
Bioactive Compounds from Marine Macroalgae: A Natural Defense Against Oxidative Stress-Related Diseases

 , , , and
, , , and 
Abstract
1. Introduction
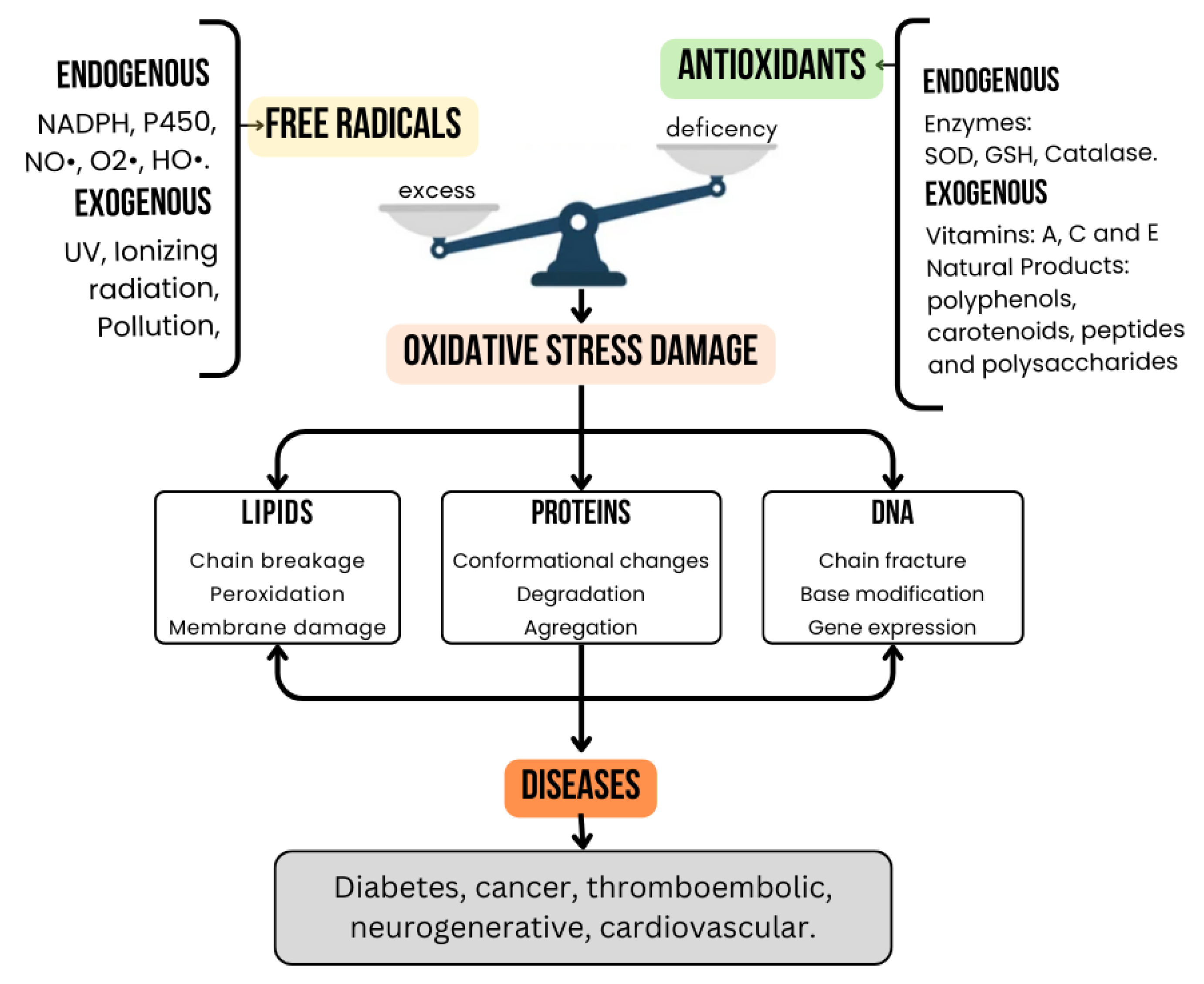
2. Oxidative Stress and Its Role in Chronic Diseases
2.1. Mechanisms of Oxidative Stress in the Body
2.2. Link Between Oxidative Stress and Chronic Diseases
3. Marine Algae as a Source of Bioactive Compounds
4. Mechanisms of Action of Algal Compounds Against Oxidative Stress
4.1. Phenolic Compounds
4.2. Carotenoids
4.3. Polysaccharides
4.4. Peptides
5. Therapeutic Potential of Marine Algal Bioactive Compounds
5.1. Diabetes

{kind=link}
| Therapeutic Area | Algae | Bioactive Compound | Observed Effects | References |
|---|---|---|---|---|
| Diabetes | Polycladia myrica, Halimeda opuntia | Carboxylic acids | Reduction in blood glucose levels, improvement in kidney and liver function, and increase in antioxidants, such as reduced glutathione and superoxide dismutase (SOD). | [86] |
| Caulerpa racemosa | - | Lower glucose levels, weight control, and reduced hepatic fat accumulation. | [87] | |
| Laurencia papillosa | Ethanolic compounds | Reduced preprandial and postprandial glucose levels, interaction with key enzymes, such as pancreatic α-amylase and the insulin receptor. | [88] | |
| Polycladia myrica, Padina antillarum | Glycosides | High α-glucosidase inhibition (IC50: ~12–13 µg/mL). | [89] | |
| Spirulina platensis | acacetin pinocembrin ω-6 PUFA | Decrease in glucose levels, HbA1c percentage as well as TNF-α and IL-6 levels. | [90,91] | |
| Ecklonia cava | Dieckol Fucodiphloroethol G Phlorofucofuroeckol A 6,6′-bieckol 7-Phloroeckol | Inhibition of α-glucosidase and α-amylase. Alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. | [92] | |
| Sargassum hemiphyllum | Fucoxanthin | Inhibits α-amylase, sucrase, maltase absorption, enhances insulin release in vitro. | [93] | |
| Ulva pinnatifida | Fucoxanthin | Significant decrease on blood glucose levels promotes translocation and induction of glucose transporter 4 in skeletal muscles of diabetic/obese KK-Ay mice. | [94] | |
| Caulerpa lentillifera | Polyphenols and sterols | Decrease dipeptidyl peptidase-IV and α-glucosidase enzyme. Increases insulin secretion and glucose uptake in 3T3-L1 adipocytes (10 μL CLE). | [95] | |
| Halimeda tuna | Flavonoids and phenol hydroquinine | Inhibits α-amylase IC50 = 0.88 mg/mL and α-glucosidase IC50 = 0.01 mg/mL. | [96] | |
| Cancer | Laminaria japonica | Fucoxanthin | Inhibition of cell migration and enhanced therapeutic efficacy when combined with gefitinib in lung cancer treatment. | [97] |
| Ulva rigida, Chaetomorpha myrica, Gracilaria foliifera | Terpenoids, polyphenols, sulfonates, polysaccharides, fatty acids, chlorophylls, amide proteins, flavonoids, carotenoids | Significant inhibition of human cancer cells (MCF-7), with IC50 values between 13 and 43 µg/mL. | [98] | |
| Lyngbya majuscula | Cucarin-A | Interferes with tubulin polymerization by competitively binding to the colchicine-binding site. | [99] | |
| Dunaliella salina | - | Anti-proliferative activity against the SW480 colon carcinoma cell line. | [100] | |
| Amphiroa anceps | Octadecanoic acid and n-Hexadecanoic acid | A 92% reduction in viability of A549 cancer cells, with no teratogenic effects in zebrafish models. | [101] | |
| Codium decorticatum | Fucoidan | Induction of apoptosis of A549 cancer cells. | [102] | |
| Ulva lactuca Ulva fasciata | Di-isooctyl Phthalate Butylated Hydroxytoluene | U. lactuca extract had strong activity against MCF-7 and Hela cell lines (IC50 10.83 ± 1.0, 12.43 ± 1.3 μg/mL, respectively), while U. fasciata had strong activity against PC3 and HePG2 cell lines (IC50 12.99 ± 1.2, 16.75 ± 1.5 μg/mL. | [103] | |
| Ecklonia cava | Caffeic acid, naringin, catechin hydrate and phloroglucinol | Suppress the growth of CT26 colon cancer by activating apoptosis, suppressing cell proliferation, inhibiting cell migration and enhancing the tumor-suppressing activity. | [104] | |
| Caulerpa racemosa | Caulersin (C2) | Identified as a potent and effective agent in fighting against non-small cell lung cancer (NSCLC). | [105] | |
| Laurencia obtusa | Terpenes and acetogenins | Potential to be used in the treatment of neoplasms, such as gastric adenocarcinoma. | [106] | |
| Costaria costata | Phlorethol CcPh | Inhibitor of the α-NaGalase of cancer cells and, therefore, has high therapeutic potential. | [107] | |
| Thromboembolic Diseases | Padina tetrastromatica, Ulva fasciata | Sulfated polysaccharides PSPS; SPS from P. tetrastromatica, USPS; SPS from U. fasciata | Significant prolongation of coagulation times (APTT and PT), comparable or superior to heparin in vitro and in SD rats. Exert antithrombotic activity through the modulation of the intrinsic coagulation pathway. | [108] |
| Monostroma nitidum | Sulfated polysaccharide MS-1 | Potent anticoagulant and thrombolytic activity by acting on antithrombin-III and factor Xa. | [109] | |
| Rhodomela confervoides | Sulfated polysaccharides | Anticoagulant activity, especially in ethanol extract fraction (prolonging clotting time to 407.97 s at 10 mg/mL. | [110] | |
| Cystoseira humilis and Sargassum vulgare | Sulfated polysaccharides | Procoagulant effects in certain fractions. | [110] | |
| Chlorella sorokiniana | Sulfated polysacharides | Prolonged clotting time more than 38 s at 10 µg/mL in a PTT test. The PT at 200 µg/mL increased clotting time (up to 14 s). | [111] | |
| Caulerpa cupressoide | Sulfated polysaccharides SP1, SP2 y SP3 | Anticoagulant. Potentiation of thrombin inhibition by antithrombin (IC50 = 10 µg/mL). | [112] | |
| Enteromorpha clathrata | Sulfated polysaccharide high arabinose containing | Prolonged the activated partial thromboplastin time and thrombin time in vitro. | [113] | |
| Monostroma angicava | Sulfated polysaccharide PF2 | Anticoagulant activity was mainly attributed to strong potentiation thrombin or factor Xa inhibition. | [114] | |
| Botryocladia occidentalis | Sulfated galactan | Enhance thrombin and factor Xa, inhibition by antihrombin and/or heparin cofactor II. | [115] | |
| Codium divaricatum | Sulfated polysaccharide CP2-1 | High anticoagulant activity assessed by activated partial thromboplastin time and thrombin time. | [116] | |
| Neurodegenerative Diseases | Microalgae | Astaxanthin | AST plays a protective role in neurons and enhances learning, memory, and cognitive abilities. | [117] |
| Sargassum fusiforme | Saringosterol | Improved memory and 81% reduction in beta-amyloid plaques in the hippocampus, with no adverse effects. | [118] | |
| - | Sodium oligomannate GV-971, seaweed-derived | Significant cognitive improvement in patients with mild to moderate Alzheimer’s; approved in China. | [119,120] | |
| Nannochloropsis oceanica | DHA, EPA | Effects against amyloid-beta (Aβ)-induced toxicity in neuronal cells. mitigated Aβ-mediated oxidative stress and upregulated the activity of key antioxidant enzymes, specifically superoxide dismutase SOD, GSH levels, and CAT, in Neuro-2A neuroblastoma cells. | [121] | |
| Sargussum horneri | Fucoxanthin | Anti-Alzheimer’s disease (AD) neuroprotective effects in vitro and in vivo. | [122] | |
| Sargassum macrocarpum | Sargachromenol | Promotes neuronal differentiation of PC12D cells and supports the survival of neuronal PC12D cells via two distinct signaling pathways. | [123] | |
| Laminaria japonica | Fucoidan | Protects against dopaminergic neuron death in vivo and in vitro, via its antioxidative activity. | [124] | |
| Marine red algae | κ-carrageenan oligosaccharides (KOS) | Has immunomodulatory effects and can be used as a potential intervention therapy for inflammatory related neurodegenerative diseases. | [125] |
5.2. Cancer
5.3. Thromboembolic Disease
5.4. Neurodegenerative Diseases
6. Novel Extraction Techniques of Algal Bioactive Compounds
6.1. Assisted Extraction Methods
6.1.1. Enzyme-Assisted Extraction (EAE)
6.1.2. Microwave-Assisted Extraction (MAE)
6.1.3. Ultrasound-Assisted Extraction (UAE)
6.1.4. Irradiation-Assisted Extraction (IAE)
6.2. Emerging Extraction Techniques
6.2.1. Supercritical Fluid Extraction (SFE)
6.2.2. Pressurized Liquid Extraction (PLE)
6.2.3. Deep Eutectic Solvents (DES)
6.2.4. Pulsed Electric Field (PEF)
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine Bacterial Secondary Metabolites: A Treasure House for Structurally Unique and Effective Antimicrobial Compounds. Mar. Drugs 2021, 19, 530. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. Algae as a source of bioactive ingredients for the formulation of functional foods and nutraceuticals. In Functional Ingredients from Algae for Foods and Nutraceuticals, 2nd ed.; Dominguez, H., Pereira, L., Kraan, S., Eds.; Woodhead Publishing: Sawston, UK, 2023; pp. 3–114. [Google Scholar] [CrossRef]
- Ganesan, A.R.; Tiwari, U.; Rajauria, G. Seaweed nutraceuticals and their therapeutic role in disease prevention. Food Sci. Hum. Wellness 2019, 8, 252–263. [Google Scholar] [CrossRef]
- Begum, R.; Howlader, S.; Mamun-Or-Rashid, A.N.M.; Rafiquzzaman, S.M.; Ashraf, G.M.; Albadrani, G.M.; Uddin, M.S. Antioxidant and Signal-Modulating Effects of Brown Seaweed-Derived Compounds Against Oxidative Stress-Associated Pathology. Oxid. Med. Cell. Longev. 2021, 1, 9974890. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cel. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil-Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Bhadange, Y.A.; Carpenter, J.; Saharan, V.K. A Comprehensive Review on Advanced Extraction Techniques for Retrieving Bioactive Components From Natural Sources. ACS Omega 2024, 9, 31274–31297. [Google Scholar] [CrossRef]
- Carvajal, C. Especies Reactivas Del Oxígeno: Formación, Función Y Estrés Oxidativo. Med. Leg. De Costa Rica 2019, 36, 91–100. [Google Scholar]
- Yoshikawa, T.; You, F. Oxidative Stress and Bio-Regulation. Int. J. Mol. Sci. 2024, 25, 3360. [Google Scholar] [CrossRef]
- Perrone, S.; Lembo, C.; Giordano, M.; Petrolini, C.; Cannavò, L.; Gitto, E. Molecular Mechanisms of Oxidative Stress-Related Neonatal Jaundice. J. Biochem. Mol. Toxicol. 2023, 37, e23349. [Google Scholar] [CrossRef]
- Adwas, A.A.; Elsayed, A.; Azab, A.E.; Quwaydir, F.A. Oxidative Stress And Antioxidant Mechanisms In Human Body. J. Appl. Biotechnol. Bioeng. 2019, 6, 43–47. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defense Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role In The Entire Antioxidant Defense Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Obeme-Nmom, J.I.; Abioye, R.O.; Flores, S.S.R.; Udenigwe, C.C. Regulation Of Redox Enzymes By Nutraceuticals: A Review Of The Roles Of Antioxidant Polyphenols And Peptides. Food Funct. 2024, 11, 10956–10980. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive Oxygen Species, Toxicity, Oxidative Stress, and Antioxidants: Chronic Diseases And Aging. Arch. Toxicol. 2023, 97, 2499–2574. [Google Scholar] [CrossRef]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several Lines of Antioxidant Defense Against Oxidative Stress: Antioxidant Enzymes, Nanomaterials With Multiple Enzyme-Mimicking Activities, and Low-Molecular-Weight Antioxidants. Arch. Toxicol. 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The Role of Oxidative Stress in Physiopathology and Pharmacological Treatment with Pro- and Antioxidant Properties in Chronic Diseases. Oxid. Med. Cell Longev. 2020, 23, 2082145. [Google Scholar] [CrossRef]
- Yang, J.; Luo, J.; Tian, X.; Zhao, Y.; Li, Y.; Wu, X. Progress in Understanding Oxidative Stress, Aging, and Aging-Related Diseases. Antioxidants 2024, 25, 394. [Google Scholar] [CrossRef]
- Solovev, I.; Sergeeva, A.; Geraskina, A.; Shaposhnikov, M.; Vedunova, M.; Borysova, O.; Moskalev, A. Aging and physiological barriers: Mechanisms of barrier integrity changes and implications for age-related diseases. Mol. Biol. Rep. 2024, 51, 917. [Google Scholar] [CrossRef]
- Chaudhary, M.R.; Chaudhary, S.; Sharma, Y.; Singh, T.A.; Mishra, A.K.; Sharma, S.; Mehdi, M.M. Aging, oxidative stress and degenerative diseases: Mechanisms, complications and emerging therapeutic strategies. Biogerontology 2023, 24, 609–662. [Google Scholar] [CrossRef]
- Sarmiento-Salinas, F.L.; Perez-Gonzalez, A.; Acosta-Casique, A.; Ix-Ballote, A.; Diaz, A.; Treviño, S.; Rosas-Murrieta, N.H.; Millán-Perez-Peña, L.; Maycotte, P. Reactive Oxygen Species: Role in Carcinogenesis, Cancer Cell Signaling and Tumor Progression. Life Sci. 2021, 284, 119942. [Google Scholar] [CrossRef]
- Nakai, K.; Tsuruta, D. What Are Reactive Oxygen Species, Free Radicals, and Oxidative Stress in Skin Diseases? Int. J. Mol. Sci. 2021, 22, 10799. [Google Scholar] [CrossRef] [PubMed]
- Hassan, H.; Ansari, F.A.; Ingle, K.N.; Singh, K.; Bux, F. Commercial Products and Environmental Benefits of Algal Diversity. In Biodiversity and Bioeconomy; Singh, K., Ribeiro, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2024; pp. 475–502. [Google Scholar] [CrossRef]
- Ahmed, N.; Sheikh, M.A.; Ubaid, M.; Chauhan, P.; Kumar, K.; Choudhary, S. Comprehensive Exploration of Marine Algae Diversity, Bioactive Compounds, Health Benefits, Regulatory Issues, and Food and Drug Applications. Meas. Food 2024, 14, 100163. [Google Scholar] [CrossRef]
- Ghallab, D.S.; Ibrahim, R.S.; Mohyeldin, M.M.; Shawky, E. Marine Algae: A Treasure Trove of Bioactive Anti-inflammatory Compounds. Mar. Pollut. Bull. 2024, 199, 116023. [Google Scholar] [CrossRef]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef] [PubMed]
- Flores-Contreras, E.A.; Araújo, R.G.; Rodríguez-Aguayo, A.A.; Guzmán-Román, M.; García-Venegas, J.C.; Nájera-Martínez, E.F.; Sosa-Hernández, J.E.; Iqbal, H.M.N.; Melchor-Martínez, E.M.; Parra-Saldivar, R. Polysaccharides from the Sargassum and Brown Algae Genus: Extraction, Purification, and Their Potential Therapeutic Applications. Plants 2023, 12, 2445. [Google Scholar] [CrossRef]
- Patel, A.K.; Albarico, F.P.J.B.; Perumal, P.K.; Vadrale, A.P.; Nian, C.T.; Chau, H.T.B.; Anwar, C.; Wani, H.M.U.D.; Pal, A.; Saini, R.; et al. Algae as an Emerging Source of Bioactive Pigments. Bioresour. Technol. 2022, 351, 126910. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Engelen, A.; Varela, J. Polyunsaturated Fatty Acids of Marine Macroalgae: Potential for Nutritional and Pharmaceutical Applications. Mar. Drugs 2012, 9, 1920–1935. [Google Scholar] [CrossRef]
- Arora, N.; Philippidis, G.P. The Prospects of Algae-Derived Vitamins and Their Precursors for Sustainable Cosmeceuticals. Processes 2023, 11, 587. [Google Scholar] [CrossRef]
- Menaa, F.; Wijesinghe, U.; Thiripuranathar, G.; Althobaiti, N.A.; Albalawi, A.E.; Khan, B.A.; Menaa, B. Marine Algae-Derived Bioactive Compounds: A New Wave of Nanodrugs? Mar. Drugs 2021, 19, 484. [Google Scholar] [CrossRef]
- Khan, S.U.; Lone, A.N.; Khan, M.S.; Virani, S.S.; Blumenthal, R.S.; Nasir, K.; Miller, M.; Michos, E.D.; Ballantyne, C.M.; Boden, W.E.; et al. Effect of Omega-3 fatty Acids on Cardiovascular Outcomes: A systematic Review and Meta-analysis. EClinicalMedicine 2021, 38, 100997. [Google Scholar] [CrossRef]
- Pradhan, B.; Nayak, R.; Patra, S.; Jit, B.P.; Ragusa, A.; Jena, M. Bioactive Metabolites from Marine Algae as Potent Pharmacophores against Oxidative Stress-Associated Human Diseases: A Comprehensive Review. Molecules 2021, 26, 37. [Google Scholar] [CrossRef] [PubMed]
- Dighriri, I.M.; Alsubaie, A.M.; Hakami, F.M.; Hamithi, D.M.; Alshekh, M.M.; Khobrani, F.A.; Dalak, F.E.; Hakami, A.A.; Alsueaadi, E.H.; Alsaawi, L.S.; et al. Effects of Omega-3 Polyunsaturated Fatty Acids on Brain Functions: A Systematic Review. Cureus 2022, 14, e30091. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Yadav, D.; Lee, P.C.; Jin, J.O. Immunomodulatory effects of polysaccharides from marine algae for treating cancer, infectious disease, and inflammation. Phytother. Res. 2022, 36, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Nigam, A.; Mishra, R.; Gupta, S.; Chaudhary, A.A.; Khan, S.-U.-D.; Almuqri, E.A.; Ahmed, Z.H.; Rustagi, S.; Singh, D.P.; et al. Novel Therapeutic Approach for Obesity: Seaweeds as an Alternative Medicine with the Latest Conventional Therapy. Med. Sci. 2024, 12, 55. [Google Scholar] [CrossRef]
- Wan-Loy, C.; Siew-Moi, P. Marine Algae as a Potential Source for Anti-Obesity Agents. Mar. Drugs 2016, 14, 222. [Google Scholar] [CrossRef]
- Jayawardhana, H.H.A.C.K.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Liyanage, N.M.; Nagahawatta, D.P.; Lee, H.-G.; Kim, J.-I.; Jeon, Y.-J. Marine Algal Polyphenols as Skin Protective Agents: Current Status and Future Prospectives. Mar. Drugs 2023, 21, 285. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.-E.; Kim, K.H.; Kang, N.J. Beneficial Effects of Marine Algae-Derived Carbohydrates for Skin Health. Mar. Drugs 2018, 16, 459. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Sosa-Hernández, J.E.; Raza, A.; Nabeel, F.; Iqbal, H.M.N. Biosorption: An Interplay between Marine Algae and Potentially Toxic Elements—A Review. Mar. Drugs 2018, 16, 65. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine Macroalgae as Sources of Protein and Bioactive Compounds in Feed for Monogastric Animals. J Sci Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef]
- Jimenez-Lopez, C.; Pereira, A.G.; Lourenço-Lopes, C.; Garcia-Oliveira, P.; Cassani, L.; Fraga-Corral, M.; Prieto, M.A.; Simal-Gandara, J. Main bioactive phenolic compounds in marine algae and their mechanisms of action supporting potential health benefits. Food Chem. 2021, 30, 128262. [Google Scholar] [CrossRef]
- Lv, Q.Z.; Long, J.T.; Gong, Z.F.; Nong, K.Y.; Liang, X.M.; Qin, T.; Huang, W.; Yang, L. Current State of Knowledge on the Antioxidant Effects and Mechanisms of Action of Polyphenolic Compounds. Nat. Prod. Commun. 2021, 16, 7. [Google Scholar] [CrossRef]
- Tziveleka, L.-A.; Tammam, M.A.; Tzakou, O.; Roussis, V.; Ioannou, E. Metabolites with Antioxidant Activity from Marine Macroalgae. Antioxidants 2021, 10, 1431. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Gao, N.; Zang, Z.; Meng, X.; Lin, Y.; Yang, S.; Yang, Y.; Jin, Z.; Li, B. Classification and antioxidant assays of polyphenols: A review. J. Fut. Foods 2024, 4, 193–204. [Google Scholar] [CrossRef]
- Dong, H.; Dong, S.; Erik Hansen, P.; Stagos, D.; Lin, X.; Liu, M. Progress of Bromophenols in Marine Algae from 2011 to 2020: Structure, Bioactivities, and Applications. Mar. Drugs 2020, 18, 411. [Google Scholar] [CrossRef]
- Zheng, H.; Zhao, Y.; Guo, L. A Bioactive Substance Derived from Brown Seaweeds: Phlorotannins. Mar. Drugs 2022, 20, 742. [Google Scholar] [CrossRef]
- Leyton, A.; Pezoa-Conte, R.; Barriga, A.; Buschmann, A.H.; Mäki-Arvela, P.; Mikkola, J.P.; Lienqueo, M.E. Identification and efficient extraction method of phlorotannins from the brown seaweed Macrocystis pyrifera using an orthogonal experimental design. Algal Res. 2016, 16, 201–208. [Google Scholar] [CrossRef]
- Taniguchi, R.; Ito, C.; Keitoku, S.; Miyake, Y.; Itoigawa, M.; Matsui, T.; Shibata, T. Analysis on the structure of phlorethols isolated from the warm-temperate brown seaweed Sargassum carpophyllum and their antioxidant properties. Nat. Prod. Commun. 2022, 17. [Google Scholar] [CrossRef]
- Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Barba, F.J.; Lorenzo, J.M. Antioxidant Potential of Extracts Obtained from Macro- (Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata) and Micro-Algae (Chlorella vulgaris and Spirulina platensis) Assisted by Ultrasound. Medicines 2018, 5, 33. [Google Scholar] [CrossRef]
- Cichoński, J.; Chrzanowski, G. Microalgae as a Source of Valuable Phenolic Compounds and Carotenoids. Molecules 2022, 27, 8852. [Google Scholar] [CrossRef]
- Andriopoulos, V.; Gkioni, M.D.; Koutra, E.; Mastropetros, S.G.; Lamari, F.N.; Hatziantoniou, S.; Kornaros, M. Total Phenolic Content, Biomass Composition, and Antioxidant Activity of Selected Marine Microalgal Species with Potential as Aquaculture Feed. Antioxidants 2022, 11, 1320. [Google Scholar] [CrossRef]
- Fernando, I.P.; Kim, M.; Son, K.T.; Jeong, Y.; Jeon, Y.J. Antioxidant Activity of Marine Algal Polyphenolic Compounds: A Mechanistic Approach. J. Med. Food 2016, 19, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, 13394. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, O.V.; do Rosário, R.C.; Teixeira-Costa, B.E. Sources of Carotenoids in Amazonian Fruits. Molecules 2024, 29, 2190. [Google Scholar] [CrossRef] [PubMed]
- Generalić Mekinić, I.; Šimat, V.; Rathod, N.B.; Hamed, I.; Čagalj, M. Algal Carotenoids: Chemistry, Sources, and Application. Foods 2023, 12, 2768. [Google Scholar] [CrossRef]
- Baeza-Morales, A.; Medina-García, M.; Martínez-Peinado, P.; Pascual-García, S.; Pujalte-Satorre, C.; López-Jaén, A.B.; Martínez-Espinosa, R.M.; Sempere-Ortells, J.M. The Antitumour Mechanisms of Carotenoids: A Comprehensive Review. Antioxidants 2024, 13, 1060. [Google Scholar] [CrossRef]
- Ren, Y.; Sun, H.; Deng, J.; Huang, J.; Chen, F. Carotenoid Production from Microalgae: Biosynthesis, Salinity Responses and Novel Biotechnologies. Mar. Drugs 2021, 19, 713. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Gómez-Villegas, P.; Gonda, M.L.; León-Vaz, A.; León, R.; Mildenberger, J.; Rebours, C.; Saravia, V.; Vero, S.; Vila, E.; et al. Microalgae, Seaweeds and Aquatic Bacteria, Archaea, and Yeasts: Sources of Carotenoids with Potential Antioxidant and Anti-Inflammatory Health-Promoting Actions in the Sustainability Era. Mar. Drugs 2023, 21, 340. [Google Scholar] [CrossRef]
- Murthy, K.C.; Vanitha, A.; Rajesha, J.; Swamy, M.M.; Sowmya, P.R.; Ravishankar, G.A. In vivo Antioxidant Activity of Carotenoids from Dunaliella salina—A Green Microalga. Life Sci. 2005, 76, 1381–1390. [Google Scholar] [CrossRef]
- Ahmad, M.T.; Shariff, M.; Yusoff, F.; Goh, Y.M.; Banerjee, S. Applications of Microalga Chlorella vulgaris in Aquaculture. Rev. Aquac. 2020, 12, 328–346. [Google Scholar] [CrossRef]
- Kurniawan, R.; Nurkolis, F.; Taslim, N.A.; Subali, D.; Surya, R.; Gunawan, W.B.; Alisaputra, D.; Mayulu, N.; Salindeho, N.; Kim, B. Carotenoids Composition of Green Algae Caulerpa racemosa and Their Antidiabetic, Anti-Obesity, Antioxidant, and Anti-Inflammatory Properties. Molecules 2023, 28, 3267. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and Chlorophylls as Antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-Y.; Huang, X.; Cheong, K.-L. Recent Advances in Marine Algae Polysaccharides: Isolation, Structure, and Activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef]
- Moreira, J.B.; Santos, T.D.; Cruz, C.G.; Silveira, J.T.d.; Carvalho, L.F.d.; Morais, M.G.d.; Costa, J.A.V. Algal Polysaccharides-Based Nanomaterials: General Aspects and Potential Applications in Food and Biomedical Fields. Polysaccharides 2023, 4, 371–389. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of In Vitro Antioxidant Activity of Polysaccharides. Oxid. Med. Cell Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef]
- Bai, L.; Xu, D.; Zhou, Y.-M.; Zhang, Y.-B.; Zhang, H.; Chen, Y.-B.; Cui, Y.-L. Antioxidant Activities of Natural Polysaccharides and Their Derivatives for Biomedical and Medicinal Applications. Antioxidants 2022, 11, 2491. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Anastyuk, S.D.; Ishina, I.A.; Isakov, V.V.; Zvyagintseva, T.N.; Thinh, P.D.; Zadorozhny, P.A.; Dmitrenok, P.S.; Ermakova, S.P. Structural Characteristics and Anticancer Activity in Vitro of Fucoidan from Brown Alga Padina boryana. Carbohydr Polym. 2018, 15, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Xu, R.Y.; Zhou, W.Z.; Sheng, X.B.; Yang, A.Y.; Cheng, J.L. Effects of Fucoidan Extracted from Brown Seaweed on Lipid Peroxidation in Mice. Acta Nutrim. Sin. 2002, 24, 389–392. [Google Scholar]
- Rocha de Souza, M.C.; Marques, C.T.; Guerra Dore, C.M.; Ferreira da Silva, F.R.; Oliveira Rocha, H.A.; Leite, E.L. Antioxidant Activities of Sulfated Polysaccharides from Brown and Red Seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Ward, F.; Deyab, M.A.; Al-Zahrani, M.; Touliabah, H.E. Chemical Composition, Antioxidant, and Antitumor Activity of Fucoidan from the Brown Alga Dictyota dichotoma. Molecules 2023, 28, 7175. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.; Zhao, R. Fucoidan: Structure and Bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef]
- Pacheco-Quito, E.M.; Ruiz-Caro, R.; Veiga, M.D. Carrageenan: Drug Delivery Systems and Other Biomedical Applications. Mar. Drugs 2020, 18, 583. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, A.; Romeis, D.; Hupfeld, E.; Sieber, V. Biocatalytic Conversion of Carrageenans for the Production of 3,6-Anhydro-D-galactose. J. Agric. Food Chem. 2024, 72, 5816–5827. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.T.; Cuong, D.X.; Thuy, L.H.; Thuan, P.T.; Tuyen, D.T.T.; Mo, V.T.; Dong, D.H. Carrageenan of Red Algae Eucheuma gelatinae: Extraction, Antioxidant Activity, Rheology Characteristics, and Physicochemistry Characterization. Molecules 2022, 27, 1268. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chang, J.S. Proteins and Bioactive Peptides From Algae: Insights Into Antioxidant, Anti-hypertensive, Anti-diabetic and Anti-cancer Activities. Trends Food Sci. Technol. 2024, 145, 104352. [Google Scholar] [CrossRef]
- López-García, G.; Dublan-García, O.; Arizmendi-Cotero, D.; Gómez Oliván, L.M. Antioxidant and Antimicrobial Peptides Derived from Food Proteins. Molecules 2022, 27, 1343. [Google Scholar] [CrossRef]
- Jafarirad, S.; Nateghi, L.; Moslemi, M.; Afshari, K.P. Functional Properties of the Bioactive Peptides Derived from Sargassum angustifolium algae. Food Meas. 2023, 17, 6588–6599. [Google Scholar] [CrossRef]
- Li, Z.; He, H.; Liu, J.; Gu, H.; Fu, C.; Zeb, A.; Che, T.; Shen, S. Preparation and Vasodilation Mechanism of Angiotensin-I-Converting Enzyme Inhibitory Peptide from Ulva prolifera Protein. Mar. Drugs 2024, 22, 398. [Google Scholar] [CrossRef]
- Fernando, R.; Sun, X.; Rupasinghe, H.P.V. Production of Bioactive Peptides from Microalgae and Their Biological Properties Related to Cardiovascular Disease. Macromol 2024, 4, 582–596. [Google Scholar] [CrossRef]
- Ko, S.C.; Kim, D.; Jeon, Y.J. Protective Effect of a Novel Antioxidative Peptide Purified from a Marine Chlorella ellipsoidea Protein against Free Radical-Induced Oxidative Stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef]
- Chénais, B. Algae and Microalgae and Their Bioactive Molecules for Human Health. Molecules 2021, 26, 1185. [Google Scholar] [CrossRef] [PubMed]
- Caturano, A.; D’Angelo, M.; Mormone, A.; Russo, V.; Mollica, M.P.; Salvatore, T.; Galiero, R.; Rinaldi, L.; Vetrano, E.; Marfella, R.; et al. Oxidative Stress in Type 2 Diabetes: Impacts from Pathogenesis to Lifestyle Modifications. Curr. Issues Mol. Biol. 2023, 45, 6651–6666. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Mabhida, S.E.; Ziqubu, K.; Nkambule, B.B.; Mazibuko-Mbeje, S.E.; Hanser, S.; Basson, A.K.; Pheiffer, C.; Kengne, A.P. Pancreatic β-cell dysfunction in type 2 diabetes: Implications of Inflammation and Oxidative Stress. World J. Diabetes 2023, 14, 130–146. [Google Scholar] [CrossRef] [PubMed]
- Giacco, F.; Brownlee, M. Oxidative Stress and Diabetic Complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Alwaleed, E.A.; Jillany, A.; Kasem, N.R.; Galal, H. Evaluation of the Pancreatoprotective Effect of Algal Extracts on Alloxan-induced Diabetic Rat. Bioact. Carbohydr. Diet. Fibre 2020, 24, 100237. [Google Scholar] [CrossRef]
- El Habitri, N.; Belkacemi, L. Antidiabetic Effect of Oral Supplementation with Caulerpa racemosa Powder. Eur. J. Biol. Res. 2022, 12, 141–152. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; Kasem, N.R.; Alsoghier, H.M.; Jillany, A.; Galal, H.; Alwaleed, E.A. Efficacy of The Marine Red Alga Laurencia papillosa Extract on Alloxan Stimulated Hyperglycemic Activity in Male Wistar Albino Rats. Bioact. Carbohydr. Diet. Fibre 2024, 31, 100403. [Google Scholar] [CrossRef]
- Alrasheedi, A.A.; Basnawi, A.I.; Althaiban, M.A. Effects of Spirulina platensis Microalgae on Renal Function and Antioxidant Defense in Rats with Streptozotocin-induced Diabetes. J. Funct. Foods 2024, 122, 106485. [Google Scholar] [CrossRef]
- Guldas, M.; Ziyanok-Demirtas, S.; Sahan, Y.; Yildiz, E.; Gurbuz, O. Antioxidant and Anti-diabetic Properties of Spirulina platensis Produced in Turkey. Food Sci Technol. 2020, 41, 615–625. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, M.H.; Heo, S.J.; Kang, S.M.; Ko, S.C.; Han, J.S.; Jeon, Y.J. Dieckol Isolated From Ecklonia cava Inhibits α-glucosidase and α-amylase in vitro and Alleviates Postprandial Hyperglycemia in Streptozotocin-induced Diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef]
- Hwang, P.A.; Hung, Y.L.; Tsai, Y.K.; Chien, S.Y.; Kong, Z.L. The Brown Seaweed Sargassum hemiphyllum Exhibits α-amylase and α-glucosidase Inhibitory Activity and Enhances Insulin Release in vitro. Cytotechnology 2015, 67, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, S.; Hosokawa, M.; Miyashita, K. Fucoxanthin Promotes Translocation and Induction of Glucose Transporter 4 in Skeletal Muscles of Diabetic/obese KK-Ay Mice. Phytomedicine 2012, 19, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.R.; Rhyu, D.Y. Anti-diabetic Effects of Caulerpa lentillifera: Stimulation of Insulin Secretion in Pancreatic β-cells and Enhancement of Glucose Uptake in Adipocytes. Asian Pac. J. Trop. Biomed. 2014, 4, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Gazali, M.; Jolanda, O.; Husni, A.; Nurjanah, F.A.; Majid, A.; Syafitri, R. In Vitro α-Amylase and α-Glucosidase Inhibitory Activity of Green Seaweed Halimeda tuna Extract from the Coast of Lhok Bubon, Aceh. Plants 2023, 12, 393. [Google Scholar] [CrossRef]
- Ming, J.X.; Wang, Z.C.; Huang, Y.; Ohishi, H.; Wu, R.J.; Shao, Y.; Wang, H.; Qin, M.Y.; Wu, Z.L.; Li, Y.Y.; et al. Fucoxanthin Extracted from Laminaria Japonica Inhibits Metastasis and Enhances the Sensitivity of Lung Cancer to Gefitinib. J. Ethnopharmacol. 2021, 265, 113302. [Google Scholar] [CrossRef]
- Senthilkumar, D.; Jayanthi, S. Partial Characterization and Anticancer Activities of Purified Glycoprotein Extracted from Green Seaweed Codium decorticatum. J. Funct. Foods 2016, 25, 323–332. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Abdelnour, S.; Alagawany, M.; Abdo, M.; Sakr, M.A.; Khafaga, A.F.; Mahgoub, S.A.; Elnesr, S.S.; Gebriel, M.G. Microalgae in Modern Cancer Therapy: Current knowledge. Biomed. Pharmacother. 2019, 111, 42–50. [Google Scholar] [CrossRef]
- Darvish, M.; Jalili, H.; Ranaei-Siadat, S.O.; Sedighi, M. Potential Cytotoxic Effects of Peptide Fractions from Dunaliella salina protein Hydrolyzed by Gastric Proteases. J. Aquat. Food Prod. Technol. 2017, 27, 165–175. [Google Scholar] [CrossRef]
- Algotiml, R.; Gab-Alla, A.; Seoudi, R.; Abulreesh, H.H.; El-Readi, M.Z.; Elbanna, K. Anticancer and Antimicrobial Activity of Biosynthesized Red Sea Marine Algal Silver Nanoparticles. Scient. Rep. 2022, 12, 2421. [Google Scholar] [CrossRef]
- Janani, G.; Girigoswami, A.; Deepika, B.; Udayakumar, S.; Girigoswami, K. Unveiling the Role of Nano-Formulated Red Algae Extract in Cancer Management. Molecules 2024, 29, 2077. [Google Scholar] [CrossRef]
- Saeed, A.; Abotaleb, S.; Alam, N.; ELMehalawy, A.; Gheda, S. In vitro Assessment of Antimicrobial, Antioxidant and Anticancer Activities of Some Marine Macroalgae. Egypt. J. Bot. 2020, 60, 81–96. [Google Scholar]
- Gong, J.E.; Kim, J.E.; Park, S.H. Anti-tumor Effects of an Aqueous Extract of Ecklonia cava in BALB/cKorl Syngeneic Mice Using Colon Carcinoma CT26 cells. Oncol. Rep. 2023, 49, 128. [Google Scholar] [CrossRef] [PubMed]
- Lau, V.; Nurkolis, F.; Park, M.N.; Heriyanto, D.S.; Taslim, N.A.; Tallei, T.E.; Permatasari, H.K.; Tjandrawinata, R.R.; Moon, S.; Kim, B. Green Seaweed Caulerpa racemosa as a Novel Non-Small Cell Lung Cancer Inhibitor in Overcoming Tyrosine Kinase Inhibitor Resistance: An Analysis Employing Network Pharmacology, Molecular Docking, and In Vitro Research. Mar. Drugs 2024, 22, 272. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, J.R.B.; Rodrigues, R.P.; Mazzuco, A.C.; de Cassia Ribeiro Gonçalves, R.; Bernardino, A.F.; Kuster, R.M.; Kitagawa, R.R. In Vitro and In Silico Evaluation of Red Algae Laurencia obtusa Anticancer Activity. Mar. Drugs 2023, 21, 318. [Google Scholar] [CrossRef] [PubMed]
- Bakunina, I.; Imbs, T.; Likhatskaya, G.; Grigorchuk, V.; Zueva, A.; Malyarenko, O.; Ermakova, S. Effect of Phlorotannins from Brown Algae Costaria costata on α-N-Acetylgalactosaminidase Produced by Duodenal Adenocarcinoma and Melanoma Cells. Mar. Drugs 2023, 21, 33. [Google Scholar] [CrossRef]
- Lekshmi, V.S.; Kurup, G.M. Anticoagulant Activities of Sulfated Polysaccharides From the Edible Marine Algae Padina tetrastromatica and Ulva fasciata: A combined in vitro and in vivo Approach. J. Pharmacogn. Phytochem. 2019, 8, 693–698. [Google Scholar]
- Cao, S.; He, X.; Qin, L.; He, M.; Yang, Y.; Liu, Z.; Mao, W. Anticoagulant and Antithrombotic Properties in Vitro and in Vivo of a Novel Sulfated Polysaccharide from Marine Green Alga Monostroma nitidum. Mar. Drugs 2019, 17, 247. [Google Scholar] [CrossRef]
- Saidani, K.; Ziani, N.; Touati, N.; Merzouk, H.; Bedjou, F. Anticoagulant Activity of Sulfated Polysaccharides and Polyphenols Extracted from Marine Algae. Curr. Bioact. Comp. 2021, 17, 246–255. [Google Scholar] [CrossRef]
- Mousavian, Z.; Safavi, M.; Azizmohseni, F.; Hadizadeh, M.; Mirdamadi, S. Characterization, Antioxidant and Anticoagulant Properties of Exopolysaccharide from Marine Microalgae. AMB Express 2022, 12, 27. [Google Scholar] [CrossRef]
- Rodrigues, J.A.G.; Queiroz, I.N.L.D.; Quinderé, A.L.G.; Vairo, B.C.; Mourão, P.A.D.S.; Benevides, N.M.B. An Antithrombin-dependent Sulfated Polysaccharide Isolated from the Green Alga Caulerpa cupressoides has in vivo Anti-and Prothrombotic Effects. Ciência Rural 2011, 41, 634–639. [Google Scholar] [CrossRef]
- Qi, X.; Mao, W.; Gao, Y.; Chen, Y.; Chen, Y.; Zhao, C.; Shan, J. Chemical Characteristic of an Anticoagulant-active Sulfated Polysaccharide from Enteromorpha clathrata. Carbohydr. Polyms. 2012, 90, 1804–1810. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Liu, X.; He, X.; Wang, S.; Cao, S.; Xia, Z.; Mao, W. Structure and Anticoagulant Property of a Sulfated Polysaccharide Isolated from the Green Seaweed Monostroma angicava. Carbohydr. Polym. 2017, 159, 195–206. [Google Scholar] [CrossRef]
- Melo, F.R.; Mourão, P.A. An Algal Sulfated Galactan has an Unusual Dual Effect on Venous Thrombosis Due to Activation of Factor XII and Inhibition of the Coagulation Proteases. Thromb. Haemost. 2008, 99, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Mao, W.; Yan, M.; Liu, X.; Xia, Z.; Wang, S.; Xiao, B.; Chen, C.; Zhang, L.; Cao, S. Structural Characterization and Anticoagulant Activity of a Sulfated Polysaccharide from the Green Alga Codium divaricatum. Carbohydr. Polym. 2015, 5, 175–182. [Google Scholar] [CrossRef]
- Liu, N.; Lyu, X.; Zhang, X.; Zhang, F.; Chen, Y.; Li, G. Astaxanthin Attenuates Cognitive Deficits in Alzheimer’s Disease Models by Reducing Oxidative Stress Via the SIRT1/PGC-1α Signaling Pathway. Cell Biosci. 2023, 13, 173. [Google Scholar] [CrossRef]
- Bogie, J.; Hoeks, C.; Schepers, M.; Tiane, A.; Cuypers, A.; Leijten, F.; Chintapakorn, Y.; Suttiyut, T.; Pornpakakul, S.; Struik, D.; et al. Dietary Sargassum fusiforme Improves Memory and Reduces Amyloid Plaque Load in an Alzheimer’s Disease Mouse Model. Sci. Rep. 2019, 9, 4908. [Google Scholar] [CrossRef]
- Xiao, S.; Chan, P.; Wang, T.; Hong, Z.; Wang, S.; Kuang, W.; He, J.; Pan, X.; Zhou, Y.; Ji, Y.; et al. A 36-week Multicenter, Randomized, Double-blind, Placebo-controlled, Parallel-group, Phase 3 Clinical Trial of Sodium Oligomannate for Mild-to-moderate Alzheimer’s Dementia. Alzheimers Res. Ther. 2021, 13, 62. [Google Scholar] [CrossRef]
- Syed, Y.Y. Sodium Oligomannate: First Approval. Drugs 2020, 80, 441–444, Erratum in Drugs 2020, 80, 445–446.. [Google Scholar] [CrossRef]
- Lai, Y.-J. Omega-3 Fatty Acid Obtained from Nannochloropsis oceanica Cultures Grown Under Low Urea Protect Against Aβ-induced Neural Damage. J. Food Sci. Technol. 2015, 52, 2982–2989. [Google Scholar] [CrossRef]
- Yang, M.; Jin, L.; Wu, Z.; Xie, Y.; Zhang, P.; Wang, Q.; Yan, S.; Chen, B.; Liang, H.; Naman, C.B.; et al. PLGA-PEG Nanoparticles Facilitate In Vivo Anti-Alzheimer’s Effects of Fucoxanthin, a Marine Carotenoid Derived from Edible Brown Algae. J. Agric. Food Chem. 2021, 69, 9764–9777. [Google Scholar] [CrossRef]
- Tsang, C.K.; Ina, A.; Goto, T.; Kamei, Y. Sargachromenol, a novel nerve growth factor-potentiating substance isolated from Sargassum macrocarpum, promotes neurite outgrowth and survival via distinct signaling pathways in PC12D cells. Neuroscience 2005, 132, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Zhang, Q.; Wang, H.; Cui, Y.; Sun, Z.; Yang, J.; Zheng, Y.; Jia, J.; Yu, F.; Wang, X.; et al. Fucoidan Protects Against Dopaminergic Neuron Death in vivo and in vitro. Eur. J. Pharmacol. 2009, 617, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.A.; Xu, L.; Jin, L.M.; Wang, B.X.; Fu, C.Z.; Bai, Y.; Wu, H.G. κ-Carrageenan Oligosaccharides Inhibit the Inflammation of Lipopolysaccharide-Activated Microglia Via TLR4/NF-κB and p38/JNK MAPKs Pathways. Neurochem Res. 2022, 47, 295–304. [Google Scholar] [CrossRef]
- Moheimanian, N.; Mirkhani, H.; Purkhosrow, A.; Sohrabipour, J.; Jassbi, A.R. In Vitro and In Vivo Antidiabetic, α-Glucosidase Inhibition and Antibacterial Activities of Three Brown Algae, Polycladia myrica, Padina antillarum, and Sargassum boveanum, and a Red Alga, Palisada perforata from the Persian Gulf. Iran J. Pharm. Res. 2023, 22, 133731. [Google Scholar] [CrossRef] [PubMed]
- Baba, A.I.; Câtoi, C. Carcinogenesis. In Comparative Oncology. Bucharest (RO): The Publishing House of the Romanian Academy; 2007. Available online: https://www.ncbi.nlm.nih.gov/books/NBK9552/ (accessed on 14 December 2024).
- Afzal, S.; Abdul; Manap, A.S.; Attiq, A.; Albokhadaim, I.; Kandeel, M.; Alhojaily, S.M. From Imbalance to Impairment: The Central Role of Reactive Oxygen Species in Oxidative Stress-induced Disorders and Therapeutic Exploration. Front. Pharmacol. 2023, 14, 1269581. [Google Scholar] [CrossRef]
- Lichota, A.; Szewczyk, E.M.; Gwozdzinski, K. Factors Affecting the Formation and Treatment of Thrombosis by Natural and Synthetic Compounds. Int. J. Mol. Sci. 2020, 21, 7975. [Google Scholar] [CrossRef]
- Masselli, E.; Pozzi, G.; Vaccarezza, M.; Mirandola, P.; Galli, D.; Vitale, M.; Carubbi, C.; Gobbi, G. ROS in Platelet Biology: Functional Aspects and Methodological Insights. Int. J. Mol. Sci. 2020, 21, 4866. [Google Scholar] [CrossRef]
- Li, P.; Ma, X.; Huang, G. Understanding Thrombosis: The Critical Role of Oxidative Stress. Hematology 2024, 29, 2301633. [Google Scholar] [CrossRef]
- Olufunmilayo, E.O.; Gerke-Duncan, M.B.; Holsinger, R.M.D. Oxidative Stress and Antioxidants in Neurodegenerative Disorders. Antioxidants 2023, 12, 517. [Google Scholar] [CrossRef]
- Perluigi, M.; Di Domenico, F.; Butterfield, D.A. Oxidative Damage in Neurodegeneration: Roles in the Pathogenesis and Progression of Alzheimer Disease. Physiol. Rev. 2024, 104, 103–197. [Google Scholar] [CrossRef]
- Dionísio, P.A.; Amaral, J.D.; Rodrigues, C.M.P. Oxidative Stress and Regulated Cell Death in Parkinson’s Disease. Ageing Res. Rev. 2021, 67, 101263. [Google Scholar] [CrossRef]
- Hemerková, P.; Vališ, M. Role of Oxidative Stress in the Pathogenesis of Amyotrophic Lateral Sclerosis: Antioxidant Metalloenzymes and Therapeutic Strategies. Biomolecules 2021, 11, 437. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, F.; Fan, X.; Chen, M.; Xu, X.; Xu, Q.; Zhu, H.; Xu, A.; Pouladi, M.A.; Xu, X. CHCHD2 up-regulation in Huntington Disease Mediates a Compensatory Protective Response Against Oxidative Stress. Cell Death Dis. 2024, 15, 126. [Google Scholar] [CrossRef] [PubMed]
- Guardado Yordi, E.; Pérez Martínez, A.; Radice, M.; Scalvenzi, L.; Abreu-Naranjo, R.; Uriarte, E.; Santana, L.; Matos, M.J. Seaweeds as Source of Bioactive Pigments with Neuroprotective and/or Anti-Neurodegenerative Activities: Astaxanthin and Fucoxanthin. Mar. Drugs 2024, 22, 327. [Google Scholar] [CrossRef] [PubMed]
- Calon, F.; Cole, G. Neuroprotective Action of Omega-3 Polyunsaturated Fatty Acids Against Neurodegenerative Diseases: Evidence from Animal Studies. Prostaglandins Leukot. Essent. Fat. Acids 2007, 77, 287–293. [Google Scholar] [CrossRef]
- Quitério, E.; Grosso, C.; Ferraz, R.; Delerue-Matos, C.; Soares, C. A Critical Comparison of the Advanced Extraction Techniques Applied to Obtain Health-Promoting Compounds from Seaweeds. Mar. Drugs 2022, 20, 677. [Google Scholar] [CrossRef]
- Michalak, I.; Chojnacka, K. Algal Extracts: Technology and Advances. Eng. Life Sci. 2014, 14, 581–591. [Google Scholar] [CrossRef]
- Streimikyte, P.; Viskelis, P.; Viskelis, J. Enzymes-Assisted Extraction of Plants for Sustainable and Functional Applications. Int. J. Mol. Sci. 2022, 23, 2359. [Google Scholar] [CrossRef]
- Jeon, Y.-J.; Wijesinghe, W.A.J.P.; Kim, S.-K. Enzyme-assisted Extraction and Recovery of Bioactive Components from Seaweeds. In Hand Book of Marine Macroalgae: Biotechnology and Applied Phycology, 1st ed.; Kim, S.-K., Ed.; John Willy & Sons, Ltd.: New York, NY, USA, 2011. [Google Scholar] [CrossRef]
- Habeebullah, S.; Alagarsamy, S.; Sattari, Z.; Al-Haddad, S.; Fakhraldeen, S.; Al-Ghumaim, A.; Al-Yamani, F. Enzyme-assisted extraction of bioactive compounds from brown seaweeds and characterization. J. Appl. Phycol. 2020, 32, 615–629. [Google Scholar] [CrossRef]
- Billakanti, J.M.; Catchpole, O.; Fenton, T.; Mitchell, K. Extraction of Fucoxanthin from Undaria pinnatifida Using Enzymatic Pre-treatment Followed by DME and EtOH co-solvent Extraction. In Proceedings of the 10th International Symposium on Supercritical Fluids, San Francisco, CA, USA, 13–16 May 2012; King, J., Ed.; CASSS: Emeryville, CA, USA, 2012. [Google Scholar]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Donnay-Moreno, C.; Bergé, J.P.; Nyvall-Collén, P.; Bourgougnon, N. Enzyme-assisted Extraction (EAE) for the Production of Antiviral and Antioxidant Extracts from the Green Seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 2016, 16, 233–239. [Google Scholar] [CrossRef]
- Nayak, N.; Bhujle, R.R.; Nanje-Gowda, N.A.; Chakraborty, S.; Siliviru, K.; Subbiah, J.; Brennan, C. Advances in the Novel and Green-assisted Techniques for Extraction of Bioactive Compounds from Millets: A comprehensive review. Heliyon 2024, 10, e30921. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Barrutia, M.B.; Iriondo-DeHond, A.; Arcia, P.; Cozzano Ferreira, S.; Castillo, M. Enzyme assisted extraction of health promoting compounds from extruded brewers’ spent grain (BSG). In Proceedings of the XXV Jornadas Internacionales de Nutrición Práctica, Online, 20–22 April 2021; Available online: http://hdl.handle.net/10261/253416 (accessed on 28 December 2024).
- Barbosa, P.P.M.; Ruviaro, A.R.; Macedo, G.A. Conditions of Enzyme-assisted Extraction to Increase the Recovery of Flavanone Aglycones from Pectin Waste. J. Food Sci. Technol. 2021, 58, 4303–4312. [Google Scholar] [CrossRef] [PubMed]
- López-Salazar, H.; Camacho-Díaz, B.H.; Ocampo, M.A.; Jiménez-Aparicio, A.R. Microwave-assisted Extraction of Functional Compounds from Plants: A Review. Bioresources 2023, 18, 6614. [Google Scholar] [CrossRef]
- André, J.; Flórez-Fernández, N.; Domínguez, H.; Torres, M.D. Microwave-assisted Extraction of Ulva spp. Including a Stage of Selective Coagulation of Ulvan Stimulated by a Bio-ionic Liquid. Int. J. Biol. Macromol. 2023, 225, 952–963. [Google Scholar] [CrossRef]
- Nesic, A.; De Bonis, M.V.; Dal Poggetto, G.; Ruocco, G.; Santagata, G. Microwave Assisted Extraction of Raw Alginate as a Sustainable and Cost-Effective Method to Treat Beach-Accumulated Sargassum Algae. Polymers 2023, 15, 2979. [Google Scholar] [CrossRef]
- Cantarino, S.J.; Coutinho, R.; Soares, A.R.; Duarte, H.M.; Martinez, S.T. Microwave irradiation is a suitable method for caulerpin extraction from the green algae Caulerpa racemosa (Chlorophyta, Caulerpaceae). Nat. Prod. Res. 2022, 36, 2149–2153. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Allen, J.D.; Kanitkar, A.; Boldor, D. Oil Extraction from Scenedesmus obliquus Using a Continuous Microwave System–Design, Optimization, and Quality Characterization. Bioresour Technol. 2011, 102, 3396–3403. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Butler, T.O.; Pandhal, J.; Vaidyanathan, S. Microwave-Assisted Extraction for Microalgae: From Biofuels to Biorefinery. Biology 2018, 7, 18. [Google Scholar] [CrossRef]
- Kumar, K.; Srivastav, S.; Sharanagat, V.S. Ultrasound assisted extraction (UAE) of Bioactive Compounds From Fruit and Vegetable Processing by-products: A review. Ultrason. Sonochem. 2021, 70, 105325. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; Smyth, T.J.; O’Donnell, C.P. Optimization of ultrasound assisted extraction of bioactive components from brown seaweed Ascophyllum nodosum using response surface methodology. Ultrason. Sonochem. 2015, 23, 308–316. [Google Scholar] [CrossRef]
- Kumar, Y.; Singhal, S.; Tarafdar, A.; Pharande, A.; Ganesan, M.; Badgujar, P.C. Ultrasound Assisted Extraction of Selected Edible Macroalgae: Effect on Antioxidant Activity and Quantitative Assessment of Polyphenols by Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS). Algal Res. 2020, 52, 102114. [Google Scholar] [CrossRef]
- Carreira-Casais, A.; Otero, P.; Garcia-Perez, P.; Garcia-Oliveira, P.; Pereira, A.G.; Carpena, M.; Soria-Lopez, A.; Simal-Gandara, J.; Prieto, M.A. Benefits and Drawbacks of Ultrasound-Assisted Extraction for the Recovery of Bioactive Compounds from Marine Algae. Int. J. Environ. Res. Public Health 2021, 18, 9153. [Google Scholar] [CrossRef] [PubMed]
- Vernès, L.; Abert-Vian, M.; El Maâtaoui, M.; Tao, Y.; Bornard, I.; Chemat, F. Application of Ultrasound for Green Extraction of Proteins from Spirulina. Mechanism, Optimization, Modeling, and Industrial Prospects. Ultrason. Sonochem. 2019, 54, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Mahali, M.; Sibi, G. Extraction Methods and Functional Properties of Protein from Arthospira platensis for Bioavailability of Algal Proteins. Int. J. Pharm. Chem. 2019, 5, 20. [Google Scholar] [CrossRef]
- Zhao, G.; Chen, X.; Wang, L.; Zhou, S.; Feng, H.; Chen, W.N.; Lau, R. Ultrasound Assisted Extraction of Carbohydrates from Microalgae as Feedstock for Yeast Fermentation. Bioresour. Technol. 2013, 128, 337–344. [Google Scholar] [CrossRef]
- Adam, F.; Abert-Vian, M.; Peltier, G.; Chemat, F. “Solvent-free” Ultrasound-assisted Extraction of Lipids from Fresh Microalgae Cells: A Green, Clean and Scalable Process. Bioresour. Technol. 2012, 114, 457–465. [Google Scholar] [CrossRef]
- Jadhav, H.B.; Gogate, P.; Annapure, U. Analyzing the Repercussions of Ultrasound on Triacylglycerols in Food. Food Chem. Adv. 2023, 2, 100332. [Google Scholar] [CrossRef]
- Madureira, J.; Barros, L.; Cabo Verde, S.; Margaça, F.M.A.; Santos-Buelga, C.; Ferreira, I.C.F.R. Ionizing Radiation Technologies to Increase the Extraction of Bioactive Compounds from Agro-Industrial Residues: A Review. J. Agric. Food Chem. 2020, 68, 11054–11067. [Google Scholar] [CrossRef]
- Rojo del Olmo, L.; Pablo, M.J.; Croce, M.E.; Perez, M.B.; Parodi, R.R.; Andreucetti, N.A. Uso de la irradiación gamma en la extracción e identificación de compuestos bioactivos presentes en algas rojas. J. Argen. Chem. Soc. 2012, 99, 1–3. [Google Scholar]
- Choi, J.I.; Kim, H.J.; Lee, J.W. Structural Feature and Antioxidant Activity of Low Molecular Weight Laminarin Degraded by Gamma Irradiation. Food Chem. 2011, 129, 520–523. [Google Scholar] [CrossRef]
- He, D.; Yan, L.; Ma, X.; Cheng, Y.; Wu, S.; Zuo, J.; Park, E.-J.; Liu, J.; Wu, M.; Choi, J.-I.; et al. Gamma-irradiation Degraded Sulfated Polysaccharide from a New Red Algal Strain Pyropia yezoensis Sookwawon 104 with In Vitro Antiproliferative Activity. Oncol. Lett. 2020, 20, 91. [Google Scholar] [CrossRef] [PubMed]
- Herzyk, F.; Piłakowska-Pietras, D.; Korzeniowska, M. Supercritical Extraction Techniques for Obtaining Biologically Active Substances from a Variety of Plant Byproducts. Foods 2024, 13, 1713. [Google Scholar] [CrossRef] [PubMed]
- Samuel, H.S.; Nweke-Maraizu, U.; Etim, E.E. Supercritical Fluids: Properties, Formation and Applications. J. Eng. Ind. Res. 2023, 4, 176–188. [Google Scholar] [CrossRef]
- Gondo, T.F.; Jönsson, M.; Karlsson, E.N.; Sandahl, M.; Turner, C. Extractability, Selectivity, and Comprehensiveness in Supercritical Fluid Extraction of Seaweed Using Ternary Mixtures of Carbon Dioxide, Ethanol, and Water. J. Chromatogr. A 2023, 1706, 464267. [Google Scholar] [CrossRef]
- Becerra, M.; Boutefnouchet, S.; Cordoba, O.; Vitorino, G.P.; Brehu, L.; Lamour, I.; Laimay, F.; Efstathiou, A.; Smirlis, D.; Michel, S.; et al. Antileishmanial Activity of Fucosterol Recovered from Lessonia vadosa Searles (Lessoniaceae) by SFE, PSE and CPC. Phytochem. Lett. 2015, 11, 418–423. [Google Scholar] [CrossRef]
- Chatterjee, D.; Bhattacharjee, P. Supercritical Carbon Dioxide Extraction of Antioxidant Rich Fraction from Phormidium valderianum: Optimization of Experimental Process Parameters. Algal Res. 2014, 3, 49–54. [Google Scholar] [CrossRef]
- Cheriyan, B.V.; Karunakar, K.K.; Anandakumar, R.; Murugathirumal, A. Eco-Friendly Extraction Technologies: A Comprehensive Review of Modern Green Analytical Methods. Sustain. Chem. Clim. Action 2024, 6, 100054. [Google Scholar] [CrossRef]
- Barp, L.; Višnjevec, A.M.; Moret, S. Pressurized Liquid Extraction: A Powerful Tool to Implement Extraction and Purification of Food Contaminants. Foods 2023, 12, 2017. [Google Scholar] [CrossRef]
- Castro-Puyana, M.; Pérez-Sánchez, A.; Valdés, A.; Ibrahim, O.H.M.; Suárez-Alvarez, S.; Ferragut, J.A.; García-Cañas, V. Pressurized Liquid Extraction of Neochloris oleoabundans for the Recovery of Bioactive Carotenoids with Anti-proliferative Activity Against Human Colon Cancer Cells. Food Res. Int. 2017, 99, 1048–1055. [Google Scholar] [CrossRef]
- Anaëlle, T.; Leon, E.S.; Laurent, V.; Elena, I.; Mendiola, J.A.; Stéphane, C.; Nelly, K.; Stéphane, L.B.; Luc, M.; Valérie, S.P. Green Improved Processes to Extract Bioactive Phenolic Compounds from Brown Macroalgae Using Sargassum muticum as Model. Talanta 2013, 104, 44–52. [Google Scholar] [CrossRef]
- Gümüş Yılmaz, G.; Gómez Pinchetti, J.L.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Comparison of Extraction Techniques and Surfactants for the Isolation of Total Polyphenols and Phlorotannins from the Brown Algae Lobophora variegata. Anal. Lett. 2019, 52, 2724–2740. [Google Scholar] [CrossRef]
- Ristivojević, P.; Krstić Ristivojević, M.; Stanković, D.; Cvijetić, I. Advances in Extracting Bioactive Compounds from Food and Agricultural Waste and By-Products Using Natural Deep Eutectic Solvents: A Circular Economy Perspective. Molecules 2024, 29, 4717. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, Q.; Hou, Y. Efficient Purification of R-phycoerythrin from Marine Algae (Porphyra yezoensis) Based on a Deep Eutectic Solvents Aqueous Two-Phase System. Mar. Drugs 2020, 18, 618. [Google Scholar] [CrossRef] [PubMed]
- Obluchinskaya, E.D.; Daurtseva, A.V.; Pozharitskaya, O.N.; Flisyuk, E.V.; Shikov, A.N. Natural Deep Eutectic Solvents as Alternatives for Extracting Phlorotannins from Brown Algae, Pharm. Chem. J. 2019, 53, 243–247. [Google Scholar] [CrossRef]
- Wils, L.; Leman-Loubière, C.; Bellin, N.; Clément-Larosière, B.; Pinault, M.; Chevalier, S.; Enguehard-Gueiffier, C.; Bodet, C.; Boudesocque-Delaye, L. Natural deep eutectic solvent formulations for spirulina: Preparation, Intensification, and Skin Impact. Algal Res. 2021, 56, 102317. [Google Scholar] [CrossRef]
- Bocker, R.; Silva, E.K. Pulsed Electric Field Assisted Extraction of Natural Food Pigments and Colorings from Plant Matrices. Food Chem. X 2022, 15, 100398. [Google Scholar] [CrossRef]
- Postma, P.R.; Fernandes, D.A.; Timmermans, R.A.H.; Vermuë, M.H.; Barbosa, M.J.; Eppink, M.H.M.; Eppink, R.H.; Olivieri, G. Pulsed Electric Field for Protein Release of the Microalgae Chlorella vulgaris and Neochloris oleoabundans. Algal Res. 2017, 24, 181–187. [Google Scholar] [CrossRef]
- Einarsdóttir, R.; Þórarinsdóttir, K.A.; Aðalbjörnsson, B.V. Extraction of Bioactive Compounds from Alaria esculenta with Pulsed Electric Field. J. Appl. Phycol. 2022, 34, 597–608. [Google Scholar] [CrossRef]
- Kokkali, M.; Martí-Quijal, F.J.; Taroncher, M.; Ruiz, M.J.; Kousoulaki, K.; Barba, F.J. Improved Extraction Efficiency of Antioxidant Bioactive Compounds from Tetraselmis chuii and Phaedoactylum tricornutum Using Pulsed Electric Fields. Molecules. 2020, 25, 3921. [Google Scholar] [CrossRef]
- Aouir, A.; Amiali, M.; Kirilova-Gachovska, T.; Benchabane, A.; Bitam, A. The Effect of Pulsed Electric Field (PEF) and Ultrasoud (US) Technologies on the Extraction of Phycopiliproteins from Arthrospira platensis. In Proceedings of the IEEE Canada International Humanitarian Technology Conference, Ottawa, ON, Canada, 31 May–4 June 2015; IEEE: Piscataway, NJ, USA; pp. 1–4. [Google Scholar] [CrossRef]
- Toepfl, S.; Heinz, V.; Knorr, D. Applications of Pulsed Electric Fields Technology for the Food Industry. In Pulsed Electric Fields Technology for the Food Industry, 2nd ed.; Raso, J., Heinz, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 197–221. [Google Scholar]
- Toepfl, S. Pulsed Electric Fields (PEF) for Permeabilization of Cell Membranes in Food- and Bioprocessing. Applications, Process and Equipment Design and Cost Analysis. Ph.D. Thesis, Technische Universität Berlin, Berlin, Germany, 2006. [Google Scholar]
| Type of Compound | Description | Antioxidant Mechanisms |
|---|---|---|
| Phenolic Compounds | Secondary metabolites found in marine algae. The most abundant phenolic compound in marine algae is typically phlorotannin as phloroglucinol. They have a hydroxyl group (-OH) attached to a phenolic ring, enabling them to capture free radicals and ROS.
| 1. Hydrogen Atom Transfer (HAT): Donates a hydrogen atom to free radicals. 2. Single Electron Transfer (SET): Transfers an electron to free radicals. 3. Sequential Proton Loss Electron Transfer (SPLET): Donates a proton followed by an electron. 4. Transition Metal Chelation: Prevents radical formation via pro-oxidant reactions. |
| Carotenoids | Natural pigments found widely in nature are responsible for yellow, red, and orange colors. They are made up of isoprene (C5) units. Examples include astaxanthin and fucoxanthin.
| 1. Electron Transfer: Neutralizes high-energy radicals like singlet oxygen. 2. Hydrogen Atom Transfer: Neutralizes peroxyl radicals. 3. Radical Cation Formation: Reacts with other molecules or is regenerated by other antioxidants like Vitamin C and α-tocopherol. |
| Polysaccharides | Algae contain polysaccharides, such as alginates, carrageenans, fucoidans, and laminarin. These compounds have antioxidant and radical scavenging properties.
| 1. Free Radical Scavenging: Neutralizes reactive oxygen and nitrogen species. 2. Metal Chelation: Inhibits free radical formation via Fenton reactions. 3. Molecular Structure Modification: Reducing molecular weight can enhance antioxidant capacity. |
| Peptides | Bioactive peptides are derived from proteins and have antioxidant activity related to their amino acid sequence and structure. Found in algae and microorganisms.
| 1. Metal Chelation: Inhibits radical production by stabilizing pro-oxidant metals. 2. Radical Inhibition: Stabilizes radicals by donating electrons. 3. Physical Shielding: Prevents lipid peroxidation. |
| Pigments | Marine algae contain a variety of pigments that allow them to capture light for photosynthesis and perform antioxidant and protective functions. Their pigment composition varies depending on the group to which they belong.
| 1. Radical Inhibition: Chlorophyll can donate electrons to free radicals, neutralizing them and preventing cellular damage. It is especially effective against hydroxyl and peroxyl radicals. 2. Metal Chelation: Chlorophyll can bind to transition metals, such as iron, which can catalyze the formation of free radicals. 3. Physical Quenching: converting it into triplet oxygen, a less harmful form. |
| Phycobiliproteins | Phycobiliproteins are accessory protein-pigments primarily found in red algae (Rhodophyta) and cyanobacteria, where they play a crucial role in light capture for photosynthesis.
| 1. Free Radical Scavenging: Selenium-containing phycocyanin scavenges free radicals like alkoxyl, superoxide, and hydrogen peroxide. The covalently-linked tetrapyrrole chromatophore phycocyanobilin is suggested to be involved in the scavenging activity of phycocyanin. |
| Vitamins | Marine algae provide essential vitamins, like B-complex, C, A, E, and K, supporting nutrition and biotechnology. Their vitamin content varies by species, environment, and cultivation conditions.
| 1. Prevention of Lipid Peroxidation: Vitamin E prevents oxidation of polyunsaturated fatty acids absorbed from the diet. 2. ROS Scavenging and Lipoprotein Protection: Act as antioxidants, blocking the formation of reactive oxygen species (ROS) and preventing the oxidation of low-density lipoproteins. 3. Lipid Peroxidation Chain Termination: α-tocopherol donates its phenolic hydrogen atom to a peroxyl radical and converts it to a hydroperoxide. The tocopheroyl radical formed is sufficiently stable and cannot continue the chain. |
| PUFA’s | Algae contain polyunsaturated fatty acids (PUFAs), such as omega-3 and omega-6, essential for human health, promoting cardiovascular, anti-inflammatory, and neuroprotective functions.
| 1. Modulation of ROS Production: Eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) can influence the expression of antioxidant enzymes, such as superoxide dismutase and catalase, which help neutralize free radicals and other ROS. 2. Inhibition of Lipid Peroxidation: Omega-3 fatty acids can be incorporated into cell membranes, where they reduce the susceptibility of lipids to peroxidation, a chain reaction that damages cell membranes by attacking lipids 3. Modulation of the Inflammatory Response: Reducing the production of pro-inflammatory mediators and increasing the production of anti-inflammatory mediators. 4. Interaction with Cellular-Signaling Proteins: Interact with proteins like NF-κB, regulating inflammation and oxidative stress gene expression. 5. Improvement of Mitochondrial Function: Omega-3s enhance mitochondrial function, reducing ROS production and protecting against oxidative damage. |
| Extraction Method | Description | Advantages | Challenges/Limitations | Example Applications/Studies | References |
|---|---|---|---|---|---|
| Enzyme-Assisted Extraction | Uses enzymes to break down cell wall components, releasing intracellular compounds. Increases surface area for solvent contact, improving yield and reducing extraction times. | High specificity; Selective targeting of molecules; High yields; Reduced extraction time. | High enzyme cost; Variability in seaweed composition; Stability of enzymes; Potential contamination; Environmental concerns. | Porphyra yezoensis, Ulva pertusa, Laminaria japonica, Callymenia perforata—Increased protein content after enzymatic treatment. Sargassum boveanum—Effective enzyme combinations for antioxidant and antimicrobial activities. | [141,142,143,144,145,146,147,148] |
| Microwave-Assisted Extraction | Microwaves selectively heat polar molecules, increasing diffusion of bioactive compounds into the solvent. Reduces solvent consumption and extraction time. | Faster extraction; Less solvent needed; Environmentally friendly; Selective heating of polar molecules. | High equipment cost; Need for precise temperature control; Degradation of thermolabile compounds. | Ulva spp.—Significant phytochemical profile change. Sargassum algae—Efficient extraction of higher molecular weight alginates. Caulerpa racemose—Higher caulerpin recovery compared to traditional methods. | [149,150,151,152,153,154] |
| Ultrasound-Assisted Extraction | High-frequency sound waves cause cavitation, disrupting the cell walls and increasing surface area for solvent contact. | Higher yields than traditional methods; Efficient extraction of high molecular weight compounds; Improved antioxidant and antimicrobial activities. | Potential for equipment damage; Need for optimization of extraction conditions; Limited research on large-scale application. | Ascophyllum nodosum—Enhanced phenolic, fucose, and uronic acid extraction. Sargassum wightii—Increased antioxidant activity and higher yields of phenolics and flavonoids. | [155,156,157,158,159,160,161,162,163] |
| Irradiation-Assisted Extraction | Ionizing radiation (e.g., gamma rays) breaks chemical bonds, creating free radicals that increase material permeability, facilitating solvent penetration and compound extraction. | Increases permeability; Can be used for complex matrices; Efficient for extracting bioactive compounds from algae. | Need for precise radiation control; Potential safety and regulatory issues; Degradation of compounds if not properly controlled. | Polysiphonia abscissa—Enhanced antioxidant activity after gamma-ray irradiation. Laminarin—Degradation into lower molecular weight fragments that enhance biological activities (antioxidant, antimicrobial). | [164,165,166,167] |
| Supercritical Fluid Extraction | Uses supercritical fluids (like CO2) to extract bioactive compounds | - High efficiency and selectivity - Low environmental impact - Selective extraction with minimal co-extraction of impurities | - High initial investment cost - Solubility limitations - Requires understanding of thermodynamic principles | Extraction of antioxidative compounds from brown seaweed (Alaria esculenta, Laminaria digitata, Ascophyllum nodosum). Recovery of fucosterol from Lessonia vadosa | [168,169,170,171,172,173] |
| Pressurized Liquid Extraction | Utilizes elevated temperatures and pressures to enhance solvent solvating power | - Reduced extraction time - Enhanced selectivity - Lower solvent consumption | - Higher levels of some compounds may not be recovered - Requires optimization of temperature, pressure, and solvents | Extraction of carotenoids from Neochloris oleoabundans with antiproliferative activity. Phenolic compounds from Sargassum muticum. | [174,175,176,177] |
| Deep Eutectic Solvents | Mixture of hydrogen bond donor and acceptor molecules with lower melting points | - Biodegradable - Tunable polarity - Low volatility - Green solvent alternative | - High viscosity hindering mass transfer - Challenges with scaling up processes - Need for further research on solvent design | Extraction of astaxanthin from Haematococcus pluvialis. Phlorotannin extraction from Fucus vesiculosus. | [178,179,180,181] |
| Pulsed Electric Field | High-intensity electric pulses cause electroporation, releasing intracellular compounds | - Non-thermal extraction - Enhanced extraction yields - Can improve product purity | - High equipment costs - Difficult scaling for industrial use - Need for careful optimization of treatment parameters | Extraction of phenolic compounds and carbohydrates from Alaria esculenta. Carotenoids from Tetraselmis chuii. Phycocyanin from Arthrospira platensis. | [182,183,184,185,186,187,188] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monroy-García, I.N.; Torres-Romero, S.; Castro-Ochoa, L.D.; Mendoza-Acosta, A.; Viveros-Valdez, E.; Ayala-Zavala, F. Bioactive Compounds from Marine Macroalgae: A Natural Defense Against Oxidative Stress-Related Diseases. Stresses 2025, 5, 22. https://doi.org/10.3390/stresses5010022
Monroy-García IN, Torres-Romero S, Castro-Ochoa LD, Mendoza-Acosta A, Viveros-Valdez E, Ayala-Zavala F. Bioactive Compounds from Marine Macroalgae: A Natural Defense Against Oxidative Stress-Related Diseases. Stresses. 2025; 5(1):22. https://doi.org/10.3390/stresses5010022
Chicago/Turabian StyleMonroy-García, Imelda Noehmi, Suleyka Torres-Romero, Lelie Denise Castro-Ochoa, Alexander Mendoza-Acosta, Ezequiel Viveros-Valdez, and Fernando Ayala-Zavala. 2025. "Bioactive Compounds from Marine Macroalgae: A Natural Defense Against Oxidative Stress-Related Diseases" Stresses 5, no. 1: 22. https://doi.org/10.3390/stresses5010022
APA StyleMonroy-García, I. N., Torres-Romero, S., Castro-Ochoa, L. D., Mendoza-Acosta, A., Viveros-Valdez, E., & Ayala-Zavala, F. (2025). Bioactive Compounds from Marine Macroalgae: A Natural Defense Against Oxidative Stress-Related Diseases. Stresses, 5(1), 22. https://doi.org/10.3390/stresses5010022