 1974.
- Smith, H.A.; Johnson, W.S.; Scott Shonkwiler, J.; Swanson, S.R. The implications of variable or constant expansion rates in invasive weed infestations. Weed Sci. 1999, 47, 62–66. [Google Scholar] [CrossRef]
- Cavers, P.B.; Qaderi, M.M.; Threadgill, P.F.; Steel, M.G. The biology of Canadian weeds. 147. Onopordum acanthium L. Can. J. Plant Sci. 2011, 91, 739–758. [Google Scholar] [CrossRef]
- Wei, C.; Zhou, S.; Shi, K.; Zhang, C.; Shao, H. Chemical profile and phytotoxic action of Onopordum acanthium essential oil. Sci. Rep. 2020, 10, 13568. [Google Scholar] [CrossRef] [PubMed]
- Michael, P.W. Control of the biennial thistle, Onopordum, by amitrole and five perennial grasses. Aust. J. Exp. Agric. Anim. Husb. 1968, 8, 331–339. [Google Scholar] [CrossRef]
- Hyde-Wyatt, B.H.; Morris, D.I. The Noxious and Secondary Weeds of Tasmania; Department of Agriculture: Tasmania, Australia, 1980.
- Meier, L.R. Variation in Seeds of Onopordum acanthium. Master’s Thesis, Department of Plant Sciences, University of Western Ontario, London, ON, Canada, 1995. [Google Scholar]
- Scifres, C.J.; McCarty, M.K. Some factors affecting germination and seedling growth of Scotch thistle. Res. Bull. Nebr. Agric. Exp. Stn. 1969, 228, 1–29. [Google Scholar]
- Young, J.A.; Evans, R.A. Germination and persistence of achenes of Scotch thistle. Weed Sci. 1972, 20, 98–101. [Google Scholar] [CrossRef]
- Pérez-García, F. Effect of the origin of cypsela on germination of Onopordum acanthium L. (Asteraceae). Seed Sci. Technol. 1993, 21, 187–195. [Google Scholar]
- Qaderi, M.M.; Cavers, P.B. Effects of dry heat on the germinability and viability of Scotch thistle (Onopordum acanthium) cypselas: Interpopulation and interposition variation. Can. J. Bot. 2003, 81, 684–697. [Google Scholar] [CrossRef]
- Sarabi, V.; Bagheri, A.; Chitband, A.A. Evaluation of different methods for breaking cypsela dormancy in two populations of Scotch thistle (Onopordum acanthium L.) collected from the west and northwest of Iran. Can J. Plant Sci. 2022, 102, 902–910. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qaderi, M.M. Environmental Regulation of Weed Seed Dormancy and Germination. Seeds 2023, 2, 259-277. https://doi.org/10.3390/seeds2030020
Qaderi MM. Environmental Regulation of Weed Seed Dormancy and Germination. Seeds. 2023; 2(3):259-277. https://doi.org/10.3390/seeds2030020
Chicago/Turabian StyleQaderi, Mirwais M. 2023. "Environmental Regulation of Weed Seed Dormancy and Germination" Seeds 2, no. 3: 259-277. https://doi.org/10.3390/seeds2030020
APA StyleQaderi, M. M. (2023). Environmental Regulation of Weed Seed Dormancy and Germination. Seeds, 2(3), 259-277. https://doi.org/10.3390/seeds2030020