

The Evolution of Human Social Behavior


{kind=link}
Abstract
1. Introduction
2. The Role of Feelings
3. Affiliations
3.1. Parent–Child
3.2. Pair-Bonding

3.3. Kinship
3.4. Social Life
- Inclusive fitness is primarily about relatives, but the associated feelings can “spill over” to non-kin.
- Reciprocity means that I help you today because I expect that you will help me later [43]. The principle stands strong in humans, where it forms the basis for commitment and obligations, but it is relevant for other species as well. The reciprocity can be indirect in that the return is given by a third party. Our concern about reputation is related to indirect reciprocity.
- Group selection is somewhat controversial [44]. It implies that natural selection can act not only on the individual but on a tribe or a troop. The idea is that the stronger groups outperform competing groups. The benefit will likely affect the genes of an individual because if the group thrives, his or her progeny will eventually benefit. I believe group selection was instrumental in establishing the foundation of human social life and perhaps the use of religion as a tool to promote prosocial behavior [6,45].
3.5. Hostility
4. Present Society
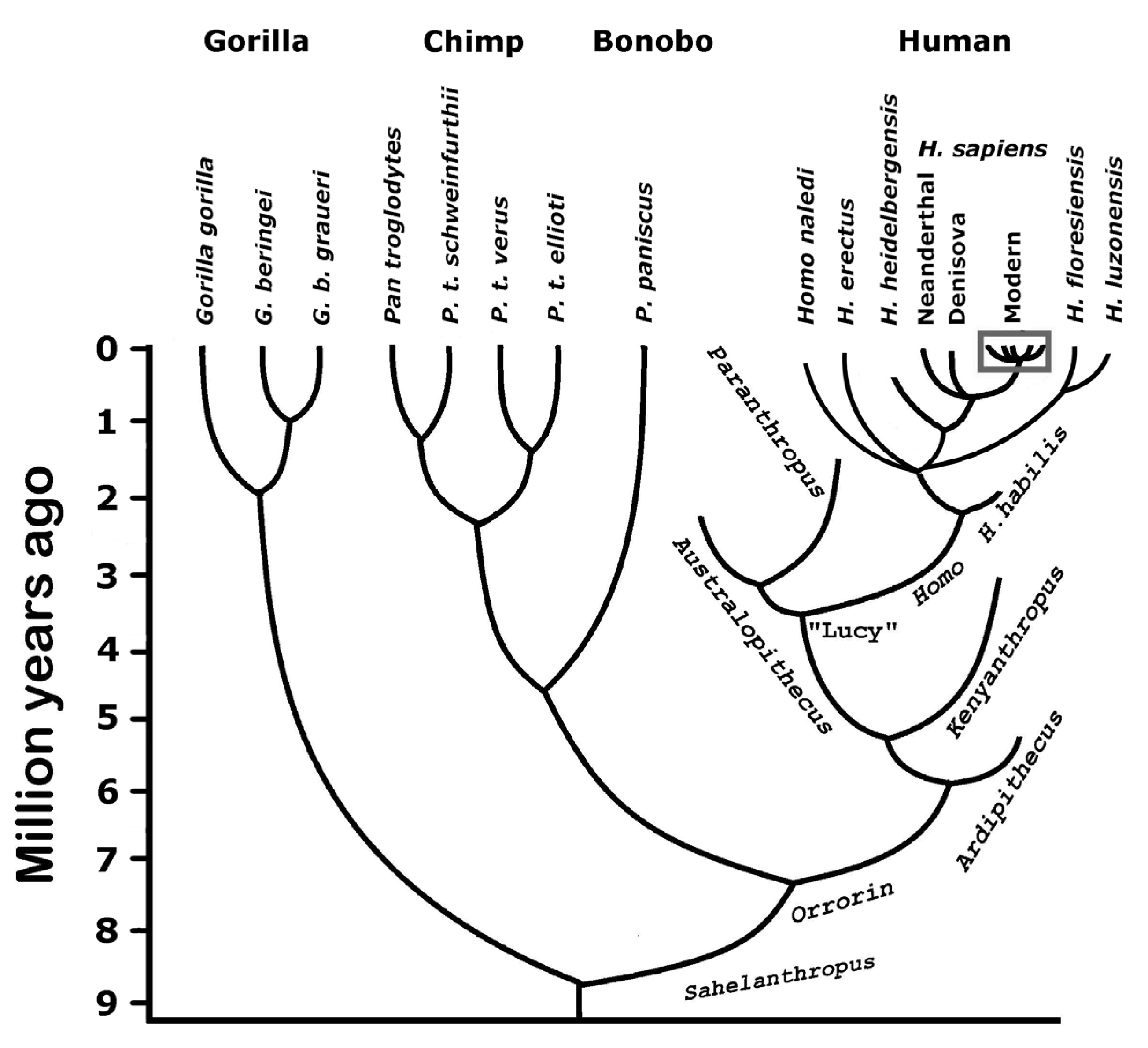
4.1. The History of Homo Sapiens
4.2. Social Discords
- A high population density. The “us and them” inclination is troublesome in a world where conflicts between groups are common.
- Present societies require that you interact with many people you do not know. We have an innate tendency to fear strangers [73]; thus, the situation is expected to increase not only hostility but also stress and anxiety.
- The large number of people, combined with how society is organized, means more competition and concomitant stress.
- We have lost the close-knit social structure of the tribe, and the consequences are loneliness, insecurity, and a lack of belonging. Family and friends do not offer complete compensation for the tribal bonds.
- While the typical tribe presumably had a relatively flat social structure, in most countries there is a distinct hierarchy. The inhabitants frequently encounter dominant individuals such as teachers, police, or government officials.
- We can no longer fulfill our needs solely by personal activity but rely on external suppliers.
5. Conclusions and Prospects
5.1. Promoting Social Behavior
5.2. Aiming for Happiness
5.3. Finale Remarks
Funding
Conflicts of Interest
References
- Dobzhansky, T. Nothing in Biology Makes Sense except in the Light of Evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Wilson, E.O. Sociobiology: The New Synthesis; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Grüter, C.; Czaczkes, T.J. Communication in Social Insects and How It Is Shaped by Individual Experience. Anim. Behav. 2019, 151, 207–215. [Google Scholar] [CrossRef]
- Dugatkin, L.A. Cheating Monkeys and Citizen Bees: The Nature of Cooperation in Animals and Humans; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Grinde, B. An Evolutionary Perspective on the Importance of Community Relations for Quality of Life. Sci. World J. 2009, 9, 588–605. [Google Scholar] [CrossRef] [PubMed]
- Atkins, P.W.; Wilson, D.S.; Hayes, S.C. Prosocial: Using Evolutionary Science to Build Productive, Equitable, and Collaborative Groups; New Harbinger Publications: New York, NY, USA, 2019. [Google Scholar]
- Grinde, B. Did Consciousness First Evolve in the Amniotes? Psychol. Conscious. 2018, 5, 239–257. [Google Scholar] [CrossRef]
- Grinde, B. Consciousness: A Strategy for Behavioral Decisions. Encyclopedia 2023, 3, 60–76. [Google Scholar] [CrossRef]
- Grinde, B. The Biology of Happiness; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Helliwell, J.F.; Layard, R.; Sachs, J.D.; Neve, J.-E.D.; Aknin, L.B.; Wang, S. World Happiness Report 2023; Sustainable Development Solutions Network: New York, NY, USA, 2023. [Google Scholar]
- Abdallah, S.; Thompson, S.; Michaelson, J.; Marks, N.; Steuer, N. The Happy Planet Index 2.0; New Economic Foundation: London, UK, 2009. [Google Scholar]
- Bertolero, M.A.; Yeo, B.T.T.; Bassett, D.S.; D’Esposito, M. A Mechanistic Model of Connector Hubs, Modularity and Cognition. Nat. Hum. Behav. 2018, 2, 765–777. [Google Scholar] [CrossRef]
- Philipson, L. Functional Modules of the Brain. J. Theor. Biol. 2002, 215, 109–119. [Google Scholar] [CrossRef]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Leknes, S.; Tracey, I. A Common Neurobiology for Pain and Pleasure. Nat. Rev. Neurosci. 2008, 9, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Kringelbach, M.L. Neuroscience of affect: Brain Mechanisms of Pleasure and Displeasure. Curr. Opin. Neurobiol. 2013, 23, 294–303. [Google Scholar] [CrossRef]
- Dalgleish, T. The Emotional Brain. Nat. Rev. Neurosci. 2004, 5, 583–589. [Google Scholar] [CrossRef]
- Balcombe, J. Animal Pleasure and Its Moral Significance. Appl. Anim. Behav. Sci. 2009, 118, 208–216. [Google Scholar] [CrossRef]
- Spence-Aizenberg, A.; Di Fiore, A.; Fernandez-Duque, E. Social Monogamy, Male–Female Relationships, and Biparental Care in Wild Titi Monkeys (Callicebus discolor). Primates 2016, 57, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Royle, N.J.; Smiseth, P.T.; Kölliker, M. The Evolution of Parental Care; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Flannery, T.F.; Rich, T.H.; Vickers-Rich, P.; Ziegler, T.; Veatch, E.G.; Helgen, K.M. A Review of Monotreme (Monotremata) Evolution. Alcheringa 2022, 46, 3–20. [Google Scholar] [CrossRef]
- Shih, H.-C.; Kuo, M.-E.; Wu, C.W.; Chao, Y.-P.; Huang, H.-W.; Huang, C.-M. The Neurobiological Basis of Love: A Meta-Analysis of Human Functional Neuroimaging Studies of Maternal and Passionate Love. Brain Sci. 2022, 12, 830. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.; Kushnick, G. Proximate and Ultimate Perspectives on Romantic Love. Front. Psychol. 2021, 12, 573123. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, S.A.; Young, L.J. The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives. Biology 2023, 12, 844. [Google Scholar] [CrossRef]
- Kleiman, D.G. Monogamy in Mammals. Q. Rev. Biol. 1977, 52, 39–69. [Google Scholar] [CrossRef]
- Lack, D. Ecological Adaptations for Breeding in Birds; Methuen and Company: London, UK, 1968. [Google Scholar]
- Lukas, D.; Clutton-Brock, T.H. The Evolution of Social Monogamy in Mammals. Science 2013, 341, 526–530. [Google Scholar] [CrossRef]
- Ford, C.S.; Beach, F.A. Patterns of Sexual Behavior; Harper: New York, NY, USA, 1951. [Google Scholar]
- Yong, J.C.; Li, N.P. Elucidating Evolutionary Principles with the Traditional Mosuo: Adaptive Benefits and Origins of Matriliny and “Walking Marriages”. Cult. Evol. 2022, 19, 22–40. [Google Scholar] [CrossRef]
- Strier, K.B. Primate Behavioral Ecology; Routledge: New York, NY, USA, 2016. [Google Scholar]
- Hasty, J.; Lewis, D.G.; Snipes, M.M. Introduction to Anthropology; OpenStax Rice University: Houston, TX, USA, 2022. [Google Scholar]
- Feldman, R. The Neurobiology of Human Attachments. Trends Cogn. Sci. 2017, 21, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Marazziti, D.; Akiskal, H.S.; Rossi, A.; Cassano, G.B. Alteration of the Platelet Serotonin Transporter in Romantic Love. Psychol. Med. 1999, 29, 741–745. [Google Scholar] [CrossRef]
- Pitkow, L.J.; Sharer, C.A.; Ren, X.; Insel, T.R.; Terwilliger, E.F.; Young, L.J. Facilitation of Affiliation and Pair-Bond Formation by Vasopressin Receptor Gene Transfer into the Ventral Forebrain of a Monogamous Vole. J. Neurosci. 2001, 21, 7392–7396. [Google Scholar] [CrossRef]
- Wood, B.; Boyle, E.K. Hominin Taxic Diversity: Fact or Fantasy? Am. J. Phys. Anthropol. 2016, 159, 37–78. [Google Scholar] [CrossRef]
- Levin, S.R.; Grafen, A. Inclusive Fitness Is an Indispensable Approximation for Understanding Organismal Design. Evolution 2019, 73, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.S.; De Moor, D.; Casanova, C.; Brent, L.J. Kinship Composition in Mammals. R. Soc. Open Sci. 2023, 10, 230486. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.T.; Steadman, L.B. Human Kinship as a Descendant-Leaving Strategy: A Solution to an Evolutionary Puzzle. J. Soc. Evol. Syst. 1997, 20, 39–51. [Google Scholar] [CrossRef]
- Fortunato, L. Lineal Kinship Organization in Cross-Specific Perspective. Philos. Trans. R. Soc. B 2019, 374, 20190005. [Google Scholar] [CrossRef] [PubMed]
- Skov, L.; Peyrégne, S.; Popli, D.; Iasi, L.N.; Devièse, T.; Slon, V.; Zavala, E.I.; Hajdinjak, M.; Sümer, A.P.; Grote, S. Genetic Insights into the Social Organization of Neanderthals. Nature 2022, 610, 519–525. [Google Scholar] [CrossRef]
- Lieberman, D.; Lobel, T. Kinship on the Kibbutz: Coresidence Duration Predicts Altruism, Personal Sexual Aversions and Moral Attitudes among Communally Reared Peers. Evol. Hum. Behav. 2012, 33, 26–34. [Google Scholar] [CrossRef]
- Krause, J.; Ruxton, G.D. Living in Groups; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Romano, A.; Saral, A.S.; Wu, J. Direct and Indirect Reciprocity among Individuals and Groups. Curr. Opin. Psychol. 2022, 43, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Smith, D. Cultural Group Selection and Human Cooperation: A Conceptual and Empirical Review. Evol. Hum. Sci. 2020, 2, e2. [Google Scholar] [CrossRef]
- Grinde, B. How Can Science Help Religion toward Optimal Benefit for Society? Zygon 2005, 40, 277–288. [Google Scholar] [CrossRef]
- Hensch, T.K. Critical Period Regulation. Annu. Rev. Neurosci. 2004, 27, 549–579. [Google Scholar] [CrossRef] [PubMed]
- Berk, L. Child Development; Pearson Higher Education: Sidney, OH, USA, 2015. [Google Scholar]
- Dombrowski, S.C.; Gischlar, K.L.; Mrazik, M.; Greer, F.W. Feral Children. In Assessing and Treating Low Incidence/High Severity Psychological Disorders of Childhood; Springer: New York, NY, USA, 2011; pp. 81–93. [Google Scholar]
- de Waal, F. Bonobo Sex and Society. Sci. Am. 2016, 25, 42–49. [Google Scholar]
- Poiani, A. Animal Homosexuality: A Biosocial Perspective; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Barron, A.B.; Hare, B. Prosociality and a Sociosexual Hypothesis for the Evolution of Same-Sex Attraction in Humans. Front. Psychol. 2020, 10, 2955. [Google Scholar] [CrossRef] [PubMed]
- Pfau, D.; Jordan, C.L.; Breedlove, S.M. The De-Scent of Sexuality: Did Loss of a Pheromone Signaling Protein Permit the Evolution of Same-Sex Sexual Behavior in Primates? Arch. Sex. Behav. 2021, 50, 2267–2276. [Google Scholar] [CrossRef] [PubMed]
- Stowers, L.; Holy, T.E.; Meister, M.; Dulac, C.; Koentges, G. Loss of Sex Discrimination and Male-Male Aggression in Mice Deficient for Trp2. Science 2002, 295, 1493–1500. [Google Scholar] [CrossRef]
- Hill, R.A.; Dunbar, R.I. Social Network Size in Humans. Hum. Nat. 2003, 14, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Peter, T. Population Dynamics and Internal Warfare: A Reconsideration. Soc. Evol. Hist. 2006, 5, 112–147. [Google Scholar]
- Rankin, D.J. Resolving the Tragedy of the Commons: The Feedback between Intraspecific Conflict and Population Density. J. Evol. Biol. 2007, 20, 173–180. [Google Scholar] [CrossRef]
- Pinker, S. The Better Angels of Our Nature; Penguin: London, UK, 2011. [Google Scholar]
- Prüfer, K.; De Filippo, C.; Grote, S.; Mafessoni, F.; Korlević, P.; Hajdinjak, M.; Vernot, B.; Skov, L.; Hsieh, P.; Peyrégne, S. A High-Coverage Neandertal Genome from Vindija Cave in Croatia. Science 2017, 358, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, N.K.; Shapiro, B.; Green, R.E. An Ancestral Recombination Graph of Human, Neanderthal, and Denisovan Genomes. Sci. Adv. 2021, 7, eabc0776. [Google Scholar] [CrossRef]
- Macciardi, F.; Martini, F. The Neanderthal Brain: Biological and Cognitive Evolution. In Updating Neanderthals; Romangnoli, F., Rivals, F., Benazzi, S., Eds.; Elsevier: New York, NY, USA, 2022; pp. 89–108. [Google Scholar]
- García-Diez, M. ‘Art’: Neanderthal Symbolic Graphic Behaviour. In Updating Neanderthals; Romangnoli, F., Rivals, F., Benazzi, S., Eds.; Elsevier: New York, NY, USA, 2022; pp. 251–260. [Google Scholar]
- Benítez-Burraco, A. The Case of Neanderthal Language (s): A Multidisciplinary Approach. PsyArXiv 2023. [Google Scholar] [CrossRef]
- Bergström, A.; McCarthy, S.A.; Hui, R.; Almarri, M.A.; Ayub, Q.; Danecek, P.; Chen, Y.; Felkel, S.; Hallast, P.; Kamm, J. Insights into Human Genetic Variation and Population History from 929 Diverse Genomes. Science 2020, 367, eaay5012. [Google Scholar] [CrossRef] [PubMed]
- Snir, A.; Nadel, D.; Groman-Yaroslavski, I.; Melamed, Y.; Sternberg, M.; Bar-Yosef, O.; Weiss, E. The Origin of Cultivation and Proto-Weeds, Long before Neolithic Farming. PLoS ONE 2015, 10, e0131422. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.; Evans, J.; Lamb, A.; Baird, D.; Hodder, I.; Marciniak, A.; Larsen, C.S.; Knüsel, C.J.; Haddow, S.D.; Pilloud, M.A. Mobility and Kinship in the World’s First Village Societies. Proc. Natl. Acad. Sci. USA 2023, 120, e2209480119. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R. Human Genetics: The Basics; Taylor & Francis: New York, NY, USA, 2016. [Google Scholar]
- Owens, K.; King, M.-C. Genomic Views of Human History. Science 1999, 286, 451–453. [Google Scholar] [CrossRef]
- Moberg, G.; Mench, J. The Biology of Animal Stress; Moberg, G., Mench, J., Eds.; CABI: Oxfordshire, UK, 2000. [Google Scholar]
- Grinde, B. Can the Concept of Discords Help Us Find the Causes of Mental Diseases? Med. Hypotheses 2009, 73, 106–109. [Google Scholar] [CrossRef]
- Eaton, S.B.; Konner, M.; Shostak, M. Stone Agers in the Fast Lane: Chronic Degenerative Diseases in Evolutionary Perspective. Am. J. Med. 1988, 84, 739–749. [Google Scholar] [CrossRef]
- Grinde, B. An Approach to the Prevention of Anxiety-Related Disorders Based on Evolutionary Medicine. Prev. Med. 2005, 40, 904–909. [Google Scholar] [CrossRef]
- Williams, G.C.; Nesse, R.M. The Dawn of Darwinian Medicine. Q. Rev. Biol. 1991, 66, 1–22. [Google Scholar] [CrossRef]
- Marks, I. The Development of Normal Fear: A Review. J. Child Psychol. Psychiatry 1987, 28, 667–697. [Google Scholar] [CrossRef]
- Moffitt, T.E.; Caspi, A.; Taylor, A.; Kokaua, J.; Milne, B.J.; Polanczyk, G.; Poulton, R. How Common Are Common Mental Disorders? Evidence That Lifetime Prevalence Rates Are Doubled by Prospective Versus Retrospective Ascertainment. Psychol. Med. 2010, 40, 899–909. [Google Scholar] [CrossRef]
- Douglas, K. Playing Fair. New Sci. 2001, 169, 38–42. [Google Scholar]
- Fowler, J.H.; Christakis, N.A. Dynamic Spread of Happiness in a Large Social Network: Longitudinal Analysis over 20 Years in the Framingham Heart Study. BMJ 2008, 337, a2338. [Google Scholar] [CrossRef]
- Nivette, A.E. Violence in Non-State Societies: A Review. Br. J. Criminol. 2011, 51, 578–598. [Google Scholar] [CrossRef]
- Hino, K.; Schneider, R.H. Planning for Crime Prevention in Japan. Built Environ. 2013, 39, 114–139. [Google Scholar] [CrossRef]
- Grinde, B. How to Form the Human Brain. PsyArXiv Prepr. 2024. [Google Scholar] [CrossRef]
- Hofmann, S.G.; Grossman, P.; Hinton, D.E. Loving-Kindness and Compassion Meditation: Potential for Psychological Interventions. Clin. Psychol. Rev. 2011, 31, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Krasnegor, N.A.; Bridges, R.S. Mammalian Parenting; Oxford University Press: Oxford, UK, 1990. [Google Scholar]
- Clark, R. The Japanese Company; Tuttle Publishing: Tokyo, Japan, 2013. [Google Scholar]
- Berry, Z.; Frederickson, J. Explanations and Implications of the Fundamental Attribution Error: A Review and Proposal. J. Integr. Soc. Sci. 2015, 5, 44–57. [Google Scholar]
- DiMaria, C.H.; Peroni, C.; Sarracino, F. Happiness Matters: Productivity Gains from Subjective Well-Being. J. Happiness Stud. 2020, 21, 139–160. [Google Scholar] [CrossRef]
- Grinde, B. Social Behavior: Making the Best of the Human Condition. Mank. Q. 2000, 41, 193–210. [Google Scholar] [CrossRef]
- Layard, R. Happiness—Lessons from a New Science; Penguin: London, UK, 2005. [Google Scholar]
- Grinde, B.; Nes, R.B.; MacDonald, I.; Wilson, D.S. Quality of Life in Intentional Communities. Soc. Indic. Res. 2017, 137, 625–640. [Google Scholar] [CrossRef]
- Gallagher, E.N.; Vella-Brodrick, D.A. Social Support and Emotional Intelligence as Predictors of Subjective Well-Being. Personal. Individ. Differ. 2008, 44, 1551–1561. [Google Scholar] [CrossRef]
- Grinde, B. Role of Happiness When Evaluating Society. Encyclopedia 2022, 2, 230–236. [Google Scholar] [CrossRef]
- Moll, J.; Krueger, F.; Zahn, R.; Pardini, M.; de Oliveira-Souza, R.; Grafman, J. Human Fronto-Mesolimbic Networks Guide Decisions about Charitable Donation. Proc. Natl. Acad. Sci. USA 2006, 103, 15623–15628. [Google Scholar] [CrossRef] [PubMed]
- Borgonovi, F. Doing Well by Doing Good. The Relationship between Formal Volunteering and Self-Reported Health and Happiness. Soc. Sci. Med. 2008, 66, 2321–2334. [Google Scholar] [CrossRef]
- Grinde, B. An Evolutionary Perspective on Free Will and Self-Consciousness. Psychol. Conscious. 2022. [Google Scholar] [CrossRef]
- Lykken, D. Happiness: The Nature and Nurture of Joy and Contentment; St. Martin’s Griffin: New York, NY, USA, 2000. [Google Scholar]
- Veenhoven, R. Well-Being in the Welfare State: Level Not Higher, Distribution Not More Equitable. J. Comp. Policy Anal. 2000, 2, 91–125. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grinde, B. The Evolution of Human Social Behavior. Encyclopedia 2024, 4, 430-443. https://doi.org/10.3390/encyclopedia4010029
Grinde B. The Evolution of Human Social Behavior. Encyclopedia. 2024; 4(1):430-443. https://doi.org/10.3390/encyclopedia4010029
Chicago/Turabian StyleGrinde, Bjørn. 2024. "The Evolution of Human Social Behavior" Encyclopedia 4, no. 1: 430-443. https://doi.org/10.3390/encyclopedia4010029
APA StyleGrinde, B. (2024). The Evolution of Human Social Behavior. Encyclopedia, 4(1), 430-443. https://doi.org/10.3390/encyclopedia4010029