Abstract
Gorgonzola is an Italian “erborinato” blue cheese from cow’s milk, bearing blue-green “parsley-like” spots due to the spread of Penicillium roqueforti mycelium. Due to its pH, water activity, and high nutrient content, as well as the environmental conditions required for its maturation, Gorgonzola constitutes an optimal ecological niche supporting the growth of both yeasts and filamentous fungi. Therefore, exploring the abundant mycobiota present in this peculiar habitat is of great interest regarding the search for new probiotic strains. The present investigation aimed to characterize the Gorgonzola mycobiota using both phenotypic (macroscopic and microscopic morphological analyses) and genotypic (DNA barcoding) analyses to find possible putative probiotic strains to be used in veterinary medicine in feed supplements. Among the different isolated filamentous fungi (Mucor and Penicillium) and yeasts (Yarrowia, Debaryomyces, Saccharomyces, and Sporobolomyces), we selected a strain of Saccharomyces cerevisiae var. boulardii. We tested its adaptation to thermal stress and its stability in feed matrices. The overall results highlight that the selected strain is stable for three months and can be considered as a possible candidate for use as a probiotic in veterinary feed supplements.
1. Introduction
Gorgonzola cheese is an Italian dairy product belonging to the class of blue cheeses and is exclusively produced in Northern Italy (Piedmont and Lombardy). It is a soft, oily cheese made from cow’s milk, with a pink-grey crust, similar in aspect and properties to the French Roquefort and the Danish Danablue bearing blue-green “parsley-like” spots due to the spread of Penicillium roqueforti mycelium [1]. There are two types of Gorgonzola characterized by different maturation times and cooking temperatures: sweet Gorgonzola, which has a higher water content and a sweeter and creamy taste with light blue spots; and spicy Gorgonzola, which has a dry texture, strong taste, and evident blue-green strips [1].
During the production of Gorgonzola, pasteurized cow milk is first inoculated with dairy Lactobacillus and Streptococcus, which activate the lactose fermentation with the release of several organic acids that constitute the optimal nutrients for filamentous fungi development [2]. Later in the production, when oxygen and temperature are low and carbon dioxide high, P. roqueforti conidia are inoculated in the clot. Their metabolic activities ensure the typical flavor and texture of blue cheese that also achieves the typical blue-green aspect after 35–40 days from the beginning of the process due to mycelial development [3,4,5,6]. Biochemical processes such as glycolysis, lipolysis, and proteolysis carried out by filamentous fungi and yeasts (but also by starter and non-starter bacteria) contribute to the final taste of this cheese [7].
Due to their intrinsic chemo-physical features (pH, water activity, high nutrient content) and the different production steps required for maturation, blue cheese provides (especially during late ripening) a rich medium allowing the growth of several yeasts and filamentous fungi, often present in the ripening rooms [8,9]. Although some fungi belonging to the genus Penicillium (in association with some lactococcal strains) can synthesize bitter peptides with a high content of hydrophobic amino acids (such as leucine, proline, and phenylalanine) [10], in general, the mycobiota confers appreciated features to the overall taste of this dairy product.
Furthermore, along with contributing to the organoleptic properties, this fungal diversity can constitute an excellent opportunity to search for new probiotic strains. Fermented food is the right place to find high quantities of beneficial microbes to be used in combination with pharmacological therapy [11]. Some filamentous fungi and yeasts like Aspergillus oryzae [12,13], Candida pintolopesii, Saccharomyces cerevisiae [14] and S. cerevisiae var. boulardii [15,16] can have probiotic properties. Among them, the best characterized one is S. cerevisiae var. boulardii, which is used in human-targeted functionalized foods like synbiotic yogurt and is obtained by adding this yeast and prebiotic inulin, resulting in stability for 28 days without syneresis events [17]. In addition, the use of probiotic fungi instead of bacteria can ensure natural resistance to the concomitant antibiotic treatments as well as fast colonization despite the presence of antibacterial molecules without the need to use resistant bacteria whose spread is questionable [18].
Moreover, extending the range of probiotics available on the market is of growing interest, especially when used to support infectious disease control both in humans [19,20] and in veterinary medicine, where pathologies caused by fungi and parasites are very frequent [21]. In the last decade, a great effort has been made to establish the potential that probiotics can have in veterinary medicine. They have been employed to treat dysbiosis [22] and described as promising antibiotic substitutes [23].
More in detail, S. cerevisiae var. boulardii has been successfully employed to treat giardiasis with a 90% reduction of the infecting charge as well as with evident lower damage of the gut mucosa, proving to be a good alternative to metronidazole [24]. The same yeast species was found to be useful in the treatment of post-antibiotic diarrhea. Dogs treated with lincomycin receiving S. cerevisiae var. boulardii supplementation showed faster recovery and better volatile fatty acids profiles [25]. Another promising aspect is the capability of S. cerevisiae var. boulardii to attenuate the effects of the mycotoxin DON (deoxynivalenol). This molecule is present at high levels (9.5 mg/kg) in forage cereals, and can trigger, at the gut level, inflammatory pathways resulting in a syndrome similar to inflammatory bowel disease (IBD). According to Oswald et al. [26], S. cerevisiae var. boulardii supplementation at a concentration of 2 × 109 colony-forming units (CFU)/kg, before supplying infected forage meals, can interfere with both the p38 MAPK and the NF-kB pathways, thus attenuating inflammatory syndromes.
To date, most mycological analyses conducted on food are just quantitative, and to the best of our knowledge, only a limited number of studies have been undertaken to combine the typing of food microbes with the discovery of potential probiotics [8,27,28,29]. Morphological evaluation of fungi growing in a pure culture supported by molecular investigations can constitute two valuable tools to shed light on the mycobiota of blue cheeses. The former is cheaper, although it requires a long time and a certain degree of experience in microscopic validation. The latter are more expensive, but allow rapid and specific identification at the species level [30].
Therefore, the present investigation aimed to identify the mycobiota of the blue cheese Gorgonzola (as well as the one present in the ripening rooms) based on both phenotypic (macroscopic and microscopic morphological analyses) and genotypic (DNA barcoding) [31] identification methods to select putative probiotic strains to be used in veterinary medicine. A second objective was the evaluation of the time-course stability of the best candidate strain for probiotic applications in a pet feed matrix.
2. Materials and Methods
2.1. Sample Collection, and Morphological Identification of Fungal Isolates
Sampling was carried out at the Biraghi S.p.A. in Cavallermaggiore (CN, Italy). Samples were collected from the Gorgonzola cheese surface using 10 different productions at the same ripening time (9 weeks) and the air of the maturation stores. The Gorgonzola cheese surface was randomly sampled through cellulose sponges fixed on a plastic handle and supplied in a sterile bag. The sponges were kept at low temperatures and transferred to the microbiological laboratory of Biraghi S.p.A. A total of 10 mL of physiological solution was then added to the sterile bags aseptically. Bags were blended in a stomacher for 30 s. Serial dilutions were prepared from each sample (from 10−2 to 10−6) and 100 μL of each dilution were inoculated in duplicate through the spread plate method in Petri dishes containing Chloramphenicol Glucose Yeast Extract (CGYE) agar or Sabouraud culture media (Oxoid Limited, Basingstoke, UK).
The environmental sampling of indoor air was performed using Surface Air System (SAS) portable equipment (VWR International Srl, Radnor, PA USA) using culture medium CYGE.
Petri dishes were then incubated at 25 ± 2 °C for 7–15 days and 125 fungal strains were isolated based on the morphological analysis. The isolated strains were classified into 47 groups (40 filamentous fungi, 7 yeasts). Except for groups composed of only one isolate, two members of each group were then inoculated on suitable culture media for micro- and macro-morphological analyses for species identification.
Isolates belonging to the genus Penicillium were inoculated on Czapek Yeast Extract Agar (CYA), 25% Glycerol Nitrate Agar (G25N), and 20% Malt Extract Agar (MEA) and incubated for 7 days at 24 °C (MEA, G25N, and CYA), then 37 °C (CYA), and 5 °C (CYA). All other isolates were inoculated on MEA and incubated at 24 °C for 7 days. The isolated strains were then identified using conventional mycological techniques [32,33,34,35,36,37,38,39].
2.2. DNA Extraction, PCR Amplification, and DNA Sequencing
To confirm the fungal identification obtained by morphological analysis, genomic DNA was extracted from one member of each group as described below.
For the DNA extraction, fungi were grown in 100 mL of liquid malt extract culture media (malt extract 20 g/L; glucose 20 g/L; peptone 2 g/L) for 7 days at 24 °C on an orbital shaker set at 150 rpm. Subsequently, fungi were filtered using sterile Whatman filter paper and the lyophilized Genomic DNA was isolated from 20 mg of lyophilized fungal tissue through the DNeasy Plant Mini Kit (Qiagen, Germantown, MD, USA) following manufacturer instructions. As a molecular marker for species identification, the ITS-rDNA region has been used with the exception of isolates belonging to the genus Penicillium. For the latter, the partial β-tubulin (BT2) region that provides a better resolution in differentiation of closely related species has been used. Specifically, for the ITS region amplification, universal primers ITS1F/ITS4 were used [40,41], while partial β-tubulin (BT2) regions have been amplified through primers Bt2a and Bt2b [42].
Amplification reactions were performed in a Biometra TRIO-Triple Powered PCR thermal cycler (Analytik Jena GmbH, Jena, Germany). In a 50 μL reaction mixture, the final concentrations or total amounts corresponded to 20 ng DNA, 1 × PCR buffer (20 mM Tris/HCl pH 8.4, 50 mM KCl), 0.4 μM of each primer, 2.5 mM MgCl2, 0.25 mM of each dNTP, 0.5 units of Taq polymerase (Promega, Madison, WI, USA). The PCR programs were set as follows: β-tubulin: 5 min at 94 °C for one cycle; 30 s at 94 °C; 45 s at 54 °C; 1 min at 72 °C for 35 cycles; and 10 min at 72 °C for one cycle. ITS region: 5 min at 94 °C for one cycle; 35 s at 94 °C; 35 s at 58 °C; 1 min at 72 °C for 40 cycles; and 5 min at 72 °C for one cycle.
PCR products were resolved on a 1.0% agarose gel and visualized by ethidium bromide staining. PCR products were purified using Wizard® SV Gel and PCR Clean-up system (Promega, Madison, WI, USA) and sequenced by Eurofins Genomic Gmbh (Ebersberg, Germany). Sequence assembly and editing were performed using SeqTrace 0.9.0 software [43].
Species identification was then achieved by comparing the ITS and β-tubulin sequences with those present in the NCBI database by BLASTN Algorithmus (BLAST-search, last accession 30 March 2023) [44] and the Westerdijk Fungal Biodiversity Institute database [45], respectively.
Sequences were deposited in GenBank under accession numbers given in Table 1.

Table 1.
Isolated fungal strains and NCBI accession number of deposited sequences.
Among the different fungal and yeast species isolated from cheese and ripening rooms, for further in-depth studies, we decided to focus on only one species based on known literature data concerning their overall probiotic potential. Being that the majority of strains belong to still controversial or not well-ascertained probiotics, we selected S. cerevisiae var. boulardii whose probiotic properties have long been assessed [15,46].
2.3. Evaluation of S. cerevisiae var. boulardii Growth Performance under Different Temperature Conditions
The strain growth at different temperatures (35 °C, 37 °C, and 40 °C) was evaluated on Sabouraud Dextrose Agar. Moreover, according to the EFSA recommendations [47] the viable cell count for feed additives was determined as follows. A cell suspension of S. cerevisiae var. boulardii in phosphate-buffered saline (PBS) with a pH of approximately 7.4 was prepared, and different dilutions were plated (100 µL) in triplicate on OGYE agar (Oxoid Limited, Hants, UK) and incubated at 35 °C. The bacterial count, as per the standards outlined in EN 15789:2022 (animal feeding stuff: methods of sampling and analysis-detection and enumeration of Saccharomyces cerevisiae used as a feed additive) [48] is required to be higher than 108 colony-forming units per gram (CFU/g). To determine the bacterial count, the following formula was applied:
CFU/g of feed matrix = (number of colonies/volume of sample plated × dilution factor) × (1/weight of the feed matrix).
2.4. S. cerevisiae var. boulardii Inoculum Preparation
Following the EFSA “Safety and efficacy of Lactobacillus acidophilus D2/CSL” (for dogs and cats), a dose of 5 × 109 CFU/kg was chosen for further experiments. Therefore, to achieve a suitable biomass, we performed a scale-up procedure starting from the pure culture of S. cerevisiae var. boulardii strain by growing the yeast in a rich liquid medium (yeast-peptone-dextrose broth) at 35 °C for 48 h. Yeast cells were recovered by centrifugation and all pellets were collected and pooled until a total biomass of 20 g dry weight (biomass level ensuring replicates) was obtained.
2.5. Time Course Evaluation of S. cerevisiae var. boulardii Growth Performances in Feed
The study utilized off-the-shelf dry food from the ‘Natural Superpremium’ line of Monge, a renowned Italian pet food brand based in Monasterolo di Savigliano, Italy. All ingredients included in this food are listed in Table 2.
Table 2.
Chemical and biochemical composition of feed matrix.
The inoculum of S. cerevisiae var. boulardi was prepared as follows. A total of 2 g of potential probiotics were added to 18 g of milled feed, and the resulting mixture was stirred for 5 min. Subsequently, 80 g of pulverized croquettes were added to the mixture and shaken for 5 min at 150 rpm. Similarly, another 100 g of milled feed was incorporated into the mixture and further stirred. The same procedure was repeated until obtaining a final mixture weighing a total of 500 g, consisting of 498 g of feed and 2 g of additive.
Following the standard EN 15789:2022 [48], 450 mL of PBS was added to 50 g of the feed matrix fortified with S. cerevisiae var. boulardii. The mixture was homogenized using a Stomacher for 1 min on speed 3 and then allowed to stand for 30 min, which allowed for the feed to absorb the liquid. Then, the Stomacher was reactivated for a further 2 min at speed 3, and the pH of the mixture was measured and corrected to a value ranging between 7.3–8.1. Finally, the Stomacher was reactivated for 1 min at speed 3. Serial dilution, starting from 10−1, was prepared in peptone salt solution (PSS) (enzymatic digest casein 1.0 g, NaCl 8.5 g, deionized water 1000 mL). The dilutions 10−6, 10−7, and 10−8 were then inoculated (100 mcL) on OGYE agar using L-shaped glass and incubated at 35 °C. According to the standard EN 15789:2022 [48], the entire procedure described above was repeated twice at week 2, as well as weeks 8 and 13 on 5 samples of 50 g of feed added to S. cerevisiae var. boulardii.
2.6. Statistical Analysis
To evaluate the presence of significant differences in the survival rate of S. cerevisiae var. boulardii in the feed mixture, the number of viable cells counted on OGYE agar was compared on 5 samples for each considered time (T0, T2, T4, T8, T13) by one-way analysis of variance (ANOVA) followed by multiple tests of Turkey comparison (* p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001). Statistical tests were carried out using Past 4 statistical analysis software [49].
3. Results
3.1. Isolation of Different Fungal and Yeast Strains from Gorgonzola Cheese and Ripening Stores
A total of 126 fungal strains were isolated from Gorgonzola cheese (Figure 1) or the air of maturing stores (Table 1). The identification at the species level was possible for 15 out of the 17 retrieved species. In two cases, despite several attempts, the obtained DNA was not suitable for PCR amplification. The 126 fungal isolates belong to three different Phyla: Ascomycota (8 genera, 13 species), Basidiomycota (1 genus, 1 species), and Mucoromycota (1 genus, 4 species) (Table 1). Regarding the Phylum Ascomycota, Penicillium was the most frequent and abundant genus (68 isolates, 5 species) followed by Debaryomyces (30 isolates, 1 species), Yarrowia (5 isolates, 2 species), Cladosporium (2 isolates, 1 species), Fusicolla (2 isolates, 1 species) Cordyceps (2 isolates, 1 species), Aspergillus (1 isolate, 1 species), and S. cerevisiae var. boulardii (1 isolate, 1 species). Regarding the Phylum Basidiomycota, the only species belonging to this Phylum was Sporobolomyces ruberrimus (3 isolates), while for the Phylum Mucoromycota we isolated 12 strains belonging to 4 different species of the genus Mucor (M. fuscus 7 isolates, M. circinelloides 1 isolate, M. lanceolatus 3 isolates, Mucor sp. 1 isolate) (Table 2). D. hansenii, M. fuscus, M. lanceolatus, P. commune and P. roqueforti were isolated from both Gorgonzola cheese and from the air of maturing stores. M. circinelloides, Mucor sp., Penicillium sp. and Y. lipolytica, Y. deformans, S. ruberrimus, and S. cerevisiae var. boulardi were isolated only from Gorgonzola cheese, whereas A. flavus, C. cladosporioides, C. farinosa, F. aquaeductuum, P. atrosanguineum, P. camemberti, were isolated from only the air of maturing stores (Table 1).

Figure 1.
Yeast and filamentous fungi isolated from Gorgonzola cheese (Biraghi S.p.a.) and grown in CYGE or Sabouraud agar. Total of 126 fungal strains belonging to phylum of Ascomycota (8 genera, 13 species), Basidiomycota (1 genus, 1 species), and Mucoromycota (1 genus, 4 species) were isolated.
3.2. Studies on the Putative Probiotic S. cerevisiae var. boulardii
Among the several isolates, five different yeast species were detected. These can be potentially included in the category of putative probiotics. However, despite several beneficial traits described for some of them, only S. cerevisiae var. boulardii has received the GRAS status without constraints (see Section 4). Therefore, in the following experiments, we focused our attention on this species.
3.2.1. Growth at Different Temperatures
Strain viability during thermal stress was tested by growing this strain at 35 °C, 37 °C, and 40 °C in Sabouraud Dextrose Agar. The temperatures of 35 °C and 37 °C supported good growth, whereas 40 °C did not allow for any growth to occur.
3.2.2. Cell Viability at 35 °C in Standard Conditions
To test the strain viability in standard conditions (before adding it to the veterinary feed matrix), a temperature of 35 °C was chosen and OGYE agar was used as a preferred medium as it guarantees good repeatability and reproducibility of the results [50]. This procedure allows for the evaluation of survival of the potential probiotic strain during shelf life in summer conditions. This approach aligns with the standards set by EFSA, which encourage conducting studies under the recommended storage conditions to provide a realistic estimate of the shelf life of each additive formulation [47]. Viable colonies were detected after 24 h growth and appear to be irregularly shaped and creamy, with a medium diameter of 1–6 mm.
3.2.3. Time-Course Stability of S. cerevisiae var. boulardii in the Feed Matrix
To establish the shelf-life of the selected S. cerevisiae var. boulardii strain in a complex feed mixture, the viable microbial population was evaluated at 35 °C during three months in a matrix whose composition is referred to in Section 2. The results show at least 108 viable cells for the sample (Table 3 and Figure 2), showing a maintained probiotic efficacy [51,52].
Table 3.
Colony-forming units (CFU/g) of S. cereviasae var. boulardii in a feed mixture during three months of monitoring were measured as viable cell count on OGYE agar.
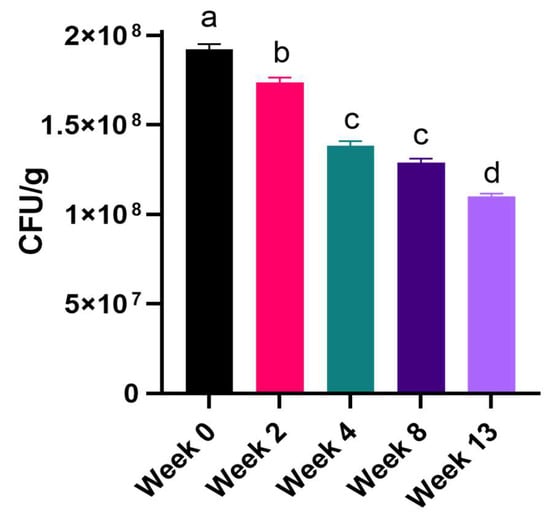
Figure 2.
Survival rate of S. cereviasae var. boulardii in feed mixture during three months of monitoring measured as viable cell count on OGYE agar. Data are results of five different samples and are expressed as CFU/g. Statistical differences to Week 0 are depicted as a different letter. Viability of putative probiotic significantly declines over time but remains above probiotic efficacy threshold of 108 CFU/g.
4. Discussion
Microbial development during cheese production and ripening is an essential and appreciated process that allows metabolic conversion of the main dairy biochemical constituents into smaller compounds conferring the peculiar aromas that characterize different cheese varieties. The bacterial evolution during cheese maturation is a well-known process [53], however, mycobiota undergo modifications in terms of quality and metabolism during ripening [54].
The choice of using a molecular approach proved to be successful for the identification of both yeasts and filamentous fungi. Yeasts are difficult to discriminate from each other and fungal strains sometimes display anomalous conidiophore formation that hampers a reliable morphology-based identification. Our strategy of exploiting barcoding regions of fungal DNA has enabled the identification of yeasts and fungi bearing macro- and micro-morphological mutations, thus highlighting the biodiversity of blue cheeses and their ripening environment.
In the present investigation, an abundant and diverse fungal population has been detected in both the Gorgonzola cheese and in the maturation stores. In the cheese matrix, apart from the expected presence of Penicillium, yeast genera were prevalent, as previously referred to [28]. Conversely, in the ripening rooms, the only yeast species found was Debaryomyces hansenii and several fungal genera, among which Penicillium, Mucor, Cladosporium, Fusicolla, and Aspergillus were detected as also reported by other investigators [55,56].
Apart from filamentous fungi, deriving from contamination or intentional seeding, food-grown yeasts deserve particular attention since they represent a valuable source of putative new probiotic strains [57,58,59]. Along with their antioxidant [58], antibacterial, immune-modulating, and nutritional roles (such as vitamin production) [59,60,61], yeasts can favor the gut colonization by well-known probiotic bacteria like L. rhamnosus [62,63] and L. paracasei [64].
In this study, as far as yeasts are concerned, 30 strains have been directly isolated from Gorgonzola cheese: S. ruberrimus, D. hansenii, Y. lipolytica, Y. deformans, and S. cerevisiae var. boulardii.
The Basidiomycete Sporobolomyces has recently gained attention [65] in the industrial field due to its ability to convert agro-food wastes into valuable products such as lipids [66], carotenoids [67], EPS [68], and several enzymes [65]. Moreover, some species of this genus are also able to produce GABA, which useful in treating mild hypertension and irritable bowel syndrome (IBS) [69]. However, despite its use as a feed supplement to enhance the intensity of the yolk color, this genus has not yet obtained the GRAS status, which is necessary to be included in the probiotic category.
Regarding Ascomycota, Debaryomyces does not show the same performances as true probiotics because it is less tolerant to acidic stress. It displays interesting properties, such as the production of alpha-galactosidase useful to hydrolyze raffinose, an anti-nutritional metabolite of soybeans [70] as well as good adhesion to Caco 2 cells and an overall immune-modulating activity [71]. On the other hand, a fish-isolated D. hansenii CBS 8339, only partially modulated some parameters of the natural immunity [72]. Therefore, despite some promising aspects, the species D. hansenii is only used in aquaculture because of its high osmotolerance (4M NaCl) [73], suggesting that no definitive conclusion on its probiotic properties can be made.
The second Ascomycete isolated in this study, Yarrowia lipolytica, shares with other yeasts the capability to produce a rose-like flavor, 2-phenylethanol, and for this aspect, it is highly appreciated at the industrial level [74]. Some studies report that Yarrowia lipolytica products, like DHA and EPA, can be employed to ameliorate the growth performances of the hybrid sturgeon in combination with Bacillus amyloliquefaciens [75]. Moreover, it has recently been proposed as a putative probiotic for its antibacterial and immune-modulating activities to enhance the growth performances of the white shrimp [76], Nile tilapia [77], and Lutjanus peru [78]. Along with aquatic animals, this yeast can be used alone or together with other putative/ascertained bacterial probiotics (Bacillus licheniformis and Bacillus subtilis) to improve the immunological parameters and growth of birds [79] and mammals [80]. The positive impact of this yeast species, on higher feed conversion rate, animal weight gain, improvement of ruminal microbial populations, and increased IgG and micronutrients (Ca, Mg, P, Fe, Zn, Cu) levels seems mainly due to the production of PUFA [80]. Nevertheless, although this yeast species has been classified as GRAS [81], the risk of possible virulent variants causing infection has been reported [82]. Therefore, safety assays for each single strain are required, as suggested by Groenewald et al. [83]. Finally, S. cerevisiae var. boulardii, whose probiotic potential is well-recognized [15,84], has also been isolated in this study. First discovered in the skins of tropical fruits such as litchis and mangosteens by French microbiologist Henri Boulard as far as 1920, this yeast was described to be able to prevent diarrhea, thus displaying probiotic properties [46]. This species is considered a variant of the best-known S. cerevisiae, showing more than 99% of gene homology according to the Average Nucleotide Identity analysis. However, some genetic traits conferring peculiar metabolic features are different. Among these, the chromosome 9 trisomy can partially explain the higher resistance of S. cerevisiae var. boulardii in acidic environments, a trait that favors the passage and survival through the gastrointestinal system [85]. This yeast also proves to tolerate higher temperatures than S. cerevisiae (37 °C rather than 30 °C) and can survive in the presence of bile salts as well [86].
S. cerevisiae var. boulardii can be employed as an anti-infective agent [87] as it can cause acidification by producing high amounts of acetic acid and, in lesser amounts, also capric acid [88]. The antimicrobial efficacy includes bacteria (Escherichia coli, Salmonella, Shigella) Rotavirus, and Candida. The peculiar anti-Candida albicans effect seems linked to the production of capric acid, which is able to prevent the filamentous growth of this yeast while inhibiting biofilm formation and adhesion [89]. However, S. cerevisiae var. boulardii is especially fitting for treating post-antibiotic diarrhea and colitis caused by C. difficile’s A toxin, since it can prevent both the binding of the A toxin to its receptor (through a proteinaceous factor) and the synthesis of the pro-inflammatory IL-8. In addition, S. cerevisiae var. boulardii can induce the production of IgA anti-A toxin. Similar effects have been observed against enteropathogenic E. coli and Helicobacter pylori as well [89]. These properties are crucial for controlling the overall pathway of inflammation at the gut epithelium level. It should be highlighted that both C. difficile-induced colitis and Candidiasis are often the results of long-term antibiotic therapy in immunocompromised patients since these two organisms are both insensitive to antibiotic treatments. Hence, the use of S. cerevisiae var. boulardii can help in overcoming this threat. In addition, it can be considered an immunomodulating agent and stabilizer of the gut barrier to treat IBS and IBD [90,91].
According to the previously described beneficial effects, the isolated S. cerevisiae var. boulardii strain has been the object of further investigations intended to establish its potential to be employed as an animal feed supplement and its stability in the feed media.
Concerning thermal stability, the temperature of 37 °C supported a growth comparable to that obtained at 35 °C, highlighting the in vivo viability of the strain and likewise ensuring the capability to face the challenges during shelf life, as for other better-characterized S. cerevisiae var. boulardii strains [86].
Regarding the time-course stability in veterinary preparations, the number of CFU was evaluated throughout 13 weeks in five time-points (week 0, 2, 4, 8, and 13) using the final commercial feed (complete pellet to be given to animals) as a matrix, according to the indications of the EFSA guidelines [47]. It is worth mentioning that time-course stability and shelf life are highly influenced by the food/feed matrix in which a probiotic strain is incorporated [46]. As expected, a constant decrease in cell number has been observed from Week 0 to Week 13, since different environmental stressors affect cell viability. Humidity, acid environment, and oxidative stress are the major causes of cell loss [92]. It is for this reason that the recommended number of living organisms to be used as probiotics should be very high (109 CFU for humans and 108 CFU for animals less than 30 kg) [49,50] to counteract both environmental stressors during the shelf-life and endogenous stressors linked to the gastrointestinal environment (competition with resident/pathogenic organisms, enzymes, acid, and bile stress). However, it should be underlined that the cell loss from Week 0 was negligible since the total count was above the threshold of 108 CFU/g (Week 0 = 1.92 (±0.07) × 108 CFU/g, vs. Week 13 = 1.10 (±0.04) × 108 CFU/g). These findings demonstrate an overall low interference due to the feed matrix and hence good cell vitality and stability in the pet-targeted mixture. Improvement in the laboratory methods (i.e., automatic mixing instead of manual mixing of probiotic and feed) could be the objective of future investigations.
Our interesting findings support the use of this food-isolated yeast in veterinary medicine, as previously reported by D’Angelo et al. [93] for another S. cerevisiae var. boulardii strain. One possible limit referred to by these authors is the low adhesion index of their strain on gut mucosa that results in a complete loss of the live probiotics four or five days after the last administration. This implies the need for continuous administration to have prolonged beneficial effects [94]. However, the advantage of using probiotic yeasts is their natural insensitivity to antibacterial targeted antibiotics and the possibility to begin the probiotic treatment simultaneously with the antibiotic therapy. Cavaglieri et al. [18] have already proposed this approach with animals treated with antibiotics (colistin) during infection.
5. Conclusions
The morphological and molecular typing of the cheese mycobiota obtained in this study revealed that the fungal population of Gorgonzola cheese is composed of three main phyla: Ascomycota (most abundant), Mucoromycota, and one species of Basidiomycota. Among filamentous fungi, the most abundant belong to the Penicillium genus, known for its contribution to cheese taste and texture. The ripening rooms displayed a less diverse mycobiota, prevalently based on filamentous fungi that should be taken into consideration since they can occasionally contaminate cheese.
The Gorgonzola cheese under study proved to be rich in interesting yeast species that deserve further investigations for their industrial and putative probiotic potential. Among them, the S. cerevisiae var. boulardii strain, selected for its already-known probiotic properties, revealed good growth performances and high stability at high temperatures and in pet feed matrices. This strain can therefore be considered a promising candidate to be exploited in veterinary medicine either alone or combined with pharmacological treatments. A possible advantage, compared to bacterial-based probiotics, is the lack of interaction between S. cerevisiae var. boulardii and antibiotics, allowing for its administration during anti-infective therapy. Finally, the absence of horizontal gene transfer between yeasts could be particularly appreciated for using S. cerevisiae var. boulardii also in intensive livestock husbandry where the high selective pressure exerted by the widespread use of antibiotics, makes each new bacterial strain a possible vector of resistance genes.
Author Contributions
Conceptualization, E.P., M.F. and N.B.; methodology, S.V., F.B., M.F. and E.M.; validation, S.V., F.B. and N.B.; formal analysis, F.B.; investigation, S.V., M.F. and F.B.; data curation, F.B.; writing—original draft preparation, E.P.; writing—review and editing, E.P., E.M., F.B. and S.V.; supervision, E.P. and N.B.; funding acquisition, E.P., M.F. and N.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research project has been completed with the financial support of Turin University (Fondi ricerca locale ex 60%, PESE_RILO 2021) and the internal grants of Biraghi S.p.A.
Data Availability Statement
Data are contained within the article. DNA sequences are deposited in GenBank under the accession numbers given in Table 1.
Acknowledgments
We are grateful to the laboratory staff, Diego Ternavasio (Gorgonzola DOP Seasoning Department Biraghi S.p.A.) for helping during the experimental part, and to Mariangela Girlanda and Edvige Calaresu for their helpful suggestions.
Conflicts of Interest
Natascia Bruni was employed by Candioli Pharma S.r.l., Beinasco (TO). The remaining authors declare that research was conducted in the absence of any commercial or financial relationships that could be conflicts.
References
- Pieri, R.; Pretolani, R. Il Sistema Agro-Alimentare della Lombardia: Rapporto 2016; Franco Angeli: Milan, Italy, 2017. [Google Scholar]
- MacGibbon, A.K.H. Milk lipids—General characteristics. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 649–654. [Google Scholar]
- Cantor, M.D.; van den Tempel, T.; Hansen, T.K.; Ardö, Y. Blue Cheese. In Cheese: Chemistry, Physics and Microbiology, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 2, pp. 929–954. [Google Scholar] [CrossRef]
- Abbas, A.; Dobson, A.D.W. Yeasts and Molds: Penicillium roqueforti. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 772–775. [Google Scholar]
- Gillot, G.; Jany, J.L.; Coton, M.; Le Floch, G.; Debaets, S.; Ropars, J.; Coton, E. Insights into Penicillium roqueforti morphological and genetic diversity. PLoS ONE 2015, 10, e0129849. [Google Scholar] [CrossRef] [PubMed]
- Gillot, G.; Jany, J.L.; Poirier, E.; Maillard, M.B.; Debaets, S.; Thierry, A.; Coton, M. Functional diversity within the Penicillium roqueforti species. Int. J. Food Microbiol. 2017, 241, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Irlinger, F.; Mounier, J. Microbial interactions in cheese: Implications for cheese quality and safety. Curr. Opin. Biotechnol. 2009, 20, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Buehler, A.J.; Evanowski, R.L.; Martin, N.H.; Boor, K.J.; Wiedmann, M. Internal transcribed spacer (ITS) sequencing reveals considerable fungal diversity in dairy products. J. Dairy Sci. 2017, 100, 8814–8825. [Google Scholar] [CrossRef] [PubMed]
- Keshri, G.; Magan, N. Detection and differentiation between mycotoxigenic and non-mycotoxigenic strains of two Fusarium spp. using volatile production profiles and hydrolytic enzymes. J. Appl. Microbiol. 2000, 89, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Boutrou, R.; Aziza, M.; Amrane, A. Enhanced proteolytic activities of Geotrichum candidum and Penicillium camembertii in mixed culture. Enzym. Microb. 2006, 39, 325–331. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rodrigues, F.C.; Stojanovic-Radic, Z.; Dimitrijevic, M.; Aleksic, A.; Neffe-Skocinska, K.; Zielinska, D.; Kolozyn-Krajewska, D.; Salehi, B.; Prabu, M.S.; et al. Probiotics: Versatile Bioactive Components in Promoting Human Health. Medicina 2020, 56, 433. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.; Eweedah, N.M.; Moustafa, E.M.; Farahat, E.M. Probiotic effects of Aspergillus oryzae on the oxidative status, heat shock protein, and immune related gene expression of Nile tilapia (Oreochromis niloticus) under hypoxia challenge. Aquaculture 2020, 520, 734669. [Google Scholar] [CrossRef]
- Konishi, H.; Isozaki, S.; Kashima, S.; Moriichi, K.; Ichikawa, S.; Yamamoto, K.; Fujiya, M. Probiotic Aspergillus oryzae produces anti-tumor mediator and exerts anti-tumor effects in pancreatic cancer through the p38 MAPK signaling pathway. Sci. Rep. 2021, 11, 11070. [Google Scholar] [CrossRef]
- Bovill, R.; Bew, J.; Robinson, S. Comparison of selective media for the recovery and enumeration of probiotic yeasts from animal feed. Int. J. Food Microbiol. 2001, 67, 55–61. [Google Scholar] [CrossRef]
- Kaźmierczak-Siedlecka, K.; Ruszkowski, J.; Fic, M.; Folwarski, M.; Makarewicz, W. Saccharomyces boulardii CNCM I-745: A Non-bacterial Microorganism Used as Probiotic Agent in Supporting Treatment of Selected Diseases. Curr. Microbiol. 2020, 77, 1987–1996. [Google Scholar] [CrossRef] [PubMed]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, A.; Aziz, T.; Al-Dalali, S.; Zhao, X.; Zhang, J.; ud Din, J.; Chen, C.; Cao, Y.; Yang, Z. Physicochemical and Microbiological Properties of Synbiotic Yogurt Made with Probiotic Yeast Saccharomyces boulardii in Combination with Inulin. Foods 2019, 8, 468. [Google Scholar] [CrossRef] [PubMed]
- Cavaglieri, L.; Montenegro, M.; Escobar, F.; Vergara, D.L.; Bainotti, M.B.; Poloni, V.L. Influence of technological procedures on viability, probiotic and anti-mycotoxin properties of Saccharomyces boulardii RC009 and biological safety studies. Curr. Res. Food Sci. 2021, 4, 132–140. [Google Scholar] [CrossRef]
- Alvarez-Olmos, M.I.; Oberhelman, R.A. Probiotic agents and infectious diseases: A modern perspective on a traditional therapy. Clin. Infect. Dis. 2001, 32, 1567–1576. [Google Scholar] [CrossRef]
- Pessione, E.; Garcia-Contreras, R. Non-conventional antimicrobial agents. In Encyclopedia of Infection and Immunity; Elsevier: Amsterdam, The Netherlands, 2021; pp. 586–607. [Google Scholar]
- Herstad, H.K.; Nesheim, B.B.; L’Abée-Lund, T.; Larsen, S.; Skancke, E. Effects of a probiotic intervention in acute canine gastroenteritis—A controlled clinical trial. J. Small Anim. Pract. 2010, 51, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Ducatelle, R.; Eeckhaut, V.; Haesebrouck, F.; Van Immerseel, F. A review on prebiotics and probiotics for the control of dysbiosis: Present status and future perspectives. Animal 2015, 9, 43–48. [Google Scholar] [CrossRef]
- Arsène, M.M.; Davares, A.K.; Andreevna, S.L.; Vladimirovich, E.A.; Carime, B.Z.; Marouf, R.; Khelifi, I. The use of probiotics in animal feeding for safe pro-duction and as potential alternatives to antibiotics. Vet. World 2021, 14, 319. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.A.; Nicoli, J.R.; Dos Santos Martins, F.; Cardoso, V.N.; Andrade, M.E.R.; Cara Machado, D.C.; Caliari, M.V.; Oliveira, D.R.; Ribeiro, M.R.S. Saccharomyces boulardii as therapeutic alternative in experimental giardiasis. J. Appl. Microbiol. 2021, 131, 460–469. [Google Scholar] [CrossRef]
- Aktas, M.; Borku, M.; Ozkanlar, Y. Efficacy of Saccharomyces boulardii as a probiotic in dogs with lincomycin induced diarrhea. Bull. Vet. Inst. Pulawy 2007, 51, 365–369. [Google Scholar]
- Oswald, I.P.; Castex, M.; Combes, S.; Lippi, Y.; Neves, M.; Hupè, J.; Pinton, P.; Alassane-Kpembi, I. Saccharomyces cerevisiae boulardii reduces the Deoxynivalenol-Induced alteration of the intestinal transcriptome. Toxins 2018, 10, 199. [Google Scholar] [CrossRef] [PubMed]
- Keshri, G.; Magan, N.; Voysey, P. Use of an electronic nose for the early detection and differentiation between spoilage fungi. Lett. Appl. Microbiol. 1998, 27, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Banjara, N.; Suhr, M.J.; Hallen-Adams, H.E. Diversity of yeast and mold species from a variety of cheese types. Curr. Microbiol. 2015, 70, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Garnier, L.; Valence, F.; Pawtowski, A.; Auhustsinava-Galerne, L.; Frotté, N.; Baroncelli, R.; Mounier, J. Diversity of spoilage fungi associated with various French dairy products. Int. J. Food Microbiol. 2017, 241, 191–197. [Google Scholar] [CrossRef]
- Hospodsky, D.; Yamamoto, N.; Peccia, J. Accuracy, precision, and method detection limits of quantitative PCR for airborne bacteria and fungi. Appl. Environ. Microbiol. 2010, 76, 7004–7012. [Google Scholar] [CrossRef] [PubMed]
- Pitkäranta, M.; Meklin, T.; Hyvärinen, A.; Paulin, L.; Auvinen, P.; Nevalainen, A.; Rintala, H. Analysis of fungal flora in indoor dust by ribosomal DNA sequence analysis, quantitative PCR, and culture. Appl. Environ. Microbiol. 2008, 74, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicilliumand Talaromyces; Academic Press Inc.: London, UK, 1979; 634p. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.H. Compendium of Soil Fungi, 2nd ed.; Taxonomically Revised; IHW-Verlag: Eching, Germany, 2007; 672p, ISBN 9783930167692/3930167697. [Google Scholar]
- von Arx, J.A. The Genera of Fungi Sporulating in Pure Culture; Cramer, J., Ed.; J. Cramer.: Vaduz, Germany, 1981; p. 424. [Google Scholar] [CrossRef]
- von Arx, J.A.; Rodrigues De Miranda, M.; Smith, T.H.; Yarrow, D. The genera of yeasts and the yeast-like fungi. Stud. Mycol. 1977, 14, 1–72. [Google Scholar]
- Samson, R.; Horkstra, E.S.; Frisvad, J.C. Introduction to Food and Airborne Fungi, 7th ed.; Centraalbureau Voor Schimmelcultures: Utrecht, The Netherlands, 2004; 389p. [Google Scholar]
- Schipper, M.A.A. Study on variability in Mucor hiemalis and related species. Stud. Mycol. 1973, 4, 1–40. [Google Scholar]
- Schipper, M.A.A. On Mucor circinelloides, Mucor racemosus and related species. Stud. Mycol. 1976, 12, 1–40. [Google Scholar]
- Schipper, M.A.A. (1). On certain species of Mucor with a key to all accepted species. (2). On the genera Rhizomucor and Parasitella. Stud. Mycol. 1978, 17, 1–70. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- Stucky, B.J. SeqTrace: A Graphical Tool for Rapidly Processing DNA Sequencing Chromatograms. J. Biomol. Tech. 2012, 23, 90–93. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Westerdijk Fungal Biodiversity Institute. Available online: http://www.westerdijkinstitute.nl (accessed on 30 March 2023).
- Lazo-Vélez, M.A.; Serna-Saldìvar, S.O.; Rosales-Medina, M.F.; Tinoco-Alvear, M.; Briones-Garcia, M. Application of Saccharomyces cerevisiae var. boulardii in food processing: A review. J. Appl. Microbiol. 2018, 125, 943–951. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Guidance on the identity, characterisation and conditions of use of feed additives. EFSA J. 2017, 15, e05023. [Google Scholar] [CrossRef] [PubMed]
- EN15789:2022; Animal Feeding Stuffs: Methods of Sampling and Analysis—Detection and Enumeration of Saccharomyces Cerevisiae Used as Feed Additive. European Committee for Standardization: Brussels, Belgium, 2022.
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Leuschner, R.G.; Bew, J.; Bertin, G. Validation of an official control method for enumeration of authorized probiotic yeast in animal feed. Syst. Appl. Microbiol. 2003, 26, 147–153. [Google Scholar] [CrossRef]
- COMMISSION IMPLEMENTING REGULATION (EU) 2015/502 of 24 March 2015 Concerning the Authorisation of the Preparation of Saccharomyces cerevisiae NCYC R404 as a Feed Additive for Dairy Cows (Holder of the Authorisation Micro Bio-System Ltd.) L 79/57 25.03.2015. Off. J. Eur. Union, 2015.
- COMMISSION IMPLEMENTING REGULATION (EU) 2019/899 of 29 May 2019 Concerning the Renewal of the Authorisation of Saccharomyces cerevisiae CNCM I-4407 as a Feed Additive for Lambs for Fattening, Dairy Goats, Dairy Sheep, Dairy Buffaloes, Horses and Pigs for Fattening and Repealing Regulations (EC) No 1447/2006, (EC) No 188/2007, (EC) No 232/2009, (EC) No 186/2007 and (EC) No 209/2008 (Holder of Authorisation S.I. Lesaffre) L144/32 3.6.2019. Off. J. Eur. Union, 2019.
- Swearingen, P.A.; O’Sullivan, D.J.; Warthesen, J.J. Isolation, characterization, and influence of native, nonstarter lactic acid bacteria on Cheddar cheese quality. J. Dairy Sci. 2001, 84, 50–59. [Google Scholar] [CrossRef]
- Ledenbach, L.H.; Marshall, R.T. Microbiological Spoilage of Dairy Products. In Compendium of the Microbiological Spoilage of Foods and Beverages. Food Microbiology and Food Safety; Sperber, W., Doyle, M., Eds.; Springer: New York, NY, USA, 2009; pp. 41–67. [Google Scholar]
- Mounier, J.; Goerges, S.; Gelsomino, R.; Vancanneyt, M.; Vandemeulebroecke, K.; Hoste, B.; Cogan, T.M. Sources of the adventitious microflora of a smear ripened cheese. J. Appl. Microbiol. 2006, 101, 668–681. [Google Scholar] [CrossRef]
- Serra, R.; Abrunhosa, L.; Kozakiewicz, Z.; Venâncio, A.; Lima, N. Use of ozone to reduce molds in a cheese ripening room. J. Food Prot. 2003, 66, 2355–2358. [Google Scholar] [CrossRef] [PubMed]
- Palma, M.L.; Zamith-Miranda, D.; Martins, F.S.; Bozza, F.A.; Nimrichter, L.; Montero-Lomeli, M.; Douradinha, B. Probiotic Saccharomyces cerevisiae strains as biotherapeutic tools: Is there room for improvement? Appl. Microbiol. Biotechnol. 2015, 99, 6563–6570. [Google Scholar] [CrossRef] [PubMed]
- Menezes, A.G.T.; Ramos, C.L.; Cenzi, G.; Melo, D.S.; Dias, D.R.; Schwan, R.F. Probiotic potential, antioxidant activity, and phytase production of indigenous yeasts isolated from indigenous fermented foods. Probiotics Antimicro. Prot. 2020, 12, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Syal, P.; Vohra, A. Probiotic potential of yeasts isolated from traditional Indian fermented foods. Int. J. Microbiol. Res. 2013, 5, 390. [Google Scholar] [CrossRef]
- Oliveira, T.; Ramalhosa, E.; Nunes, L.; Pereira, J.A.; Colla, E.; Pereira, E.L. Probiotic potential of indigenous yeasts isolated during the fermentation of table olives from Northeast of Portugal. Innov. Food Sci. Emerg. Technol. 2017, 44, 167–172. [Google Scholar] [CrossRef]
- Lara-Hidalgo, C.E.; Hernández-Sánchez, H.; Hernández-Rodríguez, C.; Dorantes-Álvarez, L. Yeasts in fermented foods and their probiotic potential. Austin J. Nutr. Metab. 2017, 4, 1045. [Google Scholar]
- Liu, S.Q.; Tsao, M. Enhancing stability of lactic acid bacteria and probiotics by Williopsis saturnus var. saturnusin fermented milks. Nutr. Food Sci. 2010, 40, 314–322. [Google Scholar] [CrossRef]
- Suharja, A.; Henriksson, A.; Liu, S.Q. Impact of Saccharomyces cerevisiae on viability of probiotic Lactobacillus rhamnosus in fermented milk under ambient conditions. J. Food Process. Preserv. 2014, 38, 326–337. [Google Scholar] [CrossRef]
- Xie, N.; Zhou, T.; Li, B. Kefir yeasts enhance probiotic potentials of Lactobacillus paracasei H9: The positive effects of co-aggregation between the two strains. Food Res. Int. 2012, 45, 394–401. [Google Scholar] [CrossRef]
- Kot, A.M.; Kieliszek, M.; Piwowarek, K.; Błażejak, S.; Mussagy, C.U. Sporobolomyces and Sporidiobolus–non-conventional yeasts for use in industries. Fungal Biol. Rev. 2021, 37, 41–58. [Google Scholar] [CrossRef]
- Patel, A.; Karageorgou, D.; Rova, E.; Katapodis, P.; Rova, U.; Christakopoulos, P.; Matsakas, L. An Overview of Potential Oleaginous Microorganisms and Their Role in Biodiesel and Omega-3 Fatty Acid-Based Industries. Microorganisms 2020, 8, 434. [Google Scholar] [CrossRef] [PubMed]
- Mussagy, C.U.; Khan, S.; Kot, A.M. Current developments on the application of microbial carotenoids as an alternative to synthetic pigments. Crit. Rev. Food Sci. Nutr. 2021, 62, 6932–6946. [Google Scholar] [CrossRef] [PubMed]
- Gientka, I.; B1azejak, S.; Stasiak-Rozanska, L.; Chlebowska-Smigiel, A. Exopolysaccharides from yeast: Insight into optimal conditions for biosynthesis, chemical composition and functional properties. Acta Sci. Pol. Technol. Aliment. 2015, 14, 283–292. [Google Scholar] [CrossRef]
- Han, S.M.; Lee, J.S. Production and its anti-hyperglycemic effects of γ-aminobutyric acid from the wild yeast strain Pichia silvicola UL6-1 and Sporobolomyces carnicolor 402-JB-1. Mycobiology 2017, 45, 199–203. [Google Scholar] [CrossRef]
- Viana, P.A.; de Rezende, S.T.; Passos, F.M.L.; Machado, S.G.; Maitan, G.P.; Coelho, V.T.D. Alpha-Galactosidases production by Debaryomyces hansenii UFV-1. Food Sci. Biotechnol. 2011, 20, 601–606. [Google Scholar] [CrossRef]
- Ochangco, H.S.; Gamero, A.; Smith, I.M.; Christensen, J.E.; Jespersen, L.; Arneborg, N. In vitro investigation of Debaryomyces hansenii strains for potential probiotic properties. World J. Microbiol. Biotechnol. 2016, 32, 141. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Salinas, I.; Cuesta, A.; Meseguer, J.; Tovar-Ramirez, D.; Ascencio-Valle, F.; Esteban, M.Á. Oral delivery of live yeast Debaryomyces hansenii modulates the main innate immune parameters and the expression of immune-relevant genes in the gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2008, 25, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Angulo, M.; Reyes-Becerril, M.; Medina-Córdova, N.; Tovar-Ramírez, D.; Angulo, C. Probiotic and nutritional effects of Debaryomyces hansenii on animals. Appl. Microbiol. Biotechnol. 2020, 104, 7689–7699. [Google Scholar] [CrossRef] [PubMed]
- Celińska, E.; Kubiak, P.; Białas, W.; Dziadas, M.; Grajek, W. Yarrowia lipolytica: The novel and promising 2-phenylethanol producer. J. Ind. Microbiol. Biotechnol. 2013, 40, 389–392. [Google Scholar] [CrossRef]
- Fei, H.; Lin, G.D.; Zheng, C.C.; Huang, M.M.; Qian, S.C.; Wu, Z.J.; Han, B.N. Effects of Bacillus amyloliquefaciens and Yarrowia lipolytica lipase 2 on immunology and growth performance of Hybrid sturgeon. Fish Shellfish Immunol. 2018, 82, 250–257. [Google Scholar] [CrossRef]
- Licona-Jain, A.; Campa-Córdova, Á.; Luna-González, A.; Racotta, I.S.; Tello, M.; Angulo, C. Dietary supplementation of marine yeast Yarrowia lipolytica modulates immune response in Litopenaeus vannamei. Fish Shellfish Immunol. 2020, 105, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Neuls, L.; de Souza, V.J.; Romão, S.; Bitencourt, T.B.; Ramos, C.J.R.; Parra, J.E.G.; Cazarolli, L.H. Immunomodulatory effects of Yarrowia lipolytica as a food additive in the diet of Nile tilapia. Fish Shellfish Immunol. 2021, 119, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Becerril, M.; Alamillo, E.; Angulo, C. Probiotic and immunomodulatory activity of marine yeast Yarrowia lipolytica strains and response against Vibrio parahaemolyticus in fish. Probiotics Antimicro. Prot. 2021, 13, 1292–1305. [Google Scholar] [CrossRef] [PubMed]
- Czech, A.; Sembratowicz, I.; Zieba, G. Effect of the use of Yarrowia lipolytica and Saccharomyces cerevisiae yeast with a probiotic in the diet of turkeys on their gut microbiota and immunity. Vet. Med. 2020, 65, 174–182. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Esteban, M.Á.; Angulo, C. Yarrowia lipolytica, health benefits for animals. Appl. Microbiol. Biotechnol. 2021, 105, 7577–7592. [Google Scholar] [CrossRef] [PubMed]
- Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; Pelaez, C.; et al. Scientific opinion on the safety of Yarrowia lipolytica yeast biomass as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, 5594. [Google Scholar] [CrossRef]
- Desnos-Ollivier, M.; Letscher-Bru, V.; Neuvéglise, C.; Dromer, F. Yarrowia lipolytica causes sporadic cases and local outbreaks of infection and colonisation. Mycoses 2020, 63, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Groenewald, M.; Boekhout, T.; Neuveglise, C.; Gaillardin, C.; Van Dijck, P.W.M.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- McFarland, L.V. Systematic review and meta-analysis of Saccharomyces boulardii in adult patients. World J. Gastroenterol. 2010, 16, 2202–2222. [Google Scholar] [CrossRef]
- Edwards-Ingram, L.; Gitsham, P.; Burton, N.; Warhurst, G.; Clarke, I.; Hoyle, D.; Oliver, G.S.; Stateva, L. Genotypic and Physiological Characterization of Saccharomyces boulardii, the probiotic strain of Saccharomyces Cerevisiae. Appl. Environ. Microbiol. 2007, 73, 2458–2467. [Google Scholar] [CrossRef]
- Fietto, J.L.; Araújo, R.S.; Valadão, F.N.; Fietto, L.G.; Brandão, R.L.; Neves, M.J.; Castro, I.M. Molecular and physiological comparisons between Saccharomyces cerevisiae and Saccharomyces boulardii. Can. J. Microbiol. 2004, 50, 615–621. [Google Scholar] [CrossRef]
- Rodrigues, A.C.; Cara, D.C.; Fretez, S.H.; Cunha, F.Q.; Vieira, E.C.; Nicoli, J.R.; Vieira, L.Q. Saccharomyces boulardii stimulates sIgA production and the phagocytic system of gnotobiotic mice. J. Appl. Microbiol. 2000, 89, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Thevelein, M.J.; Foulquié-Moreno, R.M.; De Graeve, S.; Vandercruys, P.; Offei, B. Unique genetic basis of the distinct antibiotic potency of high acetic acid production in the probiotic yeast Saccharomyces cerevisiae var. boulardii. Genome Res. 2019, 29, 1478–1494. [Google Scholar] [CrossRef]
- Rampal, P.; Czerucka, D. Diversity of Saccharomyces boulardii CNCM I-745 mechanisms of action against intestinal infections. World J. Gastroenterol. 2019, 25, 2188–2203. [Google Scholar] [CrossRef]
- Kelesidis, T. Efficacy and safety of the probiotic Saccharomyces boulardii for the prevention and therapy of gastrointestinal disorders. Ther. Adv. Gastroenterol. 2012, 5, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Soares, R.L. Irritable bowel syndrome: A clinical review. World J. Gastroenterol. 2014, 20, 12144–12160. [Google Scholar] [CrossRef] [PubMed]
- Dinkçi, N.; Akdeniz, V.; Akalin, A.S. Survival of probiotics in functional foods during shelf life. In Food Quality and Shelf Life; Charis, M.G., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 201–233. [Google Scholar]
- D’angelo, S.; Fracassi, F.; Bresciani, F.; Galuppi, R.; Diana, A.; Linta, N.; Bettini, G.; Morini, M.; Pietra, M. Effect of Saccharomyces boulardii in dog with chronic enteropathies: Double-blinded, placebo-controlled study. Vet. Rec. 2018, 182, 258. [Google Scholar] [CrossRef]
- Blehaut, H.; Massot, J.; Elmer, G.; Levy, R.H. Disposition kinetics of Saccharomyces boulardii in man and rat. Drug Dispos. 1989, 10, 353–364. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).