Abstract
In January 2020, a mortality event was documented in endangered Pallid Sturgeon (Scaphirhynchus albus) at the Neosho National Fish Hatchery (Neosho, MO, USA). Moribund fish appeared emaciated, with hemorrhages on the fins and rostrums. Mortality steadily increased, prompting an investigation into the cause. No viral or bacterial pathogens were detected; however, a high number of the novel Gyrodactylus sp. was identified on the fins and body surface of the affected fish. Treatment of the parasites with a formalin product was effective, and mortality decreased. Herein, we describe the novel species Gyrodactylus conei using a combination of morphological and molecular data. Due to the listing of the host as an endangered species, the concept of parasite conservation is also discussed.
1. Introduction
Pallid Sturgeon (Scaphirhynchus albus) is a large, ancient riverine species native to the Missouri River and lower half of the Mississippi River, preferring turbid waters with a current [1]. Habitat loss and degradation, river flow modification, pollution, and overharvest contributed to population decline, leading to the species being listed as endangered in 1990 ([2]). Extensive recovery efforts by federal and state partners have helped restore some natural reproduction in the lower Missouri River and ultimately hope to recover self-sustaining populations in reaches with suitable spawning habitats. For recovery in parts of the Missouri and the lower Mississippi Rivers, captive broods are propagated at Gavin’s Point National Fish Hatchery (NFH) (Yankton, SD, USA) and Neosho NFH (Neosho, MO, USA), with the resulting progeny grown to fingerling size prior to stocking. Careful management of these fingerling lots is required to produce healthy fish with representative genetic diversity of parent stocks for recovery in the wild.
Gyrodactylus is one of the most speciose genera of Platyhelminth parasites. It is estimated that there may be more than 20,000 species, with only a small fraction having been described [3]. Gyrodactylus has never been reported on sturgeons in North America, with the only reports of monogeneans involving three other genera (i.e., Nitzschia, Diclybothrium, and Paradiclybothrium) [4]. The only report of a Gyrodactylus infecting any sturgeon comes from a low-level infection involving an unidentified species on pond-reared Atlantic Sturgeon (Acipenser oxyrinchus oxyrinchus) (Mitchill 1815) in Poland [5].
Outbreaks related to Gyrodactylus are relatively rare, with the most studied clinical cases being associated with Gyrodactylus salaris infecting Atlantic Salmon (Salmo salar) in Europe [6]. In North America, low-level infections are commonly observed on hatchery fish and typically appear to be more of a stress-inducing nuisance than an outright pathogen [7,8]. However, in this study, elevated levels of mortality coincided with high numbers of Gyrodactylus infesting Pallid Sturgeon (Scaphirhynchus albus) at the Neosho National Fish Hatchery (Neosho, MO, USA). The mortality event culminated in the successful treatment of the ectoparasitic infection with formalin, which significantly reduced mortality.
2. Results
2.1. Diagnosis of Mortality Event
The evaluation of Moribund Sturgeon at the La Crosse Fish Health Center revealed a significant Gyrodactylus infestation, with the parasites primarily being observed on the fins. A total of 425 Gyrodactylus (left pectoral fin: 44; right pectoral fin: 96; left pelvic fin: 52; right pelvic fin: 60; caudal fin: 55; surrounding water: 118) were counted from one fish and 487 from another (left pectoral fin: 81; right pectoral fin: 30; right pelvic fin: 51; left pelvic fin: 31; caudal fin: 47; surrounding water: 247). No apparent bacterial or viral pathogens were identified. The high level of parasitism observed by Gyrodactylus on the Pallid Sturgeon appeared to a factor in the disease outbreak.
The histologic examination of a fish found changes only in the skin and branchial chamber. Few monogenes were evident, as most had fallen off during fixation/processing. The epidermis was variably irregular and mildly hyperplastic, with 6–10 cell layers and occasional mucus cells. The surface layer was eroded and ragged, with frequent intraepidermal macrophages that contained melanin granules, hypereosinophilic cells, and karyorrhectic debris (necrosis) (Figure 1A,B). Few eosinophilic granulocytes were also present in areas of erosion. The epidermal cells had hydropic changes with spongiosis and increased numbers of intercellular lymphocytes.
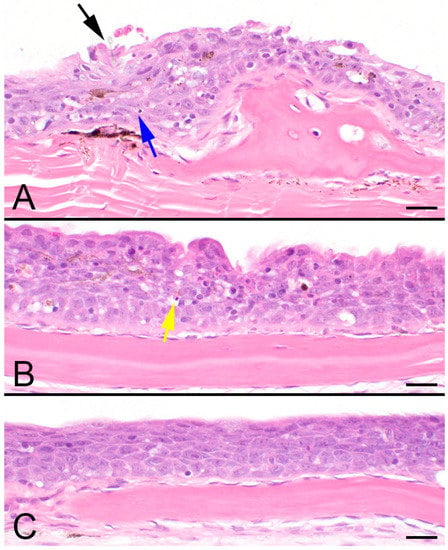
Figure 1.
Photomicrograph Pallid Sturgeon (Scaphirhynchus albus) skin. (A) The surface is irregular due to an epithelial injury overlying a dermal scale. A fragment of a monogenean hooklet (black arrow) is faintly visible. Epithelial cells with clear vacuoles both within (hydropic swelling) and between cells (spongiosis), as well as karyorrhectic debris (blue arrow), are evident. (B) The epidermis is hyperplastic, and the surface is irregular. There is increased hydropic swelling and spongiosis, with pyknotic shrunken pink cells (yellow arrow, necrosis). Lymphocytes and macrophages containing pigments are also present. (C) Normal epithelium. Scale bar is 20 μm. 600×, H&E.
2.2. Treatment
The outcome of the treatment was successful, and after the second treatment, clinical signs of external infestation by Gyrodactylus spp. were not observed. Due to the infrequent and rarely documented (if ever) use of Parasite-S on Pallid Sturgeon, the 100 ppm dosage was selected at the low range of the approved concentration for drug administration (up to 250 ppm). However, there was some potential toxicity to the treatment, as a slight increase in mortality occurred on treatment days (Figure 2). It is important to note that some fish did not return to feed after the treatments and later died. It was estimated that this outcome was caused by the associated stress of the formalin treatment rather than the parasite infestation. It is important to note that, due to the value of these fish, we did not sacrifice additional fish to reevaluate the parasite infestation but assumed the treatment was effective due to the improvement in the condition of the fish, their behavior, and decreased mortality. No recurring infestations have been observed since.
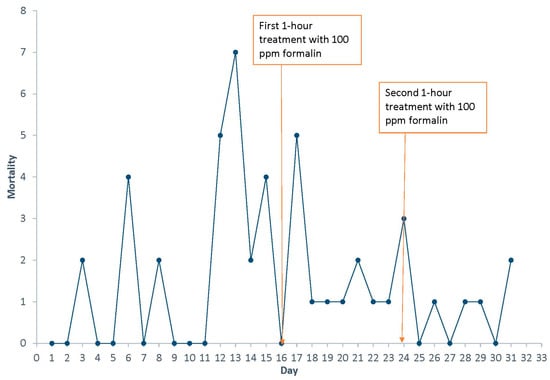
Figure 2.
Mortality of Scaphirhynchus albus during an outbreak associated with Gyrodactylus conei n. sp. Days where treatments of Parasite-S were administered are also shown.
2.3. New Species Description
Gyrodactylidae Cobbold, 1864
Gyrodactylus von Nordmann, 1832
Gyrodactylus conei n. sp.
Measurements were taken from permanently mounted (n = 9) specimens, as well as fresh specimens (n = 3), in temporary wetmounts [bracketed]. Parasites consistent with the genus Gyrodactylus were identified (Figure 3). The worms were 394 ± 44 long and 99 ± 13 wide. Prohaptor-containing glands are in each lobe. MCO present near but posterior to the pharynx. MCO are round with one large spine and seven smaller spines arranged in a single row (Figure 3d). Ovaries are round and anterior to the testes. Intestinal crura are blind and terminate near the testes. Circular haptor with stout anchors feature a straight shaft and relatively thick points (Figure 3a,b). The anchors are 55.0 ± 2.4 [60.5 ± 0.7] in length, with a 14.3 ± 2.1 [17.0 ± 1.8] root, 42.9 ± 1.8 [45.6 ± 0.8] shaft, and 23.1 ± 2.8 [26.6 ± 1.8] point. The dorsal bar is thin but consistent medially, with round posterior projections on each end. The ventral bar is 25.0 ± 2.2 [27.4 ± 0.8] in transverse length and 6.6 ± 0.4 [6.5 ± 0.4] in longitudinal width, with a 16.8 ± 1.9 [17.3 ± 0.4] membrane. The anterolateral processes are 10.4 ± 1.6 [9.2 ± 1.1] long. Marginal hooks (Figure 3c), featuring a robust sickle with a rounded heel with a pronounced shelf have a total length of 31.4 ± 1.4 [32.2 ± 0.8], with a 5.1 ± 0.4 [5.1 ± 0.3] long sickle and 26.2 ± 1.1 [27.1 ± 0.9] handle.
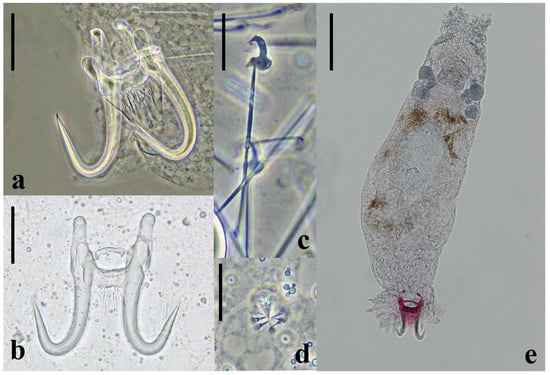
Figure 3.
Taxonomically important features of Gyrodactylus conei n. sp. (a) Anchors and ventral bar (phase contrast). (b) Anchors and ventral bar (Brightfield). (c) Marginal hook (phase contrast). (d) Male copulatory organ (phase contrast). (e) Whole body stained in Gomori’s trichrome and mounted in Canada balsam.
Type Host: Pallid Sturgeon (Scaphirhynchus albus).
Type Location: Neosho National Fish Hatchery; Neosho, MO, USA.
Site of Infection: Fins.
Type Material: Slides of the specimen types were deposited in the National Museum of Natural History (Smithsonian Institute, Washington, D.C., USA): holotypes (USNM 1606618) and paratypes (USNM 1606619).
Zoobank Registration: urn:lsid:zoobank.org:pub:E93106C1-CDE4-47D9-A0B8-9BC192F04A0E.
Etymology: The species name is in honor of Dr. David Cone, retired professor from St. Mary’s University (Nova Scotia, Canada), in honor of his work studying the parasites of fish, notably Gyrodactylids, as well as in thanks for the mentoring he has provided author EML over the years.
Molecular data: Partial SSU rRNA gene 460 bp (OR039423) and 906 bp of the ITS region (OR039424) were submitted to GenBank.
2.3.1. Molecular Identification
The partial SSU rRNA gene (460 bp) was identical to an unidentified Gyrodactylus sp. collected from a Golden Shiner (Notemigonus crysoleucas) farm in Minnesota (KT149284; e = 0) and Gyrodactylus colemanensis Mizelle and Kritsky, 1967 collected from Brook Trout (Salvelinus fontinalis) in Nova Scotia (JF836090.1; e-value = 0). All other BLAST results for the SSU rRNA gene were less than 97.50% similar. The sequence for the ITS region (906 bp) was 98.13% similar to Gyrodactylus colemanensis collected from Brook Trout in Nova Scotia (JF836142.1; e-value = 0) and 96.69% similar to the Gyrodactylus sp. collected from Golden Shiners at a farm in Minnesota (KT149288.1; e-value = 0). All other BLAST results for the ITS region were <89% similar.
2.3.2. Remarks
Both the morphological and molecular data collected from these parasites supported the description of the novel species. Gyrodactylus specimens collected from Pallid Sturgeon share a general morphological resemblance to G. colemanensis, which was described from the skin of steelhead Oncorynchus mykiss (syn: Salmo gairdneri) in a California hatchery [9]. However, these species can be differentiated based on the size of the sclerotized parts, especially the anchors and ventral bar, which were larger with the G. conei n. sp. compared to G. colemanensis (Table 1). Furthermore, examination of the holotype for G. colemanensis (USNM 1357413) revealed distinct differences among the taxonomically informative marginal hook sickles, as the heel of G. conei n. sp. displays a pronounced proximal rounding compared to the more angled heel with G. colemanensis. While the toe for both species was similarly shaped, this feature was markedly more abrupt with the G. conei n. sp. Differences in the molecular data (1.87%), which were scattered throughout the ITS region, further confirmed that G. conei was a novel species, as differences of 1% have been proposed to separate Gyrodactylus spp., although interspecific differences as low as 0.81% have been reported [10,11].

Table 1.
Comparison of the measurements (µm) of haptoral sclerotized parts for Gyrodactylus conei n. sp. and Gyrodactylus colemanensis. a-preserved specimens (wetmounts). b-reported in Mizelle and Kritsky, 1967. c-Measurements of the Holotype specimen for G. colemanensis (USNM 1357413). ALP = anterolateral processes.
The primary fish cultured, historically and currently, at the Neosho NFH has been Rainbow Trout (Oncorhynchus mykiss), which is a host of G. colemanensis and the closest genetic relative of G. conei n. sp. Initially, we considered that G. colemanensis could have colonized the new host in the hatchery; however, only G. salmonis has been identified from the trout at Neosho NFH (La Crosse Fish Health Center, unpublished data). Furthermore, the genetic change observed between G. colemanensis and G. conei was at a level nearly inconceivable to be produced in a hatchery setting over the course of the few months while the young sturgeon were being raised. For example, Leis et al. [7] found that G. magnus and G. salaris were separated by 1.95 My. While that length of time was calculated with data from mitochondrial DNA, when the ITS region sequences for those two species were compared a difference of 96 nucleotides was observed which suggests, assuming a consistent rate of molecular change, each nucleotide difference in the ITS could be associated with the passing of ~20,312 years. The difference of 17 nucleotides in the ITS between G. conei and G. colemanensis suggests a significant amount of time has passed since the initial host switching occurred, and the most likely route of infection for the young sturgeon is through wild caught, adult Pallid Sturgeon that, for spawning purposes, were brought into the facility where young sturgeon were reared. We expect that G. conei n. sp. would be found to infect wild Pallid Sturgeon in the Missouri River Basin. Furthermore, the native range for both fish species, as well as that of Golden Shiners, overlap in the Western United States [1] and illustrate that these three fish hosts are capable of inhabiting similar ecosystems within relative proximity to each other. It would seem likely that host switching among these hosts occurred sometime in the very distant past.
3. Discussion
We believe that Gyrodactylus conei n. sp. played some role in the mortality of Pallid Sturgeon that occurred at the Neosho NFH. No viruses or apparently pathogenic bacteria were detected, and the Gyrodactylus infection observed was exceptionally intense. Once treated, the mortality rate decreased. Furthermore, the tissue changes seen by light microscopy were consistent with other reports of monogenean infestations, the severity of which could explain the morbidity and death. Epithelial hyperplasia with reduced numbers of mucus cells, erosion, epithelial cell necrosis, and inflammation are typical of gyrodactylid infestations [12,13]. The fluid and electrolyte losses in the skin, coupled with the energy investment to produce mucus and repair the skin, would severely impair growth and predispose fish to bacterial infection. It is interesting to note that this level of infestation was not observed prior or since on Pallid Sturgeon at the Neosho NFH.
Gyrodactylus spp. tend to be quite host-specific [4]. However, host switching involving closely related host species or even unrelated hosts (i.e., prey to predator) has been documented [10]. While the colonization of a new host can lead to allopatric speciation, switching among closely related fish may not lead to a substantive genetic change, even among populations separated for significant periods of time (e.g., several thousand years [7]). The ability to host switch within the lineage associated with G. colemanensis and G. conei n. sp. appears exceptional. In addition to G. colemanensis, an unnamed but morphologically and molecularly similar parasite was identified from Golden Shiner at a baitfish farm in Minnesota [14]. All three of these species are known to infect the fins and skin of their hosts [9,14,15]. Given that this lineage has been able to colonize salmonids, leuciscids, and now asipenserids suggests this is a remarkable group capable of parasitizing incredibly diverse host fish and worthy of future study regarding host switching and the evolutionary changes that follow such an event.
Treatment with Parasite-S (formalin) at 100 ppm for one hour was effective in resolving the mortality associated with the heavy external infestation of Gyrodactylus conei n. sp., although caution should be exercised in administering this treatment. During this trial, some mortality, albeit very low numbers, could have been associated with the formalin treatment. An alternative treatment with 35% Perox-Aid® (hydrogen peroxide) may also be possible, as it has been shown to be effective for the treatment of Gyrodactylus spp. on multiple finfish species [8]. In the United States, participants in the Investigational New Animal Drug (INAD) Program may propose a treatment with 35% Perox-Aid® (hydrogen peroxide) under INAD #11-669 for fish species that are not already approved for labeled use under NADA 141-255 (salmonid species are approved for the treatment of Gyrodactylus spp. under NADA 141-255). Investigations to study both the efficacy and safety of Perox-Aid® (hydrogen peroxide) treatments for Gyrodactylus infestations of Pallid Sturgeon would provide valuable data that could support approval by the FDA and, if found safe and effective, would provide an important tool to improve the health of this species in the culture.
Pallid Sturgeon are listed as an endangered species, and possibly due to the status of the fish, no reports of parasite infections were found during our literature search, and none were listed in [4]. As such, G. conei n. sp. represents the only parasite described from this endangered fish, leading to an interesting question: Is G. conei also endangered? While some parasites are considered generalists that can infect a variety of hosts, species of Gyrodactylus tend to be host-specific or infect related hosts. The risk of extinction for this parasite would appear equal to, if not greater than, the risks to Pallid Sturgeon. Furthermore, if the G. conei n. sp. is endangered, should thought be given to its conservation? The concept of parasite conservation is an area that has garnered recent attention, and there are several examples of species that have gone extinct due to antiparasitic treatments during captive rearing of the host [16]. While, by definition, parasites harm the individual host in some manner, they actually contribute to properly functioning, biodiverse ecosystems through a variety of mechanisms [17,18]. Windsor [17] stated that parasites should be released back into the wild with the rehabilitated host, essentially stating that we should not intend to release parasite-sterile hosts back into their habitats. Although, in the case presented herein with Pallid Sturgeon, it was important to treat the parasitic infestation in order to save the hosts, highlighting how the conservation of parasites may involve an intricate balancing act of maintaining healthy hosts while keeping the parasites at sustainable levels in potentially densely reared captive populations. Overall, it seems important to keep in mind that each creature, no matter their size or trophic status, has a place in a properly functioning ecosystem. It seems prudent to at least consider parasite conservation during host restoration efforts, as lost biodiversity is impossible to regain.
4. Materials and Methods
4.1. Description of Mortality Event
A mortality event was observed in a production lot of 650 fingerling Pallid Sturgeon produced from captive brood stock at the Neosho NFH (Neosho, MO, USA) in January of 2020. Fish considered healthy would display relaxed swimming behavior, with the sturgeon being evenly distributed throughout the tank. However, during the outbreak, their behavior was altered, as they appeared lethargic and bunched up in groups. The water temperature was a constant 17 °C, dissolved oxygen 7.5 ppm, and a density of 5 fish/m2. The fish were an average length and weight of 195 mm and 4.95 g, respectively. Low levels of mortality were also observed, and a targeted sample of fish displaying clinical signs were sent to La Crosse Fish Health Center for diagnostic testing. The fish appeared emaciated, with marked erythema on the rostrum and pectoral fins (Figure 4). Some were moribund and pale, with excessive erosion of the skin and fin margins. A heavy infestation of Gyrodactylus spp. was noted, and the following treatment was administered under constant temperature with supplemental oxygen.
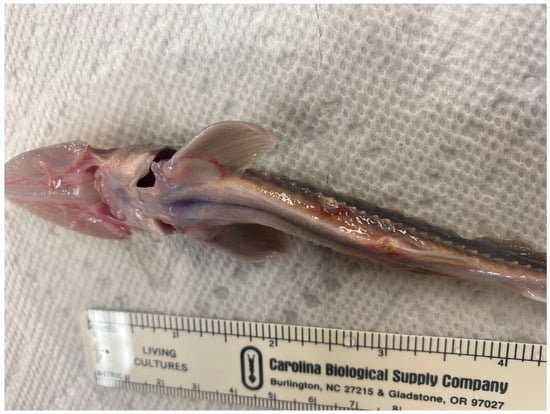
Figure 4.
Pallid Sturgeon showing signs of disease (e.g., emaciation and erythema) related to the outbreak of Gyrodactylus conei n. sp.
4.2. Treatment Protocol
A one-hour, standing bath treatment of 100 ppm Parasite-S (formalin) was conducted according to the label instructions for the control of external monogenean (Gyrodactylus spp.) parasites (NADA# 140-989). Mortality continued (up to two fish per day) posttreatment, and the treatment was repeated on day 12. After the second treatment, mortality was reduced to less than one fish per day, and the overall condition continued to improve. No further treatments were administered.
4.3. Bacteriology Methods
Bacteriology samples were taken from the kidney of moribund Pallid Sturgeon using a sterile 1 µL loop and streaked onto Tryptic soy agar (TSA). The samples were then processed (i.e., colonies picked, isolated, and biochemically assessed) according to the standard procedures for the detection of finfish bacterial pathogens outlined in the American Fisheries Society and USFWS Blue Book [19].
Kidney and spleen tissue were collected and screened for the presence of Renibacterium salmoninarum (R. sal) using the quantitative PCR methods described in [19].
4.4. Virology Methods
Moribund Sturgeon were screened for viruses as outlined in the AFS and USFWS Blue Book [19]. Briefly, kidney and spleen tissue were collected from moribund Pallid Sturgeon and placed in HBSS. The samples were then homogenized, diluted 1:1, and plated onto Chinook Salmon embryo (incubated at 15 °C; CHSE-214 [20], epithelioma papulosum cyprini (EPC) cells (incubated at 15 °C and 20 °C [21] and White Sturgeon skin cells (incubated at 20 °C (WSSK-1 [22]). The cells were monitored twice a week for the development of a cytopathic effect (CPE).
Kidney and spleen tissue homogenates were also used to screen for the presence of the Missouri River Sturgeon Iridovirus (MRSIV) using conventional PCR methods. DNA was extracted using a Qiagen DNeasy Blood & Tissue Extraction kit, and the amplification methods were followed as described in [23].
4.5. Histology Methods
Whole diseased Pallid Sturgeon were fixed in Davidson’s Fixative for 48 h and then transferred to 70% ethanol. An entire fixed fish was decalcified in formic acid, dissected, and serially sectioned; multiple sections from all areas of the body, as well as viscera, were embedded in paraffin blocks, cut, stained with hematoxylin and eosin, and coverslipped for standard light microscopy.
4.6. Gyrodactylus Identification
Temporary mounts of Gyrodactylus specimens, cleared in sodium dodecyl sulfate (Wong et al. 2006), were used for morphological identification. Permanently mounted specimens were stained with Gomori’s trichrome [24], dehydrated in a series of ethanol, cleared in clove oil, and mounted in Canada balsam [25]. Measurements, in µm, were reported as described as in [26], except we measured the transverse ventral bar length. Individual worms were also placed in 95% ethanol for molecular identification using methods similar to [25]. Briefly, the DNA was individually extracted from four worms using a Qiagen DNeasy Blood and Tissue kit. PCR reactions, using primers PBS18SF and PBS18SR [27] to amplify the small subunit (SSU) rRNA gene, as well as P3b and P4 [28] to amplify the ITS1/5.8S/ITS2 region (ITS), were added to Platinum Taq DNA Polymerase High Fidelity (Thermo Fisher, Waltham, MA, USA) Master Mix in accordance with the manufacturer’s instructions. The thermocycler parameters were as follows: 95 °C for an initial 10 min denaturation and 40 repeating cycles of a denaturation at 95 °C for 1 min, annealing at 53 °C for 1 min, a 72 °C extension for 2 min, and a final 10 min extension at 72 °C. PCR products were Sanger-sequenced by Eton Biosciences, and the resulting chromatograms were examined and assembled with Geneious 2021.1.1 (www.geneious.com; accessed on 21 March 2022). The contiguous sequences were examined for similarities with other known sequences through BLAST searches in GenBank (https://www.ncbi.nlm.nih.gov/genbank/).
Author Contributions
Conceptualization, E.L., J.B., I.S., S.D., R.K., R.M., J.P., N.E. and W.B.; methodology, E.L., J.B., I.S., S.D., R.K., R.M., J.P., N.E. and W.B.; formal analysis, E.L., J.B., I.S., S.D., R.K., R.M. and W.B.; investigation, E.L., J.B., I.S., S.D., R.K., R.M., J.P., N.E. and W.B.; data curation, E.L., J.B., I.S., S.D., R.K., R.M. and W.B.; writing—original draft preparation, E.L., J.B. and W.B.; and writing—review and editing, E.L., J.B., I.S., S.D., R.K., R.M., J.P., N.E. and W.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the U.S. Fish and Wildlife Service.
Institutional Review Board Statement
Ethical review and approval were not required for this study, and fish were euthanized in accordance with the approved methods outlined in the Guidelines for the use of Fishes in Research established by the American Fisheries Society [29].
Data Availability Statement
Genetic data are available in GenBank (18S rRNA gene: OR039423; ITS region: OR039424).
Acknowledgments
We would like to thank John Fisher (National Conservation Training Library; Shepherdstown, WV, USA) for help obtaining the literature. Any mention of trade names does not imply endorsement by the United States Government. The findings and conclusions in this article are those of the author(s) and do not necessarily represent the views of the U.S. Government.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pflieger, W.L. The Fishes of Missouri; Missouri Department of Conservation: Jefferson City, MO, USA, 1975. [Google Scholar]
- US Fish and Wildlife Service. Endangered and Threatened Wildlife and Plants; Determination of Endangered Status for the Pallid Sturgeon. Fed. Regist. 1990, 55, 36641–36647. [Google Scholar]
- Bakke, T. Host Specificity Dynamics: Observations on Gyrodactylid Monogeneans. Int. J. Parasitol. 2002, 32, 281–308. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, G.L. Parasites of North American Freshwater Fish; Cornell University Press: Ithaca, NY, USA, 1999. [Google Scholar]
- Popielarczyk, R.; Kolman, R. Preliminary Analysis of Ectoparasites of the Sturgeon Acipenser Oxyrinchus Oxyrinchus (Mitchill, 1815) Originating from Different Water Habitats. Ann. Parasitol. 2013, 59, 139–141. [Google Scholar] [PubMed]
- Johnsen, B.O.; Jensen, A.J. The Gyrodactylus Story in Norway. Aquaculture 1991, 98, 289–302. [Google Scholar] [CrossRef]
- Leis, E.; Chi, T.K.; Lumme, J. Global Phylogeography of Salmonid Ectoparasites of the Genus Gyrodactylus, with an Emphasis on the Origin of the Circumpolar Gyrodactylus salmonis (Platyhelminthes: Monogenea). Comp. Parasitol. 2021, 88, 130–143. [Google Scholar] [CrossRef]
- Tuttle-Lau, M.; Leis, E.M.; Cupp, A.; Peterman, L.; Hebert, J.; Erickson, R.; Schleis, S.; Gaikowski, M. Efficacy of Hydrogen Peroxide to Reduce Gyrodactylus Species Infestation Density on Four Fish Species. J. Aquat. Anim. Health 2023. [Google Scholar] [CrossRef]
- Mizelle, J.D.; Kritsky, D.C. Studies on Monogenetic Trematodes. XXXIII. New Species of Gyrodactylus and a Key to the North American Species. Trans. Am. Microsc. Soc. 1967, 86, 390. [Google Scholar] [CrossRef]
- Zietara, M.S.; Lumme, J. Speciation by Host Switch and Adaptive Radiation in a Fish Parasite Genus Gyrodactylus (Monogenea, Gyrodactylidae). Evolution 2002, 56, 2445–2458. [Google Scholar] [CrossRef]
- Lumme, J.; Zietara, M.S.; Lebedeva, D. Ancient and Modern Genome Shuffling: Reticulate Mito-Nuclear Phylogeny of Four Related Allopatric Species of Gyrodactylus von Nordmann, 1832 (Monogenea: Gyrodactylidae), Ectoparasites on the Eurasian Minnow Phoxinus phoxinus (L.) (Cyprinidae). Syst. Parasitol. 2017, 94, 183–200. [Google Scholar] [CrossRef]
- Bruno, D.W.; Noguera, P.A.; Poppe, T.T. A Colour Atlas of Salmonid Diseases, 2nd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Teruo, M. Color Atlas of Fish Histopathology; Shin-Suisan Shinbun-Sha, Ltd.: Tokyo, Japan, 2007; Volume 2. [Google Scholar]
- Leis, E.; King, S.; Leis, S.; Cone, D. Infections of Gyrodactylus crysoleucas and Gyrodactylus sp. (Monogenea) at a Golden Shiner (Notemigonus crysoleucas) Farm in Minnesota. Comp. Parasitol. 2016, 83, 105–110. [Google Scholar] [CrossRef]
- Cone, D.K.; Beverley-Burton, M.; Wiles, M.; McDonald, T.E. The Taxonomy of Gyrodactylus (Monogenea) Parasitizing Certain Salmonid Fishes of North America, with a Description of Gyrodactylus nerkae n. Sp. Can. J. Zool. 1983, 61, 2587–2597. [Google Scholar] [CrossRef]
- Rózsa, L.; Vas, Z. Co-Extinct and Critically Co-Endangered Species of Parasitic Lice, and Conservation-Induced Extinction: Should Lice Be Reintroduced to Their Hosts? Oryx 2015, 49, 107–110. [Google Scholar] [CrossRef]
- Windsor, D.A. Conservation of Parasites. Int. J. Parasitol. Parasites Wildl. 2021, 14, 137. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Nichols, E. Neglected Wild Life: Parasitic Biodiversity as a Conservation Target. Int. J. Parasitol. Parasites Wildl. 2013, 2, 222–227. [Google Scholar] [CrossRef] [PubMed]
- U.S. Fish and Wildlife Service. American Fisheries Society-Fish Health Section. Standard Procedures for Aquatic Animal Health Inspections. In AFS-FHS. FHS Blue Book: Suggested Procedures for the Detection and Identification of Certain Finfish and Shellfish Pathogens, 2020 Edition. Available online: https://units.fisheries.org/fhs/fish-health-section-blue-book-2020/ (accessed on 25 May 2023).
- Lannan, C.N.; Winton, J.R.; Fryer, J.L. Fish Cell Lines: Establishment and Characterization of Nine Cell Lines from Salmonids. Vitro 1984, 20, 671–676. [Google Scholar] [CrossRef]
- Winton, J.; Batts, W.; DeKinkelin, P.; LeBerre, M.; Bremont, M.; Fijan, N. Current Lineages of the Epithelioma Papulosum Cyprini (EPC) Cell Line Are Contaminated with Fathead Minnow, Pimephales Promelas, Cells: Contamination of EPC Cells. J. Fish Dis. 2010, 33, 701–704. [Google Scholar] [CrossRef]
- Watson, L.; Yun, S.; Groff, J.; Hedrick, R. Characteristics and Pathogenicity of a Novel Herpesvirus Isolated from Adult and Subadult White Sturgeon Acipenser Transmontanus. Dis. Aquat. Org. 1995, 22, 199–210. [Google Scholar] [CrossRef]
- Kurobe, T.; Kwak, K.; MacConnell, E.; McDowell, T.; Mardones, F.; Hedrick, R. Development of PCR Assays to Detect Iridovirus Infections among Captive and Wild Populations of Missouri River Sturgeon. Dis. Aquat. Org. 2010, 93, 31–42. [Google Scholar] [CrossRef]
- Kritsky, D.C.; Leiby, P.D.; Kayton, R.J. A Rapid Stain Technique for the Haptoral Bars of Gyrodactylus Species (Monogenea). J. Parasitol. 1978, 64, 172–174. [Google Scholar] [CrossRef]
- Leis, E.; Easy, R.; Cone, D. A Variant of Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013 (Monogenea) Identified from Weed Shiner (Notropis texanus) in Wisconsin Portions of the Upper Mississippi River. Syst. Parasitol. 2021, 98, 247–253. [Google Scholar] [CrossRef]
- Malmberg, G. The Excretory Systems and the Marginal Hooks as a Basis for the Systematics of Gyrodactylus (Trematoda, Monogenea). Ark. Zool. 1970, 23, 1–235. [Google Scholar]
- Cone, D.; Abbott, C.; Gilmore, S.; Burt, M. A New Genus and Species of Gyrodactylid (Monogenea) from Silver Hake, Merluccius bilinearis, in the Bay of Fundy, New Brunswick, Canada. J. Parasitol. 2010, 96, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Cable, J.; Harris, P.D.; Tinsley, R.C.; Lazarus, C.M. Phylogenetic Analysis of Gyrodactylus spp. (Platyhelminthes: Monogenea) Using Ribosomal DNA Sequences. Can. J. Zool. 1999, 77, 1439–1449. [Google Scholar]
- Use of Fishes in Research Committee (Joint committee of the American Fisheries Society, the American Institute of Fishery Research Biologists, and the American Society of Ichthyologists and Herpetologists). Guidelines for the Use of Fishes in Research; American Fisheries Society: Bethesda, MD, USA, 2014. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).