Abstract
Aflatoxin B1 (AFB1), a potent mycotoxin, poses a significant threat to animal health through contaminated feed. Our study aims to investigate the neurotoxic effects of AFB1in chickens, with a special emphasis on the brain. Seven-day-old chickens were fed AFB1-contaminated feed (5 mg of AFB1/kg of feed) for two weeks, after which neurobehavioral assessments and biochemical analyses were conducted and compared to control chickens. In the open-field test, chickens exposed to AFB1 exhibited a reduction in locomotor activity and exploratory behavior. Additionally, AFB1 exposure increased the tonic immobility response. Biochemical analyses revealed that AFB1-contaminated feed reduced whole-brain acetylcholinesterase activity, suggesting impaired cholinergic neurotransmission. Indicators of oxidative stress in the brain revealed a reduction in glutathione levels, superoxide dismutase levels, and total antioxidant capacity, alongside an increase in malondialdehyde levels, indicating heightened oxidative stress in the brain. The neurotoxic effects of AFB1 were further supported by the upregulation of pro-inflammatory cytokine genes, including interleukin-1 beta, interleukin-6, interleukin-17, and inducible nitric oxide synthase, as determined by real-time quantitative polymerase chain reaction. Our study demonstrates that AFB1-contaminated feed influences chicken neurobehavioral outcomes and brain biochemistry and represents the inaugural evidence that AFB1 exposure markedly reduces AChE activity in the whole brain of chickens.
1. Introduction
Aflatoxins are potent and prevalent mycotoxins produced by several species of Aspergillus, mainly Aspergillus flavus and Aspergillus parasiticus. They present a considerable threat to global public health, particularly through contaminated feed such as peanuts, corn, and grains [1]. Aflatoxins that contaminate chicken feed can subsequently be transmitted to eggs, meat, and other consumable components, posing a health hazard to consumers [2]. Aflatoxin B1 (AFB1) is the most common and potent aflatoxin, accounting for 75% of all aflatoxins generated in food products contaminated by fungi. A significant amount of evidence from animal and human studies indicates that AFB1 is one of the naturally occurring carcinogens, and it has been classified as a group 1 carcinogen by the International Agency for Research on Cancer [3]. In addition, AFB1 is recognized as a teratogen and a mutagen [4]. Moreover, AFB1 adversely affects the immune system in chickens by inhibiting antibody production, limiting lymphocyte proliferation, and diminishing the functionality of immune organs, thereby increasing the birds’ vulnerability to infections [5]. Furthermore, it adversely impacts growth performance by diminishing feed intake and reducing nutrient digestibility, which results in poor weight gain [6]. Consumption of AFB1-contaminated feed produces a clear toxicity to internal organs, particularly hepatotoxicity, cardiotoxicity, and nephrotoxicity [7,8].
AFB1 demonstrates neurotoxic properties, which can lead to cognitive and behavioral impairments [9]. The ability to cross the blood–brain barrier compromises brain integrity, leading to the accumulation of AFB1 in brain tissue, which promotes neuronal damage in the central nervous system and contributes to cognitive decline [10]. It is important to note that the neurotoxicity of AFB1 does not solely arise from the parent compound. Several biotransformation products, such as aflatoxin B1-8,9-epoxide, are known for their high reactivity with cellular macromolecules and induction of cellular toxicity [11]. These metabolites may vary in toxicity, and their formation heavily depends on species-specific metabolic pathways. Notably, the efficiency of AFB1 biotransformation differs among species, influencing their susceptibility to AFB1-induced toxicity [12]. In poultry, data on the neurotoxic potential of AFB1 metabolites are limited, indicating a need for further mechanistic investigation. A review of prior studies in both in vitro and animal models indicates that AFB1 neurotoxicity is linked to neurotransmitter suppression and physiological disruptions, leading to neurobehavioral impairments [10,13]. At the cellular level, AFB1 directly affects neuronal cells by inducing the production of reactive oxygen species (ROS), inflammatory responses, DNA damage, S-shape cell cycle arrest, and apoptosis [14]. Recent insights indicate that AFB1, along with T-2 toxin, can activate glial cells, disrupt synaptic transmission, and promote neuroinflammation through oxidative stress and mitochondrial dysfunction, providing a more comprehensive understanding of the mechanisms underlying AFB1-induced neurotoxicity [15].
Acetylcholinesterase (AChE), a key enzyme in cholinergic neurotransmission, has been identified as a target of various neurotoxicants [16]. However, its role in AFB1-induced neurobehavioral impairments in poultry represents a substantial gap. Given the critical role of the cholinergic system in cognitive and motor functions, investigating AChE activity in the brains of AFB1-exposed chickens may provide valuable mechanistic insights about AFB1 neurotoxicity. In addition, the possible oxidative–inflammatory changes that may be involved and exacerbate AFB1 toxicity, particularly in the brains of chickens, remain poorly understood and represent a promising area for research. Given the frequent exposure of chickens to AFB1 through contaminated feed, investigating its effects on the avian brain is necessary for expanding our understanding of AFB1 neurotoxicity, protecting poultry health, and promoting the sustainability of the poultry industry. This study aims to address this knowledge gap by determining the effects of AFB1 on neurobehavioral performance, AChE activity, oxidative stress status, and neuroinflammation in the brains of chickens. To our knowledge, this is the first study to demonstrate that AFB1 significantly reduces whole-brain AChE activity in chickens, suggesting a novel mechanism of cholinergic disruption in chickens. By integrating behavioral assessments with biochemical analyses, this research might provide critical insights into the neurotoxic mechanisms of AFB1 in the chicken brain. These findings may have important applications in the development of early biomarkers of mycotoxin-induced neurotoxicity in poultry, the improvement of feed quality control protocols, and the protection of animal welfare and productivity in commercial farming systems.
2. Results
2.1. Clinical Observation of Intoxicated Chickens
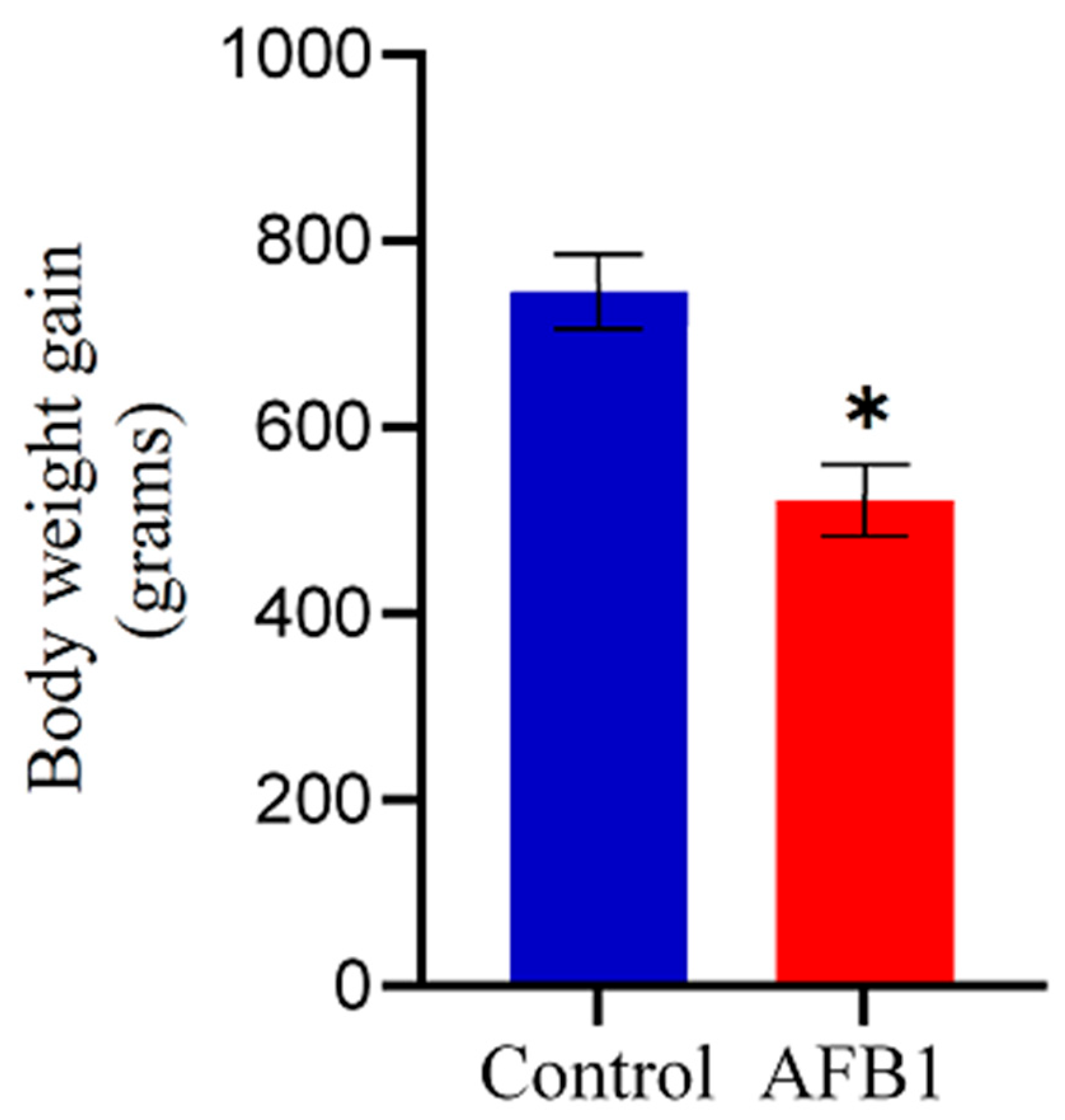
In this experiment, no broilers died; nonetheless, the chickens in the AFB1 group exhibited smaller size, diminished appetite, and slower growth compared to the control group, evidenced by their significantly lower weight gain (p < 0.05) relative to the control group (Figure 1). Moreover, the birds showed signs of depression, including dull eyes and loose and lackluster feathers, compared to the control group, which maintained smooth and healthy feathers.
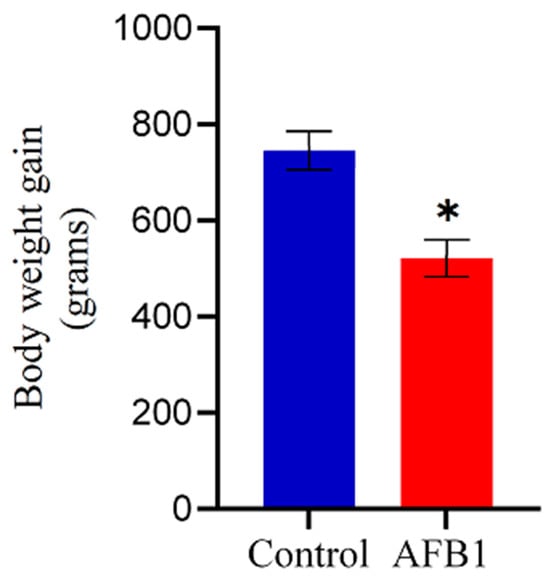
Figure 1.
Effect of AFB1 on chicken growth. Chickens treated with AFB1 exhibited significantly reduced weight gain compared to the control group. Data are shown as mean ± SD (grams) from 20 biological replicates. The asterisk (*) denotes a statistically significant difference compared to the control group with a p < 0.05.
2.2. Neurobehavioral Effects of AFB1 Based on Open-Field Activity and Tonic Immobility
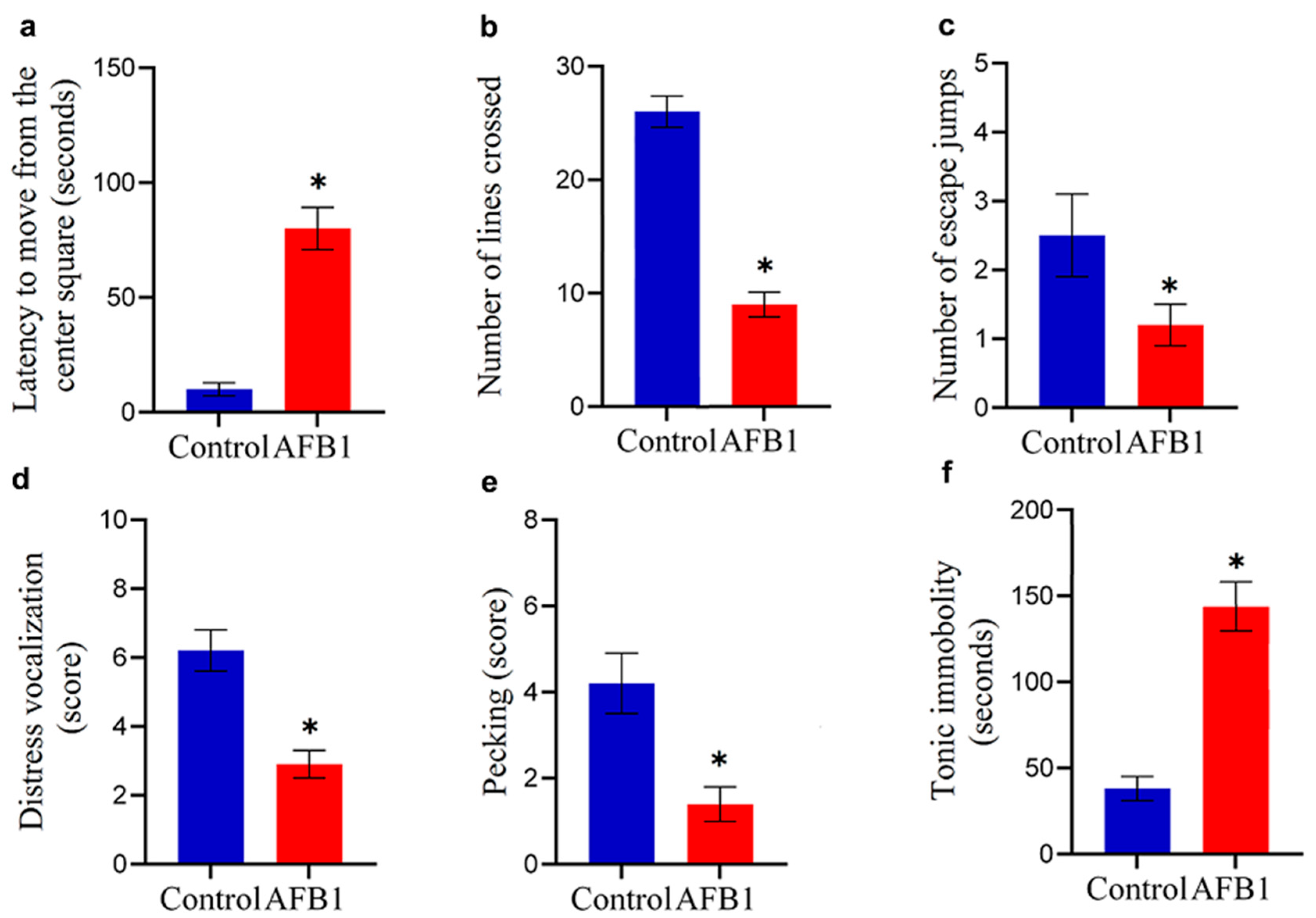
The five-minute open-field activity patterns and tonic immobility performance of chickens that were given AFB1-contaminated feed with a concentration of 5 mg of AFB1/kg of feed over two weeks are shown in Figure 2. AFB1 consumption significantly impaired locomotor activity in chickens. This was evidenced by a prolonged latency to exit the central square of the open-field arena (p < 0.05) and a marked reduction in the number of lines crossed compared to the control group (p < 0.05; Figure 2a,b). Additionally, AFB1 intoxication resulted in a significant decrease in both the number of escape jumps and the vocalization score relative to control values (p < 0.05; Figure 2c,d). The exploration behavior of chickens that ingested feed contaminated with AFB1 was notably impacted, as evidenced by a significant decrease (p < 0.05) in the score for pecking at the ground compared to the control group (Figure 2e). AFB1 significantly increased (p < 0.05) the duration of the chicks’ tonic immobility response compared to the control group (Figure 2f), indicating that it induces fear and hypoactivity in the chickens.
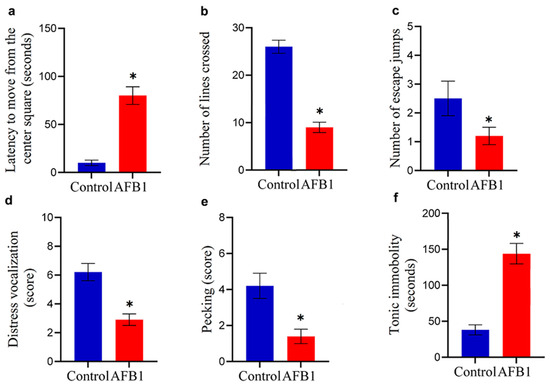
Figure 2.
Neurobehavioral effects of AFB1 in chickens. Chickens fed AFB1-contaminated feed (5 mg AFB1/kg) for two weeks exhibited reduced locomotor activity, shown by increased latency to move from the central square and fewer crossed lines in the open-field test (* p < 0.05 (a,b)). AFB1 exposure also decreased escape jumps and vocalization scores (* p < 0.05 (c,d)). Exploratory behavior was impaired, as indicated by reduced pecking activity (* p < 0.05 (e)). Additionally, AFB1 significantly prolonged tonic immobility duration, reflecting hypoactivity (* p < 0.05 (f)). Data are expressed as mean ± SD (n = 20). (*) denotes a statistically significant difference compared to the control.
2.3. Effect of AFB1 on Brain AChE Activity
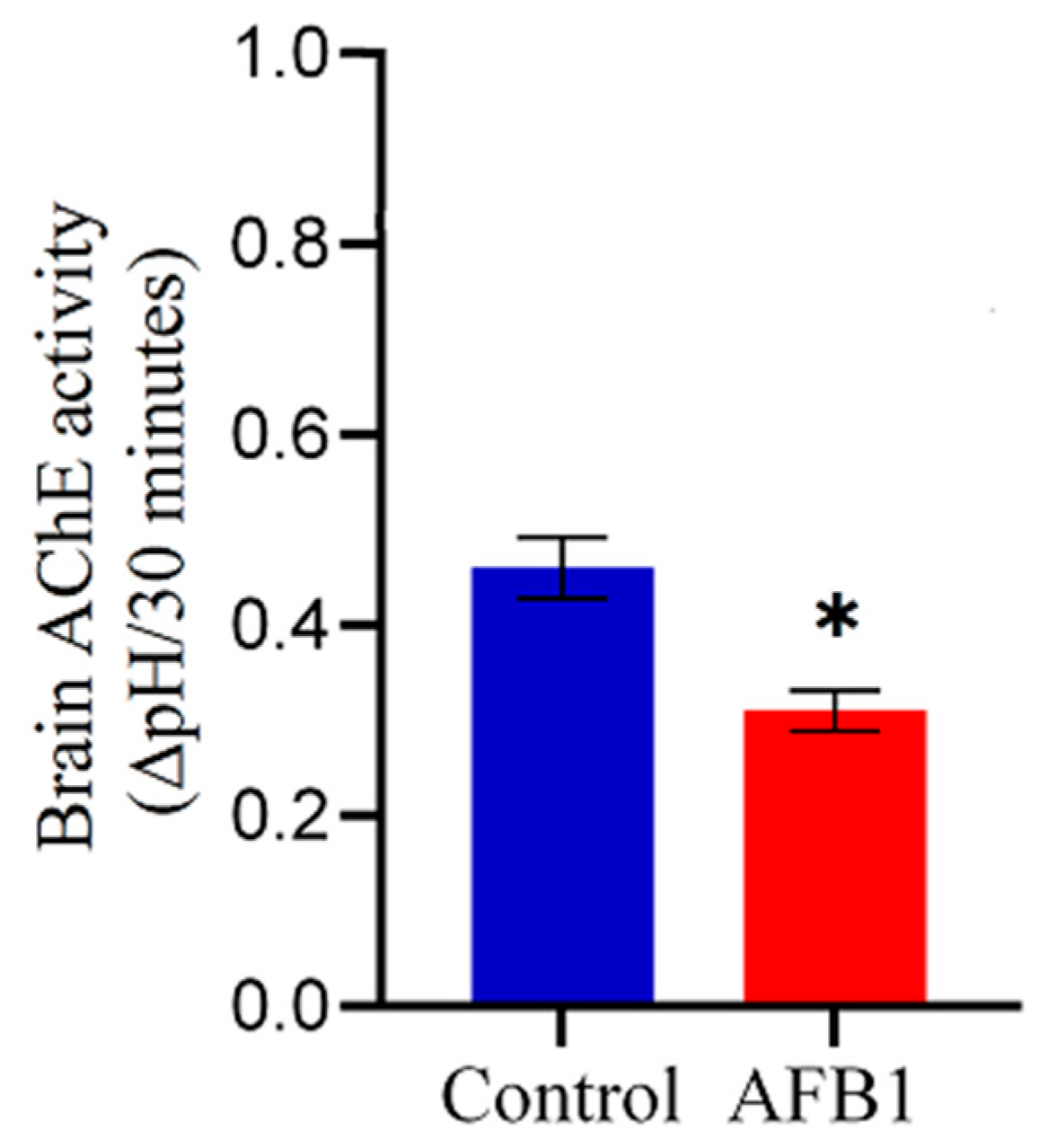
Chickens exposed to AFB1-contaminated feed (5 mg AFB1/kg) for two weeks exhibited a significant (p < 0.05) reduction in whole-brain AChE activity compared to the control group (Figure 3). Specifically, AFB1 exposure led to a 32.6% decrease in whole-brain AChE activity relative to control chickens, indicating potential neurotoxic effects of AFB1 on cholinergic function.
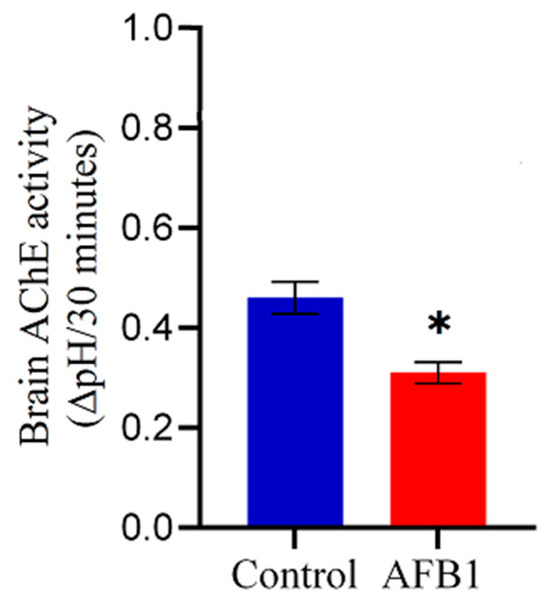
Figure 3.
Effect of AFB1 on whole-brain AChE activity in chickens. Chickens were fed AFB1-contaminated feed (5 mg AFB1/kg) for two weeks. (*) indicates a significant reduction in whole-brain AChE activity compared to the control group (p < 0.05). Data are presented as mean ± SD (n = 20).
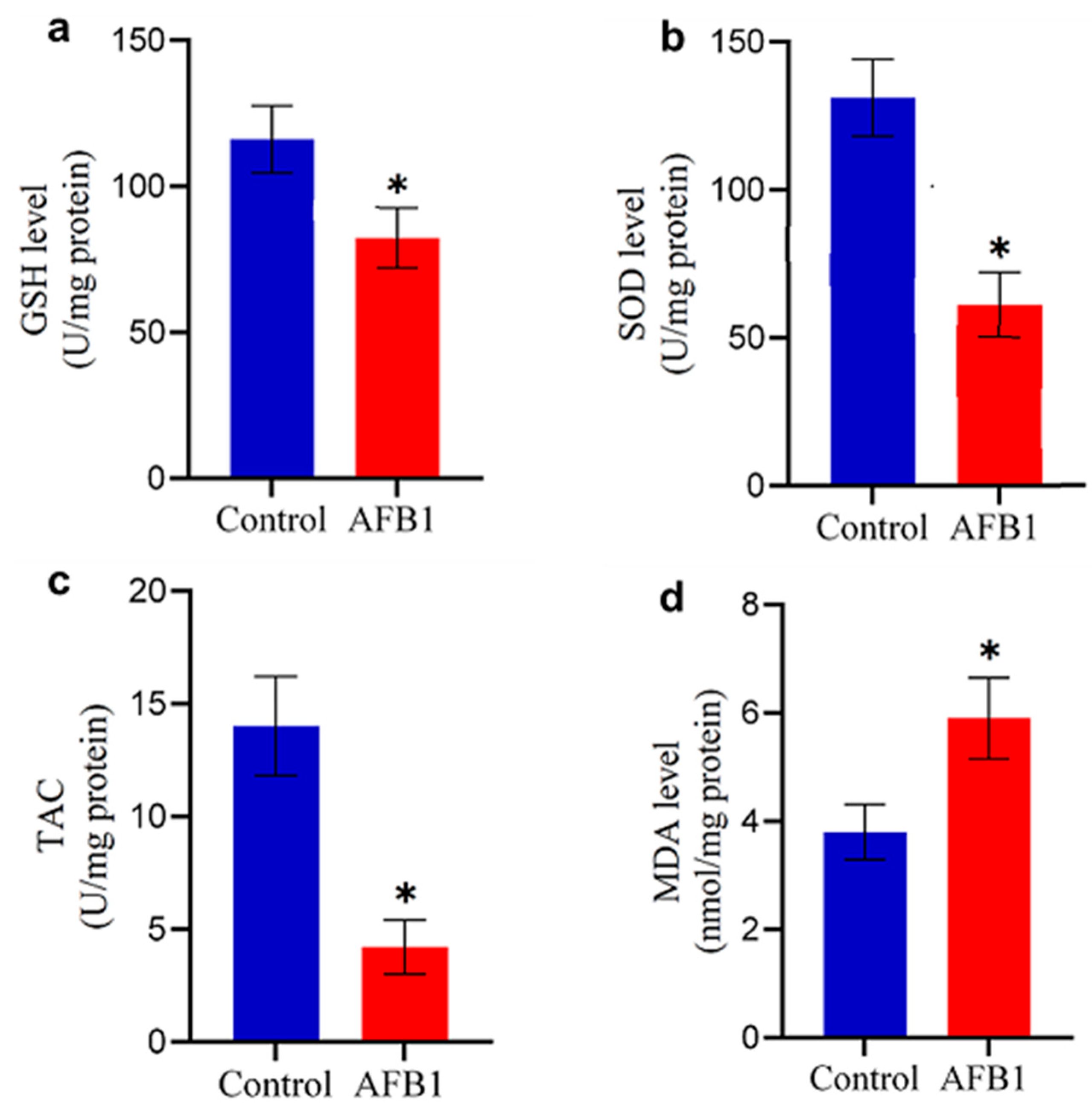
2.4. Antioxidant Imbalance in the Brain of AFB1-Treated Chickens
Exposure to AFB1-contaminated feed significantly altered oxidative stress markers and antioxidant enzyme levels in the brains of treated chickens. Compared to the control group, AFB1-exposed chickens exhibited a marked reduction in GSH content and SOD levels (p < 0.05; Figure 2b and Figure 4a), indicating compromised antioxidant defenses. Similarly, TAC was significantly diminished (p < 0.05; Figure 4c). In contrast, the MDA level was significantly elevated (p < 0.05; Figure 4d), suggesting increased oxidative damage. These results highlight the role of AFB1-induced oxidative stress in disrupting brain homeostasis, potentially contributing to neurotoxic effects.
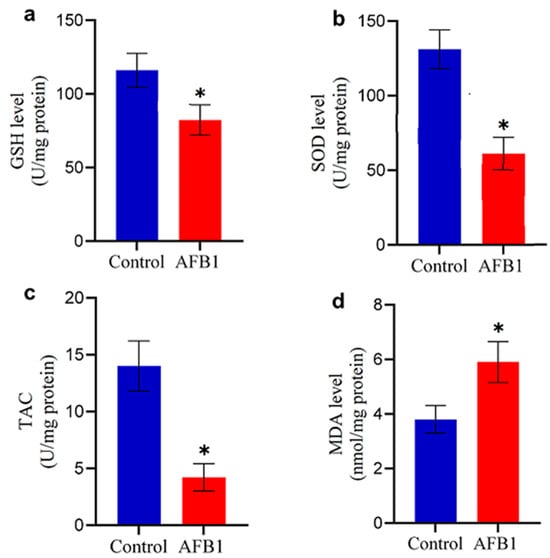
Figure 4.
Changes in oxidative stress markers in the brain of control and AFB1-treated chickens: (a) GSH level; (b) SOD level; (c) TAC; and (d) MDA content. Data are presented as mean ± S.D. (n = 20). (*) indicates a significant difference (p < 0.05) compared to the control group.
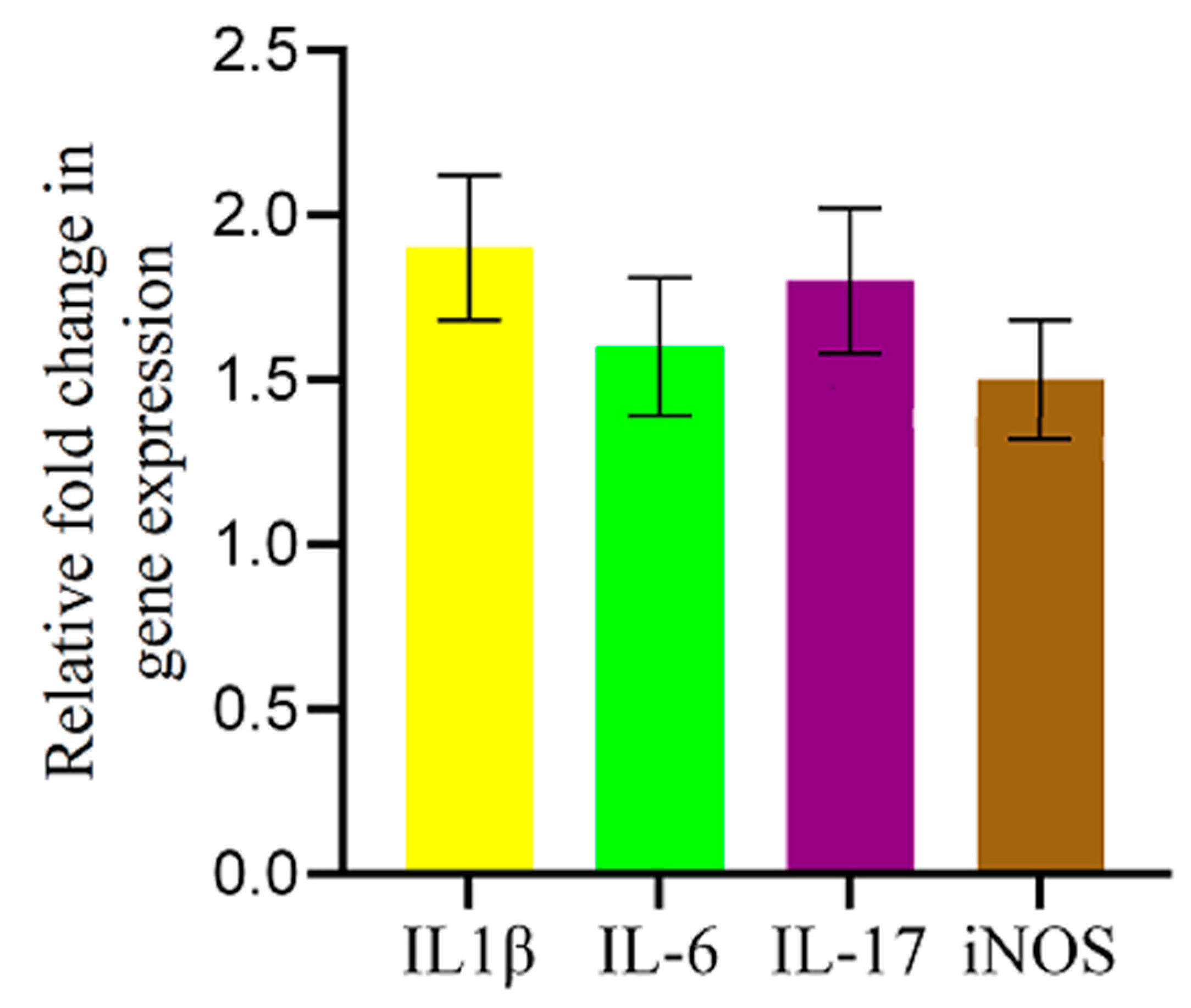
2.5. AFB1-Induced Alterations in Inflammation-Related Gene Expression in the Chicken Brain
To investigate the inflammatory response triggered by AFB1 in the chicken brain, we quantified the mRNA expression levels of key inflammatory mediators—IL-1β, IL-6, IL-17, and iNOS—using qRT-PCR. As shown in Figure 5, AFB1 exposure resulted in a significant upregulation of all four genes compared to the control group. IL-1β exhibited the highest increase (1.9-fold), followed by IL-17 (1.8-fold) and IL-6 (1.6-fold), indicating a clear pro-inflammatory response. Additionally, iNOS expression was elevated by 1.5-fold, suggesting increased nitric oxide production and associated inflammation-driven oxidative stress. These findings provide strong evidence that AFB1 induces neuroinflammation, potentially contributing to its neurotoxic effects in chickens.
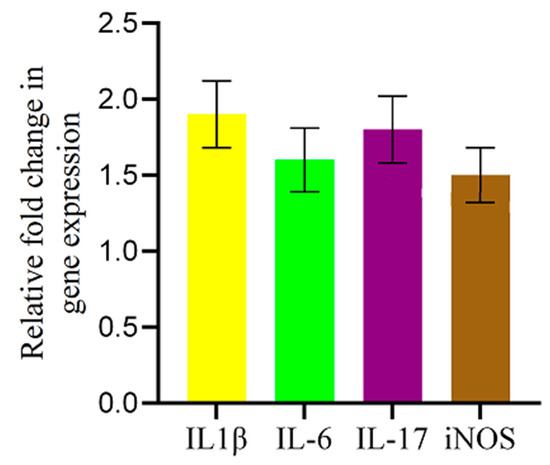
Figure 5.
AFB1-induced upregulation of inflammatory-related genes in the chicken brain. qRT-PCR analysis of IL-1β, IL-6, IL-17, and iNOS mRNA expression levels in the brains of AFB1-treated chickens. Gene expression was normalized to the 28S gene and is shown as fold change relative to controls. AFB1 significantly increased IL-1β, IL-6, IL-17, and iNOS expression. Data are presented as mean ± SD (n = 20).
3. Discussion
The contamination of poultry feed with AFB1 is considered a critical global challenge; however, its neurotoxic effects, particularly in chickens, remain poorly comprehended. We chose chickens as the model organism due to their economic importance in the poultry industry and heightened exposure and sensitivity to environmental contaminants. Given the brain’s vulnerability to toxic compounds [14], our study focused on AFB1-induced neurotoxicity, with special emphasis on the brain. We demonstrate that AFB1-contaminated feed impairs neurobehavioral function, decreases AChE activity, increases oxidative stress, and promotes neuroinflammation in the brains of chickens.
Our study demonstrates the negative effects of AFB1 on growth performance and health in poultry. The chickens exposed to AFB1-contaminated feed suffered from smaller body size, decreased appetite, and significantly lower weight gain relative to the control group. Furthermore, they displayed symptoms of dull eyes and a poor feather condition, suggesting a decline in overall health. Our findings align with previous research demonstrating that AFB1 exposure significantly reduces body weight and growth rates in broilers, which can be primarily linked to adverse effects on protein synthesis and liver function, as well as the suppression of the feeding center, which ultimately results in diminished feed intake [17].
Chickens intoxicated with AFB1 showed a marked reduction in both locomotor and exploratory activities based on the behavioral assessment using an open field test. This finding is consistent with a study demonstrating that AFB1 exposure impairs locomotor function and induces anxiety- and depression-like behaviors, as observed in rats administered 25 μg/kg/week of AFB1 for eight weeks [18]. In contrast to the continuous exposure used in our study, a study in rats reported no significant neurobehavioral impairments following intermittent AFB1 exposure [19]. This discrepancy suggests that factors such as dosage, exposure duration, and species-specific differences may play a crucial role in determining the neurotoxic effects of AFB1.
To the best of our knowledge, this study is the first to demonstrate a significant decline in whole-brain AChE activity in chickens exposed to AFB1, underscoring impaired cholinergic neurotransmission as a potential mechanism of neurotoxicity. Our novel finding aligns with previous studies highlighting the neurotoxic impacts of AFB1 on cholinergic activity. For instance, a study has shown a significant reduction in AChE activity in the brains of silver catfish following dietary AFB1 exposure [20]. Similarly, intraperitoneal administration of AFB1 at a dose of 0.6 mg/kg/day for four days resulted in a marked decrease in AChE activity in mouse brain tissue, accompanied by an increase in monoamine oxidase activity, which were linked to depression-like behaviors, memory impairments, and learning deficits, further reinforcing the role of AFB1 in impairing neurotransmission [21]. These results are further supported by in vitro studies that demonstrated that AFB1 non-competitively inhibits mouse brain AChE activity, suggesting a direct biochemical mechanism by which AFB1 may alter cholinergic signaling [22]. Although a definitive causal relationship remains unclear, prior studies have shown that reduced brain AChE activity is strongly correlated with neurobehavioral deficits, including decreased exploratory behavior and impaired learning and memory in rodents [23]. Our study suggests cholinergic dysfunction as a potential contributing factor in the behavioral disturbances observed in chickens after oral exposure to AFB1.
Chickens fed with AFB1-contaminated feed led to a reduction in the levels of essential antioxidant molecules in the brain, including GSH and SOD, as well as a reduction in TAC, coinciding with a marked increase in MDA, a marker of lipid peroxidation in tissues, indicating elevated oxidative stress. The imbalance between oxidative stress generation and antioxidant defenses highlights the increased susceptibility of the chicken brain to AFB1-induced neurotoxicity. This result suggests that oxidative stress is one mechanism responsible for brain dysfunction in chickens following AFB1 intoxication. This finding is consistent with a previous study that showed aflatoxins can cause oxidative stress in broiler chickens, leading to significant cellular damage [24]. Similarly, a study in mice reported that AFB1 exposure significantly increased MDA levels in the brain while reducing GSH, catalase, and SOD activity [25]. These collective findings highlight the conserved nature of AFB1-induced oxidative stress across species and emphasize the critical role of oxidative imbalance in its neurotoxic effects.
In our study, AFB1 exposure led to a significant upregulation of pro-inflammatory cytokines IL-1β, IL-6, and IL-17, along with increased iNOS expression in the chicken brain, indicating a robust neuroinflammatory response. These findings align with previous research in rodents, where even low-dose AFB1 exposure (20 ng/mL) has been shown to elevate the mRNA expression of key inflammatory mediators, including NF-κB, IL-1β, and TNF-α, while also promoting the secretion of IL-1β, IL-6, TNF-α, and IL-10 in microglial and astrocyte cultures [26]. Although AFB1-induced neuroinflammation is well-documented in mammalian models [10], studies on its impact in avian species remain limited. Our findings provide novel evidence that AFB1 elicits an inflammatory response in the chicken brain.
This study provides evidence that AFB1 acts as a neurotoxicant by impairing cholinergic signaling, inducing oxidative stress, and triggering inflammation—mechanisms linked to neurobehavioral disorders. These findings align with toxicological theories identifying oxidative stress and inflammation as key drivers of toxin-induced neural damage. The neurotoxic impact of AFB1 on the brain in chickens remains relatively underexplored. Our study fills this gap by proving that AFB1-induced brain toxicity not only affects mammals but also poultry, thereby posing significant risks to avian health and productivity. Our findings highlight the need for stricter regulation of AFB1 contamination in poultry feed to protect poultry health. Given AFB1’s neurotoxic effects, interventions such as antioxidant supplementation and improved feed storage practices may help mitigate oxidative stress and inflammation.
While our findings provide valuable insights into the neurotoxic effects of AFB1 in chickens, several limitations should be acknowledged. First, our biochemical analyses were conducted on whole-brain homogenates, which may have masked region-specific variations in AChE activity, oxidative stress markers, and inflammatory responses. Different brain regions may exhibit varying susceptibilities to AFB1, and future studies should investigate these localized effects. Subsequent studies employing region-specific assessments could provide a more precise understanding of AFB1-induced neurotoxicity. Second, although our study focused on the neurotoxic effects of AFB1, we did not conduct histopathological examinations of peripheral organs such as the liver and kidneys. Including such assessments in future studies could provide a more comprehensive understanding of the systemic toxicity induced by AFB1 and its correlation with neurobiological alterations. Third, this study employed a 12:12 light–dark cycle, which, while commonly used in laboratory settings, could influence physiological rhythms, behavior, and potentially the neurotoxic effects of AFB1. Although, this lighting schedule was selected for consistency with previous studies on AFB1 toxicity in chickens [27,28]. We acknowledge that different light schedules may affect the manifestation of neurotoxic effects. Future studies should consider how variations in circadian rhythms might influence the manifestation of neurotoxic effects of AFB1. Lastly, our study focused on a relatively short exposure period, whereas prolonged exposure may yield additional insights into the progressive nature of AFB1-induced neurotoxicity. Addressing these limitations in future research will further clarify the mechanisms underlying AFB1’s impact on the avian nervous system.
4. Materials and Methods
4.1. Chemicals
AFB1 from Aspergillus flavus (CAS No. 1162-65-8), chloroform (CAS No. 67-66-3), sodium barbital (CAS No. 144-02-5), sodium chloride (CAS No. 7647-14-5), potassium dihydrogen phosphate (CAS No. 7778-77-0), and acetylthiocholine iodide (CAS No. 1866-15-5) were purchased from Sigma-Aldrich (Saint Louis, MO, USA). Glutathione (GSH), superoxide dismutase (SOD), total antioxidant capacity (TAC), and malondialdehyde (MDA) kits were purchased from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). TRIzol™ reagent and PowerUp™ SYBR™ Green Master Mix were acquired from Thermo Fisher Scientific (Mount Prospect, IL, USA), whereas qScript cDNA SuperMix was procured from Quantabio (Beverly, MA, USA).
4.2. Chicken Husbandry and Treatment
Forty-one-day-old broiler chicks (Ross 308) were employed. The birds were obtained from a local commercial farm and used in this study. Upon arrival, the birds were housed in clean, well-ventilated pens under standard environmental conditions. The ambient temperature was maintained at 21–24 °C, with a 12:12 light–dark cycle. All birds had ad libitum access to a commercial finisher diet and clean drinking water from day 1 to day 21. The birds underwent a seven-day adaptation period before the experiment began. The birds received the necessary veterinary care, including routine health monitoring and treatment when needed. A standard vaccination program was applied in accordance with regional poultry health guidelines to protect against common infectious diseases. This study received ethical approval from the research ethics approval committee of the College of Veterinary Medicine at the University of Al-Qadisiyah, under approval number 4413, in November 2024.
4.3. Preparation of AFB1-Contaminated Diet
To prepare the feed contaminated with AFB1, pure crystalline AFB1 was dissolved in chloroform at 1 mg per 10 mL and subsequently mixed with an appropriate quantity of ground feed. Following the careful handling of the resultant contaminated premix feed within a fume hood, and while appropriately equipped with the necessary laboratory personal protective equipment, the premix was permitted to air-dry at ambient temperature overnight for the evaporation of the solvent. Subsequently, it was thoroughly mixed with the basal diet to attain the specified concentration of AFB1 (5 mg of AFB1/kg of feed). The AFB1 dose of 5 mg/kg of feed was selected based on preliminary data and previous studies demonstrating its effectiveness in inducing observable toxic systemic effects in poultry [27,28,29]. While the final concentration of AFB1 in the feed was based on precise theoretical calculations using known quantities of crystalline AFB1, direct analytical verification of AFB1 levels in the prepared feed was not performed in this study. To serve as a control diet, the basal diet was formulated by mixing the same ground diet with an equivalent quantity of chloroform, excluding AFB1. In the same manner, the mixture was permitted to dry overnight before being thoroughly mixed with the basal diet.
4.4. Experimental Design
Following the adaptation period, the chicks were randomly assigned to two groups. The AFB1-exposed group (n = 20) received an AFB1-contaminated diet for two weeks, while the control group (n = 20) was maintained on a basal diet. During the experiment period, the clinical signs of the birds were monitored. On the final day of the experiment, after the neurobehavioral assessments, chickens were euthanized via intravenous administration of sodium pentobarbital (100 mg/kg body weight). This method ensures rapid loss of consciousness with minimal stress, thereby preserving the integrity of downstream biochemical analyses [30].
4.5. Open Field Test
It serves as a commonly used technique in neurophysiological research to evaluate chickens’ locomotor activity and exploratory behavior. An open field box with the dimensions of (90 cm × 90 cm × 50 cm) was used. The arena was divided into 24 equal squares, with a smooth floor, a quiet environment, and consistent lighting to minimize external influences on behavior. Before the trials, we maintained the birds on regular feeding schedules and allowed them to adjust to the experimental environment before testing. The experiments were carried out between 9 and 12 a.m., and the birds were delicately handled to minimize stress. Fifty grams of wheat were dispersed across the surface of the arena. Each chick was individually placed at the center of the arena and observed for an average of five minutes. Locomotor activity was assessed using the following behavioral parameters: latency to exit the central square, number of line crossings, number of escape jumps, and distress vocalization, scored as 0 (no calls), 1 (1–2 calls), 2 (3–4 calls), and 3 (≥5 calls). Exploratory behavior was evaluated by recording ground-pecking frequency, scored as 0 (no pecking), 1 (1–2 pecks), 2 (3–4 pecks), and 3 (≥5 pecks) [31].
4.6. Tonic Immobility Test
This test evaluates the fearfulness and stress susceptibility of chickens. Birds exhibiting prolonged tonic immobility are deemed more fearful; those that quickly recover are regarded as less fearful or more resilient to stress. The test involves individually positioning each chick on its right side on a wooden surface. The hands are then quietly withdrawn, and the duration of immobility is measured until the chicken exhibits resistance and begins to right itself and stand [32].
4.7. Determination of Brain AChE Activity
The chickens were euthanized and decapitated to collect brain samples. The entire brain was homogenized in an ice bath using a glass homogenizer. Half of the homogenate (1.22 g) was used to assess AChE activity, while the remaining portion was preserved for oxidative stress and inflammatory parameter analyses. The AChE activity was assessed based on the previously published electrometric technique that was originally described by Michel [33] and subsequently refined to enhance its applicability across various animal species [34]. The technique is based on measuring the change in pH resulting from the enzymatic hydrolysis of acetylthiocholine iodide, providing a straightforward and effective approach for evaluating AChE activity in biological samples [35]. This method has been extensively validated for assessing AChE activity in chickens, demonstrating its reliability in toxicological studies [36,37,38]. Briefly, the barbital–phosphate buffer solution (BPB) was made by combining 6 mM sodium barbital, 600 mM sodium chloride, and 4.63 mM potassium dihydrogen phosphate. Hydrochloric acid was then added to adjust the pH level to 8.1. Three mL of BPB solution for every 100 mg of brain wet weight was utilized in brain homogenization. The reaction mixture was then prepared, which consisted of 3 mL distilled water, 0.2 mL brain homogenate, and 3 mL BPB. The pH of the mixture was then measured (pH 1). The mixture was then combined with 0.10 mL acetylthiocholine iodide (259 mM), which served as the AChE substrate. Next, the mixture was incubated at 37 °C for 30 min, and the pH of the reaction mixture was measured at the end of incubation (pH 2) [38].
The AChE activity in units of ∆pH was calculated as follows:
∆pH = (pH1 − pH2) − ∆pH of the blank.
The blank contains all the reaction mixture but not brain homogenate.
The percentage of AChE inhibition was calculated as follows:
%AChE inhibition = AChE activity of the control group − AChE activity of the AFB1-intoxicated group/AChE activity of the control group × 100
4.8. Assessment of Oxidative Stress Indicators in the Brain
Brain samples from the control and AFB1-treated groups were homogenized in a 0.9% NaCl solution and centrifuged at 3000× g for five minutes. Oxidative stress was evaluated by measuring GSH, SOD, TAC, and MDA levels using commercial kits, following the manufacturer’s instructions.
4.9. Gene Expression Related to Pro-Inflammatory Cytokines
The expression level of four pro-inflammatory cytokine genes in the chickens’ brains, including interleukin-1 beta (IL-1β), interleukin-6 (IL-6), interleukin-17 (IL-17), and inducible nitric oxide synthase (iNOS), was compared by performing the quantitative reverse transcription polymerase chain reaction (RT-qPCR). Total RNA was extracted using TRIzolTM according to the manufacturer’s instructions. It was then reverse-transcribed into cDNA using qScript cDNA SuperMix in line with standard procedures. The RT-qPCR amplification was conducted using the PowerUp™ SYBRTM Green Master Mix using the primers that are specified in Table 1. The relative expression per transcript was determined by normalizing to 28 S rRNA as the reference gene and comparing the control and AFB1-treated samples utilizing the 2−ΔΔCT method [39].

Table 1.
Primer sequences used in RT-qPCR.
4.10. Statistical Analysis
Parametric data, confirmed to follow a normal distribution, were analyzed using GraphPad Prism 10.4.1. An unpaired t-test was performed to compare groups and identify significant differences. Statistical significance was set at p < 0.05.
5. Conclusions
This study demonstrates that AFB1-contaminated feed significantly alters neurobehavioral outcomes in chickens, accompanied by biochemical changes in the brain. Specifically, AFB1 exposure led to reduced locomotor activity, exploratory behavior, and an increased tonic immobility response. Biochemically, to our knowledge, we present the first evidence that AFB1 exposure significantly decreases whole-brain AChE activity in chickens, indicating impaired cholinergic neurotransmission. Markers of oxidative stress revealed reduced levels of GSH and SOD, decreased TAC, and elevated MDA levels, signifying heightened oxidative stress in chicken brains. Furthermore, AFB1 exposure induced a clear inflammatory response in the brain, evidenced by the upregulation of pro-inflammatory cytokine genes, including IL-1β, IL-6, IL-17, and iNOS. These findings highlight the potential mechanisms underlying AFB1’s neurotoxic effects in the chicken brain and provide a foundation for further in-depth exploration of AFB1’s diverse effects in the avian brain.
Author Contributions
Conceptualization, W.H.S. and N.K.H.A.; methodology, W.H.S. and H.A.-K.; formal analysis, W.H.S. and N.K.H.A.; investigation, W.H.S. and H.A.-K.; data curation, W.H.S. and N.K.H.A.; writing—original draft preparation, W.H.S.; writing—review and editing, W.H.S., N.K.H.A. and H.A.-K.; visualization, W.H.S.; supervision, W.H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data that support this study will be shared upon reasonable request to the corresponding author.
Acknowledgments
The author expresses appreciation to the deanery of the College of Veterinary Medicine at the University of Al-Qadisiyah and the deanery of the College of Pharmacy at Alayen Iraqi University for their administrative and technical support in accomplishing this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Fouché, T.; Claassens, S.; Maboeta, M. Aflatoxins in the Soil Ecosystem: An Overview of Its Occurrence, Fate, Effects and Future Perspectives. Mycotoxin Res. 2020, 36, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.M.; Ruan, D.; El Senousey, H.A.K.; Chen, W.; Jiang, S.; Zheng, C. Harmful Effects and Control Strategies of Aflatoxin B1 Produced by Aspergillus Flavus and Aspergillus Parasiticus Strains on Poultry: Review. Toxins 2019, 11, 176. [Google Scholar] [CrossRef]
- Abrehame, S.; Manoj, V.R.; Hailu, M.; Chen, Y.Y.; Lin, Y.C.; Chen, Y.P. Aflatoxins: Source, Detection, Clinical Features and Prevention. Processes 2023, 11, 204. [Google Scholar] [CrossRef]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin Contamination of Commercial Maize Products during an Outbreak of Acute Aflatoxicosis in Eastern and Central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef] [PubMed]
- Yunus, A.W.; Razzazi-Fazeli, E.; Bohm, J. Aflatoxin B1 in Affecting Broiler’s Performance, Immunity, and Gastrointestinal Tract: A Review of History and Contemporary Issues. Toxins 2011, 3, 566–590. [Google Scholar] [CrossRef]
- Alharthi, A.S.; Al Sulaiman, A.R.; Aljumaah, R.S.; Alabdullatif, A.A.; Elolimy, A.A.; Alqhtani, A.H.; Al-Garadi, M.A.; Abudabos, A.M. Protective Effect of Date Pits on Growth Performance, Carcass Traits, Blood Indices, Intestinal Morphology, Nutrient Digestibility, and Hepatic Aflatoxin Residues of Aflatoxin B1-Exposed Broilers. Agriculture 2022, 12, 476. [Google Scholar] [CrossRef]
- Altyar, A.E.; Kensara, O.A.; Sayed, A.A.; Aleya, L.; Almutairi, M.H.; Zaazouee, M.S.; Elshanbary, A.A.; El-Demerdash, F.M.; Abdel-Daim, M.M. Acute Aflatoxin B1-Induced Hepatic and Cardiac Oxidative Damage in Rats: Ameliorative Effects of Morin. Heliyon 2023, 9, e21837. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of Oral Exposure to Aflatoxin B1-Induced Renal Dysfunction, Oxidative Stress, and Cell Apoptosis in Mice Kidney by Curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef]
- Subramaniam, S.; Sabran, M.R.; Stanslas, J.; Kirby, B.P. Effect of Aflatoxin B1 Exposure on the Progression of Depressive-like Behavior in Rats. Front. Nutr. 2022, 9, 1032810. [Google Scholar] [CrossRef]
- Dai, C.; Tian, E.; Li, H.; Gupta, S.D.; Hao, Z.; Wang, Z.; Velkov, T.; Shen, J. Molecular Mechanisms of Aflatoxin Neurotoxicity and Potential Neuroprotective Agents. Food Sci. Hum. Wellness 2023, 13, 2445–2455. [Google Scholar] [CrossRef]
- Cao, W.; Yu, P.; Yang, K.P.; Cao, D. Aflatoxin B1: Metabolism, Toxicology, and Its Involvement in Oxidative Stress and Cancer Development. Toxicol. Mech. Methods 2022, 32, 395–419. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.L.; Williams, D.E.; Coulombe, R.A. Species Differences in the Biotransformation of Aflatoxin B1: Primary Determinants of Relative Carcinogenic Potency in Different Animal Species. Toxins 2025, 17, 30. [Google Scholar] [CrossRef]
- Park, S.; Lee, J.Y.; You, S.; Song, G.; Lim, W. Neurotoxic Effects of Aflatoxin B1 on Human Astrocytes in Vitro and on Glial Cell Development in Zebrafish in Vivo. J. Hazard. Mater. 2020, 386, 121639. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Chen, Q.; Wang, L.; Gao, X.; Zhu, W.; Mu, P.; Deng, Y. Aflatoxin B1 Induces Neurotoxicity through Reactive Oxygen Species Generation, Dna Damage, Apoptosis, and s-Phase Cell Cycle Arrest. Int. J. Mol. Sci. 2020, 21, 6517. [Google Scholar] [CrossRef]
- Song, C.; Wang, Z.; Cao, J.; Dong, Y.; Chen, Y. Neurotoxic Mechanisms of Mycotoxins: Focus on Aflatoxin B1 and T-2 Toxin. Environ. Pollut. 2024, 356, 124359. [Google Scholar] [CrossRef]
- Pohanka, M. Cholinesterases, a Target of Pharmacology and Toxicology. Biomed. Pap. 2011, 155, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Liu, S.B.; Zhang, Q.; Tan, H.Z. Effects of Aflatoxin B1 on Growth Performance, Carcass Traits, Organ Index, Blood Biochemistry and Oxidative Status in Chinese Yellow Chickens. J. Vet. Med. Sci. 2023, 85, 1015–1022. [Google Scholar] [CrossRef]
- Aytekin Sahin, G.; Karabulut, D.; Unal, G.; Sayan, M.; Sahin, H. Effects of Probiotic Supplementation on Very Low Dose AFB1-Induced Neurotoxicity in Adult Male Rats. Life Sci. 2022, 306, 120798. [Google Scholar] [CrossRef]
- Braga, A.C.M.; Souto, N.S.; Cabral, F.L.; Dassi, M.; Rosa, É.V.F.; Guarda, N.D.S.; Royes, L.F.F.; Fighera, M.R.; Moresco, R.N.; Oliveira, M.S. Intermittent Exposure to Aflatoxin B1 Did Not Affect Neurobehavioral Parameters and Biochemical Markers of Oxidative Stress. Brain Sci. 2023, 13, 386. [Google Scholar] [CrossRef]
- Coulombe Jr, R.A.; Sharma, R.P. Effect of Repeated Dietary Exposure of Aflatoxin B1 on Brain Biogenic Amines and Metabolites in the Rat. Toxicol. Appl. Pharmacol. 1985, 80, 496–501. [Google Scholar] [CrossRef]
- Linardaki, Z.I.; Lamari, F.N.; Margarity, M. Saffron (Crocus sativus L.) Tea Intake Prevents Learning/Memory Defects and Neurobiochemical Alterations Induced by Aflatoxin B1 Exposure in Adult Mice. Neurochem. Res. 2017, 42, 2743–2754. [Google Scholar] [CrossRef] [PubMed]
- Cometa, M.F.; Lorenzini, P.; Fortuna, S.; Volpe, M.T.; Meneguz, A.; Palmery, M. In Vitro Inhibitory Effect of Aflatoxin B 1 on Acetylcholinesterase Activity in Mouse Brain. Toxicology 2005, 206, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.C.R.; Hawkins, E.; Jahr, F.M.; McClay, J.L.; Deshpande, L.S. Repeated Exposure to Chlorpyrifos Is Associated with a Dose-Dependent Chronic Neurobehavioral Deficit in Adult Rats. Neurotoxicology 2022, 90, 172–183. [Google Scholar] [CrossRef]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.-H.; Liu, W.-C. Marine Algal Polysaccharides Alleviate Aflatoxin B1-Induced Bursa of Fabricius Injury by Regulating Redox and Apoptotic Signaling Pathway in Broilers. Poult. Sci. 2021, 100, 844–857. [Google Scholar] [CrossRef]
- Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Licata, P.; Crupi, R. Evaluation of Neuroprotective Effects of Quercetin against Aflatoxin B1-Intoxicated Mice. Animals 2020, 10, 898. [Google Scholar] [CrossRef]
- Mehrzad, J.; Malvandi, A.M.; Alipour, M.; Hosseinkhani, S. Environmentally Relevant Level of Aflatoxin B1 Elicits Toxic Pro-Inflammatory Response in Murine CNS-Derived Cells. Toxicol. Lett. 2017, 279, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Sun, X.; Wang, H.; Li, W.; Wang, X.; Cheng, P.; Li, S.; Zhang, X.; Hamid, S. Curcumin Successfully Inhibited the Computationally Identified CYP2A6 Enzyme-Mediated Bioactivation of Aflatoxin B1 in Arbor Acres Broiler. Front. Pharmacol. 2017, 8, 143. [Google Scholar] [CrossRef]
- Li, S.; Han, M.; Zhang, Y.; Ishfaq, M.; Liu, R.; Wei, G.; Zhang, X.; Zhang, X. Effect of Curcumin as Feed Supplement on Immune Response and Pathological Changes of Broilers Exposed to Aflatoxin B1. Biomolecules 2022, 12, 1188. [Google Scholar] [CrossRef]
- Fernandez, A.; Verde, M.T.; Gascon, M.; Ramos, J.; Gomez, J.; Luco, D.F.; Chavez, G. Variations of Clinical Biochemical Parameters of Laying Hens and Broiler Chickens Fed Aflatoxin-Containing Feed. Avian Pathol. 1994, 23, 37–47. [Google Scholar] [CrossRef]
- Hernandez, E.; James, F.; Torrey, S.; Widowski, T.; Schwean-Lardner, K.; Monteith, G.; Turner, P.V. Evaluation of Brain Death in Laying Hens During On-Farm Killing by Cervical Dislocation Methods or Pentobarbital Sodium Injection. Front. Vet. Sci. 2019, 6, 297. [Google Scholar] [CrossRef]
- Mohammad, F.K.; Faris, G.A.M. Behavioral Effects of Acute Manganese Chloride Administration in Chickens. Biol. Trace Elem. Res. 2006, 110, 265–273. [Google Scholar] [CrossRef]
- Alnuaimi, S.I.; Al-Abdaly, Y.Z. Neurobehavioral Toxicity of Copper Sulfate Accompanied by Oxidative Stress and Histopathological Alterations in Chicks’ Brain. Iraqi J. Vet. Sci. 2023, 37, 53–60. [Google Scholar] [CrossRef]
- Michel, H.O. An Electrometric Method for the Determination of Red Blood Cell and Plasma Cholinesterase Activity. J. Lab. Clin. Med. 1949, 34, 1564–1568. [Google Scholar]
- Mohammad, F.K.; Faris, G.A.; Al-Kassim, N.A. A Modified Electrometric Method for Measurement of Erythrocyte Acetylcholinesterase Activity in Sheep. Vet. Hum. Toxicol. 1997, 39, 337–339. [Google Scholar] [PubMed]
- Mohammad, F.K.; St Omer, V.E. Modifications of Michel’s Electrometric Method for Rapid Measurement of Blood Cholinesterase Activity in Animals: A Minireview. Vet. Hum. Toxicol. 1982, 24, 119–121. [Google Scholar] [PubMed]
- Alias, A.S. The Use of an Electrometric Method for Measurement of Cholinesterase Activity in Plasma and Tissues of Local Doves. In Proceedings of the 11th Scientific Congress, Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt, 5–7 December 2004; Volume 1, pp. 241–259. [Google Scholar]
- Abass, K.S.; Mohammad, F.K. Validation of an Electrometric Method for Cholinesterase Measurement in the Plasma and Tissues of the Chicken. In Proceedings of the 11th Scientific Congress, Faculty of Veterinary Medicine, Assiut University, Assiut, Egypt, 5–7 December 2004; Volume 1, pp. 241–259. [Google Scholar]
- Al-Badrany, Y.M.A.; Mohammad, F.K. Effects of Acute and Repeated Oral Exposure to the Organophosphate Insecticide Chlorpyrifos on Open-Field Activity in Chicks. Toxicol. Lett. 2007, 174, 110–116. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Salvador, C.; Cortes, A.L.; Pandiri, A.R.; Gimeno, I.M. Cytokine Expression in the Eye and Brain of Chickens Following Infection with a Very Virulent plus Marek’s Disease Virus Strain. Vet. Immunol. Immunopathol. 2021, 237, 110277. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, J.; Jiang, C.; Cui, J.; Hong, L.; Hao, Z.; Tang, Y.; Liu, Y.; Cui, X.; Teng, X. Se Alleviated Pb-Caused Neurotoxicity in Chickens: SPS2-GPx1-GSH-IL-2/IL-17-NO Pathway, Selenoprotein Suppression, Oxidative Stress, and Inflammatory Injury. Antioxidants 2024, 13, 370. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).