

Early Life Stress and Gut Microbiome Dysbiosis: A Narrative Review


Abstract
1. Introduction
2. Method
3. ELS and Its Impact on Mental Health
3.1. Stress System Elements
3.2. Neurological Alterations in ELS
3.2.1. Alterations in Prefrontal–Hippocampal–Amygdala Circuits
3.2.2. Alterations in Prefrontal-Striatal Dopaminergic Circuits
4. Gut Microbiome in Early Life
5. Microbiological Aspects of ELS
5.1. ELS in Animal Models
5.2. ELS in Humans
5.2.1. Prenatal Stress Studies
5.2.2. Postnatal Stress Studies
6. Microbial Key Regulators of ELS
6.1. The Role of SCFAs in ELS
6.2. The Role of Amino Acids in ELS
6.3. The Role of Psychobiotics in ELS
7. Discussion
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACTH | Adrenocorticotropin |
| ACEs | Adverse childhood experiences |
| ADHD | Attention deficit hyperactivity disorder |
| ASD | Autism spectrum disorder |
| ANS | Autonomic nervous system |
| CNS | Central nervous system |
| CRH | Corticotropin-releasing hormone |
| C-section | Cesarean section |
| DA | Dopamine |
| ELS | Early life stress |
| EP | Epinephrine |
| GABA | γ-aminobutyric acid |
| Gln | Glutamine |
| Glu | Glutamate |
| GM | Gut microbiome |
| GCs | Glucocorticoids |
| GRs | Glucocorticoid receptors |
| 5-HT | Serotonin |
| 5-HTT | Serotonin transporter |
| HPA | Hypothalamic–pituitary–adrenal |
| IFN | Interferon |
| IL | Interleukin |
| Kyn | Kynurenine |
| LPS | Lipopolysaccharide |
| MDD | Major depressive disorder |
| MS | Maternally separated |
| MGB | Microbiome-gut–brain |
| NE | Norepinephrine |
| NF-κB | Nuclear factor kappa B |
| PVN | Paraventricular nucleus |
| PTSD | Post-traumatic stress disorder |
| PFC | Prefrontal cortex |
| PPD | Prenatal psychological distress |
| ROI | Recurrence of illness index |
| SCFAs | Short-chain fatty acids |
| Trp | Tryptophan |
References
- Shin, S.H.; Kim, Y.K. Early life stress, neuroinflammation, and psychiatric illness of adulthood. Adv. Exp. Med. Biol. 2023, 1411, 105–134. [Google Scholar] [CrossRef] [PubMed]
- Boullier, M.; Blair, M. Adverse childhood experiences. Paediatr. Child Health 2018, 28, 132–137. [Google Scholar] [CrossRef]
- Alhowaymel, F.; Kalmakis, K.; Jacelon, C. Developing the concept of adverse childhood experiences: A global perspective. J. Pediatr. Nurs. 2021, 56, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Merrick, M.T.; Ford, D.C.; Ports, K.A.; Guinn, A.S. Prevalence of adverse childhood experiences from the 2011–2014 behavioral risk factor surveillance system in 23 States. JAMA Pediatr. 2018, 172, 1038–1044. [Google Scholar] [CrossRef]
- Merrick, M.T.; Ports, K.A.; Ford, D.C.; Afifi, T.O.; Gershoff, E.T.; Grogan-Kaylor, A. Unpacking the impact of adverse childhood experiences on adult mental health. Child Abuse Negl. 2017, 69, 10–19. [Google Scholar] [CrossRef]
- Bishop, M.; Rosenstein, D.; Bakelaar, S.; Seedat, S. An analysis of early developmental trauma in social anxiety disorder and posttraumatic stress disorder. Ann. Gen. Psychiatry 2014, 13, 16. [Google Scholar] [CrossRef]
- Chi, X.; Jiang, W.; Guo, T.; Hall, D.L.; Luberto, C.M.; Zou, L. Relationship between adverse childhood experiences and anxiety symptoms among Chinese adolescents: The role of self-compassion and social support. Curr. Psychol. 2023, 142, 12822–12834. [Google Scholar] [CrossRef] [PubMed]
- Elmore, A.L.; Crouch, E. The association of adverse childhood experiences with anxiety and depression for children and youth, 8 to 17 years of age. Acad. Pediatr. 2020, 20, 600–608. [Google Scholar] [CrossRef]
- Lew, D.; Xian, H. Identifying distinct latent classes of adverse childhood experiences among US children and their relationship with childhood internalizing disorders. Child Psychiatry Hum. Dev. 2019, 50, 668–680. [Google Scholar] [CrossRef]
- Dye, H. The impact and long-term effects of childhood trauma. J. Hum. Behav. Soc. Environ. 2018, 28, 381–392. [Google Scholar] [CrossRef]
- Lowry, E.; McInerney, A.; Schmitz, N.; Deschênes, S.S. Adverse childhood experiences and cognitive function in adulthood: Examining the roles of depressive symptoms and inflammation in a prospective cohort study. Soc. Psychiatry Psychiatr. Epidemiol. 2022, 57, 2367–2377. [Google Scholar] [CrossRef] [PubMed]
- Dannehl, K.; Rief, W.; Euteneuer, F. Childhood adversity and cognitive functioning in patients with major depression. Child Abuse Negl. 2017, 70, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Gould, F.; Clarke, J.; Heim, C.; Harvey, P.D.; Majer, M.; Nemeroff, C.B. The effects of child abuse and neglect on cognitive functioning in adulthood. J. Psychiatr. Res. 2012, 46, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Majer, M.; Nater, U.M.; Lin, J.M.; Capuron, L.; Reeves, W.C. Association of childhood trauma with cognitive function in healthy adults: A pilot study. BMC Neurol. 2010, 10, 61. [Google Scholar] [CrossRef]
- Lee, M.; Park, K.H. Overgeneral memory in depression: Differences in with or without history of trauma, negative mood, and functional impairment. J. Korean Assn. Learn.-Cent. Curric. Instr. 2021, 21, 403–417. [Google Scholar] [CrossRef]
- Nemeroff, C.B. Neurobiological consequences of childhood trauma. J. Clin. Psychiatry 2004, 65, 18–28. [Google Scholar]
- Ho, T.C.; King, L.S. Mechanisms of neuroplasticity linking early adversity to depression: Developmental considerations. Transl. Psychiatry 2021, 11, 517. [Google Scholar] [CrossRef]
- Enoch, M.A. The role of early life stress as a predictor for alcohol and drug dependence. Psychopharmacology 2011, 214, 17–31. [Google Scholar] [CrossRef]
- Gilbert, R.; Widom, C.S.; Browne, K.; Fergusson, D.; Webb, E.; Janson, S. Burden and consequences of child maltreatment in high-income countries. Lancet 2009, 373, 68–81. [Google Scholar] [CrossRef]
- McLaughlin, K.A.; Greif Green, J.; Gruber, M.J.; Sampson, N.A.; Zaslavsky, A.M.; Kessler, R.C. Childhood adversities and first onset of psychiatric disorders in a national sample of US adolescents. Arch. Gen. Psychiatry 2012, 69, 1151–1160. [Google Scholar] [CrossRef]
- Integrative HMP (iHMP) Research Network Consortium. The integrative human microbiome project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103. [Google Scholar] [CrossRef]
- Salvucci, E. Microbiome, holobiont and the net of life. Crit. Rev. Microbiol. 2016, 42, 485–494. [Google Scholar] [CrossRef]
- Rajilić-Stojanović, M.; de Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Noh, H.; Jang, H.H.; Kim, G.; Zouiouich, S.; Cho, S.Y.; Kim, H.J.; Kim, J.; Choe, J.S.; Gunter, M.J.; Ferrari, P.; et al. Taxonomic composition and diversity of the gut microbiota in relation to habitual dietary intake in Korean adults. Nutrients 2021, 13, 366. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Morales, P.; Orellana, C.A.; Moutafis, G.; Moonen, G.; Rincon, G.; Nielsen, L.K.; Marcellin, E. Revisiting the evolution and taxonomy of Clostridia, a phylogenomic update. Genome Biol. Evol. 2019, 11, 2035–2044. [Google Scholar] [CrossRef]
- Raethong, N.; Nakphaichit, M.; Suratannon, N.; Sathitkowitchai, W.; Weerapakorn, W.; Keawsompong, S.; Vongsangnak, W. Analysis of human gut microbiome: Taxonomy and metabolic functions in Thai adults. Genes 2021, 12, 331. [Google Scholar] [CrossRef]
- Chabé, M.; Lokmer, A.; Ségurel, L. Gut protozoa: Friends or foes of the human gut microbiota? Trends Parasitol. 2017, 33, 925–934. [Google Scholar] [CrossRef]
- Kim, J.Y.; Whon, T.W.; Lim, M.Y.; Kim, Y.B.; Kim, N.; Kwon, M.S.; Kim, J.; Lee, S.H.; Choi, H.J.; Nam, I.H.; et al. The human gut archaeome: Identification of diverse haloarchaea in Korean subjects. Microbiome 2020, 8, 114. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Pargin, E.; Roach, M.J.; Skye, A.; Papudeshi, B.; Inglis, L.K.; Mallawaarachchi, V.; Grigson, S.R.; Harker, C.; Edwards, R.A.; Giles, S.K. The human gut virome: Composition, colonization, interactions, and impacts on human health. Front. Microbiol. 2023, 14, 963173. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. An updated overview on the relationship between human gut microbiome dysbiosis and psychiatric and psychological disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2024, 128, 110861. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. Neurodevelopmental disorders associated with gut microbiome dysbiosis in children. Children 2024, 11, 796. [Google Scholar] [CrossRef]
- Adamek, K.; Skonieczna-Żydecka, K.; Węgrzyn, D.; Łoniewska, B. Prenatal and early childhood development of gut microbiota. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 9667–9680. [Google Scholar] [CrossRef] [PubMed]
- Borrego-Ruiz, A.; Borrego, J.J. Human gut microbiome, diet, and mental disorders. Int. Microbiol. 2025, 28, 1–15. [Google Scholar] [CrossRef]
- Nishijima, S.; Suda, W.; Oshima, K.; Kim, S.W.; Hirose, Y.; Morita, H.; Hattori, M. The gut microbiome of healthy Japanese and its microbial and functional uniqueness. DNA Res. 2016, 23, 125–133. [Google Scholar] [CrossRef]
- Pechtel, P.; Pizzagalli, D.A. Effects of early life stress on cognitive and affective function: An integrated review of human literature. Psychopharmacology 2011, 214, 55–70. [Google Scholar] [CrossRef]
- Smith, K.E.; Pollak, S.D. Early life stress and development: Potential mechanisms for adverse outcomes. J. Neurodev. Disord. 2020, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. The resilient brain: Epigenetics, stress and the lifecourse. Psychoneuroendocrinology 2017, 83, 76. [Google Scholar] [CrossRef]
- Agorastos, A.; Pervanidou, P.; Chrousos, G.P.; Kolaitis, G. Early life stress and trauma: Developmental neuroendocrine aspects of prolonged stress system dysregulation. Hormones 2018, 17, 507–520. [Google Scholar] [CrossRef] [PubMed]
- LeMoult, J.; Humphreys, K.L.; Tracy, A.; Hoffmeister, J.A.; Ip, E.; Gotlib, I.H. Meta-analysis: Exposure to early life stress and risk for depression in childhood and adolescence. J. Am. Acad. Child Adolesc. Psychiatry 2020, 59, 842–855. [Google Scholar] [CrossRef] [PubMed]
- Nanni, V.; Uher, R.; Danese, A. Childhood maltreatment predicts unfavorable course of illness and treatment outcome in depression: A meta-analysis. Am. J. Psychiatry 2012, 169, 141–151. [Google Scholar] [CrossRef]
- Pervanidou, P.; Makris, G.; Chrousos, G.; Agorastos, A. Early life stress and pediatric posttraumatic stress disorder. Brain Sci. 2020, 10, 169. [Google Scholar] [CrossRef]
- Senner, F.; Schneider-Axmann, T.; Kaurani, L.; Zimmermann, J.; Wiltfang, J.; von Hagen, M.; Vogl, T.; Spitzer, C.; Senner, S.; Schulte, E.C.; et al. Association of early life stress and cognitive performance in patients with schizophrenia and healthy controls. Schizophr. Res. Cogn. 2023, 32, 100280. [Google Scholar] [CrossRef]
- Zatti, C.; Rosa, V.; Barros, A.; Valdivia, L.; Calegaro, V.C.; Freitas, L.H.; Ceresér, K.M.M.; da Rocha, N.S.; Bastos, A.G.; Schuch, F.B. Childhood trauma and suicide attempt: A meta-analysis of longitudinal studies from the last decade. Psychiatry Res. 2017, 256, 353–358. [Google Scholar] [CrossRef]
- Khoury, L.; Tang, Y.L.; Bradley, B.; Cubells, J.F.; Ressler, K.J. Substance use, childhood traumatic experience, and posttraumatic stress disorder in an urban civilian population. Depress. Anxiety 2010, 27, 1077–1086. [Google Scholar] [CrossRef]
- Makris, G.; Agorastos, A.; Chrousos, G.P.; Pervanidou, P. Stress system activation in children and adolescents with autism spectrum disorder. Front. Neurosci. 2022, 15, 756628. [Google Scholar] [CrossRef]
- Haykin, H.; Rolls, A. The neuroimmune response during stress: A physiological perspective. Immunity 2021, 54, 1933–1947. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, G.; Liu, Y. The molecular physiology of CRH neurons. Front. Neuroendocrinol. 2012, 33, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.H.; Abdulai-Saiku, S.; Vyas, A. Arginine vasopressin in the medial amygdala causes greater post-stress recruitment of hypothalamic vasopressin neurons. Mol. Brain 2021, 14, 141. [Google Scholar] [CrossRef] [PubMed]
- Cawley, N.X.; Li, Z.; Loh, Y.P. 60 YEARS OF POMC: Biosynthesis, trafficking, and secretion of pro-opiomelanocortin-derived peptides. J. Mol. Endocrinol. 2016, 56, T77–T97. [Google Scholar] [CrossRef]
- Atzori, M.; Cuevas-Olguin, R.; Esquivel-Rendon, E.; Garcia-Oscos, F.; Salgado-Delgado, R.C.; Saderi, N.; Miranda-Morales, M.; Treviño, M.; Pineda, J.C.; Salgado, H. Locus ceruleus norepinephrine release: A central regulator of CNS spatio-temporal activation? Front. Synaptic Neurosci. 2016, 8, 25. [Google Scholar] [CrossRef]
- Chrousos, G.P. Stress and disorders of the stress system. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef]
- Bucci, M.; Marques, S.S.; Oh, D.; Harris, N.B. Toxic stress in children and adolescents. Adv. Pediatr. 2016, 63, 403–428. [Google Scholar] [CrossRef]
- Charmandari, E.; Tsigos, C.; Chrousos, G. Endocrinology of the stress response. Annu. Rev. Physiol. 2005, 67, 259–284. [Google Scholar] [CrossRef]
- Gold, P.W.; Gabry, K.E.; Yasuda, M.R.; Chrousos, G.P. Divergent endocrine abnormalities in melancholic and atypical depression: Clinical and pathophysiologic implications. Endocrinol. Metab. Clin. North Am. 2002, 31, 37–62. [Google Scholar] [CrossRef] [PubMed]
- Lähdepuro, A.; Savolainen, K.; Lahti-Pulkkinen, M.; Eriksson, J.G.; Lahti, J.; Tuovinen, S.; Kajantie, E.; Pesonen, A.K.; Heinonen, K.; Räikkönen, K. The impact of early life stress on anxiety symptoms in late adulthood. Sci. Rep. 2019, 9, 4395. [Google Scholar] [CrossRef]
- Eichenbaum, H. Prefrontal–hippocampal interactions in episodic memory. Nat. Rev. Neurosci. 2017, 18, 547–558. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Morrison, J.H. The brain on stress: Vulnerability and plasticity of the prefrontal cortex over the life course. Neuron 2013, 79, 16–29. [Google Scholar] [CrossRef]
- Monroy, E.; Hernández-Torres, E.; Flores, G. Maternal separation disrupts dendritic morphology of neurons in prefrontal cortex, hippocampus, and nucleus accumbens in male rat offspring. J. Chem. Neuroanat. 2010, 40, 93–101. [Google Scholar] [CrossRef]
- Danielewicz, J.; Hess, G. Early life stress alters synaptic modification range in the rat lateral amygdala. Behav. Brain Res. 2014, 265, 32–37. [Google Scholar] [CrossRef]
- Malter Cohen, M.; Jing, D.; Yang, R.R.; Tottenham, N.; Lee, F.S.; Casey, B.J. Early-life stress has persistent effects on amygdala function and development in mice and humans. Proc. Natl. Acad. Sci. USA 2013, 110, 18274–18278. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E.; Way, B.M.; Seeman, T.E. Early adversity and adult health outcomes. Dev. Psychopathol. 2011, 23, 939–954. [Google Scholar] [CrossRef]
- Berman, A.K.; Lott, R.B.; Donaldson, S.T. Periodic maternal deprivation may modulate offspring anxiety-like behavior through mechanisms involving neuroplasticity in the amygdala. Brain Res. Bull. 2014, 101, 7–11. [Google Scholar] [CrossRef]
- Ishikawa, J.; Nishimura, R.; Ishikawa, A. Early-life stress induces anxiety-like behaviors and activity imbalances in the medial prefrontal cortex and amygdala in adult rats. Eur. J. Neurosci. 2015, 41, 442–453. [Google Scholar] [CrossRef]
- Wei, L.; David, A.; Duman, R.S.; Anisman, H.; Kaffman, A. Early life stress increases anxiety-like behavior in Balbc mice despite a compensatory increase in levels of postnatal maternal care. Horm. Behav. 2010, 57, 396–404. [Google Scholar] [CrossRef]
- Callaghan, B.L.; Richardson, R. Maternal separation results in early emergence of adult-like fear and extinction learning in infant rats. Behav. Neurosci. 2011, 125, 20–28. [Google Scholar] [CrossRef]
- Oomen, C.A.; Soeters, H.; Audureau, N.; Vermunt, L.; Van Hasselt, F.N.; Manders, E.M.M.; Joëls, M.; Lucassen, P.J.; Krugers, H. Severe early life stress hampers spatial learning and neurogenesis, but improves hippocampal synaptic plasticity and emotional learning under high-stress conditions in adulthood. J. Neurosci. 2010, 30, 6635–6645. [Google Scholar] [CrossRef]
- Finsterwald, C.; Alberini, C.M. Stress and glucocorticoid receptor-dependent mechanisms in long-term memory: From adaptive responses to psychopathologies. Neurobiol. Learn. Mem. 2014, 112, 17–29. [Google Scholar] [CrossRef]
- Ivy, A.S.; Rex, C.S.; Chen, Y.; Dubé, C.; Maras, P.M.; Grigoriadis, D.E.; Gall, C.M.; Lynch, G.; Baram, T.Z. Hippocampal dysfunction and cognitive impairments provoked by chronic early-life stress involve excessive activation of CRH receptors. J. Neurosci. 2010, 30, 13005–13015. [Google Scholar] [CrossRef]
- Hanson, J.L.; Nacewicz, B.M.; Sutterer, M.J.; Cayo, A.A.; Schaefer, S.M.; Rudolph, K.D.; Shirtcliff, E.A.; Pollak, S.D.; Davidson, R.J. Behavioral problems after early life stress: Contributions of the hippocampus and amygdala. Biol. Psychiatry 2015, 77, 314–323. [Google Scholar] [CrossRef]
- Teicher, M.H.; Anderson, C.M.; Ohashi, K.; Khan, A.; McGreenery, C.E.; Bolger, E.A.; Rohan, M.L.; Vitaliano, G.D. Differential effects of childhood neglect and abuse during sensitive exposure periods on male and female hippocampus. Neuroimage 2018, 169, 443–452. [Google Scholar] [CrossRef]
- Chen, M.C.; Hamilton, J.P.; Gotlib, I.H. Decreased hippocampal volume in healthy girls at risk of depression. Arch. Gen. Psychiatry 2010, 67, 270–276. [Google Scholar] [CrossRef]
- Gorka, A.X.; Hanson, J.L.; Radtke, S.R.; Hariri, A.R. Reduced hippocampal and medial prefrontal gray matter mediate the association between reported childhood maltreatment and trait anxiety in adulthood and predict sensitivity to future life stress. Biol. Mood Anxiety Disord. 2014, 4, 12. [Google Scholar] [CrossRef]
- Hanson, J.L.; van den Bos, W.; Roeber, B.J.; Rudolph, K.D.; Davidson, R.J.; Pollak, S.D. Early adversity and learning: Implications for typical and atypical behavioral development. J. Child Psychol. Psychiatry 2017, 58, 770–778. [Google Scholar] [CrossRef]
- Woon, F.L.; Hedges, D.W. Hippocampal and amygdala volumes in children and adults with childhood maltreatment-related posttraumatic stress disorder: A meta-analysis. Hippocampus 2008, 18, 729–736. [Google Scholar] [CrossRef]
- Gee, D.G.; Gabard-Durnam, L.J.; Flannery, J.; Goff, B.; Humphreys, K.L.; Telzer, E.H.; Hare, T.A.; Bookheimer, S.Y.; Tottenham, N. Early developmental emergence of human amygdala-prefrontal connectivity after maternal deprivation. Proc. Natl. Acad. Sci. USA 2013, 110, 15638–15643. [Google Scholar] [CrossRef]
- VanTieghem, M.R.; Tottenham, N. Neurobiological programming of early life stress: Functional development of amygdala prefrontal circuitry and vulnerability for stress related psychopathology. Curr. Top. Behav. Neurosci. 2018, 38, 117–136. [Google Scholar] [CrossRef]
- Wolf, R.C.; Herringa, R.J. Prefrontal–amygdala dysregulation to threat in pediatric posttraumatic stress disorder. Neuropsychopharmacology 2016, 41, 822–831. [Google Scholar] [CrossRef]
- Jedd, K.; Hunt, R.H.; Cicchetti, D.; Hunt, E.; Cowell, R.A.; Rogosch, F.A.; Toth, S.L.; Thomas, K.M. Long-term consequences of childhood maltreatment: Altered amygdala functional connectivity. Dev. Psychopathol. 2015, 27, 1577–1589. [Google Scholar] [CrossRef]
- Kim, P.; Evans, G.W.; Angstadt, M.; Ho, S.S.; Sripada, C.S.; Swain, J.E.; Liberzon, I.; Phan, K.L. Effects of childhood poverty and chronic stress on emotion regulatory brain function in adulthood. Proc. Natl. Acad. Sci. USA 2013, 110, 18442–18447. [Google Scholar] [CrossRef]
- Koss, K.J.; Gunnar, M.R. Annual Research Review: Early adversity, the hypothalamic-pituitary-adrenocortical axis, and child psychopathology. J. Child Psychol. Psychiatry 2017, 59, 327–346. [Google Scholar] [CrossRef]
- Pagliaccio, D.; Luby, J.L.; Bogdan, R.; Agrawal, A.; Gaffrey, M.S.; Belden, A.C.; Botteron, K.N.; Harms, M.P.; Barch, D.M. Amygdala functional connectivity, HPA axis genetic variation, and life stress in children and relations to anxiety and emotion regulation. J. Abnorm. Psychol. 2015, 124, 817–833. [Google Scholar] [CrossRef]
- Heim, C.M.; Binder, E.B. Current research trends in early life stress and depression: Review of human studies on sensitive periods, gene-environment interactions, and epigenetics. Exp. Neurol. 2012, 233, 102–111. [Google Scholar] [CrossRef]
- Turecki, G.; Meaney, M.J. Effects of the social environment and stress on glucocorticoid receptor gene methylation: A systematic review. Biol. Psychiatry 2016, 79, 87–96. [Google Scholar] [CrossRef]
- McEwen, B.S. Neurobiological and systemic effects of chronic stress. Chronic Stress 2017, 1, 2470547017692328. [Google Scholar] [CrossRef]
- McEwen, C.A.; McEwen, B.S. Social structure, adversity, toxic stress, and intergenerational poverty: An early childhood model. Annu. Rev. Sociol. 2017, 43, 445–472. [Google Scholar] [CrossRef]
- Birn, R.M.; Roeber, B.J.; Pollak, S.D. Early childhood stress exposure, reward pathways, and adult decision making. Proc. Natl. Acad. Sci. USA 2017, 114, 13549–13554. [Google Scholar] [CrossRef]
- Novick, A.M.; Levandowski, M.L.; Laumann, L.E.; Philip, N.S.; Price, L.H.; Tyrka, A.R. The effects of early life stress on reward processing. J. Psychiatr. Res. 2018, 101, 80–103. [Google Scholar] [CrossRef]
- Risbrough, V.B.; Glynn, L.M.; Davis, E.P.; Sandman, C.A.; Obenaus, A.; Stern, H.S.; Keator, D.B.; Yassa, M.A.; Baram, T.Z.; Baker, D.G. Does anhedonia presage increased risk of posttraumatic stress disorder?: Adolescent anhedonia and posttraumatic disorders. In Behavioral Neurobiology of PTSD; Current Topics in Behavioral Neurosciences; Springer: Cham, Switzerland, 2018; Volume 38, pp. 249–265. [Google Scholar] [CrossRef]
- Chocyk, A.; Dudys, D.; Przyborowska, A.; Majcher, I.; Maćkowiak, M.; Wędzony, K. Maternal separation affects the number, proliferation and apoptosis of glia cells in the substantia nigra and ventral tegmental area of juvenile rats. Neuroscience 2011, 173, 1–18. [Google Scholar] [CrossRef]
- Yan, C.G.; Rincón-Cortés, M.; Raineki, C.; Sarro, E.; Colcombe, S.; Guilfoyle, D.N.; Yang, Z.; Gerum, S.; Biswal, B.B.; Milham, M.P.; et al. Aberrant development of intrinsic brain activity in a rat model of caregiver maltreatment of offspring. Transl. Psychiatry 2017, 7, e1005. [Google Scholar] [CrossRef]
- Peña, C.J.; Neugut, Y.D.; Calarco, C.A.; Champagne, F.A. Effects of maternal care on the development of midbrain dopamine pathways and reward-directed behavior in female offspring. Eur. J. Neurosci. 2014, 39, 946–956. [Google Scholar] [CrossRef]
- Rodrigues, A.J.; Leão, P.; Carvalho, M.; Almeida, O.F.X.; Sousa, N. Potential programming of dopaminergic circuits by early life stress. Psychopharmacology 2011, 214, 107–120. [Google Scholar] [CrossRef]
- Dennison, M.J.; Rosen, M.L.; Sambrook, K.A.; Jenness, J.L.; Sheridan, M.A.; McLaughlin, K.A. Differential associations of distinct forms of childhood adversity with neurobehavioral measures of reward processing: A developmental pathway to depression. Child Dev. 2017, 90, e96–e113. [Google Scholar] [CrossRef]
- Marusak, H.A.; Hatfield, J.R.; Thomason, M.E.; Rabinak, C.A. Reduced ventral tegmental area-hippocampal connectivity in children and adolescents exposed to early threat. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 130–137. [Google Scholar] [CrossRef]
- Corral-Frías, N.S.; Nikolova, Y.S.; Michalski, L.J.; Baranger, D.A.A.; Hariri, A.R.; Bogdan, R. Stress-related anhedonia is associated with ventral striatum reactivity to reward and transdiagnostic psychiatric symptomatology. Psychol. Med. 2015, 45, 2605–2617. [Google Scholar] [CrossRef]
- Goff, B.; Gee, D.G.; Telzer, E.H.; Humphreys, K.L.; Gabard-Durnam, L.; Flannery, J.; Tottenham, N. Reduced nucleus accumbens reactivity and adolescent depression following early-life stress. Neuroscience 2013, 249, 129–138. [Google Scholar] [CrossRef]
- Kamkar, N.H.; Lewis, D.J.; van den Bos, W.; Morton, J.B. Ventral striatal activity links adversity and reward processing in children. Dev. Cogn. Neurosci. 2017, 26, 20–27. [Google Scholar] [CrossRef]
- Ledoux, J.E.; Daw, N.D. Surviving threats: Neural circuit and computational implications of a new taxonomy of defensive behaviour. Nat. Rev. Neurosci. 2018, 19, 269–282. [Google Scholar] [CrossRef]
- Nelson, C.A.; Scott, R.D.; Bhutta, Z.A.; Harris, N.B.; Danese, A.; Samara, M. Adversity in childhood is linked to mental and physical health throughout life. BMJ 2020, 371, m3048. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The first microbial colonizers of the human gut: Composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Cong, X.; Xu, W.; Janton, S.; Henderson, W.A.; Matson, A.; McGrath, J.M.; Maas, K.; Graf, J. Gut microbiome developmental patterns in early life of preterm infants: Impacts of feeding and gender. PLoS ONE 2016, 11, e0152751. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef]
- Morreale, C.; Giaroni, C.; Baj, A.; Folgori, L.; Barcellini, L.; Dhami, A.; Agosti, M.; Bresesti, I. Effects of perinatal antibiotic exposure and neonatal gut microbiota. Antibiotics 2023, 12, 258. [Google Scholar] [CrossRef]
- Arboleya, S.; Binetti, A.; Salazar, N.; Fernández, N.; Solís, G.; Hernandez-Barranco, A.; Margolles, A.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 2012, 79, 763–772. [Google Scholar] [CrossRef]
- Moles, L.; Gómez, M.; Heilig, H.; Bustos, G.; Fuentes, S.; de Vos, W.; Fernández, L.; Rodríguez, J.M.; Jiménez, E. Bacterial diversity in meconium of preterm neonates and evolution of their fecal microbiota during the first month of life. PLoS ONE 2013, 8, e66986. [Google Scholar] [CrossRef]
- Pantazi, A.C.; Balasa, A.L.; Mihai, C.M.; Chisnoiu, T.; Lupu, V.V.; Kassim, M.A.K.; Mihai, L.; Frecus, C.E.; Chirila, S.I.; Lupu, A.; et al. Development of gut microbiota in the first 1000 days after birth and potential interventions. Nutrients 2023, 15, 3647. [Google Scholar] [CrossRef]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O’Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K.; et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 4. [Google Scholar] [CrossRef]
- Madan, J.C.; Hoen, A.G.; Lundgren, S.N.; Farzan, S.F.; Cottingham, K.L.; Morrison, H.G.; Sogin, M.L.; Li, H.; Moore, J.H.; Karagas, M.R. Association of cesarean delivery and formula supplementation with the intestinal microbiome of 6-week-old infants. JAMA Pediatr. 2016, 170, 212–219. [Google Scholar] [CrossRef]
- Hansen, R.; Scott, K.P.; Khan, S.; Martin, J.C.; Berry, S.H.; Stevenson, M.; Okpapi, A.; Munro, M.J.; Hold, G.L. First-pass meconium samples from healthy term vaginally-delivered neonates: An analysis of the microbiota. PLoS ONE 2015, 10, e0133320. [Google Scholar] [CrossRef]
- Shao, Y.; Forster, S.C.; Tsaliki, E.; Vervier, K.; Strang, A.; Simpson, N.; Kumar, N.; Stares, M.D.; Rodger, A.; Brocklehurst, P.; et al. Stunted microbiota and opportunistic pathogen colonization in Caesarean-section birth. Nature 2019, 574, 117–121. [Google Scholar] [CrossRef]
- Coscia, A.; Bardanzellu, F.; Caboni, E.; Fanos, V.; Peroni, D.G. When a neonate is born, so is a microbiota. Life 2021, 11, 148. [Google Scholar] [CrossRef]
- Karlsson, C.L.; Molin, G.; Cilio, C.M.; Ahrné, S. The pioneer gut microbiota in human neonates vaginally born at term—A pilot study. Pediatr. Res. 2011, 70, 282–286. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Ye, Y.; Wang, F.; Chen, F.; Zheng, C. The role of microbiota in infant health: From early life to adulthood. Front. Immunol. 2021, 12, 708472. [Google Scholar] [CrossRef]
- Martin, R.; Makino, H.; Cetinyurek Yavuz, A.; Ben-Amor, K.; Roelofs, M.; Ishikawa, E.; Kubota, H.; Swinkels, S.; Sakai, T.; Oishi, K.; et al. Early-life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS ONE 2016, 11, e0158498. [Google Scholar] [CrossRef] [PubMed]
- Hesla, H.M.; Stenius, F.; Jäderlund, L.; Nelson, R.; Engstrand, L.; Alm, J.; Dicksved, J. Impact of lifestyle on the gut microbiota of healthy infants and their mothers—The ALADDIN birth cohort. FEMS Microbiol. Ecol. 2014, 90, 791–801. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, A.; Farver, M.; Smilowitz, J.T. The influence of early infant-feeding practices on the intestinal microbiome and body composition in infants. Nutr. Metab. Insights 2015, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schütte, U.M.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6, e21313. [Google Scholar] [CrossRef]
- Bezirtzoglou, E.; Tsiotsias, A.; Welling, G.W. Microbiota profile in feces of breast- and formula-fed newborns by using fluorescence in situ hybridization (FISH). Anaerobe 2011, 17, 478–482. [Google Scholar] [CrossRef]
- Yassour, M.; Vatanen, T.; Siljander, H.; Hämäläinen, A.M.; Härkönen, T.; Ryhänen, S.J.; Franzosa, E.A.; Vlamakis, H.; Huttenhower, C.; Gevers, D.; et al. Natural history of the infant gut microbiome and impact of antibiotic treatment on bacterial strain diversity and stability. Sci. Transl. Med. 2016, 8, 343ra81. [Google Scholar] [CrossRef]
- Schwartz, D.J.; Langdon, A.E.; Dantas, G. Understanding the impact of antibiotic perturbation on the human microbiome. Genome Med. 2020, 12, 82. [Google Scholar] [CrossRef]
- Fouhy, F.; Guinane, C.M.; Hussey, S.; Wall, R.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C.; et al. High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob. Agents Chemother. 2012, 56, 5811–5820. [Google Scholar] [CrossRef]
- Tanaka, S.; Kobayashi, T.; Songjinda, P.; Tateyama, A.; Tsubouchi, M.; Kiyohara, C.; Shirakawa, T.; Sonomoto, K.; Nakayama, J. Influence of antibiotic exposure in the early postnatal period on the development of intestinal microbiota. FEMS Immunol. Med. Microbiol. 2009, 56, 80–87. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Stiemsma, L.T.; Amenyogbe, N.; Brown, E.M.; Finlay, B. The intestinal microbiome in early life: Health and disease. Front. Immunol. 2014, 5, 427. [Google Scholar] [CrossRef]
- Slykerman, R.F.; Thompson, J.; Waldie, K.E.; Murphy, R.; Wall, C.; Mitchell, E.A. Antibiotics in the first year of life and subsequent neurocognitive outcomes. Acta Paediatr. 2017, 106, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Borre, Y.E.; O’Keeffe, G.W.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Microbiota and neurodevelopmental windows: Implications for brain disorders. Trends Mol. Med. 2014, 20, 509–518. [Google Scholar] [CrossRef]
- Damiani, F.; Cornuti, S.; Tognini, P. The gut-brain connection: Exploring the influence of the gut microbiota on neuroplasticity and neurodevelopmental disorders. Neuropharmacology 2023, 231, 109491. [Google Scholar] [CrossRef]
- Troller-Renfree, S.V.; Brito, N.H.; Desai, P.M.; Leon-Santos, A.G.; Wiltshire, C.A.; Motton, S.N.; Meyer, J.S.; Isler, J.; Fifer, W.P.; Noble, K.G. Infants of mothers with higher physiological stress show alterations in brain function. Dev. Sci. 2020, 23, e12976. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.L.; Fields, A.; Gee, D.G.; Gabard-Durnam, L.; Caldera, C.; Humphreys, K.L.; Goff, B.; Flannery, J.; Telzer, E.H.; Shapiro, M.; et al. Mind and gut: Associations between mood and gastrointestinal distress in children exposed to adversity. Dev. Psychopathol. 2020, 32, 309–328. [Google Scholar] [CrossRef]
- Vogel, S.C.; Brito, N.H.; Callaghan, B.L. Early life stress and the development of the infant gut microbiota: Implications for mental health and neurocognitive development. Curr. Psychiatry Rep. 2020, 22, 61. [Google Scholar] [CrossRef]
- Kemp, K.M.; Colson, J.; Lorenz, R.G.; Maynard, C.L.; Pollock, J.S. Early life stress in mice alters gut microbiota independent of maternal microbiota inheritance. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2021, 320, R663–R674. [Google Scholar] [CrossRef]
- Otaru, N.; Kourouma, L.; Pugin, B.; Constancias, F.; Braegger, C.; Mansuy, I.M.; Lacroix, C. Transgenerational effects of early life stress on the fecal microbiota in mice. Commun. Biol. 2024, 7, 670. [Google Scholar] [CrossRef] [PubMed]
- Tao, E.; Wu, Y.; Hu, C.; Zhu, Z.; Ye, D.; Long, G.; Chen, B.; Guo, R.; Shu, X.; Zheng, W.; et al. Early life stress induces irritable bowel syndrome from childhood to adulthood in mice. Front. Microbiol. 2023, 14, 1255525. [Google Scholar] [CrossRef]
- Jašarević, E.; Howard, C.D.; Misic, A.M.; Beiting, D.P.; Bale, T.L. Stress during pregnancy alters temporal and spatial dynamics of the maternal and offspring microbiome in a sex-specific manner. Sci. Rep. 2017, 7, 44182. [Google Scholar] [CrossRef]
- Amini-Khoei, H.; Haghani-Samani, E.; Beigi, M.; Soltani, A.; Mobini, G.R.; Balali-Dehkordi, S.; Haj-Mirzaian, A.; Rafieian-Kopaei, M.; Alizadeh, A.; Hojjati, M.R.; et al. On the role of corticosterone in behavioral disorders, microbiota composition alteration and neuroimmune response in adult male mice subjected to maternal separation stress. Int. Immunopharmacol. 2019, 66, 242–250. [Google Scholar] [CrossRef]
- Pusceddu, M.M.; El Aidy, S.; Crispie, F.; O’Sullivan, O.; Cotter, P.; Stanton, C.; Kelly, P.; Cryan, J.F.; Dinan, T.G. N-3 polyunsaturated fatty acids (PUFAs) reverse the impact of early-life stress on the gut microbiota. PLoS ONE 2015, 10, e0139721. [Google Scholar] [CrossRef]
- Kuti, D.; Winkler, Z.; Horváth, K.; Juhász, B.; Paholcsek, M.; Stágel, A.; Gulyás, G.; Czeglédi, L.; Ferenczi, S.; Kovács, K.J. Gastrointestinal (non-systemic) antibiotic rifaximin differentially affects chronic stress-induced changes in colon microbiome and gut permeability without effect on behavior. Brain Behav. Immun. 2020, 84, 218–228. [Google Scholar] [CrossRef]
- Rincel, M.; Aubert, P.; Chevalier, J.; Grohard, P.A.; Basso, L.; de Oliveira, C.M.; Helbling, J.C.; Lévy, É.; Chevalier, G.; Leboyer, M.; et al. Multi-hit early life adversity affects gut microbiota, brain and behavior in a sex-dependent manner. Brain Behav. Immun. 2019, 80, 179–192. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, S.A.; Kang, W.S.; Kim, J.W. Early-life stress modulates gut microbiota and peripheral and central inflammation in a sex-dependent manner. Int. J. Mol. Sci. 2021, 22, 1899. [Google Scholar] [CrossRef] [PubMed]
- El Aidy, S.; Ramsteijn, A.S.; Dini-Andreote, F.; van Eijk, R.; Houwing, D.J.; Salles, J.F.; Olivier, J.D.A. Serotonin transporter genotype modulates the gut microbiota composition in young rats, an effect augmented by early life stress. Front. Cell. Neurosci. 2017, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Ke, S.; Hartmann, J.; Ressler, K.J.; Liu, Y.Y.; Koenen, K.C. The emerging role of the gut microbiome in posttraumatic stress disorder. Brain Behav. Immun. 2023, 114, 360–370. [Google Scholar] [CrossRef]
- Yadav, S.K.; Ahmad, R.; Moshfegh, C.M.; Sankarasubramanian, J.; Joshi, V.; Elkhatib, S.K.; Chhonker, Y.S.; Murry, D.J.; Talmon, G.A.; Guda, C.; et al. Repeated social defeat stress induces an inflammatory gut milieu by altering the mucosal barrier integrity and gut microbiota homeostasis. Biol. Psychiatry Glob. Open Sci. 2023, 3, 824–836. [Google Scholar] [CrossRef]
- Aatsinki, A.K.; Keskitalo, A.; Laitinen, V.; Munukka, E.; Uusitupa, H.M.; Lahti, L.; Kortesluoma, S.; Mustonen, P.; Rodrigues, A.J.; Coimbra, B.; et al. Maternal prenatal psychological distress and hair cortisol levels associate with infant fecal microbiota composition at 2.5 months of age. Psychoneuroendocrinology 2020, 119, 104754. [Google Scholar] [CrossRef]
- Zijlmans, M.A.; Korpela, K.; Riksen-Walraven, J.M.; de Vos, W.M.; de Weerth, C. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology 2015, 53, 233–245. [Google Scholar] [CrossRef]
- Jahnke, J.R.; Roach, J.; Azcarate-Peril, M.A.; Thompson, A.L. Maternal precarity and HPA axis functioning shape infant gut microbiota and HPA axis development in humans. PLoS ONE 2021, 16, e0251782. [Google Scholar] [CrossRef] [PubMed]
- Grant-Beurmann, S.; Jumare, J.; Ndembi, N.; Matthew, O.; Shutt, A.; Omoigberale, A.; Martin, O.A.; Fraser, C.M.; Charurat, M. Dynamics of the infant gut microbiota in the first 18 months of life: The impact of maternal HIV infection and breastfeeding. Microbiome 2022, 12, 61. [Google Scholar] [CrossRef]
- D’Agata, A.L.; Wu, J.; Welandawe, M.K.V.; Dutra, S.V.O.; Kane, B.; Groer, M.W. Effects of early life NICU stress on the developing gut microbiome. Dev. Psychobiol. 2019, 61, 650–660. [Google Scholar] [CrossRef]
- Hermes, G.D.A.; Eckermann, H.A.; de Vos, W.M.; de Weerth, C. Does entry to center-based childcare affect gut microbial colonization in young infants? Sci. Rep. 2020, 24, 10235. [Google Scholar] [CrossRef]
- Reid, B.M.; Horne, R.; Donzella, B.; Szamosi, J.C.; Coe, C.L.; Foster, J.A.; Gunnar, M.R. Microbiota-immune alterations in adolescents following early life adversity: A proof of concept study. Dev. Psychobiol. 2021, 63, 851–863. [Google Scholar] [CrossRef]
- Hemmings, S.M.J.; Malan-Müller, S.; van den Heuvel, L.L.; Demmitt, B.A.; Stanislawski, M.A.; Smith, D.G.; Bohr, A.D.; Stamper, C.E.; Hyde, E.R.; Morton, J.T.; et al. The microbiome in posttraumatic stress disorder and trauma-exposed controls: An exploratory study. Psychosom. Med. 2017, 79, 936–946. [Google Scholar] [CrossRef]
- Malan-Muller, S.; Valles-Colomer, M.; Foxx, C.L.; Vieira-Silva, S.; van den Heuvel, L.L.; Raes, J.; Seedat, S.; Lowry, C.A.; Hemmings, S.M.J. Exploring the relationship between the gut microbiome and mental health outcomes in a posttraumatic stress disorder cohort relative to trauma-exposed controls. Eur. Neuropsychopharmacol. 2022, 56, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Mulder, R.H.; Kraaij, R.; Schuurmans, I.K.; Frances-Cuesta, C.; Sanz, Y.; Medina-Gomez, C.; Duijts, L.; Rivadeneira, F.; Tiemeier, H.; Jaddoe, V.W.; et al. Early-life stress and the gut microbiome: A comprehensive population-based investigation. Brain Behav. Immun. 2024, 118, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Hantsoo, L.; Jašarević, E.; Criniti, S.; McGeehan, B.; Tanes, C.; Sammel, M.D.; Elovitz, M.A.; Compher, C.; Wu, G.; Epperson, C.N. Childhood adversity impact on gut microbiota and inflammatory response to stress during pregnancy. Brain Behav. Immun. 2019, 75, 240–250. [Google Scholar] [CrossRef]
- Michels, N.; Van de Wiele, T.; Fouhy, F.; O’Mahony, S.; Clarke, G.; Keane, J. Gut microbiome patterns depending on children’s psychosocial stress: Reports versus biomarkers. Brain Behav. Immun. 2019, 80, 751–762. [Google Scholar] [CrossRef]
- Flannery, J.E.; Stagaman, K.; Burns, A.R.; Hickey, R.J.; Roos, L.E.; Giuliano, R.J.; Fisher, P.A.; Sharpton, T.J. Gut feelings begin in childhood: The gut metagenome correlates with early environment, caregiving, and behavior. mBio 2020, 11, e02780-19. [Google Scholar] [CrossRef] [PubMed]
- Keskitalo, A.; Aatsinki, A.K.; Kortesluoma, S.; Pelto, J.; Korhonen, L.; Lahti, L.; Lukkarinen, M.; Munukka, E.; Karlsson, H.; Karlsson, L. Gut microbiota diversity but not composition is related to saliva cortisol stress response at the age of 2.5 months. Stress 2021, 24, 551–560. [Google Scholar] [CrossRef]
- Laue, H.E.; Karagas, M.R.; Coker, M.O.; Bellinger, D.C.; Baker, E.R.; Korrick, S.A.; Madan, J.C. Sex-specific relationships of the infant microbiome and early-childhood behavioral outcomes. Pediatr. Res. 2022, 92, 580–591. [Google Scholar] [CrossRef]
- Kraaij, R.; Schuurmans, I.K.; Radjabzadeh, D.; Tiemeier, H.; Dinan, T.G.; Uitterlinden, A.G.; Hillegers, M.; Jaddoe, V.W.; Duijts, L.; Moll, H.; et al. The gut microbiome and child mental health: A population-based study. Brain Behav. Immun. 2023, 108, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, M.C.; Verosky, B.; Chen, H.J.; Davis, O.; Gur, T.L. The maternal microbiome as a map to understanding the impact of prenatal stress on offspring psychiatric health. Biol. Psychiatry 2024, 95, 300–309. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Pessa-Morikawa, T.; Husso, A.; Kärkkäinen, O.; Koistinen, V.; Hanhineva, K.; Iivanainen, A.; Niku, M. Maternal microbiota-derived metabolic profile in fetal murine intestine, brain and placenta. BMC Microbiol. 2022, 22, 46. [Google Scholar] [CrossRef]
- Wu, M.; Tian, T.; Mao, Q.; Zou, T.; Zhou, C.; Xie, J.; Chen, J. Associations between disordered gut microbiota and changes of neurotransmitters and short-chain fatty acids in depressed mice. Transl. Psychiatry 2020, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: Microbial metabolites that alleviate stress-induced brain-gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef]
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and non-immune functions. Immunity 2021, 54, 2194–2208. [Google Scholar] [CrossRef]
- Thion, M.S.; Low, D.; Silvin, A.; Chen, J.; Grisel, P.; Schulte-Schrepping, J.; Blecher, R.; Ulas, T.; Squarzoni, P.; Hoeffel, G.; et al. Microbiome influences prenatal and adult microglia in a sex-specific manner. Cell 2018, 172, 500–516.e16. [Google Scholar] [CrossRef] [PubMed]
- Caetano-Silva, M.E.; Rund, L.; Hutchinson, N.T.; Woods, J.A.; Steelman, A.J.; Johnson, R.W. Inhibition of inflammatory microglia by dietary fiber and short-chain fatty acids. Sci. Rep. 2023, 13, 2819. [Google Scholar] [CrossRef]
- Erny, D.; Dokalis, N.; Mezö, C.; Castoldi, A.; Mossad, O.; Staszewski, O.; Frosch, M.; Villa, M.; Fuchs, V.; Mayer, A.; et al. Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab. 2021, 33, 2260–2276.e7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhong, Z.; Shi, L.; Huang, L.; Lin, C.; He, Y.; Xia, X.; Zhang, T.; Ding, W.; Yang, Y. Gut microbiota mediate early life stress-induced social dysfunction and anxiety-like behaviors by impairing amino acid transport at the gut. Gut Microbes 2024, 16, 2401939. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, I.C.; Yu, H.R.; Huang, L.T.; Tiao, M.M.; Tain, Y.L. Maternal tryptophan supplementation protects adult rat offspring against hypertension programmed by maternal chronic kidney disease: Implication of tryptophan-metabolizing microbiome and aryl hydrocarbon receptor. Int. J. Mol. Sci. 2020, 21, 4552. [Google Scholar] [CrossRef]
- Galley, J.D.; Chen, H.J.; Antonson, A.M.; Gur, T.L. Prenatal stress-induced disruptions in microbial and host tryptophan metabolism and transport. Behav. Brain Res. 2021, 414, 113471. [Google Scholar] [CrossRef]
- Fung, T.C.; Vuong, H.E.; Luna, C.D.G.; Pronovost, G.N.; Aleksandrova, A.A.; Riley, N.G.; Vavilina, A.; McGinn, J.; Rendon, T.; Forrest, L.R.; et al. Intestinal serotonin and fluoxetine exposure modulate bacterial colonization in the gut. Nat. Microbiol. 2019, 4, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Höglund, E.; Øverli, Ø.; Winberg, S. Tryptophan metabolic pathways and brain serotonergic activity: A comparative review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef]
- Steenbergen, L.; Jongkees, B.J.; Sellaro, R.; Colzato, L.S. Tryptophan supplementation modulates social behavior: A review. Neurosci. Biobehav. Rev. 2016, 64, 346–358. [Google Scholar] [CrossRef]
- Keane, J.M.; Khashan, A.S.; McCarthy, F.P.; Kenny, L.C.; Collins, J.M.; O’Donovan, S.; Brown, J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; et al. Identifying a biological signature of prenatal maternal stress. JCI Insight 2021, 6, e143007. [Google Scholar] [CrossRef]
- Chen, L.W.; Wang, S.T.; Wang, L.W.; Kao, Y.C.; Chu, C.L.; Wu, C.C.; Chiang, C.H.; Huang, C.C. Early neurodevelopmental trajectories for autism spectrum disorder in children born very preterm. Pediatrics 2020, 146, e20200297. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Feng, Z.; Cheng, H.W. Perspective: Gestational tryptophan fluctuation altering neuroembryogenesis and psychosocial development. Cells 2022, 11, 1270. [Google Scholar] [CrossRef] [PubMed]
- Cappadocia, M.C.; Weiss, J.A.; Pepler, D. Bullying experiences among children and youth with autism spectrum disorders. J. Autism Dev. Disord. 2012, 42, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R.M.; Fedina, C. Cyber bullying in ADHD and Asperger syndrome populations. Res. Autism Spectr. Disord. 2011, 5, 1201–1208. [Google Scholar] [CrossRef]
- Zinner, S.H.; Conelea, C.A.; Glew, G.M.; Woods, D.W.; Budman, C.L. Peer victimization in youth with Tourette syndrome and other chronic tic disorders. Child Psychiatry Hum. Dev. 2012, 43, 124–136. [Google Scholar] [CrossRef]
- Higazi, A.M.; Kamel, H.M.; Abdel-Naeem, E.A.; Abdullah, N.M.; Mahrous, D.M.; Osman, A.M. Expression analysis of selected genes involved in tryptophan metabolic pathways in Egyptian children with autism spectrum disorder and learning disabilities. Sci. Rep. 2021, 11, 6931. [Google Scholar] [CrossRef]
- Roussin, L.; Prince, N.; Perez-Pardo, P.; Kraneveld, A.D.; Rabot, S.; Naudon, L. Role of the gut microbiota in the pathophysiology of autism spectrum disorder: Clinical and preclinical evidence. Microorganisms 2020, 8, 1369. [Google Scholar] [CrossRef]
- Santana-Coelho, D. Does the kynurenine pathway play a pathogenic role in autism spectrum disorder? Brain Behav. Immun.-Health 2024, 40, 100839. [Google Scholar] [CrossRef]
- Plitman, E.; Iwata, Y.; Caravaggio, F.; Nakajima, S.; Chung, J.K.; Gerretsen, P.; Kim, J.; Takeuchi, H.; Chakravarty, M.M.; Remington, G.; et al. Kynurenic acid in schizophrenia: A systematic review and meta-analysis. Schizophr. Bull. 2017, 43, 764–777. [Google Scholar] [CrossRef]
- Oh, C.M.; Park, S.; Kim, H. Serotonin as a new therapeutic target for diabetes mellitus and obesity. Diabetes Metab. J. 2016, 40, 89–98. [Google Scholar] [CrossRef]
- Wei, G.Z.; Martin, K.A.; Xing, P.Y.; Agrawal, R.; Whiley, L.; Wood, T.K.; Hejndorf, S.; Ng, Y.Z.; Low, J.Z.Y.; Rossant, J.; et al. Tryptophan-metabolizing gut microbes regulate adult neurogenesis via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. USA 2021, 118, e2021091118. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Li, G.; Zheng, Q.; Gu, X.; Shi, Q.; Su, Y.; Chu, Q.; Yuan, X.; Bao, Z.; Lu, J.; et al. Tryptophan metabolism in health and disease. Cell Metab. 2023, 35, 1304–1326. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, S.; Ma, N.; Johnston, L.J.; Wu, C.; Ma, X. Metabolites of microbiota response to tryptophan and intestinal mucosal immunity: A therapeutic target to control intestinal inflammation. Med. Res. Rev. 2021, 41, 1061–1088. [Google Scholar] [CrossRef]
- Kaur, H.; Bose, C.; Mande, S.S. Tryptophan metabolism by gut microbiome and gut-brain-axis: An in silico analysis. Front. Neurosci. 2019, 13, 1365. [Google Scholar] [CrossRef]
- Roager, H.M.; Licht, T.R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 2018, 9, 3294. [Google Scholar] [CrossRef]
- Young, S.N. The effect of raising and lowering tryptophan levels on human mood and social behaviour. Philos. Trans. R. Soc. London B Biol. Sci. 2013, 368, 20110375. [Google Scholar] [CrossRef]
- Ansari, F.; Neshat, M.; Pourjafar, H.; Jafari, S.M.; Samakkhah, S.A.; Mirzakhani, E. The role of probiotics and prebiotics in modulating of the gut-brain axis. Front. Nutr. 2023, 10, 1173660. [Google Scholar] [CrossRef] [PubMed]
- Rusch, J.A.; Layden, B.T.; Dugas, L.R. Signalling cognition: The gut microbiota and hypothalamic-pituitary-adrenal axis. Front. Endocrinol. 2023, 14, 1130689. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Duran, N.L.; Olson, S.L.; Hajal, N.J.; Felt, B.T.; Vazquez, D.M. Hypothalamic pituitary adrenal axis functioning in reactive and proactive aggression in children. J. Abnorm. Child Psychol. 2009, 37, 169–182. [Google Scholar] [CrossRef]
- Buske-Kirschbaum, A.; Trikojat, K.; Tesch, F.; Schmitt, J.; Roessner, V.; Luksch, H.; Rösen-Wolff, A.; Plessow, F. Altered hypothalamus-pituitary-adrenal axis function: A relevant factor in the comorbidity of atopic eczema and attention deficit/hyperactivity disorder? Psychoneuroendocrinology 2019, 105, 178–186. [Google Scholar] [CrossRef]
- Menke, A. Is the HPA axis as target for depression outdated, or is there a new hope? Front. Psychiatry 2019, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Schalbroeck, R.; van Velden, F.H.P.; de Geus-Oei, L.F.; Yaqub, M.; van Amelsvoort, T.; Booij, J.; Selten, J.P. Striatal dopamine synthesis capacity in autism spectrum disorder and its relation with social defeat: An [18F]-FDOPA PET/CT study. Transl. Psychiatry 2021, 11, 47. [Google Scholar] [CrossRef]
- Lewis, R.G.; Florio, E.; Punzo, D.; Borrelli, E. The brain’s reward system in health and disease. In Circadian Clock in Brain Health and Disease; Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2021; Volume 1344, pp. 57–69. [Google Scholar] [CrossRef]
- Goldstein, D.S. Adrenal responses to stress. Cell. Mol. Neurobiol. 2010, 30, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Garcia-Olivares, J.; Candler, S.; Schwabe, S.; Maletic, V. New insights into the mechanism of action of viloxazine: Serotonin and norepinephrine modulating properties. J. Exp. Pharmacol. 2020, 12, 285–300. [Google Scholar] [CrossRef]
- Bacchini, D.; Affuso, G.; Trotta, T. Temperament, ADHD and peer relations among schoolchildren: The mediating role of school bullying. Aggress. Behav. 2008, 34, 447–459. [Google Scholar] [CrossRef]
- Stenseng, F.; Skalická, V.; Skaug, S.S.; Belsky, J.; Wichstrøm, L. Attention-deficit hyperactivity disorder symptoms and bullying victimization from childhood to adolescence—A within-person cross-lagged approach. Dev. Psychopathol. 2024; 1–11, Advance online publication. [Google Scholar] [CrossRef]
- Dalley, J.W.; Roiser, J.P. Dopamine, serotonin and impulsivity. Neuroscience 2012, 215, 42–58. [Google Scholar] [CrossRef]
- Seo, D.; Patrick, C.J.; Kennealy, P.J. Role of serotonin and dopamine system interactions in the neurobiology of impulsive aggression and its comorbidity with other clinical disorders. Aggress. Violent Behav. 2008, 13, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, A.; Lingrand, L.; Maillard, M.; Feuz, B.; Tompkins, T.A. The effects of psychobiotics on the microbiota-gut-brain axis in early-life stress and neuropsychiatric disorders. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 105, 110142. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Borrego, J.J. Psychobiotics: A new perspective on the treatment of stress, anxiety, and depression. Anxiety Stress 2024, 30, 79–93. [Google Scholar] [CrossRef]
- Lyte, M. Probiotics function mechanistically as delivery vehicles for neuroactive compounds: Microbial endocrinology in the design and use of probiotics. BioEssays 2011, 33, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.S.M.; Stylianakis, A.A.; Richardson, R. Early-life stress, microbiota, and brain development: Probiotics reverse the effects of maternal separation on neural circuits underpinning fear expression and extinction in infant rats. Dev. Cogn. Neurosci. 2019, 37, 100627. [Google Scholar] [CrossRef]
- Karen, C.; Shyu, D.J.H.; Rajan, K.E. Lactobacillus paracasei supplementation prevents early life stress-induced anxiety and depressive-like behavior in maternal separation model-possible involvement of microbiota-gut-brain axis in differential regulation of microRNA124a/132 and glutamate receptors. Front. Neurosci. 2021, 15, 719933. [Google Scholar] [CrossRef]
- De Santa, F.; Strimpakos, G.; Marchetti, N.; Gargari, G.; Torcinaro, A.; Arioli, S.; Mora, D.; Petrella, C.; Farioli-Vecchioli, S. Effect of a multi-strain probiotic mixture consumption on anxiety and depression symptoms induced in adult mice by postnatal maternal separation. Microbiome 2024, 12, 29. [Google Scholar] [CrossRef]
- Huo, R.; Zeng, B.; Zeng, L.; Cheng, K.; Li, B.; Luo, Y.; Wang, H.; Zhou, C.; Fang, L.; Li, W.; et al. Microbiota modulate anxiety-like behavior and endocrine abnormalities in hypothalamic-pituitary-adrenal axis. Front. Cell. Infect. Microbiol. 2017, 7, 489. [Google Scholar] [CrossRef]
- Agusti, A.; Lamers, F.; Tamayo, M.; Benito-Amat, C.; Molina-Mendoza, G.V.; Penninx, B.W.J.H.; Sanz, Y. The gut microbiome in early life stress: A systematic review. Nutrients 2023, 15, 2566. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Vasupanrajit, A.; Jirakran, K.; Klomkliew, P.; Chanchaem, P.; Tunvirachaisakul, C.; Plaimas, K.; Suratanee, A.; Payungporn, S. Adverse childhood experiences and reoccurrence of illness impact the gut microbiome, which affects suicidal behaviours and the phenome of major depression: Towards enterotypic phenotypes. Acta Neuropsychiatr. 2023, 35, 328–345. [Google Scholar] [CrossRef]
- Borrego-Ruiz, A.; Fernández, S. Humiliation and its relationship with bullying victimization: A narrative review. Psychol. Soc. Educ. 2024, 16, 42–51. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Study | Design | Characteristics | Outcomes |
|---|---|---|---|
| PRENATAL | |||
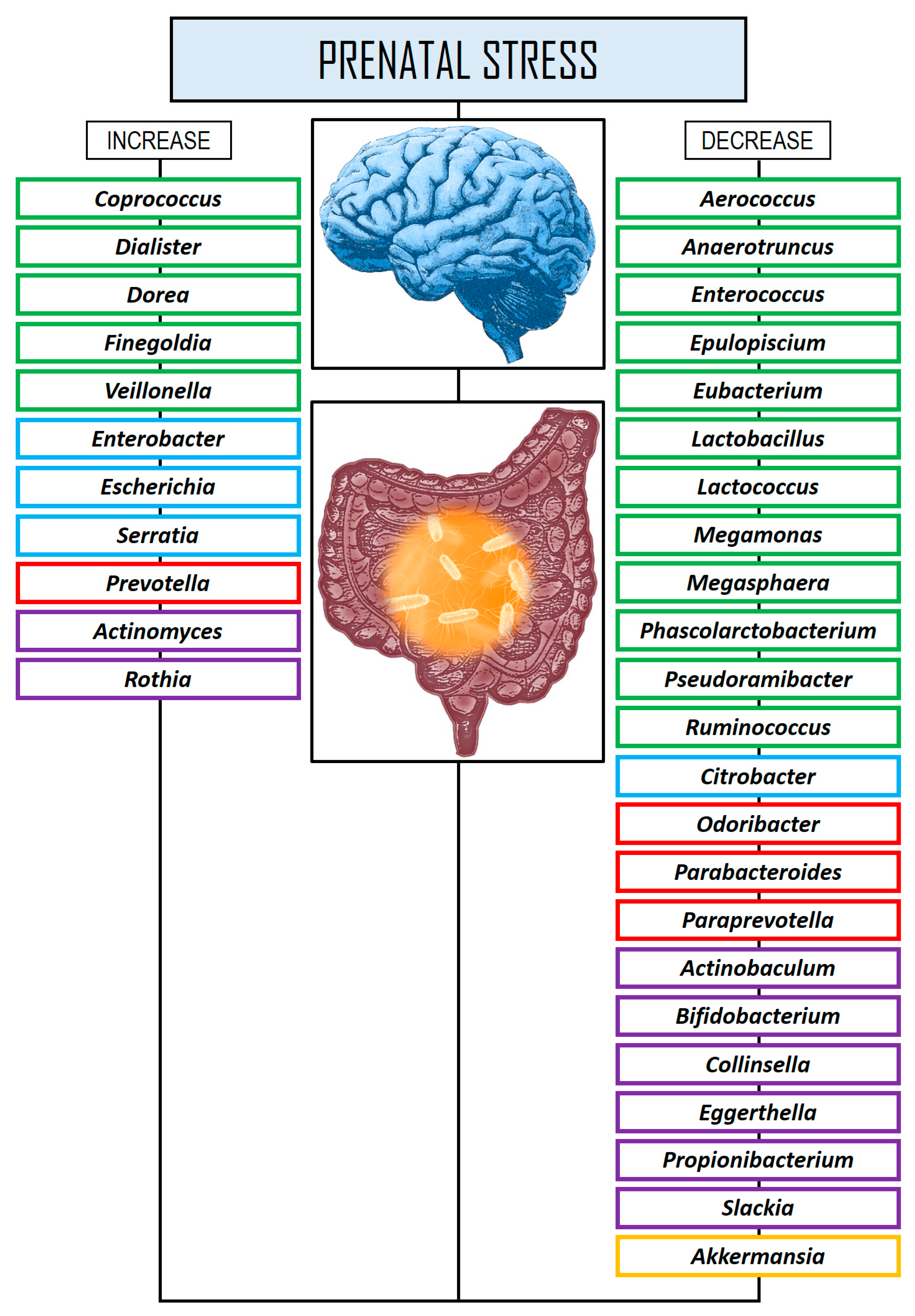
| Aatsinki et al. [150] | Longitudinal | N = 446 mother–infant (2.5 months of age) pairs. Determination of prenatal psychological distress (PPD) (N = 399) and hair cortisol concentrations (HCC) (N = 115). 16S rRNA amplicon sequencing. | Both maternal PPD and HCC were linked to alterations in the GM of infants. Chronic PPD was negatively associated with the abundance of Actinobaculum, Anaerotruncus, Akkermansia, Epulopiscium, Eubacterium, Megamonas, Megasphera, Odoribacter, Parabacteroides, Paraprevotella, Phascolarctobacterium, Propionibacterium, Pseudoramibacter, and Slackia, and positively with Actinomyces, Coprococcus, Dialister, Dorea, Finegoldia, Rothia, and Veillonella abundance. Maternal depression indicators were positively correlated with the abundance of Butyricimonas and Prevotella. Maternal HCC were associated with changes in the infant’s GM, showing negative correlations with Actinobaculum, Anaerotruncus, Butyricimonas, Citrobacter, Enterococcus, Lactobacillus, Paraprevotella, Phascolarctobacter, Ruminococcus, and Slackia. |
| Jahnke et al. [152] | Longitudinal | N = 25 mother–infant dyads. Maternal precariousness and HPA axis dysregulation were assessed both during and after pregnancy using validated scales. 16S rRNA amplicon sequencing. | Both maternal precarious exposure and HPA dysregulation were consistently linked to alterations in the infants’ GM. Maternal precarity was associated with lower diversity and higher relative abundance of Enterobacteriaceae and Streptococcaceae and a lower relative abundance of Bifidobacterium and Lachnospiraceae. Maternal HPA axis dysregulation during pregnancy was also associated with a greater relative abundance of Veillonella. |
| POSTNATAL | |||
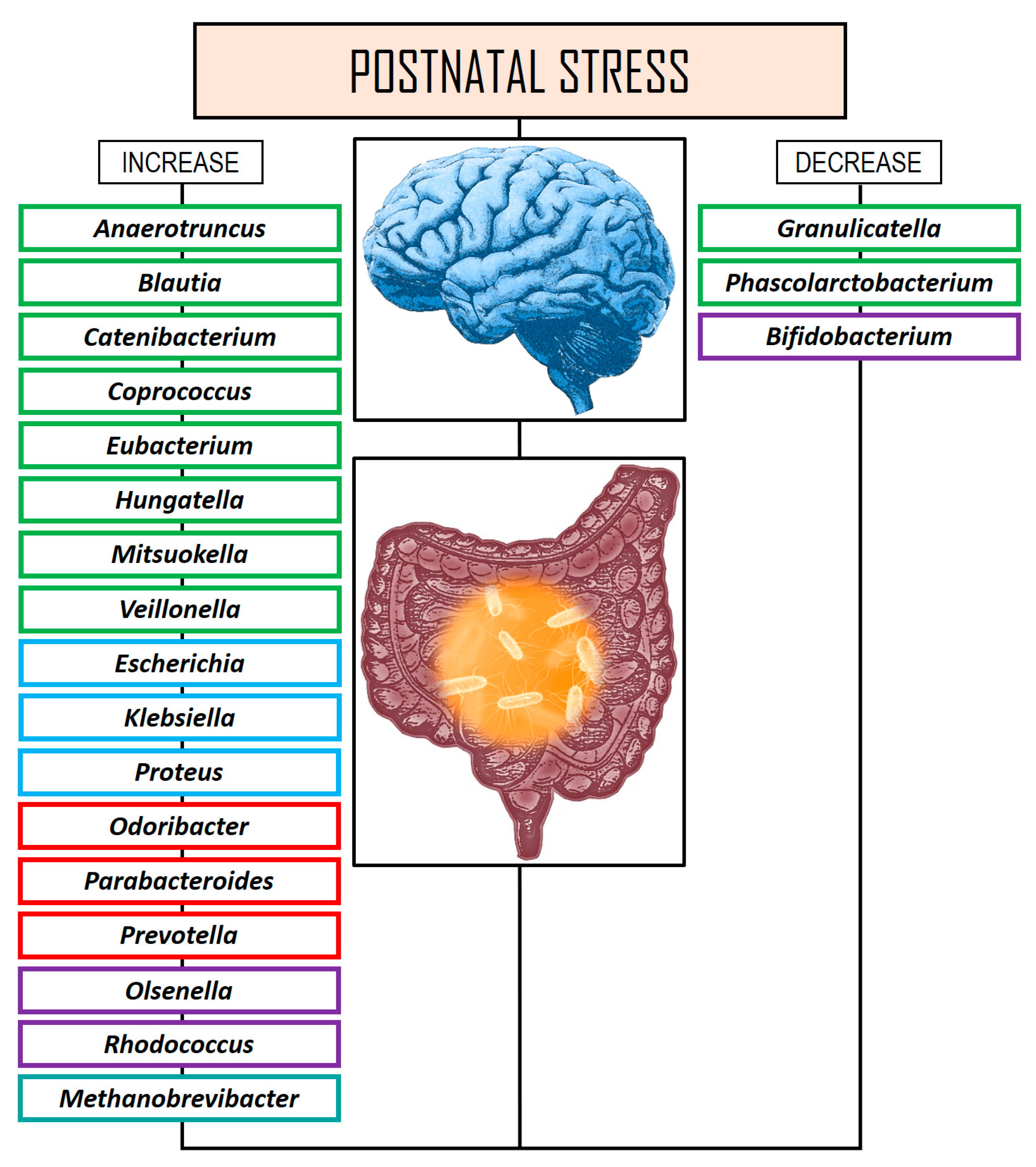
| D’Agata et al. [154] | Longitudinal | N = 82 infants stay in a neonatal intensive care unit, where stay can be stressful due to the separation of parents and infants as well as medical procedures. 16S rRNA amplicon sequencing. | Higher stress scores on the Neonatal Infant Stressor Scale were associated with higher relative abundances of Proteus and Veillonella. |
| Reid et al. [156] | Case-control | N = 17 adolescents (aged 13–21) who had been internationally adopted from orphanages, comparing them with non-institutionalized adolescents (N = 18). 16S rRNA amplicon sequencing. | The stressed group had significantly higher abundances of several bacterial taxa, including the genera Bacteroides, Coprococcus, Escherichia, Prevotella, and Streptococcus, compared to the non-institutionalized group. |
| Malan-Muller et al. [158] | Case-control | N = 79 participants with PTSD (>18 years old). 16S rRNA amplicon sequencing. | Random forest models identified a consortium of the following four genera Mitsuokella, Odoribacter, Catenibacterium, and Olsenella could distinguish PTSD status with 66.4% accuracy. The relative abundance of this consortium was higher in the PTSD group and correlated positively with CAPS-5 and CTQ scores. MDD diagnosis was also associated with increased relative abundance of the Bacteroidota phylum. |
| Hantsoo et al. [160] | Longitudinal | N = 48 of healthy women between 20 and 26 weeks of gestation compared a low ACEs control group with a high ACEs group. 16S rRNA amplicon sequencing. | Women in the high ACEs group exhibited a higher differential abundance of Prevotella and a lower abundance of Erysipelotrichaceae and Phascolarctobacterium compared to those in the low ACEs group |
| Michels et al. [161] | Cross-sectional | N = 93 children-adolescents (8–16 years old) with psychosocial stress. 16S rRNA amplicon sequencing. | High stress was associated with lower Bacillota at the phylum level and higher Bacteroides, Parabacteroides, Rhodococcus, Methanobrevibacter, and Roseburia but lower Phascolarctobacterium at genus level. |
| Flannery et al. [162] | Cross-sectional | N = 40 children (mean age 6.12 years). Shotgun metagenomics analyses using Illumina GAIIx platform, taxonomically profiled using MetaPhlAn, | Both socioeconomic risk exposure and child behaviors associate with the relative abundances of specific taxa, such as Bacteroides spp. and Bifidobacterium spp. as well as functional modules encoded in their genomes (e.g., monoamine metabolism) that have been linked to cognition and health. B. fragilis was associated with reduction in aggressively, emotional reactivity, sadness, and impulsivity. Coprococcus comes and Eubacterium rectale were associated with increase in anxious depression and less inhibitory control. Roseburia inulinivorans was associated with reduced depressive problems. |
| Keskitalo et al. [163] | Case-control | N = 193 infants (2.5-month-old) with both fecal sample and salivary stress response measurement. 16S rRNA amplicon sequencing. | The findings indicated that a reduced cortisol response to the stressor was weakly linked to GM diversity, while stronger associations were observed between cortisol levels and the specific taxonomic composition of the children’s fecal microbiome. |
| Laue et al. [164] | Longitudinal | N = 260 children. 16S rRNA amplicon sequencing. | The findings indicated that while most outcomes were not linked to changes in beta-diversity, a higher diversity in the microbiome at 6 weeks was associated with lower depression levels in the entire sample, as well as reduced anxiety and improved internalizing behaviors, particularly in boys. In boys, better adaptive functioning scores correlated positively with Bifidobacterium spp., Bacteroides vulgatus, and Streptococcus spp. abundance, but negatively with the abundance of Klebsiella oxytoca, Clostridium spp., and Haemophilus spp. Better depression score was associated with the abundance of Tyzzerella nexelis, and negative relationship with Eggerthella lenta abundance. In girls, Granulicatella was associated with higher anxiety scores at 6 weeks, while Streptococcus peroris was linked to better internalizing behavior at 1 year. Furthermore, certain Blautia spp. were related to higher hyperactivity scores, with a stronger association observed in girls. |
| Kraaij et al. [165] | Cross-sectional | N = 1784 children (10-year-old) with mental problems. 16S rRNA amplicon sequencing. | Lower GM diversity and richness was related to internalizing problems and anxious/depressed conduct problems. Although a definitive evidence linking GM diversity, taxonomic traits, or functions, and mental health problems in the pediatric population was not established, a reduction in the genera that have previously been related to psychiatric disorders, including Anaerotruncus, Hungatella, and Oscillospiraceae was found. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borrego-Ruiz, A.; Borrego, J.J. Early Life Stress and Gut Microbiome Dysbiosis: A Narrative Review. Stresses 2025, 5, 38. https://doi.org/10.3390/stresses5020038
Borrego-Ruiz A, Borrego JJ. Early Life Stress and Gut Microbiome Dysbiosis: A Narrative Review. Stresses. 2025; 5(2):38. https://doi.org/10.3390/stresses5020038
Chicago/Turabian StyleBorrego-Ruiz, Alejandro, and Juan J. Borrego. 2025. "Early Life Stress and Gut Microbiome Dysbiosis: A Narrative Review" Stresses 5, no. 2: 38. https://doi.org/10.3390/stresses5020038
APA StyleBorrego-Ruiz, A., & Borrego, J. J. (2025). Early Life Stress and Gut Microbiome Dysbiosis: A Narrative Review. Stresses, 5(2), 38. https://doi.org/10.3390/stresses5020038