
Temperature and Temperature Humidity Index Changes during Summer to Autumn in a Temperate Zone May Affect Liveweight Gain and Feed Efficiency in Sheep

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics and Experimental Design
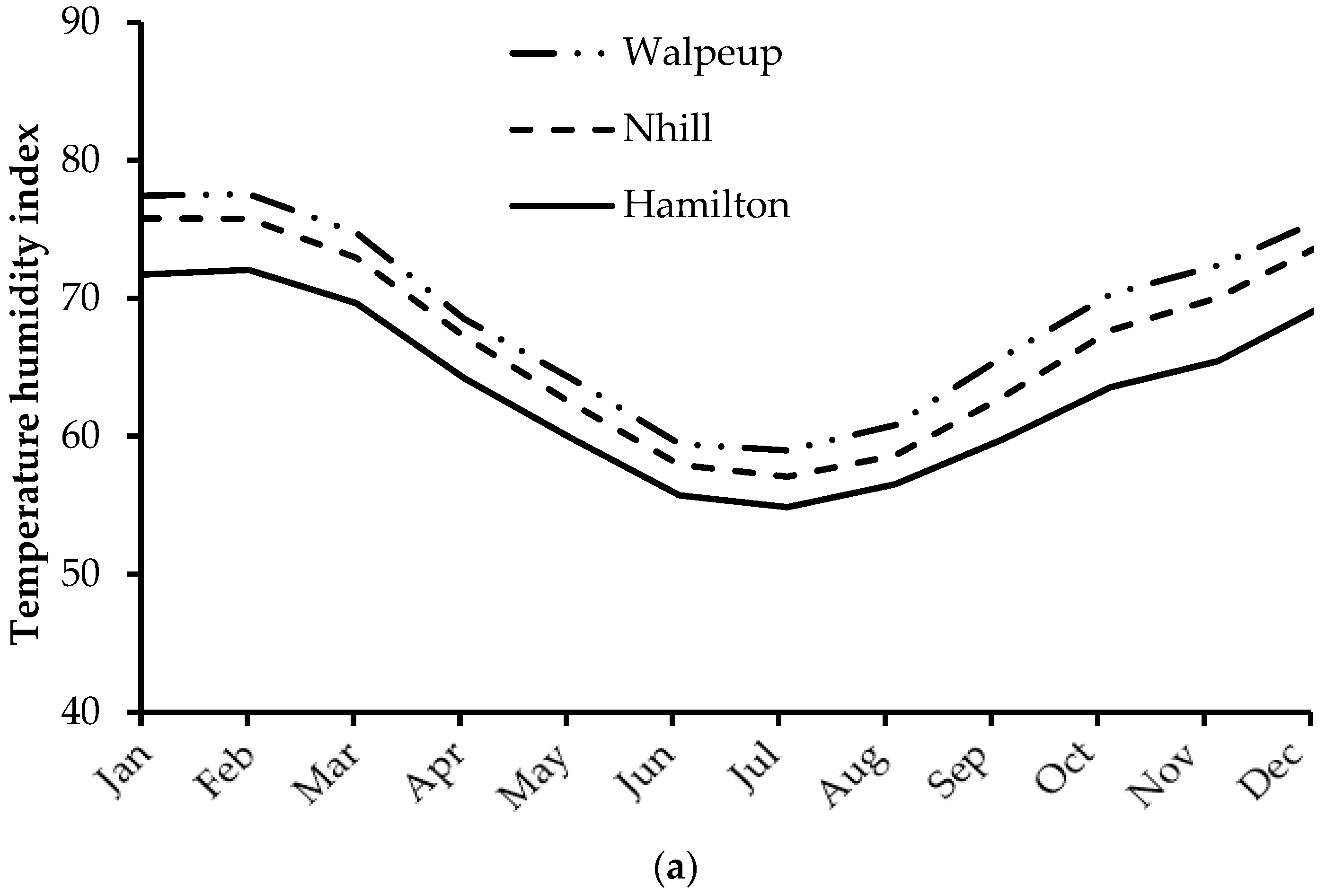
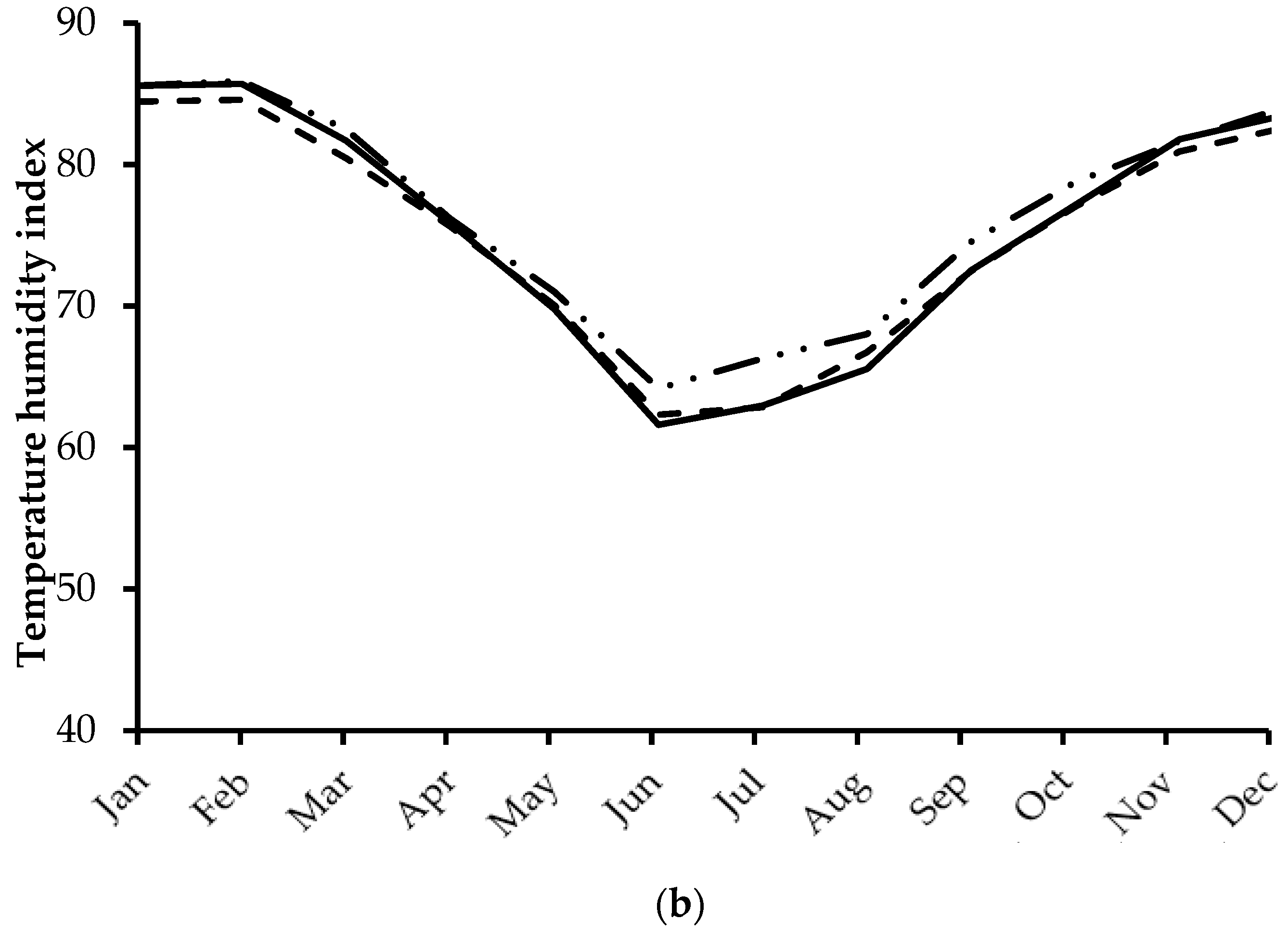
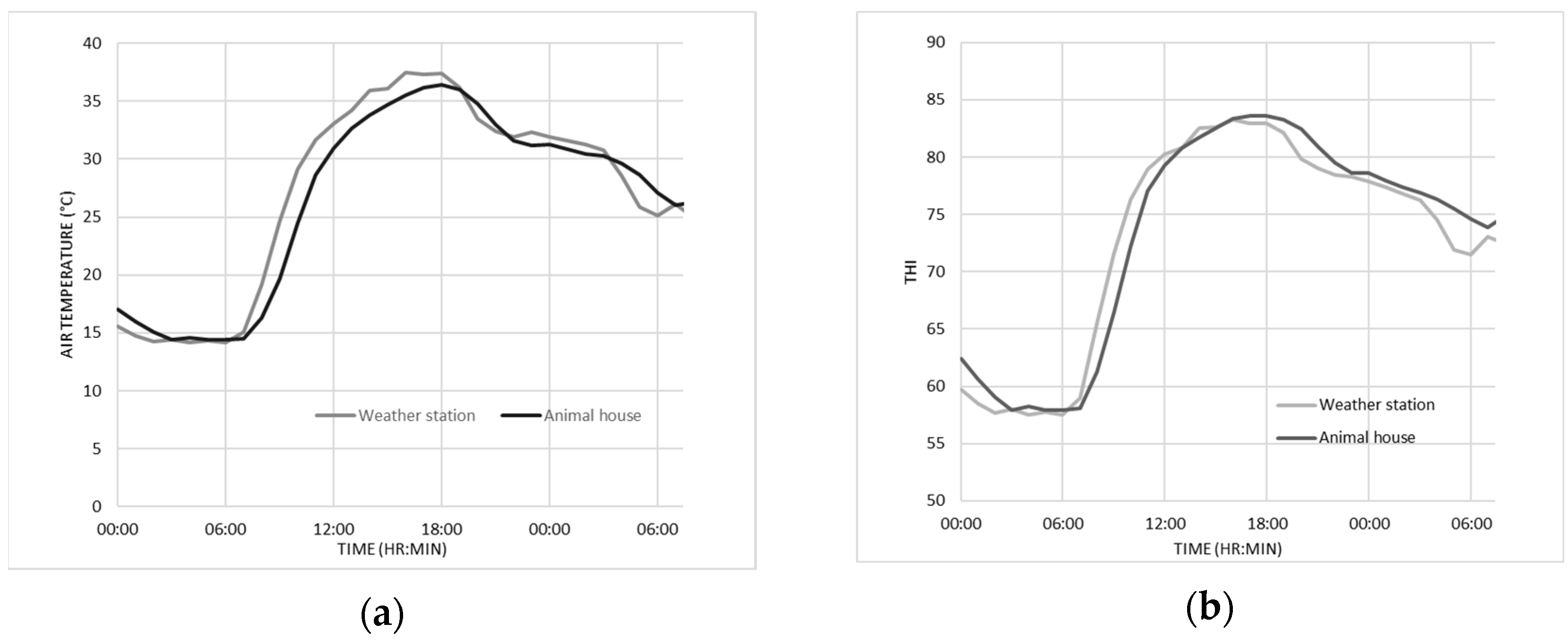
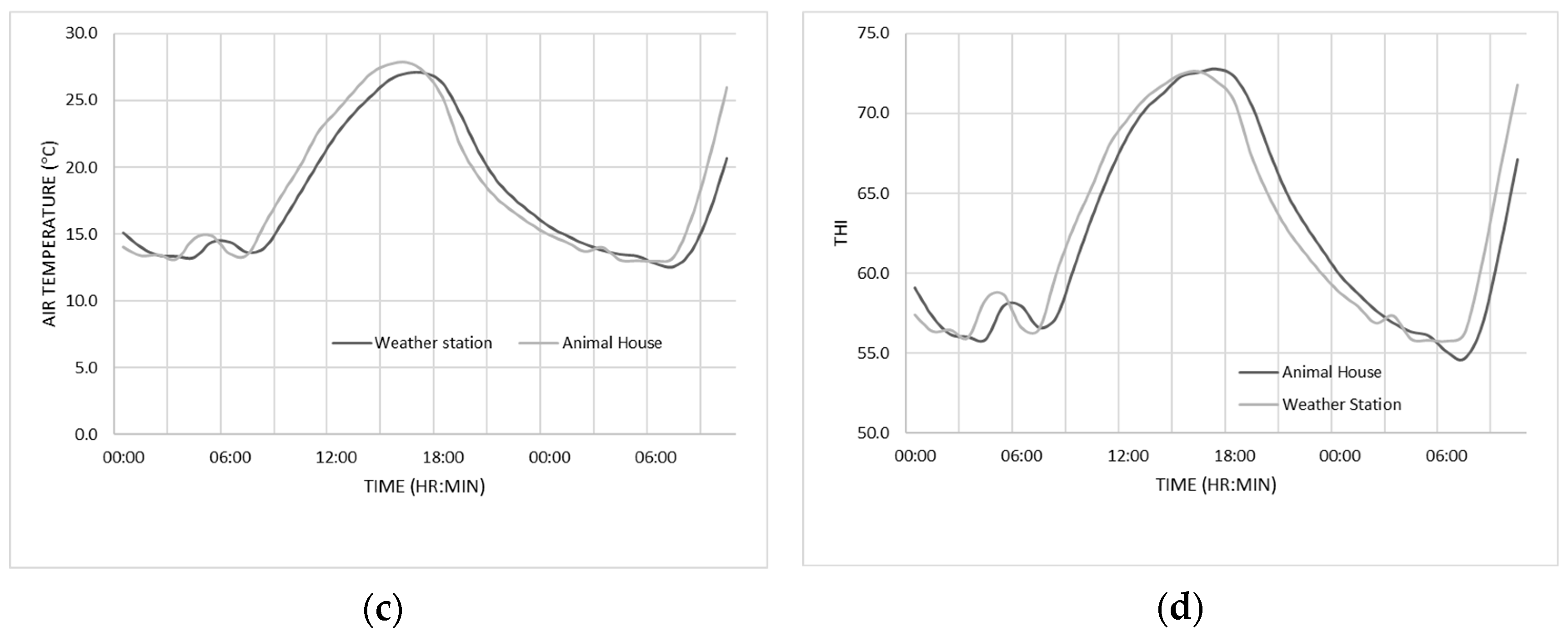
2.2. Environmental Conditions
Calculation of Temperature Humidity Index
2.3. Animal Feeding and Measurements
2.4. Selection of Temperature and THI Thresholds as Reference Values
2.5. Statistical Analysis
3. Results
3.1. Environmental Conditions and Ranges of Temperature and THI Observed during the Experiment
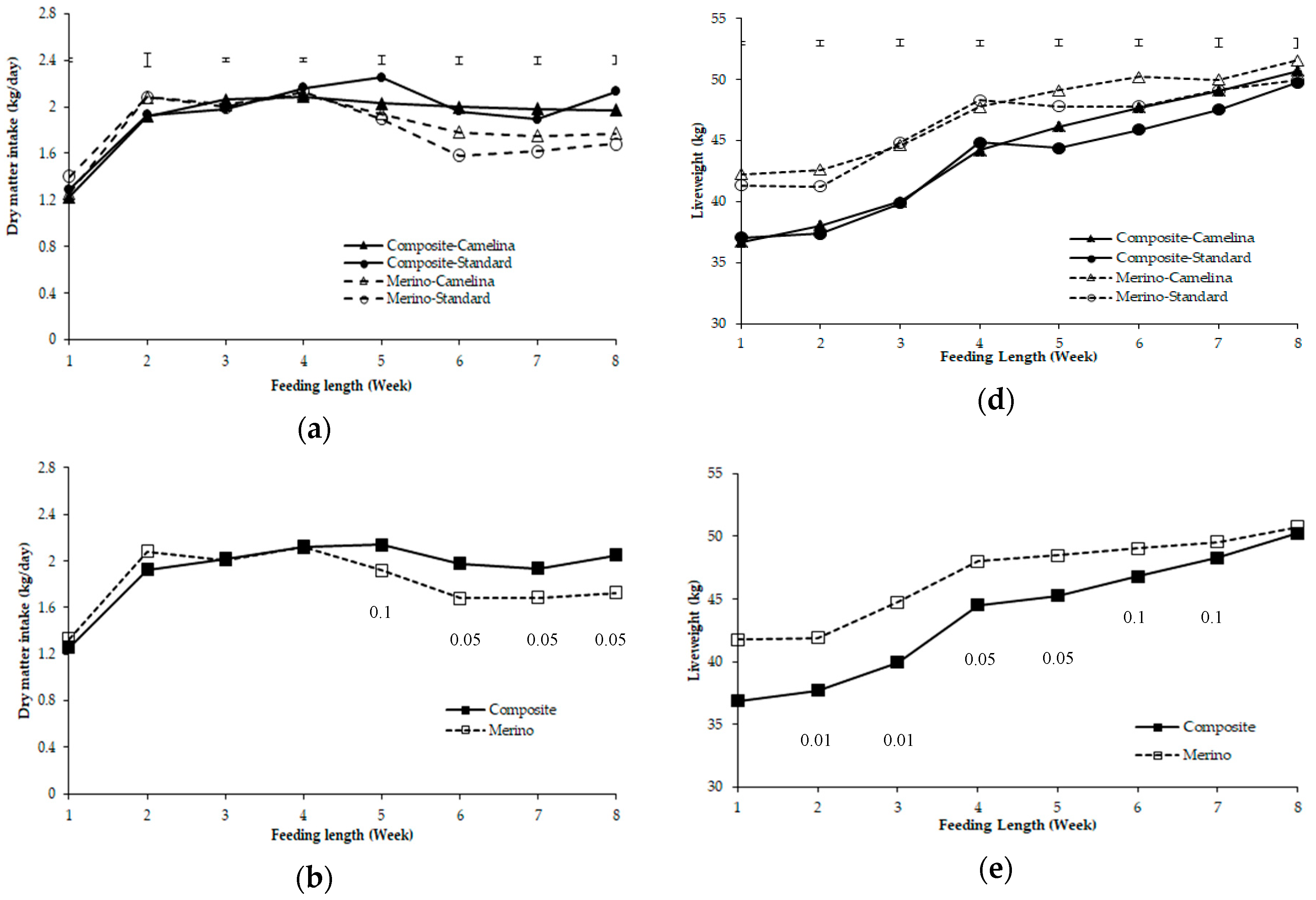
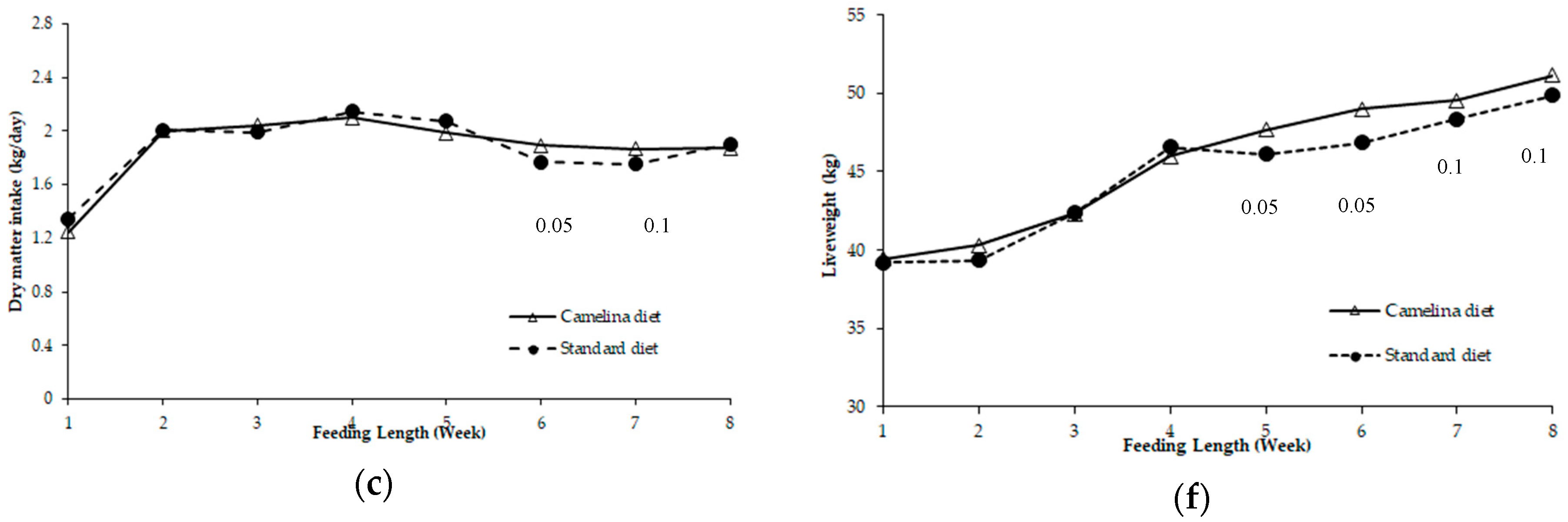
3.2. Dry Matter Intake and Liveweight of Composite Lambs and Merino Yearlings during Weeks 1–8 Period
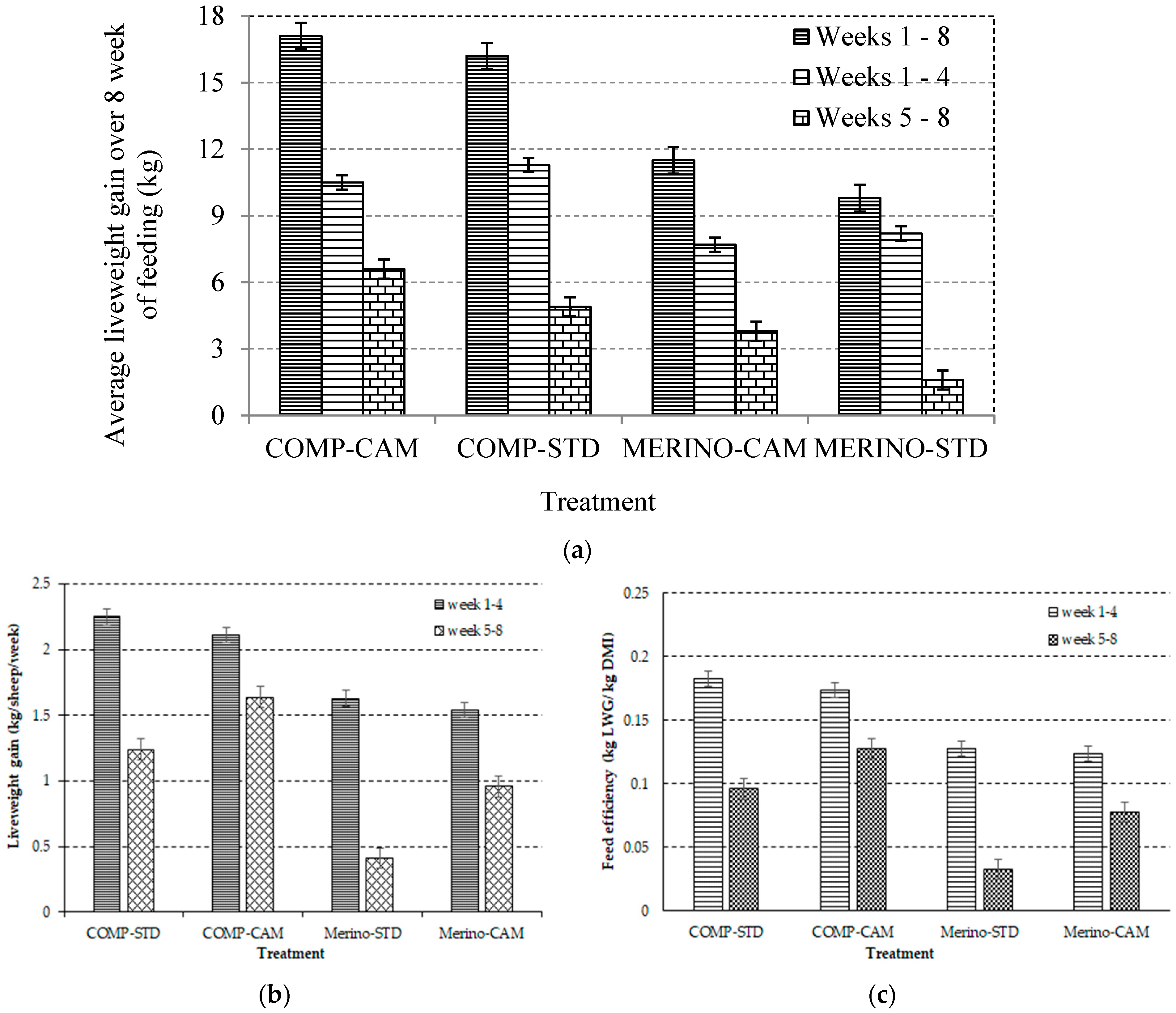
3.3. Effect of Changing Environmental Temperature and THI on Liveweight Gain and Feed Efficiency
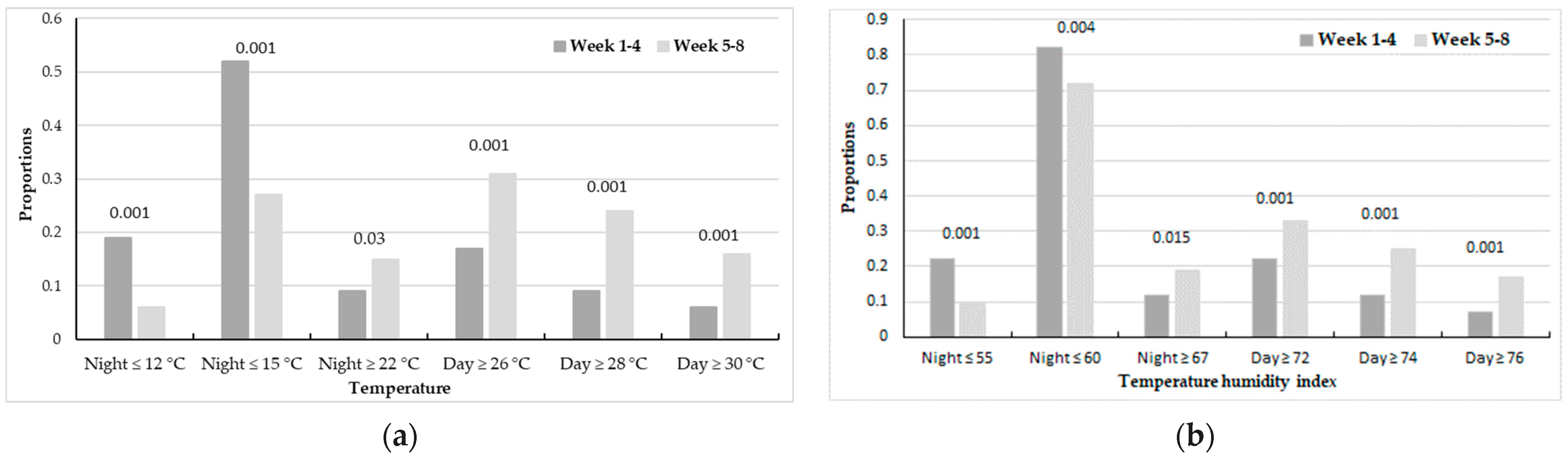
3.4. Proportions of Temperature and THI Thresholds Observed as Reference Values during Weeks 1–4 and Weeks 5–8 Periods
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Court, J.; Webb Ware, J.; Hides, S. Sheep Farming for Meat and Wool; CSIRO: Melbourne, Australia, 2010. [Google Scholar]
- Cullen, B.R.; Johnson, I.R.; Eckard, R.J.; Lodge, G.M.; Walker, R.G.; Rawnsley, R.P.; McCaskill, M.R. Climate change effects on pasture systems in south-eastern Australia. Crop Pasture Sci. 2009, 60, 933–942. [Google Scholar] [CrossRef]
- The Intergovernmental Panel on Climate Change. Climate Change 2021: The Physical Science Basis. Working Group 1 Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. Available online: https://www.ipcc.ch/report/ar6/wg1/ (accessed on 8 August 2021).
- Hahn, G.L.; Mader, T.; Eigenberg, R.A. Perspective on development of thermal indices for animal studies and management. EAAP Tech. Ser. 2003, 7, 31–44. [Google Scholar]
- Roenfeldt, S. You can’t afford to ignore heat stress. Dairy Herd Manag. 1998, 35, 6–12. [Google Scholar]
- Mishkind, M.; Vermeer, J.E.; Darwish, E.; Munnik, T. Heat stress activates phospholipase D and triggers PIP accumulation at the plasma membrane and nucleus. Plant J. Cell Mol. Biol. 2009, 60, 10–21. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Sinclair, A.J.; Holman, B.W.B. The Sources, Synthesis and Biological Actions of Omega-3 and Omega-6 Fatty Acids in Red Meat: An Overview. Foods 2021, 10, 1358. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Dunshea, F.R.; Plozza, T.E.; Hopkins, D.L.; Ponnampalam, E.N. The Impact of Antioxidant Supplementation and Heat Stress on Carcass Characteristics, Muscle Nutritional Profile and Functionality of Lamb Meat. Animals 2020, 10, 1286. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Celi, P.; Leury, B.J.; Dunshea, F.R. High dietary selenium and vitamin E supplementation ameliorates the impacts of heat load on oxidative status and acid-base balance in sheep. J. Anim. Sci. 2015, 93, 3342–3354. [Google Scholar] [CrossRef]
- Dale, N.; Fuller, H. Effect of diet composition on feed intake and growth of chicks under heat stress: II. Constant vs. cycling temperatures. Poult. Sci. 1980, 59, 1434–1441. [Google Scholar] [CrossRef]
- Coffey, M.; Seerley, R.; Funderburke, D.; McCampbell, H. Effect of heat increment and level of dietary energy and environmental temperature on the performance of growing-finishing swine. J. Anim. Sci. 1982, 54, 95–105. [Google Scholar] [CrossRef]
- Ilian, M.; Razzaque, M.; Salman, A. Use of fat in diets of sheep in hot environments. II. Effect on rumen metabolism. Anim. Feed. Sci. Technol. 1988, 19, 343–350. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmed, Z.I.; Ahmad, R.; Ashraf, M.Y.; Saifullah; Naeem, M.; Rengel, Z. ‘Camelina sativa’, a climate proof crop, has high nutritive value and multiple-uses: A review. Aust. J. Crop Sci. 2013, 7, 1551–1559. [Google Scholar]
- Obour, A.; Sintim, H.; Obeng, E.; Jeliazkov, D. Oilseed camelina (Camelina sativa L Crantz): Production systems, prospects and challenges in the USA Great Plains. Adv. Plants Agric. Res. 2015, 2, 43. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Kerr, M.G.; Butler, K.L.; Cottrell, J.J.; Dunshea, F.R.; Jacobs, J.L. Filling the out of season gaps for lamb and hogget production: Diet and genetic influence on carcass yield, carcass composition and retail value of meat. Meat Sci. 2019, 148, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Knight, M.I.; Moate, P.J.; Jacobs, J.L. An alternative approach for sustainable sheep meat production: Implications for food security. J. Anim. Sci. Biotechnol. 2020, 11, 83. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australia, 2013. [Google Scholar]
- Brash, L.D.; Fogarty, N.; Gilmour, A.R. Genetic parameters for Australian maternal and dual-purpose meatsheep breeds. II. Liveweight, wool and reproduction in Corriedale sheep. Aust. J. Agric. Res. 1994, 45, 469–480. [Google Scholar] [CrossRef]
- Thom, E.C. The Discomfort Index. Weatherwise 1959, 12, 57–61. [Google Scholar] [CrossRef]
- Roberts, C.A.; Workman, J., Jr.; Reeves, J.B., III. Near-Infrared Spectroscopy in Agriculture; American Society of Agronomy: Madison, WI, USA, 2004. [Google Scholar]
- Freer, M.; Dove, H.; Nolan, J.V. Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Melbourne, Australia, 2007. [Google Scholar]
- VSN International. Genstat Reference Manual (Release 18), Part 1 Summary; VSN International: Hemel Hempstead, UK, 2015. [Google Scholar]
- Al-Dawood, A. Towards Heat Stress Management in Small Ruminants—A Review. Ann. Anim. Sci. 2016, 17, 59–88. [Google Scholar] [CrossRef]
- Henry, B.K.; Eckard, R.J.; Beauchemin, K.A. Review: Adaptation of ruminant livestock production systems to climate changes. Animal 2018, 12, s445–s456. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Ponnampalam, E.N.; Celi, P.; Hopkins, D.L.; Leury, B.J.; Dunshea, F.R. High dietary vitamin E and selenium improves feed intake and weight gain of finisher lambs and maintains redox homeostasis under hot conditions. Small Rumin. Res. 2016, 137, 17–23. [Google Scholar] [CrossRef]
- Kadzere, C.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Sci. 2013, 77, 59–91. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Bonner, S.; Loxton, I.; Mader, T.L.; Lisle, A.; Lawrence, R. Effect of shade on body temperature and performance of feedlot steers. J. Anim. Sci. 2010, 88, 4056–4067. [Google Scholar] [CrossRef] [PubMed]
- Pardo, G.; del Prado, A. Guidelines for small ruminant production systems under climate emergency in Europe. Small Rumin. Res. 2020, 193, 106261. [Google Scholar] [CrossRef]
- Bernabucci, U.; Lacetera, N.; Danieli, P.P.; Bani, P.; Nardone, A.; Ronchi, B. Influence of different periods of exposure to hot environment on rumen function and diet digestibility in sheep. Int. J. Biometeorol. 2009, 53, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.M.; Thomas, R.; Holmes, J.H.G. Interactions between heat stress and nutrition in sheep fed roughage diets. J. Agric. Sci. 1999, 132, 351–359. [Google Scholar] [CrossRef]
- Mahjoubi, E.; Amanlou, H.; Mirzaei-Alamouti, H.R.; Aghaziarati, N.; Yazdi, M.H.; Noori, G.R.; Yuan, K.; Baumgard, L.H. The effect of cyclical and mild heat stress on productivity and metabolism in Afshari lambs. J. Anim. Sci. 2014, 92, 1007–1014. [Google Scholar] [CrossRef]
- Igono, M.O.; Bjotvedt, G.; Sanford-Crane, H.T. Environmental profile and critical temperature effects on milk production of Holstein cows in desert climate. Int. J. Biometeorol. 1992, 36, 77–87. [Google Scholar] [CrossRef]
- Muller, C.J.C.; Botha, J.A.; Smith, W.A. Effect of shade on various parameters of Friesian cows in a Mediterranean climate in South Africa. 1. Feed and water intake, milk production and milk composition. S. Afr. J. Anim. Sci. 1994, 24, 49–55. [Google Scholar]
- Gauly, M.; Bollwein, H.; Breves, G.; Brügemann, K.; Dänicke, S.; Daş, G.; Demeler, J.; Hansen, H.; Isselstein, J.; König, S.; et al. Future consequences and challenges for dairy cow production systems arising from climate change in Central Europe—A review. Animal 2013, 7, 843–859. [Google Scholar] [CrossRef]
- Smith, D.L.; Smith, T.; Rude, B.J.; Ward, S.H. Short communication: Comparison of the effects of heat stress on milk and component yields and somatic cell score in Holstein and Jersey cows. J. Dairy Sci. 2013, 96, 3028–3033. [Google Scholar] [CrossRef]
- West, J.W. Effects of heat-stress on production in dairy cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- Yazdi, M.H.; Mirzaei-Alamouti, H.R.; Amanlou, H.; Mahjoubi, E.; Nabipour, A.; Aghaziarati, N.; Baumgard, L.H. Effects of heat stress on metabolism, digestibility, and rumen epithelial characteristics in growing Holstein calves. J. Anim. Sci. 2016, 94, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Mahjoubi, E.; Yazdi, M.H.; Aghaziarati, N.; Noori, G.R.; Afsarian, O.; Baumgard, L.H. The effect of cyclical and severe heat stress on growth performance and metabolism in Afshari lambs. J. Anim. Sci. 2015, 93, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Cao, Y.; Zhou, D.W. Effects of shade on welfare and meat quality of grazing sheep under high ambient temperature. J. Anim. Sci. 2012, 90, 4764–4770. [Google Scholar] [CrossRef] [PubMed]
- Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod. Sci. 2000, 67, 1–18. [Google Scholar] [CrossRef]
- Walsberg, G. Coat Color and Solar Heat Gain in Animals. Bioscience 1983, 33, 88–91. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Warner, R.D.; Dunshea, F.R. Basal and hormone-stimulated metabolism in lambs varies with breed and diet quality. Domest. Anim. Endocrinol. 2012, 42, 94–102. [Google Scholar] [CrossRef]
- Williams, S.R.O.; Milner, T.C.; Garner, J.B.; Moate, P.J.; Jacobs, J.L.; Hannah, M.C.; Wales, W.J.; Marett, L.C. Dietary fat and betaine supplements offered to lactating cows affect dry matter intake, milk production and body temperature responses to an acute heat challenge. Animals 2021, 11, 3110. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Dietary Ingredients Used (% w/w) | |
|---|---|---|
| CAM Forage (CAM) | Standard Forage (STD) | |
| Lupins | 30 | 30 |
| Barley grain | 10 | 20 |
| Oat grain | 15 | 5 |
| Oaten hay | 0 | 45 |
| Camelina-Oat-Barley hay, 33:33:33 w/w/w | 45 | 0 |
| Nutritive Characteristics of Diet 1 | ||
|---|---|---|
| CAM | STD | |
| Dry matter, g/100 g DM | 88.85 | 89.35 |
| Crude protein, % DM | 15.20 | 14.80 |
| Metabolisable energy, MJ/kg DM | 10.83 | 10.80 |
| Crude fat, % DM | 2.91 | 2.11 |
| Acid detergent fibre, % DM | 19.93 | 19.03 |
| Neutral detergent fibre, % DM | 34.03 | 34.23 |
| Lignin, % DM | 4.20 | 4.30 |
| Phosphorus, % DM | 0.42 | 0.47 |
| Potassium, % DM | 1.42 | 1.47 |
| Sulphur, % DM | 0.21 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponnampalam, E.N.; McCaskill, M.; Giri, K.; Muir, S.K.; Cameron, F.; Jacobs, J.L.; Knight, M.I. Temperature and Temperature Humidity Index Changes during Summer to Autumn in a Temperate Zone May Affect Liveweight Gain and Feed Efficiency in Sheep. Ruminants 2022, 2, 390-406. https://doi.org/10.3390/ruminants2040027
Ponnampalam EN, McCaskill M, Giri K, Muir SK, Cameron F, Jacobs JL, Knight MI. Temperature and Temperature Humidity Index Changes during Summer to Autumn in a Temperate Zone May Affect Liveweight Gain and Feed Efficiency in Sheep. Ruminants. 2022; 2(4):390-406. https://doi.org/10.3390/ruminants2040027
Chicago/Turabian StylePonnampalam, Eric N., Malcolm McCaskill, Khageswor Giri, Stephanie K. Muir, Fiona Cameron, Joe L. Jacobs, and Matthew I. Knight. 2022. "Temperature and Temperature Humidity Index Changes during Summer to Autumn in a Temperate Zone May Affect Liveweight Gain and Feed Efficiency in Sheep" Ruminants 2, no. 4: 390-406. https://doi.org/10.3390/ruminants2040027
APA StylePonnampalam, E. N., McCaskill, M., Giri, K., Muir, S. K., Cameron, F., Jacobs, J. L., & Knight, M. I. (2022). Temperature and Temperature Humidity Index Changes during Summer to Autumn in a Temperate Zone May Affect Liveweight Gain and Feed Efficiency in Sheep. Ruminants, 2(4), 390-406. https://doi.org/10.3390/ruminants2040027