
Plasmodial Transcription Factors and Chromatin Modifiers as Drug Targets

 ,
,  and
and
Abstract
:1. Introduction
2. Malaria Parasite Chromatin Dynamics and Therapeutic Prospects
2.1. Histone Acetylases and Histone Deacetylases
2.2. Histone Methyltransferases and Histone Demethylases
2.3. ApiAP2 Transcription Factors as Drug Targets: Implications for Malaria Control
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. Word Malaria Report 2021; WHO: Genva, Switzerland, 2021. [Google Scholar]
- Burda, P.C.; Roelli, M.A.; Schaffner, M.; Khan, S.M.; Janse, C.J.; Heussler, V.T. A Plasmodium Phospholipase Is Involved in Disruption of the Liver Stage Parasitophorous Vacuole Membrane. PLoS Pathog. 2015, 11, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Tavares, J.; Formaglio, P.; Thiberge, S.; Mordelet, E.; Van Rooijen, N.; Medvinsky, A.; Ménard, R.; Amino, R. Role of Host Cell Traversal by the Malaria Sporozoite during Liver Infection. J. Exp. Med. 2013, 210, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A.; Amino, R.; van de Sand, C.; Regen, T.; Retzlaff, S.; Rennenberg, A.; Krueger, A.; Pollok, J.-M.; Menard, R.; Heussler, V.T. Manipulation of Host Hepatocytes by the Malaria Parasite for Delivery into Liver Sinusoids. Science 2006, 313, 1287–1290. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.S.Y.; Blackman, M.J. Malaria Parasite Egress at a Glance. J. Cell Sci. 2021, 134. [Google Scholar] [CrossRef] [PubMed]
- Sinden, R.E. The Biology of Malaria Transmission. In Recent Advances in Malaria Research; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 87–124. ISBN 9781118493816. [Google Scholar]
- Miao, J.; Fan, Q.; Cui, L.; Li, J.; Li, J.; Cui, L. The Malaria Parasite Plasmodium Falciparum Histones: Organization, Expression, and Acetylation. Gene 2006, 369, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.; Papenfuss, A.T.; Mair, G.R.; Janse, C.J.; Vlachou, D.; Waters, A.P.; Cowman, A.F.; Crabb, B.S.; de Koning-Ward, T.F. Molecular Genetics and Comparative Genomics Reveal RNAi Is Not Functional in Malaria Parasites. Nucleic Acids Res. 2009, 37, 3788–3798. [Google Scholar] [CrossRef]
- Trelle, M.B.; Salcedo-Amaya, A.M.; Cohen, A.M.; Stunnenberg, H.G.; Jensen, O.N. Global Histone Analysis by Mass Spectrometry Reveals a High Content of Acetylated Lysine Residues in the Malaria Parasite Plasmodium Falciparum. J. Proteome Res. 2009, 8, 3439–3450. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The Language of Covalent Histone Modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin Modifications and Their Function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef]
- Jabeena, C.A.; Rajavelu, A. Epigenetic Players of Chromatin Structure Regulation in Plasmodium Falciparum. Chembiochem 2019, 20, 1225–1230. [Google Scholar] [CrossRef]
- Callebaut, I.; Prat, K.; Meurice, E.; Mornon, J.-P.; Tomavo, S. Prediction of the General Transcription Factors Associated with RNA Polymerase II in Plasmodium Falciparum: Conserved Features and Differences Relative to Other Eukaryotes. BMC Genom. 2005, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Coulson, R.M.R.; Hall, N.; Ouzounis, C.A. Comparative Genomics of Transcriptional Control in the Human Malaria Parasite Plasmodium Falciparum. Genome Res. 2004, 14, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Balaji, S.; Babu, M.M.; Iyer, L.M.; Aravind, L. Discovery of the Principal Specific Transcription Factors of Apicomplexa and Their Implication for the Evolution of the AP2-Integrase DNA Binding Domains. Nucleic Acids Res. 2005, 33, 3994–4006. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP Family of Plant Transcription Factors. Biol. Chem. 1998, 379, 633–654. [Google Scholar] [CrossRef]
- Jeninga, M.D.; Quinn, J.E.; Petter, M. ApiAP2 Transcription Factors in Apicomplexan Parasites. Pathogens 2019, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- De Silva, E.K.; Gehrke, A.R.; Olszewski, K.; León, I.; Chahal, J.S.; Bulyk, M.L.; Llinás, M. Specific DNA-Binding by Apicomplexan AP2 Transcription Factors. Proc. Natl. Acad. Sci. USA 2008, 105, 8393–8398. [Google Scholar] [CrossRef] [PubMed]
- Campbell, T.L.; De Silva, E.K.; Olszewski, K.L.; Elemento, O.; Llinás, M. Identification and Genome-Wide Prediction of DNA Binding Specificities for the ApiAP2 Family of Regulators from the Malaria Parasite. PLoS Pathog. 2010, 6, e1001165. [Google Scholar] [CrossRef] [PubMed]
- Bartfai, R.; Hoeijmakers, W.A.; Salcedo-Amaya, A.M.; Smits, A.H.; Janssen-Megens, E.; Kaan, A.; Treeck, M.; Gilberger, T.W.; Francoijs, K.J.; Stunnenberg, H.G. H2A.Z Demarcates Intergenic Regions of the Plasmodium Falciparum Epigenome That Are Dynamically Marked by H3K9ac and H3K4me3. PLoS Pathog. 2010, 6, e1001223. [Google Scholar] [CrossRef]
- Foley, M.; Tilley, L. Quinoline Antimalarials: Mechanisms of Action and Resistance and Prospects for New Agents. Pharmacol. Ther. 1998, 79, 55–87. [Google Scholar] [CrossRef]
- Malaria Policy Advisory Committee Meeting. Available online: https://www.who.int/groups/malaria-policy-advisory-group (accessed on 11 January 2023).
- Ma, N.; Zhang, Z.; Liao, F.; Jiang, T.; Tu, Y. The Birth of Artemisinin. Pharmacol. Ther. 2020, 216, 107658. [Google Scholar] [CrossRef] [PubMed]
- Uwimana, A.; Legrand, E.; Stokes, B.H.; Ndikumana, J.M.; Warsame, M.; Umulisa, N.; Ngamije, D.; Munyaneza, T.; Mazarati, J.B.; Munguti, K.; et al. Emergence and Clonal Expansion of in Vitro Artemisinin-Resistant Plasmodium Falciparum Kelch13 R561H Mutant Parasites in Rwanda. Nat. Med. 2020, 26, 1602–1608. [Google Scholar] [CrossRef]
- Cui, L.; Miao, J. Chromatin-Mediated Epigenetic Regulation in the Malaria Parasite Plasmodium Falciparum. Eukaryot. Cell 2010, 9, 1138–1149. [Google Scholar] [CrossRef]
- Joshi, M.B.; Lin, D.T.; Chiang, P.H.; Goldman, N.D.; Fujioka, H.; Aikawa, M.; Syin, C. Molecular Cloning and Nuclear Localization of a Histone Deacetylase Homologue in Plasmodium Falciparum. Mol. Biochem. Parasitol. 1999, 99, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.B.; Leaver, D.J.; Hermans, S.J.; Kelly, G.L.; Brennan, M.S.; Downer, N.L.; Nguyen, N.; Wichmann, J.; McRae, H.M.; Yang, Y.; et al. Inhibitors of Histone Acetyltransferases KAT6A/B Induce Senescence and Arrest Tumour Growth. Nature 2018, 560, 253–257. [Google Scholar] [CrossRef]
- Sen, U.; Nayak, A.; Khurana, J.; Sharma, D.; Gupta, A. Inhibition of PfMYST Histone Acetyltransferase Activity Blocks Plasmodium Falciparum Growth and Survival. Antimicrob. Agents Chemother. 2020, 65. [Google Scholar] [CrossRef]
- Kumar, A.; Bhowmick, K.; Vikramdeo, K.S.; Mondal, N.; Subbarao, N.; Dhar, S.K. Designing Novel Inhibitors against Histone Acetyltransferase (HAT: GCN5) of Plasmodium falciparum. Eur. J. Med. Chem. 2017, 138, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Miao, J.; Cui, L. Cytotoxic Effect of Curcumin on Malaria Parasite Plasmodium Falciparum: Inhibition of Histone Acetylation and Generation of Reactive Oxygen Species. Antimicrob. Agents Chemother. 2007, 51, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Kanyal, A.; Rawat, M.; Gurung, P.; Choubey, D.; Anamika, K.; Karmodiya, K. Genome-Wide Survey and Phylogenetic Analysis of Histone Acetyltransferases and Histone Deacetylases of Plasmodium falciparum. FEBS J. 2018, 285, 1767–1782. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.; Pradhan, A.; Shah, F.; Tekwani, B.L.; Avery, M.A. Structural Insights into the Plasmodium Falciparum Histone Deacetylase 1 (PfHDAC-1): A Novel Target for the Development of Antimalarial Therapy. Bioorg Med. Chem. 2008, 16, 5254–5265. [Google Scholar] [CrossRef] [PubMed]
- Chaal, B.K.; Gupta, A.P.; Wastuwidyaningtyas, B.D.; Luah, Y.H.; Bozdech, Z. Histone Deacetylases Play a Major Role in the Transcriptional Regulation of the Plasmodium falciparum Life Cycle. PLoS Pathog. 2010, 6, e1000737. [Google Scholar] [CrossRef]
- Hu, G.; Cabrera, A.; Kono, M.; Mok, S.; Chaal, B.K.; Haase, S.; Engelberg, K.; Cheemadan, S.; Spielmann, T.; Preiser, P.R.; et al. Transcriptional Profiling of Growth Perturbations of the Human Malaria Parasite Plasmodium falciparum. Nat. Biotechnol. 2010, 28, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Andrews, K.T.; Gupta, A.P.; Tran, T.N.; Fairlie, D.P.; Gobert, G.N.; Bozdech, Z. Comparative Gene Expression Profiling of P. falciparum Malaria Parasites Exposed to Three Different Histone Deacetylase Inhibitors. PLoS ONE 2012, 7, e31847. [Google Scholar] [CrossRef]
- Tonkin, C.J.; Carret, C.K.; Duraisingh, M.T.; Voss, T.S.; Ralph, S.A.; Hommel, M.; Duffy, M.F.; Silva, L.M.d.; Scherf, A.; Ivens, A.; et al. Sir2 Paralogues Cooperate to Regulate Virulence Genes and Antigenic Variation in Plasmodium falciparum. PLoS Biol. 2009, 7, e84. [Google Scholar] [CrossRef]
- Coleman, B.I.; Skillman, K.M.; Jiang, R.H.Y.; Childs, L.M.; Altenhofen, L.M.; Ganter, M.; Leung, Y.; Goldowitz, I.; Kafsack, B.F.C.; Marti, M.; et al. A Plasmodium falciparum Histone Deacetylase Regulates Antigenic Variation and Gametocyte Conversion. Cell Host Microbe 2014, 16, 177–186. [Google Scholar] [CrossRef]
- Mancio-Silva, L.; Slavic, K.; Grilo Ruivo, M.T.; Grosso, A.R.; Modrzynska, K.K.; Vera, I.M.; Sales-Dias, J.; Gomes, A.R.; MacPherson, C.R.; Crozet, P.; et al. Nutrient Sensing Modulates Malaria Parasite Virulence. Nature 2017, 547, 213–216. [Google Scholar] [CrossRef]
- Harris, C.T.; Tong, X.; Campelo, R.; Marreiros, I.M.; Vanheer, L.N.; Nahiyaan, N.; Zuzarte-Luís, V.A.; Deitsch, K.W.; Mota, M.M.; Rhee, K.Y.; et al. Sexual Differentiation in Human Malaria Parasites Is Regulated by Competition between Phospholipid Metabolism and Histone Methylation. Nat. Microbiol. 2023, 8, 1280–1292. [Google Scholar] [CrossRef]
- Huang, Z.; Li, R.; Tang, T.; Ling, D.; Wang, M.; Xu, D.; Sun, M.; Zheng, L.; Zhu, F.; Min, H.; et al. A Novel Multistage Antiplasmodial Inhibitor Targeting Plasmodium falciparum Histone Deacetylase 1. Cell Discov. 2020, 6, 93. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.E.; Lee, J.W.; Bohmer, M.J.; Welden, J.D.; Arshadi, A.K.; Du, L.; Cichewicz, R.H.; Chakrabarti, D. Cyclic Tetrapeptide HDAC Inhibitors with Improved Plasmodium falciparum Selectivity and Killing Profile. ACS Infect. Dis. 2021, 7, 2889–2903. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Ling, D.; Tang, T.; Huang, Z.; Wang, M.; Ding, Y.; Liu, T.; Wei, H.; Xu, W.; Mao, F.; et al. Discovery of Novel Plasmodium falciparum HDAC1 Inhibitors with Dual-Stage Antimalarial Potency and Improved Safety Based on the Clinical Anticancer Drug Candidate Quisinostat. J. Med. Chem. 2021, 64, 2254–2271. [Google Scholar] [CrossRef] [PubMed]
- Hansen, F.; Sumanadasa, S.; Stenzel, K.; Duffy, S.; Meister, S.; Marek, L.; Schmetter, R.; Kuna, K.; Hamacher, A.; Mordmüller, B.; et al. Discovery of HDAC Inhibitors with Potent Activity against Multiple Malaria Parasite Life Cycle Stages. Eur. J. Med. Chem. 2014, 82, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Andrews, K.T.; Haque, A.; Jones, M.K. HDAC Inhibitors in Parasitic Diseases. Immunol. Cell Biol. 2012, 90, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Chua, M.J.; Arnold, M.S.J.; Xu, W.; Lancelot, J.; Lamotte, S.; Späth, G.F.; Prina, E.; Pierce, R.J.; Fairlie, D.P.; Skinner-Adams, T.S.; et al. Effect of Clinically Approved HDAC Inhibitors on Plasmodium, Leishmania and Schistosoma Parasite Growth. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Sumanadasa, S.D.M.; Goodman, C.D.; Lucke, A.J.; Skinner-Adams, T.; Saham, I.; Haque, A.; Do, T.A.; McFadden, G.I.; Fairlie, D.P.; Andrews, K.T. Antimalarial Activity of the Anticancer Histone Deacetylase Inhibitor SB939. Antimicrob. Agents Chemother. 2012, 56, 3849–3856. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tang, T.; Li, R.; Huang, Z.; Ling, D.; Zheng, L.; Ding, Y.; Liu, T.; Xu, W.; Zhu, F.; et al. Drug Repurposing of Quisinostat to Discover Novel Plasmodium falciparum HDAC1 Inhibitors with Enhanced Triple-Stage Antimalarial Activity and Improved Safety. J. Med. Chem. 2022, 65, 4156–4181. [Google Scholar] [CrossRef] [PubMed]
- Marek, L.; Hamacher, A.; Hansen, F.K.; Kuna, K.; Gohlke, H.; Kassack, M.U.; Kurz, T. Histone Deacetylase (HDAC) Inhibitors with a Novel Connecting Unit Linker Region Reveal a Selectivity Profile for HDAC4 and HDAC5 with Improved Activity against Chemoresistant Cancer Cells. J. Med. Chem. 2013, 56, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, N.C.; Andrews, K.T.; Tran, T.L.; Lucke, A.J.; Reid, R.C.; Fairlie, D.P. Antimalarial Histone Deacetylase Inhibitors Containing Cinnamate or NSAID Components. Bioorg Med. Chem. Lett. 2010, 20, 7080–7084. [Google Scholar] [CrossRef]
- Andrews, K.T.; Tran, T.N.; Lucke, A.J.; Kahnberg, P.; Le, G.T.; Boyle, G.M.; Gardiner, D.L.; Skinner-Adams, T.S.; Fairlie, D.P. Potent Antimalarial Activity of Histone Deacetylase Inhibitor Analogues. Antimicrob. Agents Chemother. 2008, 52, 1454–1461. [Google Scholar] [CrossRef] [PubMed]
- Audia, J.E.; Campbell, R.M. Histone Modifications and Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef]
- Cui, L.; Fan, Q.; Miao, J. Histone Lysine Methyltransferases and Demethylases in Plasmodium falciparum. Int. J. Parasitol. 2008, 38, 1083–1097. [Google Scholar] [CrossRef]
- Martin, C.; Zhang, Y. The Diverse Functions of Histone Lysine Methylation. Nat. Rev. Mol. Cell Biol. 2005, 6, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Hollin, T.; Le Roch, K.G. From Genes to Transcripts, a Tightly Regulated Journey in Plasmodium. Front. Cell Infect. Microbiol. 2020, 10, 618454. [Google Scholar] [CrossRef]
- Connacher, J.; von Grüning, H.; Birkholtz, L. Histone Modification Landscapes as a Roadmap for Malaria Parasite Development. Front. Cell Dev. Biol. 2022, 10, 848797. [Google Scholar] [CrossRef]
- Coetzee, N.; Sidoli, S.; Van Biljon, R.; Painter, H.; Llinás, M.; Garcia, B.A.; Birkholtz, L.M. Quantitative Chromatin Proteomics Reveals a Dynamic Histone Post-Translational Modification Landscape That Defines Asexual and Sexual Plasmodium falciparum Parasites. Sci. Rep. 2017, 7, 607. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.F.; Selvarajah, S.A.; Josling, G.A.; Petter, M. Epigenetic Regulation of the Plasmodium falciparum Genome. Brief. Funct. Genom. 2014, 13, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rubio, J.J.; Mancio-Silva, L.; Scherf, A. Genome-Wide Analysis of Heterochromatin Associates Clonally Variant Gene Regulation with Perinuclear Repressive Centers in Malaria Parasites. Cell Host Microbe 2009, 5, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Comeaux, C.A.; Duraisingh, M.T. Unravelling a Histone Code for Malaria Virulence. Mol. Microbiol. 2007, 66, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Malmquist, N.A.; Moss, T.A.; Mecheri, S.; Scherf, A.; Fuchter, M.J. Small-Molecule Histone Methyltransferase Inhibitors Display Rapid Antimalarial Activity against All Blood Stage Forms in Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2012, 109, 16708–16713. [Google Scholar] [CrossRef]
- Volz, J.C.; Bártfai, R.; Petter, M.; Langer, C.; Josling, G.A.; Tsuboi, T.; Schwach, F.; Baum, J.; Rayner, J.C.; Stunnenberg, H.G.; et al. PfSET10, a Plasmodium falciparum Methyltransferase, Maintains the Active Var Gene in a Poised State during Parasite Division. Cell Host Microbe 2012, 11, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Scherf, A.; Hernandez-Rivas, R.; Buffet, P.; Bottius, E.; Benatar, C.; Pouvelle, B.; Gysin, J.; Lanzer, M. Antigenic Variation in Malaria: In Situ Switching, Relaxed and Mutually Exclusive Transcription of Var Genes during Intra-Erythrocytic Development in Plasmodium falciparum. EMBO J. 1998, 17, 5418–5426. [Google Scholar] [CrossRef]
- Chen, Q.; Fernandez, V.; Sundstrom, A.; Schlichtherle, M.; Datta, S.; Hagblom, P.; Wahlgren, M. Developmental Selection of Var Gene Expression in Plasmodium falciparum. Nature 1998, 394, 392–395. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, Y.; Zhang, Y.; Fang, X.; Claes, A.; Duchateau, M.; Namane, A.; Lopez-Rubio, J.J.; Pan, W.; Scherf, A. A Critical Role of Perinuclear Filamentous Actin in Spatial Repositioning and Mutually Exclusive Expression of Virulence Genes in Malaria Parasites. Cell Host Microbe 2011, 10, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Ralph, S.A.; Scheidig-Benatar, C.; Scherf, A. Antigenic Variation in Plasmodium falciparum Is Associated with Movement of Var Loci between Subnuclear Locations. Proc. Natl. Acad. Sci. USA 2005, 102, 5414–5419, ISBN 0027-8424 (Print) 0027-8424 (Linking). [Google Scholar] [CrossRef]
- Duraisingh, M.T.; Voss, T.S.; Marty, A.J.; Duffy, M.F.; Good, R.T.; Thompson, J.K.; Freitas-Junior, L.H.; Scherf, A.; Crabb, B.S.; Cowman, A.F. Heterochromatin Silencing and Locus Repositioning Linked to Regulation of Virulence Genes in Plasmodium falciparum. Cell 2005, 121, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Mu, J.; Zhang, Q.; Ni, T.; Srinivasan, P.; Rayavara, K.; Yang, W.; Turner, L.; Lavstsen, T.; Theander, T.G.; et al. PfSETvs Methylation of Histone H3K36 Represses Virulence Genes in Plasmodium falciparum. Nature 2013, 499, 223–227. [Google Scholar] [CrossRef]
- Read, D.F.; Cook, K.; Lu, Y.Y.; Le Roch, K.G.; Noble, W.S. Predicting Gene Expression in the Human Malaria Parasite Plasmodium falciparum Using Histone Modification, Nucleosome Positioning, and 3D Localization Features. PLoS Comput. Biol. 2019, 15, e1007329. [Google Scholar] [CrossRef]
- Volz, J.; Carvalho, T.G.; Ralph, S.A.; Gilson, P.; Thompson, J.; Tonkin, C.J.; Langer, C.; Crabb, B.S.; Cowman, A.F. Potential Epigenetic Regulatory Proteins Localise to Distinct Nuclear Sub-Compartments in Plasmodium falciparum. Int. J. Parasitol. 2010, 40, 109–121. [Google Scholar] [CrossRef]
- Matthews, K.A.; Senagbe, K.M.; Nötzel, C.; Gonzales, C.A.; Tong, X.; Rijo-Ferreira, F.; Bhanu, N.V.; Miguel-Blanco, C.; Lafuente-Monasterio, M.J.; Garcia, B.A.; et al. Disruption of the Plasmodium falciparum Life Cycle through Transcriptional Reprogramming by Inhibitors of Jumonji Demethylases. ACS Infect. Dis. 2021, 6, 1058–1075. [Google Scholar] [CrossRef] [PubMed]
- Poran, A.; Nötzel, C.; Aly, O.; Mencia-Trinchant, N.; Harris, C.T.; Guzman, M.L.; Hassane, D.C.; Elemento, O.; Kafsack, B.F.C. Single-Cell RNA Sequencing Reveals a Signature of Sexual Commitment in Malaria Parasites. Nature 2017, 551, 95. [Google Scholar] [CrossRef]
- Salcedo-Amaya, A.M.; van Driel, M.A.; Alako, B.T.; Trelle, M.B.; van den Elzen, A.M.; Cohen, A.M.; Janssen-Megens, E.M.; van de Vegte-Bolmer, M.; Selzer, R.R.; Iniguez, A.L.; et al. Dynamic Histone H3 Epigenome Marking during the Intraerythrocytic Cycle of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2009, 106, 9655–9660. [Google Scholar] [CrossRef]
- Copeland, R.A.; Solomon, M.E.; Richon, V.M. Protein Methyltransferases as a Target Class for Drug Discovery. Nat. Rev. Drug Discov. 2009, 8, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, N.; von Grüning, H.; Opperman, D.; van der Watt, M.; Reader, J.; Birkholtz, L.M. Epigenetic Inhibitors Target Multiple Stages of Plasmodium falciparum Parasites. Sci. Rep. 2020, 10, 2355. [Google Scholar] [CrossRef] [PubMed]
- Ngwa, C.J.; Kiesow, M.J.; Orchard, L.M.; Farrukh, A.; Llinás, M.; Pradel, G. The G9a Histone Methyltransferase Inhibitor BIX-01294 Modulates Gene Expression during Plasmodium falciparum Gametocyte Development and Transmission. Int. J. Mol. Sci. 2019, 20, 5087. [Google Scholar] [CrossRef]
- Iwanaga, S.; Kaneko, I.; Kato, T.; Yuda, M. Identification of an AP2-Family Protein That Is Critical for Malaria Liver Stage Development. PLoS ONE 2012, 7, e47557. [Google Scholar] [CrossRef]
- Kaneko, I.; Iwanaga, S.; Kato, T.; Kobayashi, I.; Yuda, M. Genome-Wide Identification of the Target Genes of AP2-O, a Plasmodium AP2-Family Transcription Factor. PLoS Pathog. 2015, 11, e1004905. [Google Scholar] [CrossRef]
- Modrzynska, K.; Pfander, C.; Chappell, L.; Yu, L.; Suarez, C.; Dundas, K.; Gomes, A.R.; Goulding, D.; Rayner, J.C.; Choudhary, J.; et al. A Knockout Screen of ApiAP2 Genes Reveals Networks of Interacting Transcriptional Regulators Controlling the Plasmodium Life Cycle. Cell Host Microbe 2017, 21, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Yuda, M.; Iwanaga, S.; Shigenobu, S.; Mair, G.R.; Janse, C.J.; Waters, A.P.; Kato, T.; Kaneko, I. Identification of a Transcription Factor in the Mosquito-Invasive Stage of Malaria Parasites. Mol. Microbiol. 2009, 71, 1402–1414. [Google Scholar] [CrossRef] [PubMed]
- Yuda, M.; Iwanaga, S.; Shigenobu, S.; Kato, T.; Kaneko, I. Transcription Factor AP2-Sp and Its Target Genes in Malarial Sporozoites. Mol. Microbiol. 2010, 75, 854–863. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.M.; Macpherson, C.R.; Claes, A.; Scheidig-Benatar, C.; Sakamoto, H.; Yam, X.Y.; Preiser, P.; Goel, S.; Wahlgren, M.; Sismeiro, O.; et al. An ApiAP2 Member Regulates Expression of Clonally Variant Genes of the Human Malaria Parasite Plasmodium falciparum. Sci. Rep. 2017, 7, 14042. [Google Scholar] [CrossRef]
- Russell, T.J.; De Silva, E.K.; Crowley, V.M.; Shaw-Saliba, K.; Dube, N.; Josling, G.; Pasaje, C.F.A.; Kouskoumvekaki, I.; Panagiotou, G.; Niles, J.C.; et al. Inhibitors of ApiAP2 Protein DNA Binding Exhibit Multistage Activity against Plasmodium Parasites. PLoS Pathog. 2022, 18, e1010887. [Google Scholar] [CrossRef] [PubMed]
- Flueck, C.; Bartfai, R.; Niederwieser, I.; Witmer, K.; Alako, B.T.F.; Moes, S.; Bozdech, Z.; Jenoe, P.; Stunnenberg, H.G.; Voss, T.S. A Major Role for the Plasmodium falciparum ApiAP2 Protein PfSIP2 in Chromosome End Biology. PLoS Pathog. 2010, 6, e1000784. [Google Scholar] [CrossRef]
- Santos, J.M.; Josling, G.; Ross, P.; Joshi, P.; Orchard, L.; Campbell, T.; Schieler, A.; Cristea, I.M.; Llinás, M. Red Blood Cell Invasion by the Malaria Parasite Is Coordinated by the PfAP2-I Transcription Factor. Cell Host Microbe 2017, 21, 731–741.e10. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Miranda, M.; Vembar, S.S.; Delgadillo, D.M.; Ávila-López, P.A.; Herrera-Solorio, A.M.; Lozano Amado, D.; Vargas, M.; Hernandez-Rivas, R. PfAP2Tel, Harbouring a Non-Canonical DNA-Binding AP2 Domain, Binds to Plasmodium falciparum Telomeres. Cell Microbiol. 2017, 19, e12742. [Google Scholar] [CrossRef]
- Cubillos, E.F.G.; Prata, I.O.; Fotoran, W.L.; Ranford-Cartwright, L.; Wunderlich, G. The Transcription Factor PfAP2-O Influences Virulence Gene Transcription and Sexual Development in Plasmodium falciparum. Front. Cell Infect. Microbiol. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Sinha, A.; Hughes, K.R.; Modrzynska, K.K.; Otto, T.D.; Pfander, C.; Dickens, N.J.; Religa, A.A.; Bushell, E.; Graham, A.L.; Cameron, R.; et al. A Cascade of DNA-Binding Proteins for Sexual Commitment and Development in Plasmodium. Nature 2014, 507, 253–257. [Google Scholar] [CrossRef]
- Kafsack, B.F.C.; Rovira-Graells, N.; Clark, T.G.; Bancells, C.; Crowley, V.M.; Campino, S.G.; Williams, A.E.; Drought, L.G.; Kwiatkowski, D.P.; Baker, D.A.; et al. A Transcriptional Switch Underlies Commitment to Sexual Development in Malaria Parasites. Nature 2014, 507, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Yuda, M.; Iwanaga, S.; Kaneko, I.; Kato, T. Global Transcriptional Repression: An Initial and Essential Step for Plasmodium Sexual Development. Proc. Natl. Acad. Sci. USA 2015, 112, 12824–12829. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Cui, H.; Jiang, Y.; Yang, Z.; Wang, X.; Gao, H.; Liu, C.; Zhang, S.; Su, X.Z.; et al. Systematic CRISPR-Cas9-Mediated Modifications of Plasmodium Yoelii ApiAP2 Genes Reveal Functional Insights into Parasite Development. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Shen, S.; Tang, J.; He, X.; Zhao, Y.; Wang, C.; He, X.; Guo, G.; Liu, M.; Wang, L.; et al. A Cascade of Transcriptional Repression Determines Sexual Commitment and Development in Plasmodium falciparum. Nucleic Acids Res. 2021, 49, 9264–9279. [Google Scholar] [CrossRef]
- Zanghì, G.; Vembar, S.S.; Baumgarten, S.; Ding, S.; Guizetti, J.; Bryant, J.M.; Mattei, D.; Jensen, A.T.R.; Rénia, L.; Goh, Y.S.; et al. A Specific PfEMP1 Is Expressed in P. Falciparum Sporozoites and Plays a Role in Hepatocyte Infection. Cell Rep. 2018, 22, 2951–2963. [Google Scholar] [CrossRef]
- Josling, G.A.; Petter, M.; Oehring, S.C.; Gupta, A.P.; Dietz, O.; Wilson, D.W.; Schubert, T.; Längst, G.; Gilson, P.R.; Crabb, B.S.; et al. A Plasmodium falciparum Bromodomain Protein Regulates Invasion Gene Expression. Cell Host Microbe 2015, 17, 741–751. [Google Scholar] [CrossRef]
- Oladejo, D.O.; Duselu, G.O.; Dokunmu, T.M.; Isewon, I.; Oyelade, J.; Okafor, E.; Iweala, E.E.; Adebiyi, E. In Silico Structure Prediction, Molecular Docking, and Dynamic Simulation of Plasmodium falciparum AP2-I Transcription Factor. Bioinform. Biol. Insights 2023, 17, 11779322221149616. [Google Scholar] [CrossRef] [PubMed]
- Bushell, E.; Gomes, A.R.; Sanderson, T.; Anar, B.; Girling, G.; Herd, C.; Metcalf, T.; Modrzynska, K.; Schwach, F.; Martin, R.E.; et al. Functional Profiling of a Plasmodium Genome Reveals an Abundance of Essential Genes. Cell 2017, 170, 260–272.e8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Casandra, D.; Mayho, M.; et al. Uncovering the Essential Genes of the Human Malaria Parasite Plasmodium falciparum by Saturation Mutagenesis. Science 2018, 360, eaap7847. [Google Scholar] [CrossRef]
- Carrington, E.; Cooijmans, R.H.M.; Keller, D.; Toenhake, C.G.; Bártfai, R.; Voss, T.S. The ApiAP2 Factor PfAP2-HC Is an Integral Component of Heterochromatin in the Malaria Parasite Plasmodium falciparum. iScience 2021, 24, 102444. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Toledo, K.; Rojas-Meza, A.P.; Mancio-Silva, L.; Hernández-Cuevas, N.A.; Delgadillo, D.M.; Vargas, M.; Martínez-Calvillo, S.; Scherf, A.; Hernandez-Rivas, R. Plasmodium falciparum Heterochromatin Protein 1 Binds to Tri-Methylated Histone 3 Lysine 9 and Is Linked to Mutually Exclusive Expression of Var Genes. Nucleic Acids Res. 2009, 37, 2596–2606. [Google Scholar] [CrossRef]
- Shang, X.; Wang, C.; Fan, Y.; Guo, G.; Wang, F.; Zhao, Y.; Sheng, F.; Tang, J.; He, X.; Yu, X.; et al. Genome-Wide Landscape of ApiAP2 Transcription Factors Reveals a Heterochromatin-Associated Regulatory Network during Plasmodium falciparum Blood-Stage Development. Nucleic Acids Res. 2022, 50, 3413–3431. [Google Scholar] [CrossRef]
- Baruch, D.I.; Pasloske, B.L.; Singh, H.B.; Bi, X.; Ma, X.C.; Feldman, M.; Taraschi, T.F.; Howard, R.J. Cloning the P. Falciparum Gene Encoding PfEMP1, a Malarial Variant Antigen and Adherence Receptor on the Surface of Parasitized Human Erythrocytes. Cell 1995, 82, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Quansah, E.; Pappoe, F.; Shen, J.; Liu, M.; Yang, S.; Yu, L.; Zhang, C. ApiAP2 Gene-Network Regulates Gametocytogenesis in Plasmodium Parasites. Cell Microbiol. 2022, 2022, 5796578. [Google Scholar] [CrossRef]
- Droll, D.; Wei, G.; Guo, G.; Fan, Y.; Baumgarten, S.; Zhou, Y.; Xiao, Y.; Scherf, A.; Zhang, Q. Disruption of the RNA Exosome Reveals the Hidden Face of the Malaria Parasite Transcriptome. RNA Biol. 2018, 15, 1206–1214. [Google Scholar] [CrossRef]
- Bancells, C.; Llorà-Batlle, O.; Poran, A.; Nötzel, C.; Rovira-Graells, N.; Elemento, O.; Kafsack, B.F.C.; Cortés, A. Revisiting the Initial Steps of Sexual Development in the Malaria Parasite Plasmodium falciparum. Nat. Microbiol. 2019, 4, 144–154. [Google Scholar] [CrossRef]
- Brancucci, N.M.B.; Bertschi, N.L.; Zhu, L.; Niederwieser, I.; Chin, W.H.; Wampfler, R.; Freymond, C.; Rottmann, M.; Felger, I.; Bozdech, Z.; et al. Heterochromatin Protein 1 Secures Survival and Transmission of Malaria Parasites. Cell Host Microbe 2014, 16, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Yuda, M.; Kaneko, I.; Murata, Y.; Iwanaga, S.; Nishi, T. Mechanisms of Triggering Malaria Gametocytogenesis by AP2-G. Parasitol. Int. 2021, 84, 102403. [Google Scholar] [CrossRef]
- Josling, G.A.; Russell, T.J.; Venezia, J.; Orchard, L.; van Biljon, R.; Painter, H.J.; Llinás, M. Dissecting the Role of PfAP2-G in Malaria Gametocytogenesis. Nat. Commun. 2020, 11, 1503. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Santos, J.M.; Orchard, L.M.; Yamada, N.; van Biljon, R.; Painter, H.J.; Mahony, S.; Llinás, M. The PfAP2-G2 Transcription Factor Is a Critical Regulator of Gametocyte Maturation. Mol. Microbiol. 2021, 115, 1005–1024. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qiao, D.; Wen, Y.; Bi, Y.; Chen, Y.; Huang, Z.; Cui, L.; Guo, J.; Cao, Y. PfAP2-G2 Is Associated to Production and Maturation of Gametocytes in Plasmodium falciparum via Regulating the Expression of PfMDV-1. Front. Microbiol. 2021, 11, 631444. [Google Scholar] [CrossRef]
- Usui, M.; Prajapati, S.K.; Ayanful-Torgby, R.; Acquah, F.K.; Cudjoe, E.; Kakaney, C.; Amponsah, J.A.; Obboh, E.K.; Reddy, D.K.; Barbeau, M.C.; et al. Plasmodium falciparum Sexual Differentiation in Malaria Patients Is Associated with Host Factors and GDV1-Dependent Genes. Nat. Commun. 2019, 10, 2140. [Google Scholar] [CrossRef] [PubMed]
- Yuda, M.; Kaneko, I.; Iwanaga, S.; Murata, Y.; Kato, T. Female-Specific Gene Regulation in Malaria Parasites by an AP2-Family Transcription Factor. Mol. Microbiol. 2020, 113, 40–51. [Google Scholar] [CrossRef]
- Miao, J.; Wang, C.; Lucky, A.B.; Liang, X.; Min, H.; Adapa, S.R.; Jiang, R.; Kim, K.; Cui, L. A Unique GCN5 Histone Acetyltransferase Complex Controls Erythrocyte Invasion and Virulence in the Malaria Parasite Plasmodium falciparum. PLoS Pathog. 2021, 17, e1009351. [Google Scholar] [CrossRef]
- Subudhi, A.K.; Green, J.L.; Satyam, R.; Salunke, R.P.; Lenz, T.; Shuaib, M.; Isaioglou, I.; Abel, S.; Gupta, M.; Esau, L.; et al. DNA-Binding Protein PfAP2-P Regulates Parasite Pathogenesis during Malaria Parasite Blood Stages. Nat. Microbiol. 2023. [Google Scholar] [CrossRef]
- Akkaya, M.; Bansal, A.; Sheehan, P.W.; Pena, M.; Molina-Cruz, A.; Orchard, L.M.; Cimperman, C.K.; Qi, C.F.; Ross, P.; Yazew, T.; et al. A Single-Nucleotide Polymorphism in a Plasmodium berghei ApiAP2 Transcription Factor Alters the Development of Host Immunity. Sci. Adv. 2020, 6, eaaw6957. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cui, H.; Guan, J.; Liu, C.; Yang, Z.; Yuan, J. Plasmodium Transcription Repressor AP2-O3 Regulates Sex-specific Identity of Gene Expression in Female Gametocytes. EMBO Rep. 2021, 22, e51660. [Google Scholar] [CrossRef] [PubMed]
- Birnbaum, J.; Scharf, S.; Schmidt, S.; Jonscher, E.; Maria Hoeijmakers, W.A.; Flemming, S.; Toenhake, C.G.; Schmitt, M.; Sabitzki, R.; Bergmann, B.; et al. A Kelch13-Defined Endocytosis Pathway Mediates Artemisinin Resistance in Malaria Parasites. Science 2020, 367, 51–59. [Google Scholar] [CrossRef] [PubMed]
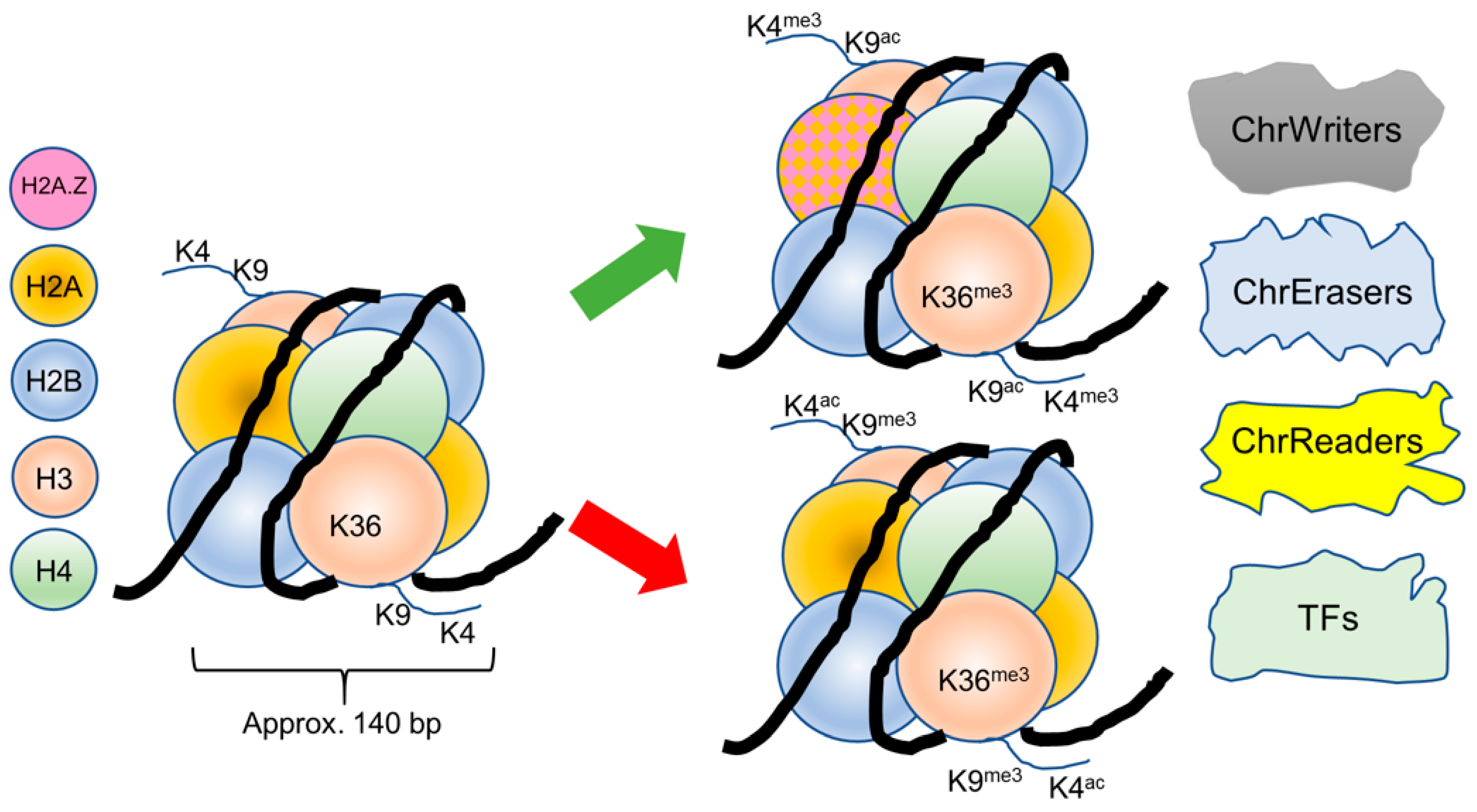

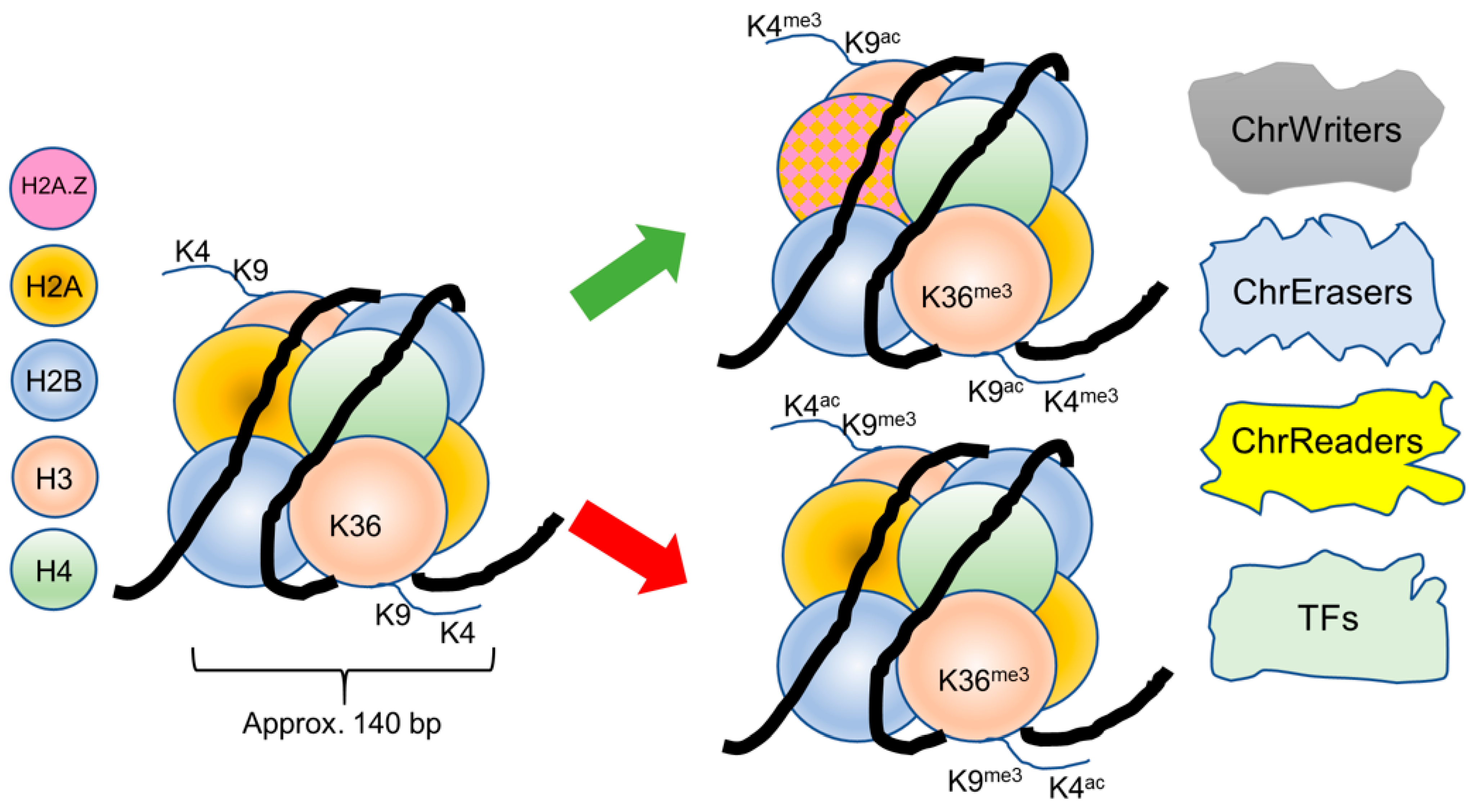{kind=link}
| Gene Encoding ApiAP2 | Stage(s) of Impact | ApiAP2 as Drug Target | Reference | |||
|---|---|---|---|---|---|---|
| Name | P. falciparum | Ortholog in P. berghei | Ortholog in P. yoelii | |||
| AP2-SP2 | PF3D7_0404100 | PBANKA_1001800 | PY17X_1003200 | Oocyst-sporozoites | n.a. | [78,90] |
| n.a. | PF3D7_0420300 | PBANKA_0521700 | PY17X_0523100 | n.a. | n.a. | [90] |
| AP2-O2 | PF3D7_0516800 | PBANKA_1231600 | PY17X_1235000 | Zygote-ookinete | n.a. | [78,90] |
| SIP2 | PF3D7_0604100 | PBANKA_0102900 | PY17X_0104500 | Schizont | n.a. | [83] |
| AP2-EXP2 | PF3D7_0611200 | PBANKA_0109500 | PY17X_0111100 | Trophozoite/schizont | n.a. | [99,113] |
| n.a. | PF3D7_0613800 | PBANKA_0112100 | PY17X_0113700 | n.a. | n.a. | [113] |
| AP2-Tel/SP3 | PF3D7_0622900 | PBANKA_1121800 | PY17X_1123200 | Oocyst-sporozoites | n.a. | [78,85,90] |
| AP2-L | PF3D7_0730300 | PBANKA_0214400 | PY17X_0215800 | Liver stage sporozoites | n.a. | [76] |
| AP2-LT | PF3D7_0802100 | PBANKA_1228100 | PY17X_1231600 | Schizont | n.a. | [18,19,111] |
| n.a. | PF3D7_0934400 | PBANKA_0835200 | PY17X_0838600 | n.a | n.a. | n.a. |
| AP2-I | PF3D7_1007700 | PBANKA_1205900 | PY17X_1209100 | Trophozoite/schizont | [94] | [84] |
| AP2-P | PF3D7_1107800 | PBANKA_0939100 | PY17X_0941600 | Trophozoite/schizont | n.a. | [112] |
| n.a. | PF3D7_1115500 | PBANKA_0932300 | PY17X_0934300 | n.a | n.a. | n.a |
| AP2-G5 | PF3D7_1139300 | PBANKA_0909600 | PY17X_0911000 | During blood stage | n.a. | [99] |
| AP2-O | PF3D7_1143100 | PBANKA_0905900 | PY17X_0907300 | Zygote, ookinete, blood stage, gametocyte | n.a. | [78,79,86,90] |
| AP2-G4 | PF3D7_1222400 | n.a. | n.a. | n.a. | n.a. | [91] |
| AP2-G | PF3D7_1222600 | PBANKA_1437500 | PY17X_1440000 | Asexual-gametocyte | n.a. | [88,90,106]] |
| n.a. | PF3D7_1239200 | PBANKA_1453700 | PY17X_1456200 | n.a. | n.a. | n.a |
| n.a. | PF3D7_1305200 | PBANKA_1403700 | PY17X_1405400 | n.a. | n.a. | n.a |
| AP2-G3/FG | PF3D7_1305200 | PBANKA_1403700 | PY17X_1405400 | Gametocyte | n.a. | [90] |
| n.a. | PF3D7_1342900 | PBANKA_1356000 | PY17X_1361700 | n.a. | n.a. | n.a |
| AP2-O4 | PF3D7_1350900 | PBANKA_1363700 | PY17X_1369400 | Zygote-ookinete | n.a. | [78,90] |
| AP2-G2 | PF3D7_1408200 | PBANKA_1034300 | PY17X_1036700 | Asexual-gametocyte | n.a. | [78,87,89] |
| AP2-O3 | PF3D7_1429200 | PBANKA_1015500 | PY17X_1017000 | Zygote-ookinete | n.a. | [78,90,114] |
| AP2-O5 | PF3D7_1449500 | PBANKA_1313200 | PY17X_1317000 | Zygote-ookinete | n.a. | [90,97] |
| AP2-HC | PF3D7_1456000 | PBANKA_1319700 | PY17X_1323500 | n.a. | n.a. | [78,90,97,99] |
| AP2-SP/EXP | PF3D7_1466400 | PBANKA_1329800 | PY17X_1334500 | Asexual-gametocyte | [82] | [78,79,81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sepulveda, L.F.O.; de Oliveira, G.M.; Chagas, E.H.N.; Wild, N.; da Paz, F.S.; Wrenger, C.; Wunderlich, G. Plasmodial Transcription Factors and Chromatin Modifiers as Drug Targets. Future Pharmacol. 2023, 3, 846-861. https://doi.org/10.3390/futurepharmacol3040051
Sepulveda LFO, de Oliveira GM, Chagas EHN, Wild N, da Paz FS, Wrenger C, Wunderlich G. Plasmodial Transcription Factors and Chromatin Modifiers as Drug Targets. Future Pharmacology. 2023; 3(4):846-861. https://doi.org/10.3390/futurepharmacol3040051
Chicago/Turabian StyleSepulveda, Luisa Fernanda Ortega, Gabriela Mendes de Oliveira, Elaine Hellen Nunes Chagas, Nele Wild, Franciarli Silva da Paz, Carsten Wrenger, and Gerhard Wunderlich. 2023. "Plasmodial Transcription Factors and Chromatin Modifiers as Drug Targets" Future Pharmacology 3, no. 4: 846-861. https://doi.org/10.3390/futurepharmacol3040051
APA StyleSepulveda, L. F. O., de Oliveira, G. M., Chagas, E. H. N., Wild, N., da Paz, F. S., Wrenger, C., & Wunderlich, G. (2023). Plasmodial Transcription Factors and Chromatin Modifiers as Drug Targets. Future Pharmacology, 3(4), 846-861. https://doi.org/10.3390/futurepharmacol3040051