

Comparative Transcriptomic Analysis Reveals Variable Responses to the Brown Planthopper Nilaparvata lugens in Different Rice Cultivars

 , and
, and 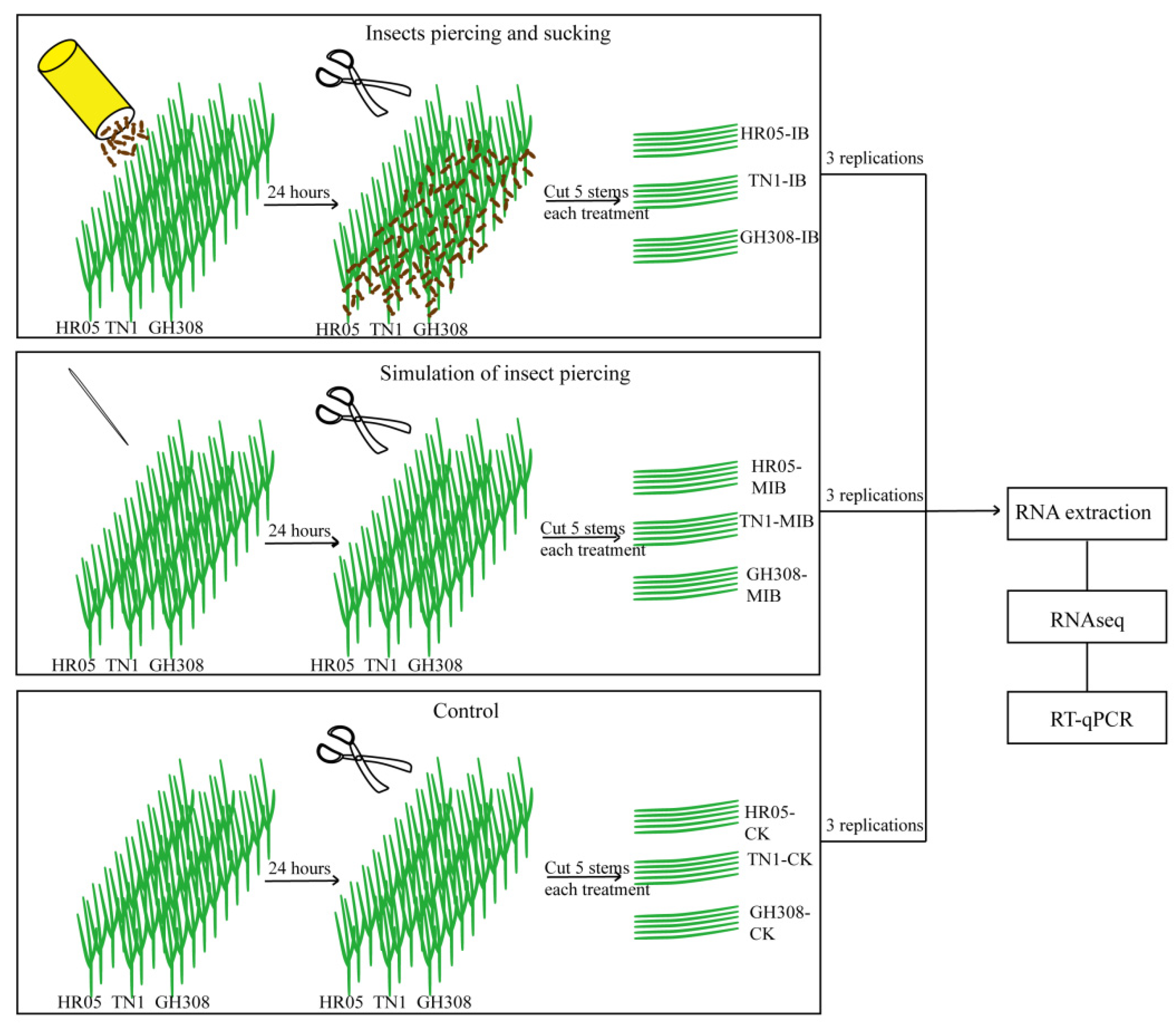{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Plants
2.2. Sample Collection
2.3. RNA Extraction and Sequencing
2.4. Quality Control and Correlation Analysis of Transcriptome Data
2.5. Transcriptome Analysis
2.6. Trend and WGCNA Analysis
2.7. Real-Time Fluorescence Quantitative PCR
3. Results
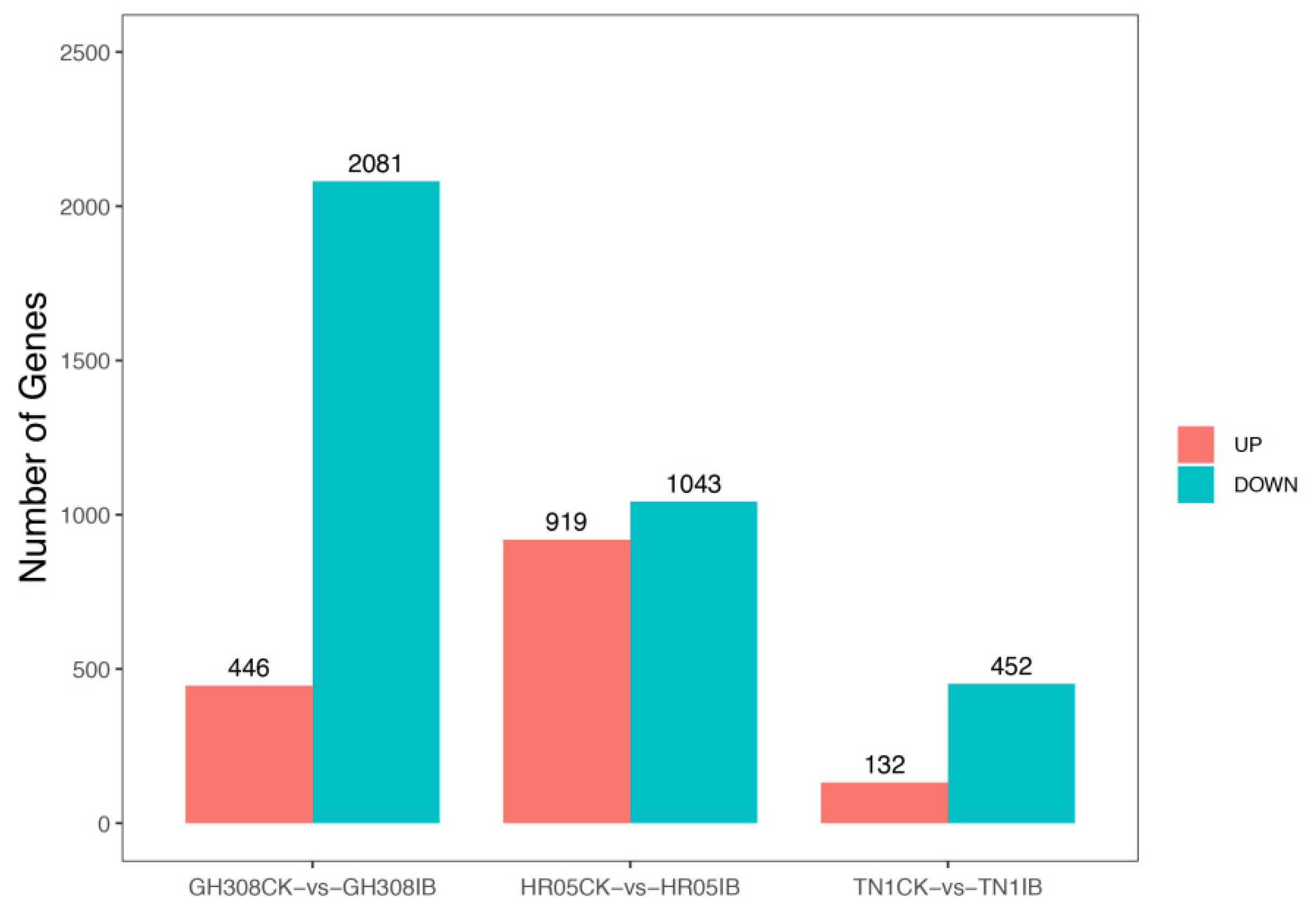
3.1. Identification of DEGs in Response to BPH
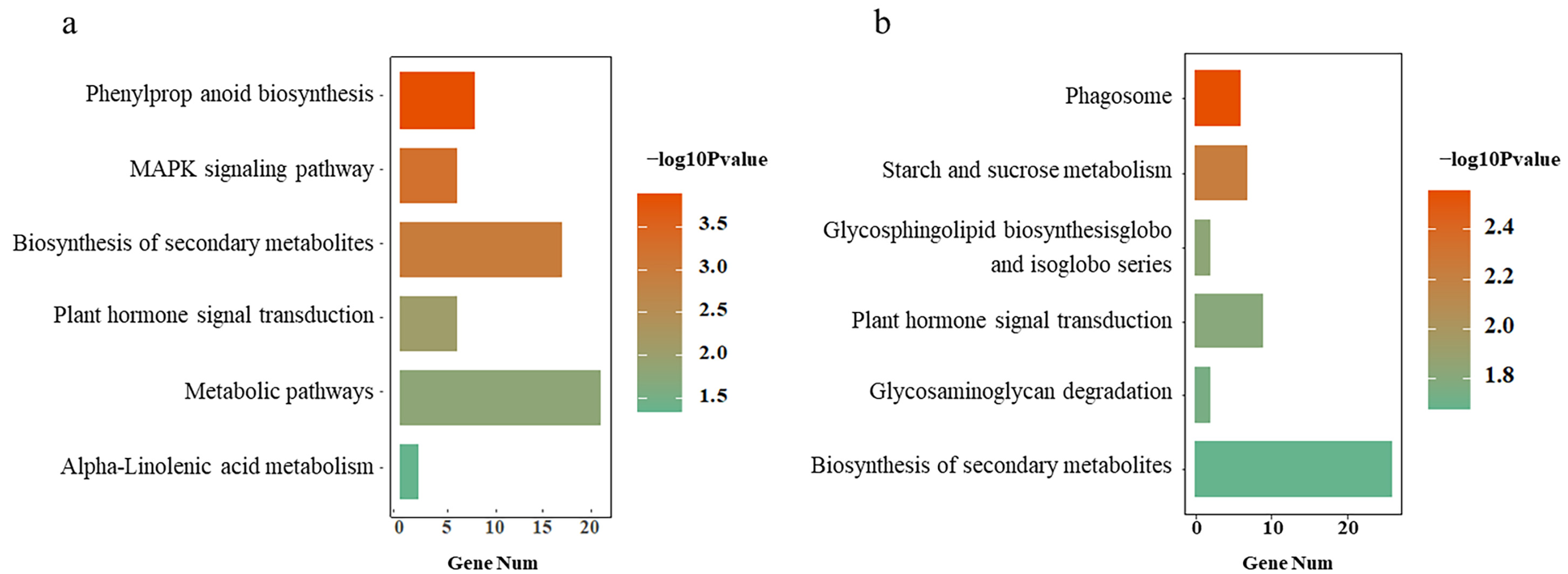
3.2. Consistently Regulated Genes in the Three Rice Cultivars under BPH Stress
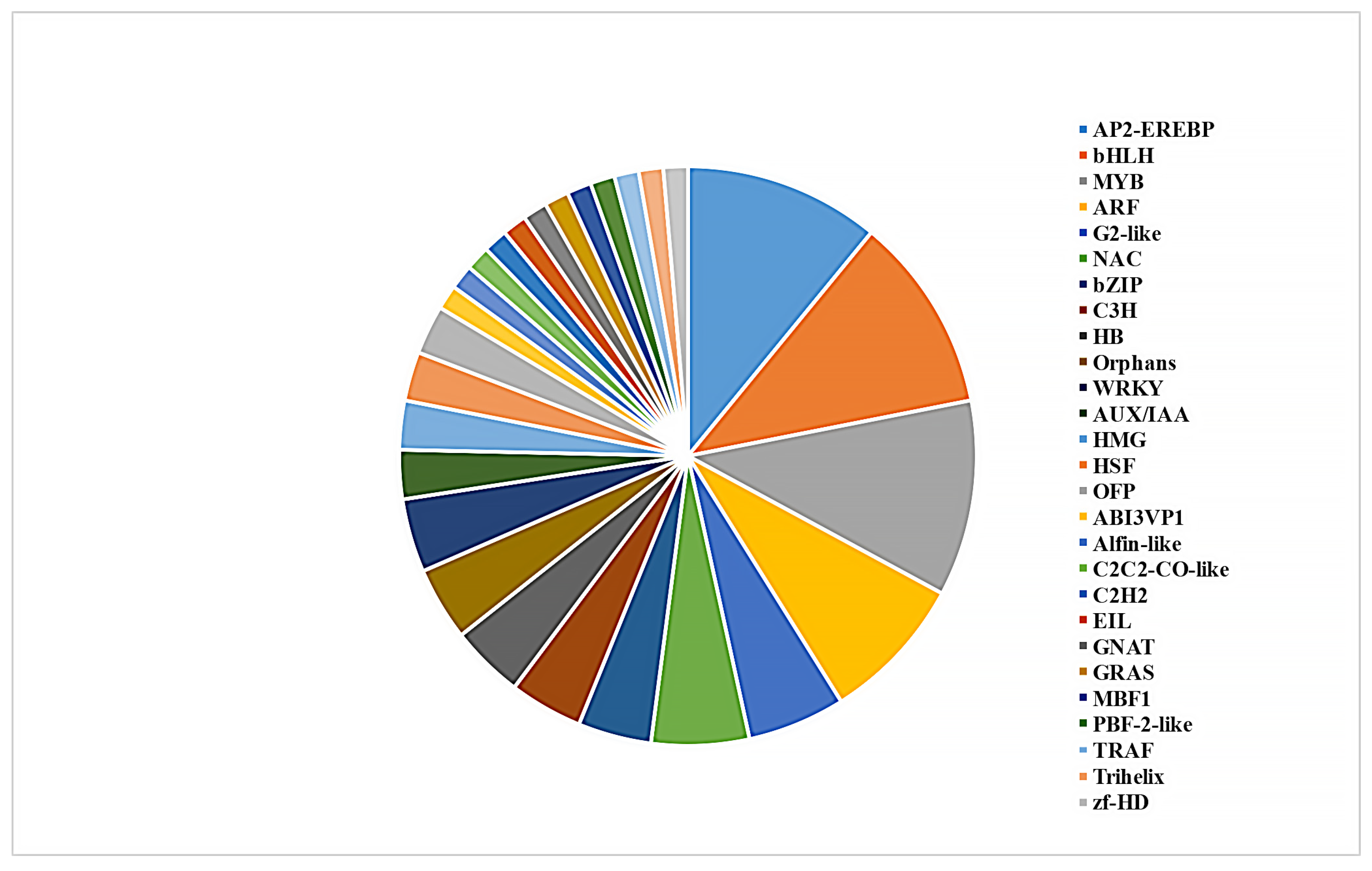
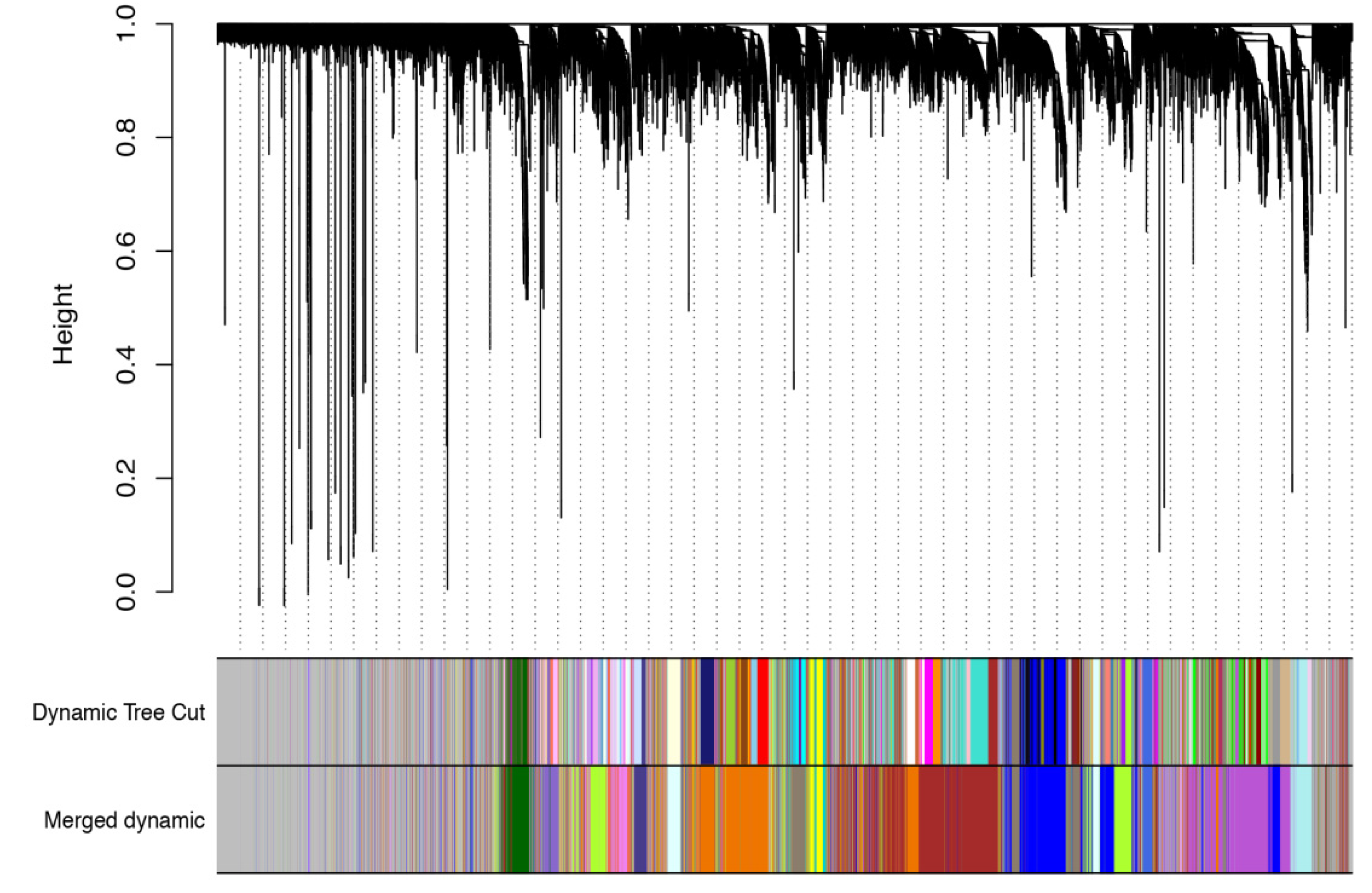
3.3. WGCNA Analysis of Different Experimental Conditions
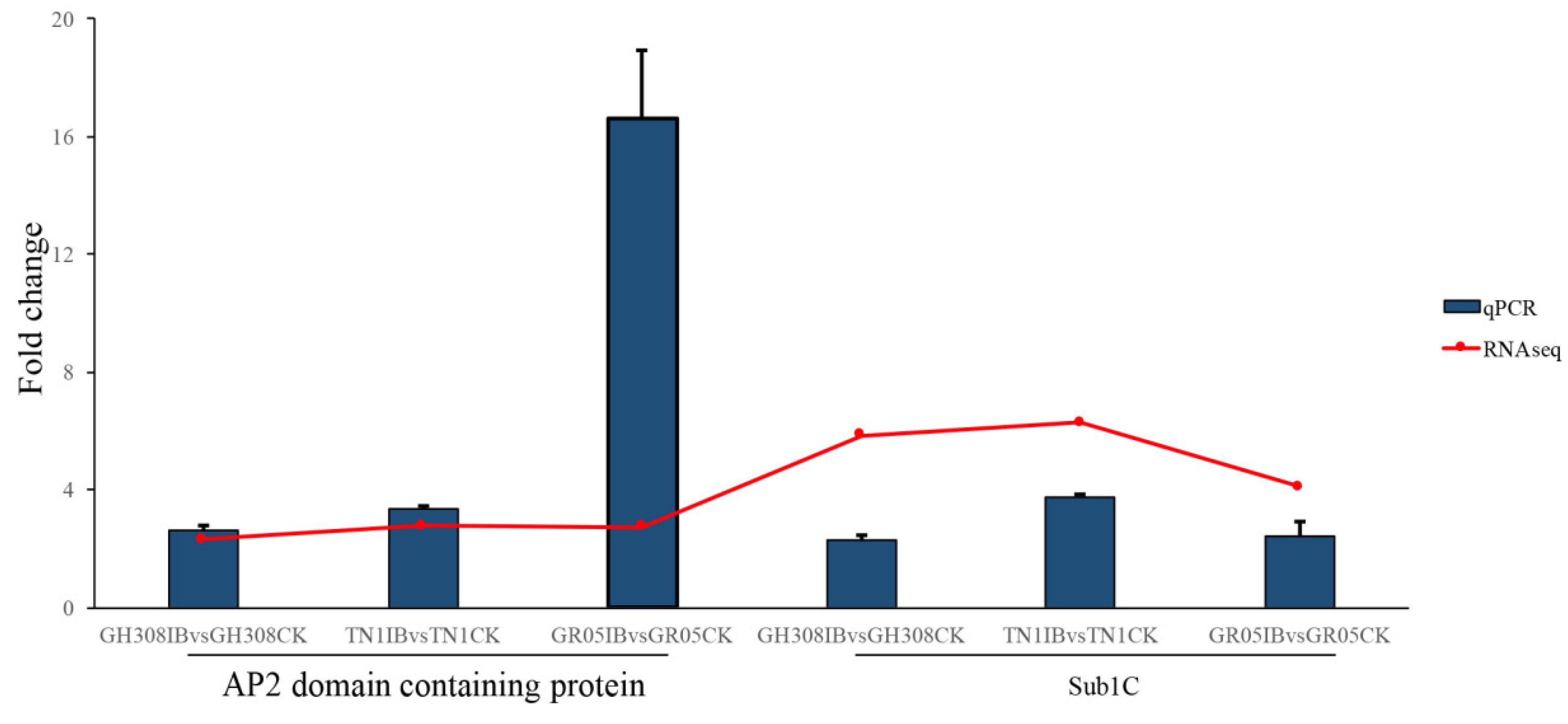
3.4. Technical Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Khush, G.S. Green revolution: Preparing for the 21st century. Genome 1999, 42, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Wing, R.A.; Ruan, J.; Zhang, G.; Wei, C.; Alexandrov, N.; McNally, K.L.; et al. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Xu, Q.; Xu, Z.J.; Chen, W.F. Effect of rice breeding process on improvement of yield and quality in China. Rice Sci. 2020, 27, 363–367. [Google Scholar] [CrossRef]
- Patel, S.; Singh, C.P. Seasonal incidence of rice stem borer, Scirpophaga incertulas (Walker) on different varieties of rice in relation to weather parameters. J. Entomol. Zool. 2017, 5, 80–83. [Google Scholar]
- Brar, D.S.; Virk, P.S.; Jena, K.K.; Khush, G.S. Breeding for resistance to planthoppers in rice. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; International Rice Research Institute: Metro Manila, Philippines, 2009; pp. 401–428. [Google Scholar]
- Haliru, B.S.; Rafii, M.Y.; Mazlan, N.; Ramlee, S.I.; Bashir, Y.R. Recent strategies for detection and improvement of brown planthopper resistance genes in rice: A review. Plants 2020, 9, 1202. [Google Scholar] [CrossRef]
- Cabauatan, P.Q.; Cabunagan, R.C.; Choi, I.R. Rice viruses transmitted by the brown planthopper Nilaparvata lugens (Stål). In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; International Rice Research Institute: Metro Manila, Philippines, 2009; pp. 357–368. [Google Scholar]
- Cheng, X.Y.; Zhu, L.L.; He, G.C. Towards understanding of molecular interactions between rice and the brown planthopper. Mol. Plant 2013, 6, 621–634. [Google Scholar] [CrossRef]
- Jing, S.L.; Zhao, Y.; Du, B.; Chen, R.Z.; Zhu, L.L.; He, G.C. Genomics of interaction between the brown planthopper and rice. Curr. Opin. Insect Sci. 2017, 19, 82–87. [Google Scholar] [CrossRef]
- Ghosh, A.; Chatterjee, M.L.; Samanta, A. Field evaluation of some new insecticides against brown planthopper Nilaparvata lugens (stål) in rice. J. Entomol. Res. 2010, 34, 35–37. [Google Scholar]
- Prashant, S.T.; Gowda, D. Evaluation of efficacy of insecticides against brown planthopper (bph), Nilaparvata lugens (stal.) in rice ecosystem. Environ. Ecol. 2015, 33, 228–231. [Google Scholar]
- Patil, S.D.; Patil, H.M.; Bhoite, K.D.; Kusalkar, D.V. Evaluation of insecticides against brown plant hopper, Nilaparvata lugens (stal) in rice, Oryza sativa L. J. Pharmacogn. Phytochem. 2020, 9, 1865–1868. [Google Scholar]
- Visarto, P.; Zalucki, M.P.; Nesbitt, H.J.; Jahn, G.C. Effect of fertilizer, pesticide treatment, and plant variety on the realized fecundity and survival rates of brown planthopper, Nilaparvata lugens (Stål) (Homoptera: Delphacidae)-generating outbreaks in Cambodia. J. Asia Pac. Entomol. 2001, 4, 75–84. [Google Scholar] [CrossRef]
- Zhang, X.; Liao, X.; Mao, K.; Zhang, K.; Wan, H.; Li, J. Insecticide resistance monitoring and correlation analysis of insecticides in field populations of the brown planthopper Nilaparvata lugens (Stål) in China 2012–2014. Pestic. Biochem. Phys. 2016, 132, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.J.; Xue, J.; Lu, B.; Zhang, X.C.; Zhou, J.C.; He, S.F.; Ma, X.F.; Jiang, Y.Q.; Fan, H.W.; Xu, J.Y.; et al. Two insulin receptors determine alternative wing morphs in planthoppers. Nature 2015, 519, 464–467. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.J.; Zhang, C.X. Insulin receptors and wing dimorphism in rice planthoppers. Phil. Tran. R. Soc. B. 2017, 372, 20150489. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Jiang, C.; Zhang, Y.; Yang, X.; Cheng, D. Radar observations of the seasonal migration of brown planthopper Nilaparvata lugens (Stål) in Southern China. Bull. Entomol. Res. 2014, 104, 731–741. [Google Scholar] [CrossRef]
- Bao, Y.X.; Ding, W.W.; Xie, X.J.; Lan, P.; Lu, M.H. Statistical analysis on the influence of the landfalling strong tropical cyclones in the catastrophic migrations of Nilaparvata lugens (Stål) in China. J. Trop. Meteorol. 2014, 1, 8–16. [Google Scholar]
- Sharma, H.C.; Ortiz, R. Host plant resistance to insects: An eco-friendly approach for pest management and environment conservation. J. Environ. Biol. 2002, 23, 111–135. [Google Scholar]
- Savary, S.; Horgan, F.; Willocquet, L.; Heong, K.L. A review of principles for sustainable pest management in rice. Crop Prot. 2012, 32, 54–63. [Google Scholar] [CrossRef]
- Trapero, C.; Wilson, I.W.; Stiller, W.N.; Wilson, L.J. Enhancing integrated pest management in GM cotton systems using host plant resistance. Front. Plant Sci. 2016, 7, 500. [Google Scholar] [CrossRef]
- Véronique, L.; Boissot, N.; Gallois, J.L. Host plant resistance to pests and pathogens, the genetic leverage in integrated pest and disease management. Integr. Pest Dis. Manag. Greenh. Crops 2020, 9, 259–283. [Google Scholar]
- Mookiah, S.; Sivasubramaniam, B.; Thangaraj, T.; Govindaraj, S. Host Plant Resistance. In Molecular Approaches for Sustainable Insect Pest Management; Omkar, Ed.; Springer: Singapore, 2021; pp. 1–56. [Google Scholar]
- Eyles, A.; Bonello, P.; Ganley, R.; Mohammed, C. Induced resistance to pests and pathogens in trees. New Phytol. 2010, 185, 893–908. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, R.K.; Sarao, P.S.; Sharma, N. Antibiosis in wild rice accessions induced by Nilaparvata lugens (Stål) feeding. Phytoparasitica 2020, 48, 801–812. [Google Scholar] [CrossRef]
- Zandt, P.A.V. Plant defense, growth, and habitat: A comparative assessment of constitutive and induced resistance. Ecology 2007, 88, 1984–1993. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, M.; Hao, P.; Yang, Z.; He, G. Penetration into rice tissues by brown planthopper and fine structure of the salivary sheaths. Entomol. Exp. Appl. 2008, 129, 295–307. [Google Scholar] [CrossRef]
- Ji, R.; Yu, H.; Fu, Q.; Chen, H.; Ye, W.; Li, S. Comparative transcriptome analysis of salivary glands of two populations of rice brown planthopper, Nilaparvata lugens, that differ in virulence. PLoS ONE 2013, 8, e79612. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Liu, C.; Wang, Y.; Chen, R.; Tang, M.; Du, B. Herbivore-induced callose deposition on the sieve plates of rice: An important mechanism for host resistance. Plant Physiol. 2008, 146, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Jena, K.K.; Kim, S.M. Current status of brown planthopper (bph) resistance and genetics. Rice 2010, 3, 161–171. [Google Scholar] [CrossRef]
- Li, C.Y.; Luo, C.; Zhou, Z.H.; Wang, R.; Ling, F.; Xiao, L.T.; Lin, Y.J.; Chen, H. Gene expression and plant hormone levels in two contrasting rice genotypes responding to brown planthopper infestation. BMC Plant Biol. 2017, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Li, H.P.; Zhou, Z.H.; Hua, H.X.; Ma, W.H. Comparative transcriptome analysis of defense response of rice to Nilaparvata lugens and Chilo suppressalis infestation. Int. J. Biol. Macromol. 2020, 163, 2270–2285. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Bao, Y.Y.; Li, B.L.; Cheng, Y.B.; Peng, Z.Y.; Liu, H.; Xu, H.J.; Zhu, Z.R.; Lou, Y.G.; Cheng, J.A.; et al. Transcriptome analysis of the brown planthopper Nilaparvata lugens. PloS ONE 2010, 5, e14233. [Google Scholar] [CrossRef]
- Sun, X.Y.; Wang, Z.Y.; Gu, Q.S.; Li, H.L.; Han, W.L.; Shi, Y. Transcriptome analysis of Cucumis sativus infected by Cucurbit chlorotic yellows virus. Virol. J. 2017, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Seifi, A.; Visser, R.G.F.; Bai, Y. How to effectively deploy plant resistances to pests and pathogens in crop breeding. Euphytica 2013, 190, 321–334. [Google Scholar] [CrossRef]
- Hammond, K.K.E.; Parker, J.E. Deciphering plant-pathogen communication: Fresh perspectives for molecular resistance breeding. Curr. Opin. Biotech. 2003, 14, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Hairmansis, A.; Nasution, A.; Utami, D.W. Introgression of bacterial blight resistance gene Xa7 into popular indonesian rice varieties through backcross and molecular breeding. IOP Conf. Ser. Earth Environ. Sci. 2020, 457, 012050. [Google Scholar] [CrossRef]
- Xiao, N.; Wu, Y.Y.; Li, A.H. Strategy for use of rice blast resistance genes in rice molecular breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar]
- Lu, Y.H.; Wu, K.M.; Jiang, Y.Y.; Guo, Y.Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Wang, X.; Yan, F.; Wang, X.; Li, R.; Cheng, J.; Lou, Y.G. Genome-wide transcriptional changes and defence-related chemical profiling of rice in response to infestation by the rice striped stem borer Chilo suppressalis. Physiol. Plant. 2011, 143, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Ralph, S.; Oddy, C.; Cooper, D.; Yueh, H.; Jancsik, S.; Kolosova, N.; Bohlmann, J. Genomics of hybrid poplar (Populus trichocarpa × deltoides) interacting with forest tent caterpillars (Malacosoma disstria): Normalized and full-length cDNA libraries, expressed sequence tags, and a cDNA microarray for the study of insect-induced defences in poplar. Mol. Ecol. 2006, 15, 1275–1297. [Google Scholar]
- Donze-Reiner, T.; Palmer, N.A.; Scully, E.D.; Prochaska, T.J.; Koch, K.G.; Heng-Moss, T.; Sarath, G. Transcriptional analysis of defense mechanisms in upland tetraploid switchgrass to greenbugs. BMC Plant. Boil. 2017, 17, 46. [Google Scholar] [CrossRef]
- Lu, J.; Ju, H.P.; Zhou, G.X.; Zhu, C.S.; Erb, M.; Wang, X.P.; Wang, P.; Lou, Y.G. An EAR-motif-containing ERF transcription factor affects herbivore-induced signaling, defense and resistance in rice. Plant. J. 2011, 68, 583–596. [Google Scholar] [CrossRef]
- De Vos, M.; Denekamp, M.; Dicke, M.; Vuylsteke, M.; Van Loon, L.C.; Smeekens, S.C.; Pieterse, C. The Arabidopsis thaliana transcription factor AtMYB102 functions in defense against the insect herbivore Pieris rapae. Plant. Signal. Behav. 2006, 1, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Guo, H.M.; Li, H.C.; Zhang, H.; Miao, X.X. Identification of transcription factors potential related to brown planthopper resistance in rice via microarray expression profiling. BMC Genom. 2012, 13, 687. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; He, H.; Zhang, J.; Jiang, Z.; Qiu, L.; Zhan, Z. Comparative Transcriptomic Analysis Reveals Variable Responses to the Brown Planthopper Nilaparvata lugens in Different Rice Cultivars. Crops 2023, 3, 40-52. https://doi.org/10.3390/crops3010005
Shi L, He H, Zhang J, Jiang Z, Qiu L, Zhan Z. Comparative Transcriptomic Analysis Reveals Variable Responses to the Brown Planthopper Nilaparvata lugens in Different Rice Cultivars. Crops. 2023; 3(1):40-52. https://doi.org/10.3390/crops3010005
Chicago/Turabian StyleShi, Longqing, Haifang He, Junian Zhang, Zhaowei Jiang, Liangmiao Qiu, and Zhixiong Zhan. 2023. "Comparative Transcriptomic Analysis Reveals Variable Responses to the Brown Planthopper Nilaparvata lugens in Different Rice Cultivars" Crops 3, no. 1: 40-52. https://doi.org/10.3390/crops3010005
APA StyleShi, L., He, H., Zhang, J., Jiang, Z., Qiu, L., & Zhan, Z. (2023). Comparative Transcriptomic Analysis Reveals Variable Responses to the Brown Planthopper Nilaparvata lugens in Different Rice Cultivars. Crops, 3(1), 40-52. https://doi.org/10.3390/crops3010005