
Exploring Seed Priming as a Strategy for Enhancing Abiotic Stress Tolerance in Cereal Crops

 , ,
, ,  and
and
Abstract
1. Introduction
2. Overview of Seed Priming
3. Abiotic Stresses in Cereal Crops
4. Seed Priming Applications in Cereal Crops for Mitigating Environmental Stresses
4.1. Role of SP in Cereals Exposed to Salt Stress
4.2. Role of SP in Cereals Exposed to Drought
4.3. Seed Priming as a Strategy to Mitigate Heavy Metal Stress in Cereals

{kind=link}
{kind=link}
| Stress Type | Priming Method | Seed Species | Effects | Ref. |
|---|---|---|---|---|
| Salt stress | Chitosan priming | Triticum durum L. | -Improved germination percentage, seedling length, and fresh/dry mass. -Increased GABA shunt metabolites (GABA, glutamate, alanine). -Reduced oxidative damage (lower MDA levels than untreated seeds). | [87] |
| Chitosan nanoparticles and chitosan priming | Oryza sativa L. | -Enhanced germination potential and seedling vigor. -Improved morphological, physiological, and biochemical responses. -Increased antioxidant activity. | [73] | |
| Ascorbic acid, potassium silicate, proline, spermidine, and saline water | Triticum durum L. | -Enhanced germination percentage and rate. | [71] | |
| Salicylic acid | Zea mays L. | -Improved growth, root dry matter, leaf relative water content, and free proline content. | [88] | |
| Salicylic acid (SA), Gibberellic acid (GA), and Sodium chloride | Hordeum vulgare L. | SA: Improved antioxidant defense mechanisms; -Increased proline, sugar, and ascorbic acid levels. -Reduced ROS accumulation and lipid peroxidation. -Supported seed adaptation to salinity stress. GA: Most effective for improving germination, shoot/root growth, and photosynthesis under salinity. | [76] | |
| Kinetin, Gibberellic acid, Iron, Auxin, and Potassium nitrate | Triticum aestivum L. | -Improved germination rate and seedling growth under salt stress. -Enhanced coleoptile and radicle growth. -Stimulated root development and seedling growth. | [89] | |
| Osmopriming with Ca2+ and K+ | Chenopodium quinoa | -Enhanced salinity tolerance through antioxidant enzyme activation (CAT, APX, SOD, GPX, PPO). -Increased antioxidant metabolites (phenolics, flavonoids, ascorbic acid), proline, glycine betaine, soluble carbohydrates. -Improved K+/Na+ homeostasis and Na+ exclusion. | [90] | |
| Zinc oxide nanoparticles + 24-epibrassinolide | Zea mays L. | -Improved root length, root surface area, stem diameter, relative leaf water content, total chlorophyll content, photosynthetic rate, and uptake of Zn and K+. -Reduced Na+ accumulation and Na+/K+ ratio. | [91] | |
| Hydrogen peroxide | Zea mays L. | -Enhanced germination, growth, and physiological traits. -Improved antioxidant defense (APX, CAT, POD, AsA). -Increased leaf water status, soluble proteins, amino acids, proline, sugars, IAA, and GA. -Reduced MDA and H2O2 levels. -Limited Na+ and Cl− uptake while improving Ca2+, K+, and Mg2+ content. | [92] | |
| Glutathione + Zinc | Zea mays L. | -Improved germination and seedling emergence -Enhanced antioxidant defense. -Reduced ROS and MDA levels. -Improved nutrient uptake (K+, Ca2+), and reduced Na+ accumulation and toxicity. -Increased K+/Na+ and Ca2+/Na+ ratios | [93] | |
| Biopriming with Bacillus sp. | Oryza sativa L. | -Enhanced germination percentage, seedling growth, and photosynthetic pigment content. | [94] | |
| Priming with PVP-coated silver nanoparticles | Hordeum vulgare L. | -Restored seed germination under salt stress; -Reduced ROS accumulation. -Increased antioxidant enzymes (SOD, CAT, GR, GPX). -Upregulation of antioxidant genes (HvSOD, HvCAT, HvGR, HvGPX). | [95] | |
| Drought stress | Selenium | Chenopodium quinoa | -Increased main panicle length, panicle weight, and grain weight. -Enhanced gas exchange parameters (photosynthesis, stomatal conductance). -Increased chlorophyll content, total phenol content, and water relations. -Improved grain quality (P, K, proteins). | [96] |
| Biopriming with Trichoderma harzianum | Oryza sativa L. | -Improved drought tolerance by reducing leaf rolling, increasing chlorophyll content, leaf area index, membrane stability index, and relative water content. Proline levels were minimized. -Enhanced morphological, physiological, and biochemical responses and delayed drought effects during tillering stage. | [97] | |
| Biopriming with Bacillus subtilis | Triticum aestivum L. | -Improved germination and growth. -Increased chlorophyll a and b, carotenoids, water-holding capacity, and salicylic acid content were observed. -Reduced proline, lipid peroxidation, and electrolyte leakage. | [98] | |
| Zinc Oxide Nanoparticles | Triticum aestivum L. | -Enhanced antioxidant enzyme activities (POX, CAT, GR), total phenolics, flavonoids, and sugars under drought. -ROS detoxification was improved. | [99] | |
| γ-Aminobutyric acid (GABA) seed priming | Triticum aestivum L. | -Improved germination, seedling biomass, water content, and photosystem efficiency. -Boosted antioxidant enzyme activities, leaf free proline, glycine betaine, soluble phenolics, and endogenous GABA levels. | [100] | |
| Halopriming with NaCl | Zea mays L. | -Improved seed germination uniformity, speed, and overall vigor (increased germination percentage, index, and seedling dry weight). -Improved water use efficiency. | [101] | |
| Ethephon seed priming | Triticum aestivum L. | -Enhanced drought stress memory at the tillering stage by maintaining leaf water; -Decreased MDA levels; -Improved root-to-leaf ABA signaling, ROS scavenging, and osmotic regulation; -Upregulated genes in ethylene-mediated pathways (carbon metabolism, glutathione metabolism, phenylpropanoid biosynthesis). | [102] | |
| Hydropriming | Chenopodium quinoa Willd | -Enhanced growth and seed yield. -Increase pigments and proline contents. | [103] | |
| Osmopriming with KNO3, Mg(NO3)2, GA3, Hydropriming | Zea mays L. | -Improved the antioxidative defense system. -Improved morpho-physiological traits such as total chlorophyll, and chlorophyll a and b. -Increased proline and catalase activity and decreased MDA content. | [104] | |
| Copper nanoparticle priming | Zea mays L. | -Enhanced drought tolerance in maize by increasing leaf water content, biomass, and yield components. -Elevated anthocyanin, chlorophyll, and carotenoid contents. -ROS accumulation was reduced through activation of antioxidant enzymes. | [105] | |
| Silver oxide nanoparticle | Triticum aestivum L. | -Significant improvement in germination, seedling growth, and biomass under drought conditions; -Positive correlation between root length and other traits; -Increased chlorophyll content; -Identification of 261 single-nucleotide polymorphisms and key genes (TraesCS1A02G049700) linked to drought tolerance. | [106] | |
| Cobalt (Co) toxicity | ZnO nanoparticle priming | Zea mays L. | -Improved plant growth, biomass, and photosynthesis. -Reduced ROS and MDA levels, limited Co uptake, stabilized ultrastructures and photosynthetic machinery, and enhanced nutrient content and antioxidant enzyme activities. | [107] |
| Cadmium (Cd) toxicity | Multiwall carbon nanotubes (MWCNTs) priming | Zea mays L. | -Improved seed germination rate, root and shoot growth, and antioxidant enzyme activities (POD, SOD, CAT). | [108] |
| Bacillus subtilis NA2, Aspergillus niger PMI-118, and L-proline priming | Triticum aestivum L. | -Improved plant biomass, shoot length, root length, chlorophyll, total sugars, proteins, and ascorbic acid. -Reduced antioxidant enzyme activities (CAT, APX) and oxidative stress (H2O2). | [109] | |
| Copper (Cu) toxicity | Silicon, melatonin, salicylic acid, glycine betaine, and ascorbic acid priming | Triticum aestivum L. | -Inhanced plant growth, biomass, and photosynthetic traits. -Reduced oxidative stress (MDA, H2O2), and boosted antioxidant enzymes. -Improved proline metabolism, AsA-GSH cycle, and gene expression. | [110] |
| Copper (Cu) toxicity | Seed priming with silver ions | Oryza sativa L. | -Increased fresh biomass, reduced Cu content in roots and shoots, and improved nutrient uptake (Ca, Fe, Mg, Mn). -Improved root cell viability, maintained root morphology, reduced malondialdehyde accumulation, and activated key signaling pathways (MAPK, phytohormone) for defense response. | [111] |
| Arsenic (As) toxicity | Seed priming with zinc | Oryza sativa L. | -Restored seedling growth, reduce As uptake, and limite oxidative stress through modulating redox homeostasis. -Reduced ROS production and protected antioxidant enzymes. | [112] |
| Aluminum (Al) toxicity | Seed priming with 24-epibrassinolide | Oryza sativa (rice) | -Enhanced seed germination, root and shoot length, and biomass. -Reduced MDA and H2O2 levels, enhanced antioxidant enzyme activities (SOD, CAT, APX), and improved photosynthetic pigments. | [113] |
| Lead (Pb) toxicity | Seed priming with calcium and salicylic acid | Triticum aestivum L. | -Alleviated oxidative stress, restored osmoprotectants, reduced Pb ion content, and enhanced antioxidant enzyme activities. They also downregulated genes overexpressed under Pb stress, suggesting protective mechanisms. | [114] |
| Lead (Pb) toxicity | Seed priming with nano–graphene oxide, nano–molybdenum, nano–selenium, nano–zinc oxide, and nano–silica. | Hordeum vulgare L. | -Improved plant growth and biomass. -Reduced oxidative stress, decreased MDA and H2O2 accumulation, enhanced enzymatic and non-enzymatic antioxidants, and supported better gas exchange and gene expression. | [115] |
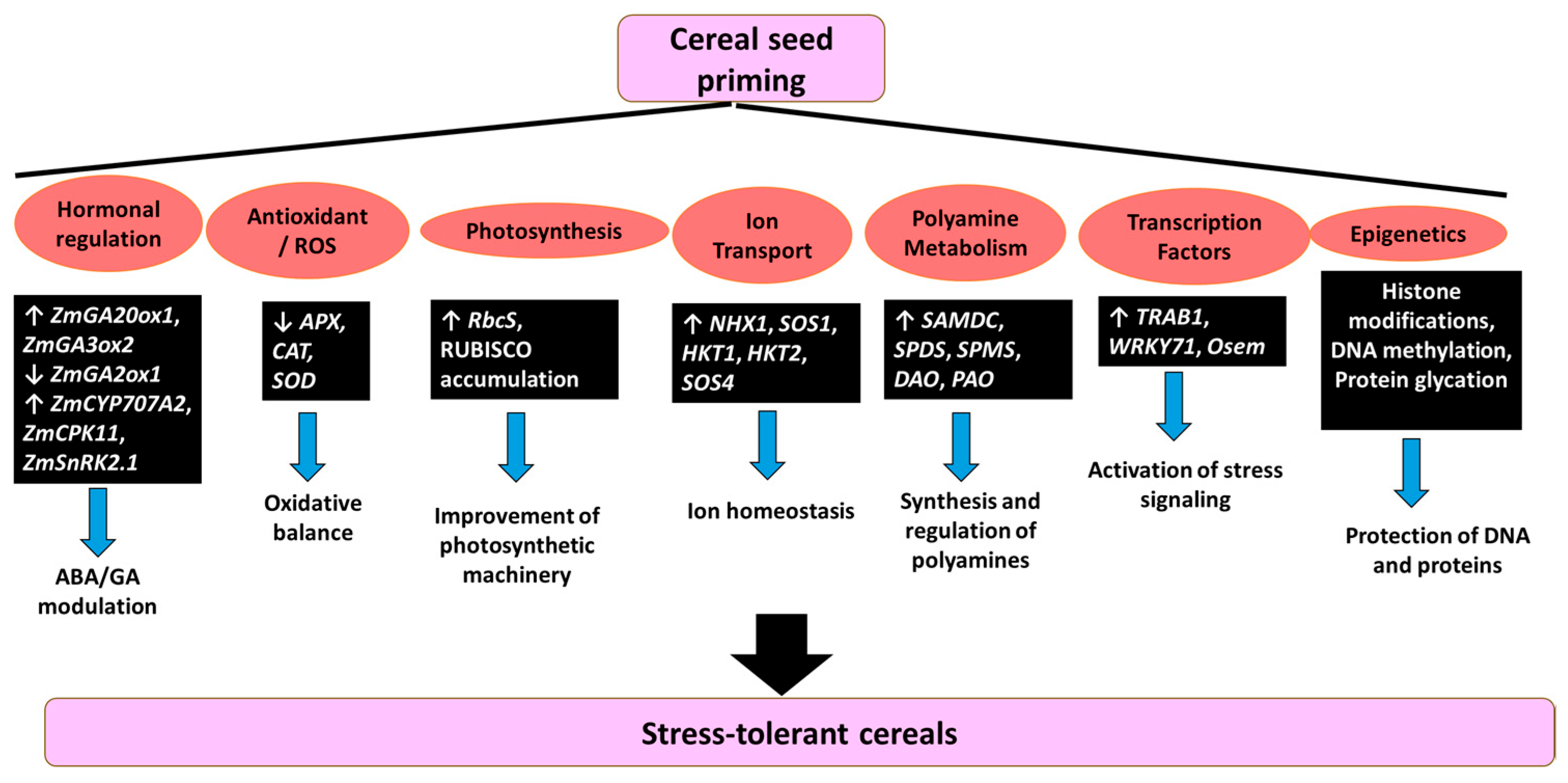
5. Molecular Signaling Pathways and Gene Regulatory Networks Mediated by Seed Priming in Cereal Crops to Enhance Stress Tolerance
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization. Agricultural Production Statistics 2010–2023; Food and Agriculture Organization: Rome, Italy, 2024. [Google Scholar]
- Guo, H.; Wu, H.; Sajid, A.; Li, Z. Whole Grain Cereals: The Potential Roles of Functional Components in Human Health. Crit. Rev. Food Sci. Nutr. 2022, 62, 8388–8402. [Google Scholar] [CrossRef]
- Nahi, O.; Siankevich, S. Upcycling of Cereal Byproducts: A Sustainable Opportunity to Valorize Wasted Nutrients and Derive Bioactive Compounds for Humans and Animals Nutrition and Health. Chimia 2023, 77, 858–866. [Google Scholar] [CrossRef]
- Noort, M.W.J.; Renzetti, S.; Linderhof, V.; du Rand, G.E.; Marx-Pienaar, N.J.M.M.; de Kock, H.L.; Magano, N.; Taylor, J.R.N. Towards Sustainable Shifts to Healthy Diets and Food Security in Sub-Saharan Africa with Climate-Resilient Crops in Bread-Type Products: A Food System Analysis. Foods 2022, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Jeyasri, R.; Muthuramalingam, P.; Satish, L.; Pandian, S.K.; Chen, J.T.; Ahmar, S.; Wang, X.; Mora-Poblete, F.; Ramesh, M. An Overview of Abiotic Stress in Cereal Crops: Negative Impacts, Regulation, Biotechnology and Integrated Omics. Plants 2021, 10, 1472. [Google Scholar] [CrossRef]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Alam, M.A.; Syed, M.A.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P.; et al. Consequences and Mitigation Strategies of Abiotic Stresses in Wheat (Triticum aestivum L.) under the Changing Climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]
- Mthiyane, P.; Aycan, M.; Mitsui, T. Strategic Advancements in Rice Cultivation: Combating Heat Stress through Genetic Innovation and Sustainable Practices—A Review. Stresses 2024, 4, 452–480. [Google Scholar] [CrossRef]
- Sharma, V.; Garg, N. Organic Solutes in Cereals Under Abiotic Stress. In Sustainable Remedies for Abiotic Stress in Cereals; Springer Nature: Berlin/Heidelberg, Germany, 2022; pp. 29–50. [Google Scholar] [CrossRef]
- Hameed, A.; Hussain, S.; Nisar, F.; Rasheed, A.; Shah, S.Z. Seed Priming as an Effective Technique for Enhancing Salinity Tolerance in Plants: Mechanistic Insights and Prospects for Saline Agriculture with a Special Emphasis on Halophytes. Seeds 2025, 4, 14. [Google Scholar] [CrossRef]
- Thakur, M.; Sharma, P.; Anand, A. Seed priming-induced early vigor in crops: An alternate strategy for abiotic stress tolerance. In Priming and Pretreatment of Seeds and Seedlings: Implication in Plant Stress Tolerance and Enhancing Productivity in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2019; pp. 163–180. [Google Scholar]
- Ali, K.; Mubasher, H.M.; Sher, A.; Sattar, A.; Manaf, A. Seed Priming for Abiotic Stress Tolerance. In Climate-Resilient Agriculture; Springer International Publishing: Berlin/Heidelberg, Germany, 2023; Volume 2, pp. 641–665. [Google Scholar] [CrossRef]
- Jatana, B.S.; Grover, S.; Ram, H.; Baath, G.S. Seed Priming: Molecular and Physiological Mechanisms Underlying Biotic and Abiotic Stress Tolerance. Agronomy 2024, 14, 2901. [Google Scholar] [CrossRef]
- Liu, X.; Quan, W.; Bartels, D. Stress Memory Responses and Seed Priming Correlate with Drought Tolerance in Plants: An Overview. Planta 2022, 255, 1–14. [Google Scholar] [CrossRef]
- Koushal, S.; Mankar, A.A.; Anbarasan, S.; Kumar, V.; Kumari, J.; Nagarjuna, S.; Jahan, R.; Kishan Kumar, R.; Satapathy, S.N. Mechanism and Methodologies of Seed Priming: Enhancing Germination and Crop Resilience. Plant Cell Biotechnol. Mol. Biol. 2024, 25, 185–194. [Google Scholar] [CrossRef]
- Kubala, S.; Lechowska, K.; Wojtyla, L.; Kosmala, A.; Quinet, M.; Lutts, S.; Garnczarska, M. Transcriptome and proteome changes accompanying increased vigor of osmoprimed rape (Brassica napus L.) seeds. BioTechnologia. J. Biotechnol. Comput. Biol. Bionanotechnol. 2013, 3, 94. [Google Scholar]
- El-Maarouf-bouteau, H. The Seed and the Metabolism Regulation. Biology 2022, 11, 168. [Google Scholar] [CrossRef]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-Still a Mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Jacobsen, J.V.; Gubler, F.; Chandler, P.M. Gibberellin Action in Germinated Cereal Grains. In Plant Hormones; Springer: Berlin/Heidelberg, Germany, 1995; pp. 246–271. [Google Scholar] [CrossRef]
- Ma, Z.; Bykova, N.V.; Igamberdiev, A.U. Cell Signaling Mechanisms and Metabolic Regulation of Germination and Dormancy in Barley Seeds. Crop J. 2017, 5, 459–477. [Google Scholar] [CrossRef]
- Farooq, M.; Usman, M.; Nadeem, F.; Rehman, H.U.; Wahid, A.; Basra, S.M.A.; Siddique, K.H.M. Seed Priming in Field Crops: Potential Benefits, Adoption and Challenges. Crop Pasture Sci. 2019, 70, 731–771. [Google Scholar] [CrossRef]
- Corbineau, F.; Taskiran-Özbingöl, N.; El-Maarouf-Bouteau, H. Improvement of Seed Quality by Priming: Concept and Biological Basis. Seeds 2023, 2, 101–115. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Tania, S.S.; Imran, S.; Rauf, F.; Kibria, M.G.; Ye, W.; Hasanuzzaman, M.; Murata, Y. Seed Priming with Nanoparticles: An Emerging Technique for Improving Plant Growth, Development, and Abiotic Stress Tolerance. J. Soil. Sci. Plant Nutr. 2022, 22, 4047–4062. [Google Scholar] [CrossRef]
- Yadav, N.; Bora, S.; Devi, B.; Upadhyay, C.; Singh, P. Nanoparticle-Mediated Defense Priming: A Review of Strategies for Enhancing Plant Resilience against Biotic and Abiotic Stresses. Plant Physiol. Biochem. 2024, 213, 108796. [Google Scholar] [CrossRef]
- Singh, H.; Jassal, R.K.; Kang, J.S.; Sandhu, S.S.; Kang, H.; Grewal, K. Seed Priming Techniques in Field Crops—A Review. Agric. Rev. 2015, 36, 251–264. [Google Scholar] [CrossRef]
- Afzal, S.; Sharma, D.; Singh, N.K. Eco-Friendly Synthesis of Phytochemical-Capped Iron Oxide Nanoparticles as Nano-Priming Agent for Boosting Seed Germination in Rice (Oryza sativa L.). Environ. Sci. Pollut. Res. 2021, 28, 40275–40287. [Google Scholar] [CrossRef]
- Sharma, D.; Afzal, S.; Singh, N.K. Nanopriming with Phytosynthesized Zinc Oxide Nanoparticles for Promoting Germination and Starch Metabolism in Rice Seeds. J. Biotechnol. 2021, 336, 64–75. [Google Scholar] [CrossRef]
- Batool, R.; Umer, M.J.; Shabbir, M.Z.; Wang, Y.; Ahmed, M.A.; Guo, J.; He, K.; Zhang, T.; Bai, S.; Chen, J.; et al. Seed Myco-Priming Improves Crop Yield and Herbivory Induced Defenses in Maize by Coordinating Antioxidants and Jasmonic Acid Pathway. BMC Plant Biol. 2022, 22, 1–17. [Google Scholar] [CrossRef]
- Biswas, S.; Seal, P.; Majumder, B.; Biswas, A.K. Efficacy of Seed Priming Strategies for Enhancing Salinity Tolerance in Plants: An Overview of the Progress and Achievements. Plant Stress. 2023, 9, 100186. [Google Scholar] [CrossRef]
- Chen, S.; Liu, H.; Yangzong, Z.; Gardea-Torresdey, J.L.; White, J.C.; Zhao, L. Seed Priming with Reactive Oxygen Species-Generating Nanoparticles Enhanced Maize Tolerance to Multiple Abiotic Stresses. Environ. Sci. Technol. 2023, 57, 19932–19941. [Google Scholar] [CrossRef]
- Faisal, S.; Muhammad, S.; Luqman, M.; Hasnain, M.; Rasool, A.; Awan, M.U.F.; Khan, Z.I.; Hussain, I. Effects of Priming on Seed Germination, Physico-Chemistry and Yield of Late Sown Wheat Crop (Triticum aestivum L.). Pol. J. Environ. Stud. 2023, 32, 1113–1124. [Google Scholar] [CrossRef]
- Janah, I.; Elhasnaoui, A.; Abouloifa, H.; Ait-El-Mokhtar, M.; Ben Laouane, R. Hormonal Priming to Increase Germination of Stevia rebaudiana Bertoni Seeds in Saline Environments. Int. J. Plant Biol. 2025, 16, 2. [Google Scholar] [CrossRef]
- Khalequzzaman; Ullah, H.; Himanshu, S.K.; Islam, N.E.T.; Tisarum, R.; Cha-um, S.; Datta, A. Seed Priming Improves Germination, Yield, and Water Productivity of Cotton Under Drought Stress. J. Soil. Sci. Plant Nutr. 2023, 23, 2418–2432. [Google Scholar] [CrossRef]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed Priming: State of the Art and New Perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed Priming for Abiotic Stress Tolerance: An Overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Sen, A.; Puthur, J.T. Influence of Different Seed Priming Techniques on Oxidative and Antioxidative Responses during the Germination of Oryza sativa Varieties. Physiol. Mol. Biol. Plants 2020, 26, 551–565. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Gao, G.; Ali, I.; Wu, X.; Tang, M.; Chen, L.; Jiang, L.; Liang, T. Effects of Various Seed Priming on Morphological, Physiological, and Biochemical Traits of Rice under Chilling Stress. Front. Plant Sci. 2023, 14, 1146285. [Google Scholar] [CrossRef]
- Subedi, R.; Maharjan, B.; Adhikari, R. Effect of Different Priming Methods in Rice (Oryza sativa). J. Agric. Environ. 2015, 16, 152–160. [Google Scholar] [CrossRef]
- Nawaz, F.; Ashraf, M.Y.; Ahmad, R.; Waraich, E.A. Selenium (Se) Seed Priming Induced Growth and Biochemical Changes in Wheat under Water Deficit Conditions. Biol. Trace Elem. Res. 2013, 151, 284–293. [Google Scholar] [CrossRef]
- Alam, A.U.; Ullah, H.; Himanshu, S.K.; Tisarum, R.; Cha-um, S.; Datta, A. Seed Priming Enhances Germination and Morphological, Physio-Biochemical, and Yield Traits of Cucumber under Water-Deficit Stress. J. Soil. Sci. Plant Nutr. 2023, 23, 3961–3978. [Google Scholar] [CrossRef]
- Sun, H.; Li, L.; Wang, X.; Wu, S.; Wang, X. Ascorbate-Glutathione Cycle of Mitochondria in Osmoprimed Soybean Cotyledons in Response to Imbibitional Chilling Injury. J. Plant Physiol. 2011, 168, 226–232. [Google Scholar] [CrossRef]
- Jisha, K.C.; Puthur, J.T. Seed Priming with BABA (β-Amino Butyric Acid): A Cost-Effective Method of Abiotic Stress Tolerance in Vigna radiata (L.) Wilczek. Protoplasma 2016, 253, 277–289. [Google Scholar] [CrossRef]
- Thornton, J.M.; Collins, A.R.S.; Powell, A.A. The Effect of Aerated Hydration on DNA Synthesis in Embryos of Brassica oleracea L. Seed Sci. Res. 1993, 3, 195–199. [Google Scholar] [CrossRef]
- Jian, H.; Wang, J.; Wang, T.; Wei, L.; Li, J.; Liu, L. Identification of Rapeseed MicroRNAs Involved in Early Stage Seed Germination under Salt and Drought Stresses. Front. Plant Sci. 2016, 7, 658. [Google Scholar] [CrossRef]
- Oshunsanya, S.O.; Nwosu, N.J.; Li, Y. Abiotic Stress in Agricultural Crops Under Climatic Conditions. In Sustainable Agriculture, Forest and Environmental Management; Springer: Singapore, 2019; pp. 71–100. [Google Scholar] [CrossRef]
- Balfagón, D.; Zandalinas, S.I.; Gómez-Cadenas, A. High Temperatures Change the Perspective: Integrating Hormonal Responses in Citrus Plants under Co-Occurring Abiotic Stress Conditions. Physiol. Plant 2019, 165, 183–197. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H. Sustainable Remedies for Abiotic Stress in Cereals; Springer Nature: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Fahad, S.; Noor, M.; Adnan, M.; Khan, M.A.; Rahman, I.U.; Alam, M.; Nasim, W. Abiotic stress and rice grain quality. In Advances in Rice Research for Abiotic Stress Tolerance; Woodhead Publishing: Sawston, UK, 2019; pp. 571–583. [Google Scholar]
- Djanaguiraman, M.; Prasad, P.V.; Ciampitti, I.A.; Talwar, H.S. Impacts of abiotic stresses on sorghum physiology. In Sorghum in the 21st Century: Food–Fodder–Feed–Fuel for a Rapidly Changing World; Springer: Singapore, 2020; pp. 157–188. [Google Scholar]
- Bali, A.S.; Sidhu, G.P.S. Abiotic stress-induced oxidative stress in wheat. In Wheat Production in Changing Environments: Responses, Adaptation and Tolerance; Springer: Berlin/Heidelberg, Germany, 2019; pp. 225–239. [Google Scholar]
- Salika, R.; Riffat, J. Abiotic stress responses in maize: A review. Acta Physiol. Plant. 2021, 43, 130. [Google Scholar] [CrossRef]
- Li, P.; Yang, X.; Wang, H.; Pan, T.; Wang, Y.; Xu, Y.; Yang, Z. Genetic control of root plasticity in response to salt stress in maize. Theor. Appl. Genet. 2021, 134, 1475–1492. [Google Scholar] [CrossRef]
- Thiruvengadam, R.; Venkidasamy, B.; Easwaran, M.; Chi, H.Y.; Thiruvengadam, M.; Kim, S.H. Dynamic Interplay of Reactive Oxygen and Nitrogen Species (ROS and RNS) in Plant Resilience: Unveiling the Signaling Pathways and Metabolic Responses to Biotic and Abiotic Stresses. Plant Cell Rep. 2024, 43, 1–24. [Google Scholar] [CrossRef]
- Calixto, C.P.G. Molecular Aspects of Heat Stress Sensing in Land Plants. Plant J. 2025, 121, e70069. [Google Scholar] [CrossRef]
- Chakraborty, S.; Roychoudhury, A. Morphological, Architectural and Biochemical Modifications of Cereal Crops During Abiotic Stress. In Omics Approach to Manage Abiotic Stress in Cereals; Springer: Singapore, 2022; pp. 1–20. [Google Scholar] [CrossRef]
- Rane, J.; Singh, A.K.; Kumar, M.; Boraiah, K.M.; Meena, K.K.; Pradhan, A.; Vara Prasad, P.V. The Adaptation and Tolerance of Major Cereals and Legumes to Important Abiotic Stresses. Int. J. Mol. Sci. 2021, 22, 12970. [Google Scholar] [CrossRef]
- Shahzadi, A.; Noreen, Z.; Alamery, S.; Zafar, F.; Haroon, A.; Rashid, M.; Aslam, M.; Younas, A.; Attia, K.A.; Mohammed, A.A.; et al. Effects of Biochar on Growth and Yield of Wheat (Triticum aestivum L.) under Salt Stress. Sci. Rep. 2024, 14, 20024. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Javed, T.; Hussain, S.; Raza, A.; Shabbir, R.; Mora-Poblete, F.; Saeed, T.; Zulfiqar, F.; Ali, M.M.; et al. Screening of Wheat (Triticum aestivum L.) Genotypes for Drought Tolerance through Agronomic and Physiological Response. Agronomy 2022, 12, 287. [Google Scholar] [CrossRef]
- Nazir, A.; Rafique, F.; Ahmed, K.; Khan, S.A.; Khan, N.; Akbar, M.; Zafar, M. Evaluation of Heavy Metals Effects on Morpho-Anatomical Alterations of Wheat (Triticum aestivum L.) Seedlings. Microsc. Res. Tech. 2021, 84, 2517–2529. [Google Scholar] [CrossRef]
- Shahzad, H.; Ullah, S.; Iqbal, M.; Bilal, H.M.; Shah, G.M.; Ahmad, S.; Zakir, A.; Ditta, A.; Farooqi, M.A.; Ahmad, I. Salinity Types and Level-Based Effects on the Growth, Physiology and Nutrient Contents of Maize (Zea mays). Ital. J. Agron. 2019, 14, 199–207. [Google Scholar] [CrossRef]
- Cao, L.; Lu, X.; Wang, G.; Zhang, P.; Fu, J.; Wang, Z.; Wei, L.; Wang, T. Transcriptional Regulatory Networks in Response to Drought Stress and Rewatering in Maize (Zea mays L.). Mol. Genet. Genom. 2021, 296, 1203–1219. [Google Scholar] [CrossRef]
- Oudghiri, M.; Yamani, B.; Benlemlih, N.; El Aammouri, S.; Abid, N.; Brhadda, N.; Bouhassoun, S.; Ziri, R.; Chriqui, A.; Aoujil, F.Z.; et al. Effect of Heavy Metals on the Morphological and Physiological Responses of the Torro Plus Variant of Zea Mays. J. Environ. Earth Sci. 2025, 7, 165–179. [Google Scholar] [CrossRef]
- Torun, H.; Novák, O.; Mikulík, J.; Strnad, M.; Ayaz, F.A. The Effects of Exogenous Salicylic Acid on Endogenous Phytohormone Status in Hordeum vulgare L. under Salt Stress. Plants 2022, 11, 618. [Google Scholar] [CrossRef]
- Ferioun, M.; Srhiouar, N.; Bouhraoua, S.; El Ghachtouli, N.; Louahlia, S. Physiological and Biochemical Changes in Moroccan Barley (Hordeum vulgare L.) Cultivars Submitted to Drought Stress. Heliyon 2023, 9, e13643. [Google Scholar] [CrossRef]
- Selim, S.; Abuelsoud, W.; Al-Sanea, M.M.; AbdElgawad, H. Elevated CO2 Differently Suppresses the Arsenic Oxide Nanoparticles-Induced Stress in C3 (Hordeum vulgare) and C4 (Zea maize) Plants via Altered Homeostasis in Metabolites Specifically Proline and Anthocyanin Metabolism. Plant Physiol. Biochem. 2021, 166, 235–245. [Google Scholar] [CrossRef]
- Rodríguez Coca, L.I.; García González, M.T.; Gil Unday, Z.; Jiménez Hernández, J.; Rodríguez Jáuregui, M.M.; Fernández Cancio, Y. Effects of Sodium Salinity on Rice (Oryza sativa L.) Cultivation: A Review. Sustainability 2023, 15, 1804. [Google Scholar] [CrossRef]
- Bhandari, U.; Gajurel, A.; Khadka, B.; Thapa, I.; Chand, I.; Bhatta, D.; Poudel, A.; Pandey, M.; Shrestha, S.; Shrestha, J. Morpho-Physiological and Biochemical Response of Rice (Oryza sativa L.) to Drought Stress: A Review. Heliyon 2023, 9, e13744. [Google Scholar] [CrossRef]
- Bari, M.A.; El-Shehawi, A.M.; Elseehy, M.M.; Naheen, N.N.; Rahman, M.M.; Kabir, A.H. Molecular Characterization and Bioinformatics Analysis of Transporter Genes Associated with Cd-Induced Phytotoxicity in Rice (Oryza sativa L.). Plant Physiol. Biochem. 2021, 167, 438–448. [Google Scholar] [CrossRef]
- Azeem, M.; Sultana, R.; Mahmood, A.; Qasim, M.; Siddiqui, Z.S.; Mumtaz, S.; Javed, T.; Umar, M.; Adnan, M.Y.; Siddiqui, M.H. Ascorbic and Salicylic Acids Vitalized Growth, Biochemical Responses, Antioxidant Enzymes, Photosynthetic Efficiency, and Ionic Regulation to Alleviate Salinity Stress in Sorghum bicolor. J. Plant Growth Regul. 2023, 42, 5266–5279. [Google Scholar] [CrossRef]
- Amoah, J.N.; Antwi-Berko, D. Comparative Physiological, Biochemical and Transcript Response to Drought in Sorghum Genotypes. Biotechnol. J. Int. 2020, 24, 1–14. [Google Scholar] [CrossRef]
- Singh, A.; Parihar, S.; Shekhawat, G.S. Cd-Induced Cytotoxicity and Its HO-1 and ROS Quenching Enzyme-Mediated Regulation in 2–3 Leaf Stage Seedlings of Sorghum Bicolor: An Important Millet Crop of the Arid & Semi-Arid Regions. J. Trace Elem. Miner. 2024, 9, 100165. [Google Scholar] [CrossRef]
- Feghhenabi, F.; Hadi, H.; Khodaverdiloo, H.; van Genuchten, M.T. Seed Priming Alleviated Salinity Stress during Germination and Emergence of Wheat (Triticum aestivum L.). Agric. Water Manag. 2020, 231, 106022. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, R.; Xing, Y.; Jiang, B.; Li, B.; Xu, X.; Zhou, Y. The Efficacy of Different Seed Priming Agents for Promoting Sorghum Germination under Salt Stress. PLoS ONE 2021, 16, e0245505. [Google Scholar] [CrossRef] [PubMed]
- Soni, A.T.; Rookes, J.E.; Arya, S.S. Chitosan Nanoparticles as Seed Priming Agents to Alleviate Salinity Stress in Rice (Oryza sativa L.) Seedlings. Polysaccharides 2023, 4, 129–141. [Google Scholar] [CrossRef]
- Yang, X.; Xiong, J.; Du, X.; Sun, M.; Ding, L.; Mei, W.; Sun, Z.; Feng, N.; Zheng, D.; Shen, X. Effects of Exogenous Spermidine on Seed Germination and Physiological Metabolism of Rice Under NaCl Stress. Plants 2024, 13, 3599. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Kamal, A.; Singh, A.; Ashfaque, F.; Alamri, S.; Siddiqui, M.H.; Khan, M.I.R. Seed priming with gibberellic acid induces high salinity tolerance in Pisum sativum through antioxidants, secondary metabolites and up-regulation of antiporter genes. Plant Biol. 2021, 23, 113–121. [Google Scholar] [CrossRef]
- Ellouzi, H.; Zorrig, W.; Amraoui, S.; Oueslati, S.; Abdelly, C.; Rabhi, M.; Siddique, K.H.M.; Hessini, K. Seed Priming with Salicylic Acid Alleviates Salt Stress Toxicity in Barley by Suppressing ROS Accumulation and Improving Antioxidant Defense Systems, Compared to Halo- and Gibberellin Priming. Antioxidants 2023, 12, 1779. [Google Scholar] [CrossRef]
- Choi, J.-Y.; Ju, Y.-H.; Nakamichi, A.; Cho, S.-W.; Woo, S.-H.; Sakagami, J.-I. Effect of Seed Hydropriming on the Elongation of Plumule and Radicle During the Germination Process and Changes in Enzyme Activity Under Water-Deficient Conditions. Plants 2024, 13, 3537. [Google Scholar] [CrossRef]
- Ren, M.; Tan, B.; Xu, J.; Yang, Z.; Zheng, H.; Tang, Q.; Wang, W. Priming methods affected deterioration speed of primed rice seeds by regulating reactive oxygen species accumulation, seed respiration and starch degradation. Front. Plant Sci. 2023, 14, 1267103. [Google Scholar] [CrossRef]
- Ali, L.G.; Nulit, R.; Ibrahim, M.H.; Yien, C.Y.S. Efficacy of KNO3, SiO2 and SA Priming for Improving Emergence, Seedling Growth and Antioxidant Enzymes of Rice (Oryza sativa), under Drought. Sci. Rep. 2021, 11, 3864. [Google Scholar] [CrossRef]
- Tounekti, T.; Mahdhi, M.; Al-Faifi, Z.; Khemira, H. Priming Improves Germination and Seed Reserve Utilization, Growth, Antioxidant Responses and Membrane Stability at Early Seedling Stage of Saudi Sorghum Varieties under Drought Stress. Not. Bot. Horti Agrobot. Cluj. Napoca 2020, 48, 938–953. [Google Scholar] [CrossRef]
- Mim, M.F.; Chowdhury, M.Z.H.; Rohman, M.M.; Naz, A.; Bhuiyan, A.U.A.; Mohi-Ud-Din, M.; Haque, M.A.; Islam, S.M.N. Metarhizium Anisopliae (MetA1) Seed Priming Improves Photosynthesis, Growth, Plant Defense and Yield of Wheat under Drought Stress. Plant Physiol. Biochem. 2024, 217, 109239. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan, M.; Ali, Q.; Ali, S. Seed Priming with Silicon Nanoparticles Improved the Biomass and Yield While Reduced the Oxidative Stress and Cadmium Concentration in Wheat Grains. Environ. Sci. Pollut. Res. 2019, 26, 7579–7588. [Google Scholar] [CrossRef]
- Nawaz, F.; Naeem, M.; Akram, A.; Ashraf, M.Y.; Ahmad, K.S.; Zulfiqar, B.; Sardar, H.; Shabbir, R.N.; Majeed, S.; Shehzad, M.A.; et al. Seed Priming with KNO3 Mediates Biochemical Processes to Inhibit Lead Toxicity in Maize (Zea mays L.). J. Sci. Food Agric. 2017, 97, 4780–4789. [Google Scholar] [CrossRef]
- Al-Hazmi, N.E.; Naguib, D.M. Amylase Properties and Its Metal Tolerance during Rice Germination Improved by Priming with Rhizobacteria. Rhizosphere 2022, 22, 100518. [Google Scholar] [CrossRef]
- Dabral, S.; Yashaswee; Varma, A.; Choudhary, D.K.; Bahuguna, R.N.; Nath, M. Biopriming with Piriformospora Indica Ameliorates Cadmium Stress in Rice by Lowering Oxidative Stress and Cell Death in Root Cells. Ecotoxicol. Environ. Saf. 2019, 186, 109741. [Google Scholar] [CrossRef]
- Basit, F.; Bhat, J.A.; Ulhassan, Z.; Noman, M.; Zhao, B.; Zhou, W.; Kaushik, P.; Ahmad, A.; Ahmad, P.; Guan, Y. Seed Priming with Spermine Mitigates Chromium Stress in Rice by Modifying the Ion Homeostasis, Cellular Ultrastructure and Phytohormones Balance. Antioxidants 2022, 11, 1704. [Google Scholar] [CrossRef] [PubMed]
- Al-Quraan, N.A.; Samarah, N.H.; Rasheed, E.I. The Role of Chitosan Priming in Induction of GABA Shunt Pathway during Wheat Seed Germination under Salt Stress. Biol. Plant 2023, 67, 234–248. [Google Scholar] [CrossRef]
- Islam, A.T.M.T.; Ullah, H.; Himanshu, S.K.; Tisarum, R.; Cha-um, S.; Datta, A. Effect of Salicylic Acid Seed Priming on Morpho-Physiological Responses and Yield of Baby Corn under Salt Stress. Sci. Hortic. 2022, 304, 111304. [Google Scholar] [CrossRef]
- Hadia, E.; Slama, A.; Romdhane, L.; Cheikh M’Hamed, H.; Fahej, M.A.S.; Radhouane, L. Seed Priming of Bread Wheat Varieties with Growth Regulators and Nutrients Improves Salt Stress Tolerance Particularly for the Local Genotype. J. Plant Growth Regul. 2023, 42, 304–318. [Google Scholar] [CrossRef]
- Mamedi, A.; Sharifzadeh, F.; Maali-Amiri, R.; Divargar, F.; Rasoulnia, A. Seed Osmopriming with Ca2+ and K+ Improves Salt Tolerance in Quinoa Seeds and Seedlings by Amplifying Antioxidant Defense and Ameliorating the Osmotic Adjustment Process. Physiol. Mol. Biol. Plants 2022, 28, 251–274. [Google Scholar] [CrossRef]
- Ahmad, A.; Tola, E.K.; Alshahrani, T.S.; Seleiman, M.F. Enhancement of Morphological and Physiological Performance of Zea mays L. under Saline Stress Using ZnO Nanoparticles and 24-Epibrassinolide Seed Priming. Agronomy 2023, 13, 771. [Google Scholar] [CrossRef]
- Chattha, M.U.; Hassan, M.U.U.; Khan, I.; Nawaz, M.; Shah, A.N.; Sattar, A.; Hashem, M.; Alamri, S.; Aslam, M.T.; Alhaithloul, H.A.S.; et al. Hydrogen Peroxide Priming Alleviates Salinity Induced Toxic Effect in Maize by Improving Antioxidant Defense System, Ionic Homeostasis, Photosynthetic Efficiency and Hormonal Crosstalk. Mol. Biol. Rep. 2022, 49, 5611–5624. [Google Scholar] [CrossRef] [PubMed]
- Kasana, R.A.; Iqbal, M.; Ali, Q.; Saeed, F.; Rizwan, M.; Perveen, R.; Yong, J.W.H. Synergistic Effects of Glutathione and Zinc Seed Priming in Alleviating Salt Stress on Maize Seed Germination, Metabolite Levels, Seedling Vigor, and Nutrient Acquisition. Plant Stress. 2025, 15, 100767. [Google Scholar] [CrossRef]
- Dutta, B.; Datta, A.; Dey, A.; Ghosh, A.K.; Bandopadhyay, R. Establishment of Seed Biopriming in Salt Stress Mitigation of Rice Plants by Mangrove Derived Bacillus Sp. Biocatal. Agric. Biotechnol. 2023, 48, 102626. [Google Scholar] [CrossRef]
- Cembrowska-Lech, D.; Rybak, K. Nanopriming of Barley Seeds—A Shotgun Approach to Improve Germination under Salt Stress Conditions by Regulating of Reactive Oxygen Species. Plants 2023, 12, 405. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Aslam, M.U.; Valipour, M.; Iqbal, R.; Haider, I.; Mustafa, A.E.Z.M.A.; Elshikh, M.S.; Ali, I.; Roy, R.; Elshamly, A.M.S. Seed Priming with Selenium Improves Growth and Yield of Quinoa Plants Suffering Drought. Sci. Rep. 2024, 14, 886. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.; Madhu, G.S.; Singh, V.P. Seed Biopriming with Trichoderma harzianum for Growth Promotion and Drought Tolerance in Rice (Oryza sativus). Agric. Res. 2023, 12, 154–162. [Google Scholar] [CrossRef]
- Lastochkina, O.; Garshina, D.; Ivanov, S.; Yuldashev, R.; Khafizova, R.; Allagulova, C.; Fedorova, K.; Avalbaev, A.; Maslennikova, D.; Bosacchi, M. Seed Priming with Endophytic Bacillus subtilis Modulates Physiological Responses of Two Different Triticum aestivum L. Cultivars Under Drought Stress. Plants 2020, 9, 1810. [Google Scholar] [CrossRef]
- Rizk, R.; Ahmed, M.; Abdul-Hamid, D.; Zedan, M.; Tóth, Z.; Decsi, K. Resulting Key Physiological Changes in Triticum aestivum L. Plants Under Drought Conditions After Priming the Seeds with Conventional Fertilizer and Greenly Synthesized Zinc Oxide Nanoparticles from Corn Wastes. Agronomy 2025, 15, 211. [Google Scholar] [CrossRef]
- Al Ghafri, S.H.; Al-Busaidi, W.M.; Farooq, M. Enhancing Drought Tolerance in Bread Wheat Through GABA Seed Priming and Optimized Storage Temperatures. J. Plant Growth Regul. 2025. [Google Scholar] [CrossRef]
- El-Sanatawy, A.M.; Ash-Shormillesy, S.M.A.I.; Qabil, N.; Awad, M.F.; Mansour, E. Seed Halo-Priming Improves Seedling Vigor, Grain Yield, and Water Use Efficiency of Maize under Varying Irrigation Regimes. Water 2021, 13, 2115. [Google Scholar] [CrossRef]
- Yang, H.; Hu, W.; Zhao, J.; Huang, X.; Zheng, T.; Fan, G. Genetic Improvement Combined with Seed Ethephon Priming Improved Grain Yield and Drought Resistance of Wheat Exposed to Soil Water Deficit at Tillering Stage. Plant Growth Regul. 2021, 95, 399–419. [Google Scholar] [CrossRef]
- Nadali, F.; Asghari, H.R.; Abbasdokht, H.; Dorostkar, V.; Bagheri, M. Improved Quinoa Growth, Physiological Response, and Yield by Hydropriming Under Drought Stress Conditions. Gesunde Pflanz. 2021, 73, 53–66. [Google Scholar] [CrossRef]
- Thongbam, S.; Sinam, V.; Mentada, B.A.S.; Kalangutkar, A.M.; Siddique, A. Priming-Mediated Triggering of Antioxidative Response to Induce Drought Tolerance in Maize (Zea mays L.). Plant Sci. Today 2023, 10, 247–252. [Google Scholar] [CrossRef]
- Van Nguyen, D.; Nguyen, H.M.; Le, N.T.; Nguyen, K.H.; Nguyen, H.T.; Le, H.M.; Nguyen, A.T.; Dinh, N.T.T.; Hoang, S.A.; Van Ha, C. Copper Nanoparticle Application Enhances Plant Growth and Grain Yield in Maize Under Drought Stress Conditions. J. Plant Growth Regul. 2022, 41, 364–375. [Google Scholar] [CrossRef]
- Elkelish, A.; Alqudah, A.M.; Alammari, B.S.; Alsubeie, M.S.; Hamed, S.M.; Thabet, S.G. Exploring genetic determinants of silver oxide nanoparticle-induced seed priming for drought tolerance in wheat. Genet. Resour. Crop Evol. 2025, 72, 3203–3218. [Google Scholar] [CrossRef]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed Priming with Zinc Oxide Nanoparticles Downplayed Ultrastructural Damage and Improved Photosynthetic Apparatus in Maize under Cobalt Stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, X.; Yang, W.; Xie, H.; Ashraf, U.; Mo, Z.; Liu, J.; Li, G.; Li, W. Seed Priming with Multiwall Carbon Nanotubes (MWCNTs) Modulates Seed Germination and Early Growth of Maize Under Cadmium (Cd) Toxicity. J. Soil. Sci. Plant Nutr. 2021, 21, 1793–1805. [Google Scholar] [CrossRef]
- Bashir, S.; Javed, S.; Al-Anazi, K.M.; Farah, M.A.; Ali, S. Bioremediation of Cadmium Toxicity in Wheat (Triticum aestivum L.) Plants Primed with L-Proline, Bacillus subtilis and Aspergillus niger. Int. J. Environ. Res. Public Health 2022, 19, 12683. [Google Scholar] [CrossRef]
- Ma, J.; Zou, M.; Peijnenburg, W.; Chen, F. Priming Agents Combat Copper Stress in Wheat (Triticum aestivum L.) under Hydroponic Conditions: Insights in Impacts on Morpho–Physio–Biochemical Traits and Health Risk Assessment. Ecotoxicol. Environ. Saf. 2025, 291, 117899. [Google Scholar] [CrossRef]
- Mu, C.; Huang, D.; Wang, M.; Li, Y.; Wang, X.; Si, D.; Cheng, C.; Ge, C.; Zhao, L.; Zhou, D. Seed Priming with Silver Ions Improves Growth and Physicochemical Features of Rice Plants (Oryza sativa L.) under Copper Stress. ACS Agric. Sci. Technol. 2024, 4, 711–722. [Google Scholar] [CrossRef]
- Choudhury, S.; Moulick, D.; Mazumder, M.K.; Pattnaik, B.K.; Ghosh, D.; Vemireddy, L.R.; Aldhahrani, A.; Soliman, M.M.; Gaber, A.; Hossain, A. An In Vitro and In Silico Perspective Study of Seed Priming with Zinc on the Phytotoxicity and Accumulation Pattern of Arsenic in Rice Seedlings. Antioxidants 2022, 11, 1500. [Google Scholar] [CrossRef] [PubMed]
- Basit, F.; Liu, J.; An, J.; Chen, M.; He, C.; Zhu, X.; Li, Z.; Hu, J.; Guan, Y. Seed Priming with Brassinosteroids Alleviates Aluminum Toxicity in Rice via Improving Antioxidant Defense System and Suppressing Aluminum Uptake. Environ. Sci. Pollut. Res. 2022, 29, 10183–10197. [Google Scholar] [CrossRef] [PubMed]
- Sobhy, S.E.; Abo-Kassem, E.E.M.; Sewelam, N.A.; Saad-Allah, K.M.; Aseel, D.G.; Saleh, A.A.; Hafez, E.E. Growth, Physiological and Molecular Response of Calcium and Salicylic Acid Primed Wheat under Lead Stress. Mol. Biol. Rep. 2025, 52, 133. [Google Scholar] [CrossRef] [PubMed]
- AL-Huqail, A.A.; Alatawi, A.; Alghanem, S.M.S.; Khan, K.A.; Abeed, A.H.A. Innovative Approach Using Different Nano—Primers to Enhance Stress Tolerance in Barley (Hordeum vulgare L.) Under Lead Toxicity. J. Soil. Sci. Plant Nutr. 2025. [Google Scholar] [CrossRef]
- Li, Z.; Xu, J.; Gao, Y.; Wang, C.; Guo, G.; Luo, Y.; Huang, Y.; Hu, W.; Sheteiwy, M.S.; Guan, Y.; et al. The Synergistic Priming Effect of Exogenous Salicylic Acid and H2O2 on Chilling Tolerance Enhancement during Maize (Zea mays L.) Seed Germination. Front. Plant Sci. 2017, 8, 1153. [Google Scholar] [CrossRef]
- Salah, S.M.; Yajing, G.; Dongdong, C.; Jie, L.; Aamir, N.; Qijuan, H.; Weimin, H.; Mingyu, N.; Jin, H. Seed Priming with Polyethylene Glycol Regulating the Physiological and Molecular Mechanism in Rice (Oryza sativa L.) under Nano-ZnO Stress. Sci. Rep. 2015, 5, 14278. [Google Scholar] [CrossRef]
- Paul, S.; Roychoudhury, A. Seed Priming with Spermine and Spermidine Regulates the Expression of Diverse Groups of Abiotic Stress-Responsive Genes during Salinity Stress in the Seedlings of Indica Rice Varieties. Plant Gene 2017, 11, 124–132. [Google Scholar] [CrossRef]
- Alzahrani, O.; Abouseadaa, H.; Abdelmoneim, T.K.; Alshehri, M.A.; El-Mogy, M.M.; El-Beltagi, H.S.; Atia, M.A.M. Agronomical, Physiological and Molecular Evaluation Reveals Superior Salt-Tolerance in Bread Wheat through Salt-Induced Priming Approach. Not. Bot. Horti Agrobot. Cluj. Napoca 2021, 49, 1–21. [Google Scholar] [CrossRef]
- Hidayah, A.; Nisak, R.R.; Susanto, F.A.; Nuringtyas, T.R.; Yamaguchi, N.; Purwestri, Y.A. Seed Halopriming Improves Salinity Tolerance of Some Rice Cultivars During Seedling Stage. Bot. Stud. 2022, 63, 24. [Google Scholar] [CrossRef]
- Forti, C.; Shankar, A.; Singh, A.; Balestrazzi, A.; Prasad, V.; Macovei, A. Hydropriming and Biopriming Improve Medicago truncatula Seed Germination and Upregulate DNA Repair and Antioxidant Genes. Genes 2020, 11, 242. [Google Scholar] [CrossRef]
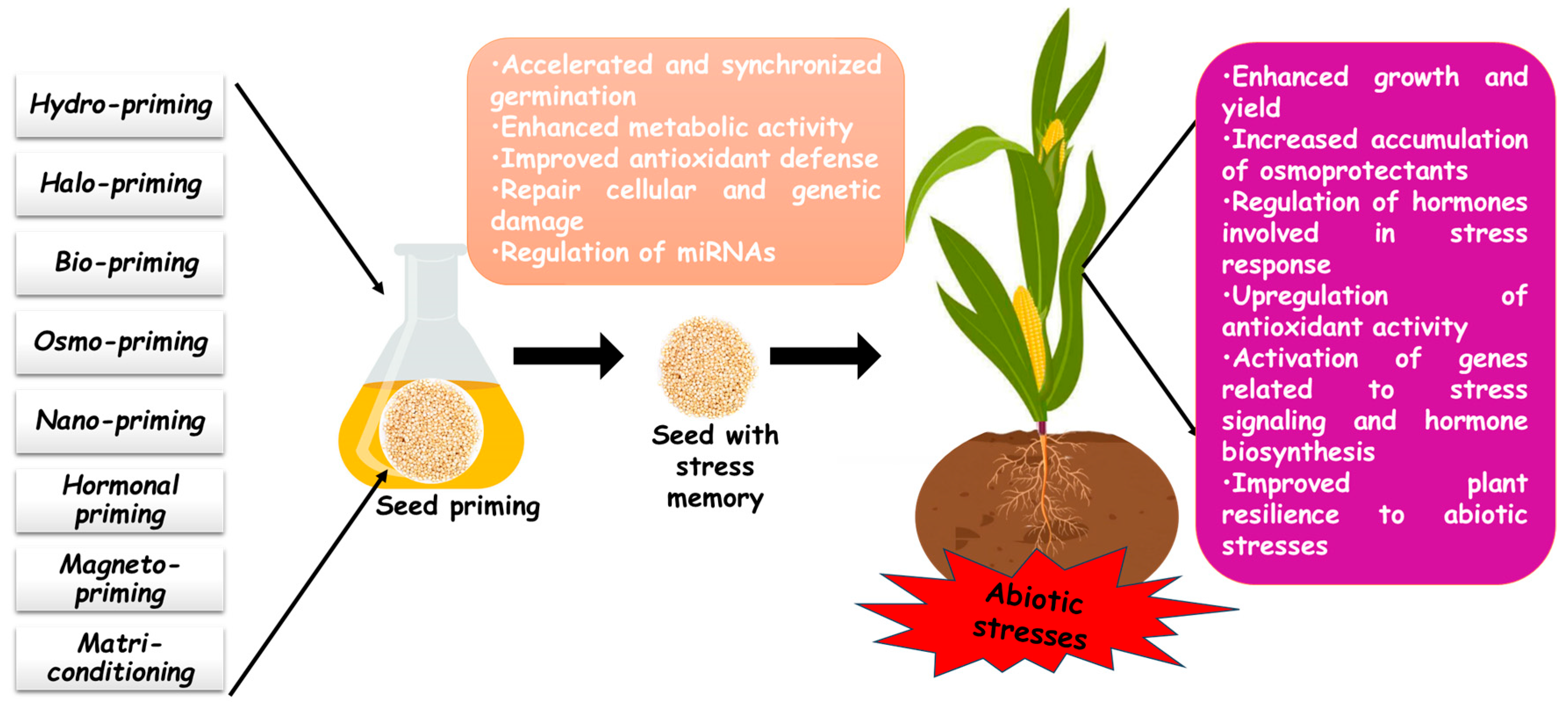
| Cereal Species | Abiotic Stress | Main Effects | Ref. |
|---|---|---|---|
| Triticum aestivum L. | Salt stress | -Negatively affected growth, yield, and physiological and biochemical traits (e.g., Decreased Ca2+ and K+ levels, increased Na+, Cl−, H2O2, MDA, and membrane permeability). -Increased antioxidant enzyme activities (SOD, CAT, APX, POD). | [56] |
| Drought stress | -Reduced morpho-physiological and biochemical traits (e.g., chlorophyll content, RWC, membrane stability, NPK levels). -Increased yield-related traits (tillers/plant, spikelets/spike, grains/spike, seed and biological yield) were negatively affected. | [57] | |
| Heavy metals toxicity | -Reduced seedling length, biomass, and root and shoot growth. -Higher accumulation of metals in roots than shoots. -Structural damage observed in root epidermis, cortical cells, and xylem vessels. -Reductions in stomatal size/number, long and short cells, and trichomes. | [58] | |
| Zea mays L. | Salt stress | -Reduced seed germination, shoot/root growth, and biomass. -Reduced physiological traits (chlorophyll, rwc, photosynthesis, transpiration). -High accumulation of Na+ and Cl− and reduced K+ uptake. | [59] |
| Drought stress | -Significant alterations in stomatal conductance, transpiration rate, photosynthesis, antioxidant enzymes, and proline levels. | [60] | |
| Heavy metals toxicity | -Reduction in morphological parameters: chlorophyll a, b, and total content. -Accumulation of secondary metabolites such as proline and flavonoids. | [61] | |
| Hordeum vulgare L. | Salt stress | -Reduced relative water content, leaf water potential, and growth parameters. -Increased proline levels in both leaves and roots, ABA, and ethylene levels. -Reduction in jasmonic acid levels. -Increased H2O2 and lipid peroxidation. | [62] |
| Drought stress | -Reduced relative water content, shoot dry weight, and chlorophyll content (SPAD index). -Increased oxidative damage indicators: hydrogen peroxide and malondialdehyde. -Increased electrolyte leakage (membrane damage). -Elevated osmoprotectants: water-soluble carbohydrates and soluble proteins. -Enhanced antioxidant enzyme activities: catalase and ascorbate peroxidase. | [63] | |
| Heavy metals toxicity | -Growth inhibition and oxidative damage (increased H2O2 and lipid peroxidation). | [64] | |
| Oryza sativa L. | Salt stress | -Reduced root growth, tissue death, and stunted development. -Chlorosis, leaf curling, leaf scorching. -Spikelet sterility, flowering abortion, embryo senescence, and yield reduction. -Stomatal closure, inhibited photosynthesis, impaired enzymatic activity, and protein synthesis. -Altered cell metabolism and increased oxidative stress. | [65] |
| Drought stress | -Stomatal closure, constrained cell division and elongation, reduced photosynthesis, lower turgor pressure, and yield loss. -Reduced seed germination, fewer tillers, early flowering, and reduced biomass. -Increased accumulation of reactive oxygen species, stress-related metabolites, ABA, and antioxidative enzyme activity. | [66] | |
| Heavy metals toxicity | -Reduced plant height, biomass, SPAD index, PSII efficiency, and photosynthetic performance. -Increased Cd and Fe in roots and shoots. -Upregulation of Cd and Fe transporters (e.g., OSHMA2, OSHMA3, OSNRAMP1, OSNRAMP5, OSIRT1, OSFRO1). -Connections with proteins involved in Fe homeostasis (e.g., MTP1, YSL6, IRO2, NAS2). | [67] | |
| Sorghum bicolor L. | Salt stress | -Oxidative stress markers and membrane damage increased with salt stress. -Chlorophyll content and PSII performance decreased due to salinity. | [68] |
| Drought stress | -Reduction in plant height, leaf water content, and chlorophyll content. -Increase in proline, malondialdehyde, soluble sugar, electrolyte leakage, hydrogen peroxide, and antioxidant enzyme activity. -Upregulation of drought-responsive genes related to antioxidants, osmolytes biosynthesis, dehydrins, photosystem, and transcription. | [69] | |
| Heavy metals toxicity | -Growth inhibition under increasing Cd concentration. -Oxidative stress: increased MDA and H2O2 levels. -Heme Oxygenase-1 (HO-1) activity peaked at 150 µm CdCl2. -Enhanced antioxidant defense (APX, GPX, CAT). -High HO-1 activity correlated with ROS scavenging. | [70] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janah, I.; Elhasnaoui, A.; Laouane, R.B.; Ait-El-Mokhtar, M.; Anli, M. Exploring Seed Priming as a Strategy for Enhancing Abiotic Stress Tolerance in Cereal Crops. Stresses 2025, 5, 39. https://doi.org/10.3390/stresses5020039
Janah I, Elhasnaoui A, Laouane RB, Ait-El-Mokhtar M, Anli M. Exploring Seed Priming as a Strategy for Enhancing Abiotic Stress Tolerance in Cereal Crops. Stresses. 2025; 5(2):39. https://doi.org/10.3390/stresses5020039
Chicago/Turabian StyleJanah, Iman, Abdelhadi Elhasnaoui, Raja Ben Laouane, Mohamed Ait-El-Mokhtar, and Mohamed Anli. 2025. "Exploring Seed Priming as a Strategy for Enhancing Abiotic Stress Tolerance in Cereal Crops" Stresses 5, no. 2: 39. https://doi.org/10.3390/stresses5020039
APA StyleJanah, I., Elhasnaoui, A., Laouane, R. B., Ait-El-Mokhtar, M., & Anli, M. (2025). Exploring Seed Priming as a Strategy for Enhancing Abiotic Stress Tolerance in Cereal Crops. Stresses, 5(2), 39. https://doi.org/10.3390/stresses5020039