
Filling the Gap in Global Morphotype Set of Filamentous Cyanobacteria: A Novel Case of True Branching in a Non-Heterocytous Cyanobacterium Edaphifilum ginni gen. et sp. nov. (Leptolyngbyales) Isolated from a Semi-Arid Terrain of India

 , , and
, , and
Abstract
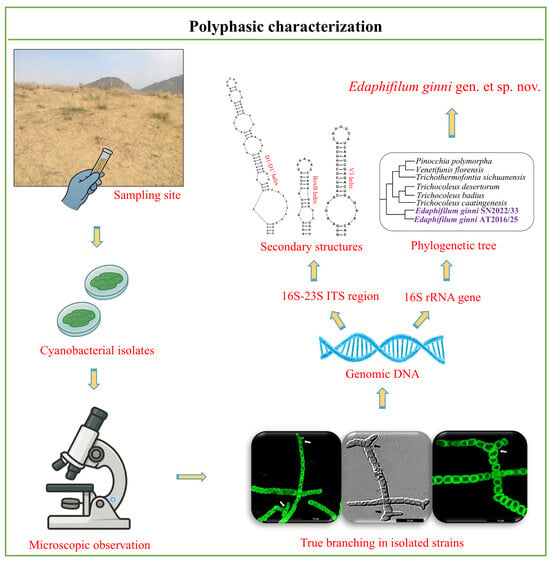
1. Introduction
2. Materials and Methods
2.1. Sampling and Cultivation
2.2. Morphological Analysis
2.3. Developmental Frequency of Branching
2.4. Molecular Characterization
2.5. Phylogenetic Analysis
3. Results
3.1. Taxonomic Description
3.2. Developmental Frequency of Branching
3.3. Molecular and Phylogenetic Analysis
3.4. Analysis of 16S-23S ITS Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olson, J.M. ‘Evolution of photosynthesis’ (1970), re-examined thirty years later. Photosynth. Res. 2001, 68, 95–112. [Google Scholar] [CrossRef]
- Kulasooriya, S. Cyanobacteria: Pioneers of planet earth. Ceylon J. Sci. Biol. Sci. 2012, 40, 71–88. [Google Scholar] [CrossRef]
- Sonam, S.; Dadheech, P.K. Cyanobacteria—The pioneering photoautotrophs. In Cyanobacteria; Elsevier: Amsterdam, The Netherlands, 2024; pp. 1–18. [Google Scholar] [CrossRef]
- Castenholz, R.W. Oxygenic photosynthetic bacteria (Sect. 19). Group l. Cyanobacteria. Bergey’s Man. Syst. Bacteriol. 1989, 3, 1710–1799. [Google Scholar]
- Whitton, B.A.; Potts, M. Introduction to the cyanobacteria. In Ecology of Cyanobacteria II; Springer: Dordrecht, The Netherlands, 2012; pp. 1–13. [Google Scholar]
- Waterbury, J.B. The cyanobacteria—Isolation, purification and identification. In The Prokaryotes; Springer: New York, NY, USA, 2006; pp. 1053–1073. [Google Scholar]
- Seckbach, J. Genesis—In the Beginning; Seckbach, J., Ed.; Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 1999; Volume 22, ISBN 978-94-007-2940-7. [Google Scholar]
- Dvořák, P.; Hašler, P.; Casamatta, D.A.; Poulíčková, A. Underestimated cyanobacterial diversity: Trends and perspectives of research in tropical environments. Fottea 2021, 21, 110–127. [Google Scholar] [CrossRef]
- Chakraborty, S.; Maruthanayagam, V.; Achari, A.; Pramanik, A.; Jaisankar, P.; Mukherjee, J. Euryhalinema mangrovii gen. nov., sp. nov. and Leptoelongatus litoralis gen. nov., sp. nov. (Leptolyngbyaceae) isolated from an Indian mangrove forest. Phytotaxa 2019, 422, 58–74. [Google Scholar] [CrossRef]
- Chavadar, M.; Saraf, A.; Suradkar, A.; Mishra, D.; Kumar, N.; Singh, P. Constrictifilum karadense gen. et sp. nov., a new nostocalean genus from Maharashtra, India. FEMS Microbiol. Lett. 2021, 368, fnab057. [Google Scholar] [CrossRef]
- Kumar, N.; Saraf, A.; Pal, S.; Mishra, D.; Singh, P. Insights into the phylogenetic inconsistencies of the genus Amazonocrinis and description of epilithic Amazonocrinis malviyae sp. nov. (Cyanobacteria, Nostocales) from Jammu and Kashmir, India. Int. J. Syst. Evol. Microbiol. 2022, 72, 005658. [Google Scholar] [CrossRef]
- Sonam, S.; Anand, S.; Pareek, N.; Singh, P.; Casamatta, D.A.; Dadheech, P.K. The novel halotolerant, filamentous cyanobacterium Krienitziella sambharensis gen. et sp. nov. (Nodosilineales, Cyanobacteriophyta) isolated from an Indian wetland (Sambhar Salt Lake, India). Diversity 2026, 18, 181. [Google Scholar] [CrossRef]
- Akagha, M.U.; Pietrasiak, N.; Bustos, D.F.; Vondrášková, A.; Lamb, S.C.; Johansen, J.R. Albertania and Egbenema gen. nov. from Nigeria and the United States, expanding biodiversity in the oculatellaceae (Cyanobacteria). J. Phycol. 2023, 59, 1217–1236. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, R.; Luz, R.; Lage, S.; Menezes, C.; Dias, E.; Flores, C.; Fonseca, A.; Gonçalves, V. Metabolomic and taxonomic characterization of Haloleptolyngbya lusitanica sp. nov. (Cyanobacteria, Synechococcales). Phycologia 2024, 63, 225–234. [Google Scholar] [CrossRef]
- Jusko, B.M.; Johansen, J.R. Description of six new cyanobacterial species from soil biocrusts on san nicolas island, california, in three genera previously restricted to Brazil. J. Phycol. 2024, 60, 133–151. [Google Scholar] [CrossRef]
- Pal, S.; Saraf, A.; Kumar, N.; Singh, P. Igniting taxonomic curiosity: The amazing story of Amazonocrinis with the description of a new genus Ahomia gen. nov. and novel species of Ahomia, Amazonocrinis, and Dendronalium from the biodiversity-rich northeast region of India. J. Phycol. 2024, 60, 387–408. [Google Scholar] [CrossRef]
- Pal, S.; Saraf, A.; Kumar, N.; Pant, H.; Badruddin, S.S.A.; Sajibulla, S.M.A.; Nijamuddin, S.K.; Prajapati, A.; Talukdar, U.; Kohar, N.; et al. Polyphasic characterization of 15 heterocytous cyanobacterial isolates from different habitats of India and description of 9 novel species belonging to the genera Desikacharya, Aliinostoc, and Desmonostoc. Algal Res. 2025, 86, 103873. [Google Scholar] [CrossRef]
- Pant, H.; Kumar, N.; Pal, S.; Singh, P. Exploring cyanobacteria from diverse habitats of the Konkan region of India, unveiling novel species of the genera Desikacharya, Pseudoaliinostoc, and Chlorogloeopsis using a polyphasic approach. J. Phycol. 2025, 61, 194–217. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Komárek, J. A polyphasic approach for the taxonomy of cyanobacteria: Principles and applications. Eur. J. Phycol. 2016, 51, 346–353. [Google Scholar] [CrossRef]
- Strunecký, O.; Ivanova, A.P.; Mareš, J. An updated classification of cyanobacterial orders and families based on phylogenomic and polyphasic analysis. J. Phycol. 2023, 59, 12–51. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Mai, T.; Johansen, J.R.; Pietrasiak, N.; Bohunická, M.; Martin, M.P. Revision of the synechococcales (cyanobacteria) through recognition of four families including Oculatellaceae fam. nov. and Trichocoleaceae fam. nov. and six new genera containing 14 species. Phytotaxa 2018, 365, 1–59. [Google Scholar] [CrossRef]
- Anagnostidis, K. Nomenclatural changes in cyanoprokaryotic order oscillatoriales. Preslia 2001, 73, 359–375. [Google Scholar]
- Tang, J.; Zhou, H.; Jiang, Y.; Yao, D.; Waleron, K.F.; Du, L.-M.; Daroch, M. Characterization of a novel thermophilic cyanobacterium within Trichocoleusaceae, Trichothermofontia sichuanensis gen. et sp. nov., and Its CO2-concentrating mechanism. Front. Microbiol. 2023, 14, 1111809. [Google Scholar] [CrossRef] [PubMed]
- Luz, R.; Cordeiro, R.; Kaštovský, J.; Johansen, J.R.; Dias, E.; Fonseca, A.; Urbatzka, R.; Vasconcelos, V.; Gonçalves, V. New terrestrial cyanobacteria from the Azores Islands: Description of Venetifunis gen. nov. and new species of Albertania, Kovacikia and Pegethrix. Phycologia 2023, 62, 483–498. [Google Scholar] [CrossRef]
- Johansen, J.R.; Casamatta, D.A. Recognizing cyanobacterial diversity through adoption of a new species paradigm. Algol. Stud. 2005, 117, 71–93. [Google Scholar] [CrossRef]
- Komárek, J. Cyanoprokaryota 2. Teil/2nd Part: Oscillatoriales; Büdel, B., Krienitz, L., Gartner, G., Schagerl, M., Eds.; Süsswasserflora von Mitteleuropa; Elsevier GmbH: Amsterdam, The Netherlands, 2005; Volume 19. [Google Scholar]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Iteman, I.; Rippka, R.; Tandeau de Marsac, N.; Herdman, M. Comparison of conserved structural and regulatory domains within divergent 16S rRNA–23S rRNA spacer sequences of cyanobacteria. Microbiology 2000, 146, 1275–1286. [Google Scholar] [CrossRef]
- Pal, S.; Saraf, A.; Kumar, N.; Singh, P. Phycological exploration of the global biodiversity hotspots of northeast India: Discovery of a new species of soil-dwelling cyanobacteria, Desikacharya kailashaharensis sp. nov. FEMS Microbiol. Lett. 2022, 369, fnac099. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S Ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A. Figtree, Version 1.4.4; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2018. Available online: https://tree.bio.ed.ac.uk/software/figtree/ (accessed on 12 February 2026).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Reuter, J.S.; Mathews, D.H. RNAstructure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Inkscape Project Inkscape, Version 1.2.1; Inkscape: Boston, MA, USA, 2022. Available online: https://Inkscape.Org (accessed on 12 February 2026).
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular Blue-Green Algae (Order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Golubić, S.; Hernández-Mariné, M.; Hoffmann, L. Developmental aspects of branching in filamentous cyanophyta/cyanobacteria. Algol. Stud. 1996, 83, 303–329. [Google Scholar] [CrossRef]
- Gugger, M.F.; Hoffmann, L. Polyphyly of true branching cyanobacteria (Stigonematales). Int. J. Syst. Evol. Microbiol. 2004, 54, 349–357. [Google Scholar] [CrossRef]
- Kabirnataj, S.; Nematzadeh, G.A.; Talebi, A.F.; Tabatabaei, M.; Singh, P. Neowestiellopsis gen. nov., a new genus of true branched cyanobacteria with the description of Neowestiellopsis persica sp. nov. and Neowestiellopsis bilateralis sp. nov., isolated from Iran. Plant Syst. Evol. 2018, 304, 501–510. [Google Scholar] [CrossRef]
- Kilgore, C.; Johansen, J.R.; Mai, T.; Hauer, T.; Casamatta, D.A.; Sheil, C. Molecular characterization of Geitleria appalachiana sp. nov. (Nostocales, Cyanobacteria) and formation of Geitleriaceae fam. nov. Fottea 2018, 18, 150–163. [Google Scholar] [CrossRef]
- Casamatta, D.A.; Villanueva, C.D.; Garvey, A.D.; Stocks, H.S.; Vaccarino, M.; Dvořák, P.; Hašler, P.; Johansen, J.R. Reptodigitus chapmanii (Nostocales, Hapalosiphonaceae) gen. nov.: A unique nostocalean (cyanobacteria) genus based on a polyphasic approach. J. Phycol. 2020, 56, 425–436. [Google Scholar] [CrossRef]
- Song, G.; Jiang, Y.; Li, R. Scytolyngbya timoleontis, gen. et sp. nov. (Leptolyngbyaceae, Cyanobacteria): A novel false branching cyanobacteria from China. Phytotaxa 2015, 224, 72–84. [Google Scholar] [CrossRef]
- Jaiswal, T.P.; Chakraborty, S.; Singh, P.; Mishra, A.K.; Singh, S.S. Description of hot spring dwelling Mastigocladus ambikapurensis sp. nov., using a polyphasic approach. Plant Syst. Evol. 2021, 307, 33. [Google Scholar] [CrossRef]
- Pokorný, J.; Vondrášková, A.; Wipplingerová, M.; Kaštovský, J. Four novel taxa of cyanobacteria from a unique thermal cave habitat in Vromoner Canyon, Albania. J. Phycol. 2025, 61, 1394–1422. [Google Scholar] [CrossRef] [PubMed]
- Muhlsteinova, R.; Johansen, J.R.; Pietrasiak, N.; Martin, M.P.; Osorio-Santos, K.; Warren, S.D. Polyphasic characterization of Trichocoleus desertorum sp. nov. (Pseudanabaenales, Cyanobacteria) from desert soils and phylogenetic placement of the genus Trichocoleus. Phytotaxa 2014, 163, 241–261. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152. [Google Scholar]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.-H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Komárek, J. Recent changes (2008) in cyanobacteria taxonomy based on a combination of molecular background with phenotype and ecological consequences (Genus and Species Concept). Hydrobiologia 2010, 639, 245–259. [Google Scholar] [CrossRef]
- Boyer, S.L.; Flechtner, V.R.; Johansen, J.R. Is the 16S-23S rRNA internal transcribed spacer region a good tool for use in molecular systematics and population genetics? A case study in cyanobacteria. Mol. Biol. Evol. 2001, 18, 1057–1069. [Google Scholar] [CrossRef]
- Johansen, J.R.; Kovacik, L.; Casamatta, D.A.; Iková, K.F.; Kaštovský, J. Utility of 16S-23S ITS sequence and secondary structure for recognition of intrageneric and intergeneric limits within cyanobacterial taxa: Leptolyngbya corticola sp. nov. (Pseudanabaenaceae, Cyanobacteria). Nova Hedwig. 2011, 92, 283–302. [Google Scholar] [CrossRef]
- Strunecký, O.; Komárek, J.; Elster, J. Biogeography of Phormidium Autumnale (Oscillatoriales, Cyanobacteria) in western and central spitsbergen. Pol. Polar Res. 2012, 33, 369–382. [Google Scholar] [CrossRef]
- Dvořák, P.; Jahodářová, E.; Hašler, P.; Gusev, E.; Poulíčková, A. A New Tropical Cyanobacterium Pinocchia polymorpha gen. et sp. nov. derived from the genus Pseudanabaena. Fottea 2015, 15, 113–120. [Google Scholar] [CrossRef]
- Singh, J.S.; Kumar, A.; Rai, A.N.; Singh, D.P. Cyanobacteria: A precious bio-resource in agriculture, ecosystem, and environmental sustainability. Front. Microbiol. 2016, 7, 529. [Google Scholar] [CrossRef]
- Mehta, D.J.; Yadav, S.M. Long-Term trend analysis of climate variables for arid and semi-arid regions of an Indian State Rajasthan. Int. J. Hydrol. Sci. Technol. 2022, 13, 191. [Google Scholar] [CrossRef]
- Singh, R.B.; Kumar, A. Climate variability and water resource scarcity in drylands of Rajasthan, India. Geoenviron. Disasters 2015, 2, 7. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Isayenkov, S.V. Metabolites facilitating adaptation of desert cyanobacteria to extremely arid environments. Plants 2022, 11, 3225. [Google Scholar] [CrossRef]
- Feng, S.; Fu, Q. Expansion of global drylands under a warming climate. Atmos. Chem. Phys. 2013, 13, 10081–10094. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Change 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Burrell, A.L.; Evans, J.P.; De Kauwe, M.G. Anthropogenic climate change has driven over 5 million km2 of drylands towards desertification. Nat. Commun. 2020, 11, 3853. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Genus | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1. E. ginni SN2022/33 | - | |||||||||
| 2. E. ginni AT2016/25 | 100 | |||||||||
| 3. Trichocoleus sp. ACSSI 315 MT425936.1 | 94.01 | 94.01 | ||||||||
| 4. T. desertorum SIK19 clone 2 MZ677340.1 | 93.43 | 93.43 | 94.67 | |||||||
| 5. T. desertorum ATA4-8-CV2 KF307604.1 | 94.62 | 94.62 | 95.93 | 98.98 | ||||||
| 6. T. caatingensis CATCD2 NR172612.1 | 94.81 | 94.81 | 96.39 | 97.96 | 97.87 | |||||
| 7. T. badius CRS-1 EF429296.1 | 94.71 | 94.71 | 95.64 | 99.44 | 98.89 | 97.96 | ||||
| 8. Venetifunis florensis BACA0587 NR191033.1 | 93.41 | 93.41 | 93.99 | 92.47 | 93.87 | 94.34 | 94.33 | |||
| 9. Pinocchia polymorpha E5 KP640605.1 | 92.92 | 92.92 | 92.55 | 92.55 | 93.02 | 92.64 | 92.45 | 93.20 | ||
| 10. Trichothermofontia sichuanensis B231 CP110848.1 | 91.57 | 91.57 | 92.72 | 92.42 | 93.52 | 93.42 | 93.33 | 92.32 | 92.74 | - |
| E. ginni (This Study) | Trichocoleus delicatulus | T. desertorum ATA4-8-CV | T. caatingensis CATCD2 | T. badius | Trichothermofontia sichuanensis B231 | Venetifunis florensis BACA0587 | Pinocchia polymorpha E5 | |
|---|---|---|---|---|---|---|---|---|
| Filament | Straight to slightly wavy, solitary | Solitary | Straight to slightly wavy | Entangled | Densely entangled, unbranched | Solitary, entangle, or curved | Straight to slightly wavy, solitary | Solitary or in colony (mats) |
| True branching | Present (V & T type) | Absent | Absent | Absent | Absent | Absent | Absent | Absent |
| Sheath | Thick in old culture, colorless, non-lamellated, rarely 2 trichomes in sheath | Mucilaginous, colorless, numerous trichomes in sheath | Thick or thin, lamellated, multiple trichomes in sheath | Firm, thin, colorless, hyaline | Firm, wide, open, 1 or 2 trichomes in sheath | Thin, colorless | Diffluent mucilage | Thin, colorless and facultative |
| Cell width × length (µm) | 1.5–2.5 (3) × 2.0–3.5 | 1.5–2.5 | 2.5–3.8 × 1.5–5.5 | 2.0 × 2.0–4.0 | 1.0–1.3 × 2.0 | 2.0–2.4 × 2.0–5.0 | 0.7–1.2 × 2.1–5.8 | 1.09–2.86 × 1.28–8.63 (12) |
| Cell shape | Slightly longer than wide or isodiametric | Somewhat longer than wide | Quadratic as well as shorter or much longer than wide | Isodiametric | Longer than wide | Longer than wide | Longer than wide | Longer than wide |
| Constrictions at cross-wall | Distinct | Not available | Distinct | Distinct | Distinct | Non-constricted | Distinct | Distinct |
| Necridic cells | Present | Not available | Present | Not available | Not available | Not available | Absent | Absent |
| Apical cell | Obtuse-rounded to conical, sharply pointed, 3.3–6.0 µm long | Cylindrical and rounded | Conical to sharply pointed, 3.5–9.3 µm long, 1.3–2.8 µm wide | Conical-obtuse to sharply pointed, 4.0–5.0 µm long | Bluntly rounded | Conical, sharply pointed | Rounded | Conical, or rounded, pointed |
| Extrusions | Present | Absent | Absent | Absent | Absent | Absent | Absent | Absent |
| Motility | Absent | Not available | Present | Not available | Not available | Not available | Present | Present |
| Occurrence | Biological crust on sandy soil, Pushkar, Rajasthan, India | Freshwater, metaphytic and epiphytic, England | Desert soils, Atacama Desert, Chile | Topsoil, Brazil | Quartzite seep wall, North Carolina | Thermal spring, Sichuan Province, China | Aerophytic in rock substrate, Flores Island, Portugal | Plankton and periphyton, Lake Hồ Đâu Co, Vietnam |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Share and Cite
Tomer, A.K.; Sonam, S.; Pareek, N.; Anand, S.; Singh, P.; Casamatta, D.A.; Dadheech, P.K. Filling the Gap in Global Morphotype Set of Filamentous Cyanobacteria: A Novel Case of True Branching in a Non-Heterocytous Cyanobacterium Edaphifilum ginni gen. et sp. nov. (Leptolyngbyales) Isolated from a Semi-Arid Terrain of India. Phycology 2026, 6, 56. https://doi.org/10.3390/phycology6020056
Tomer AK, Sonam S, Pareek N, Anand S, Singh P, Casamatta DA, Dadheech PK. Filling the Gap in Global Morphotype Set of Filamentous Cyanobacteria: A Novel Case of True Branching in a Non-Heterocytous Cyanobacterium Edaphifilum ginni gen. et sp. nov. (Leptolyngbyales) Isolated from a Semi-Arid Terrain of India. Phycology. 2026; 6(2):56. https://doi.org/10.3390/phycology6020056
Chicago/Turabian StyleTomer, Anuj Kumar, Sonam Sonam, Nidhi Pareek, Shaubhik Anand, Prashant Singh, Dale A. Casamatta, and Pawan K. Dadheech. 2026. "Filling the Gap in Global Morphotype Set of Filamentous Cyanobacteria: A Novel Case of True Branching in a Non-Heterocytous Cyanobacterium Edaphifilum ginni gen. et sp. nov. (Leptolyngbyales) Isolated from a Semi-Arid Terrain of India" Phycology 6, no. 2: 56. https://doi.org/10.3390/phycology6020056
APA StyleTomer, A. K., Sonam, S., Pareek, N., Anand, S., Singh, P., Casamatta, D. A., & Dadheech, P. K. (2026). Filling the Gap in Global Morphotype Set of Filamentous Cyanobacteria: A Novel Case of True Branching in a Non-Heterocytous Cyanobacterium Edaphifilum ginni gen. et sp. nov. (Leptolyngbyales) Isolated from a Semi-Arid Terrain of India. Phycology, 6(2), 56. https://doi.org/10.3390/phycology6020056