

Immunoglobulin A and Physiologic Correlates of Well-Being in Asian Elephants

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals
2.2. Musth
2.3. Sample Collection
2.4. Hormone Immunoassays
2.5. Statistical Methods
3. Results
3.1. Descriptive Statistics
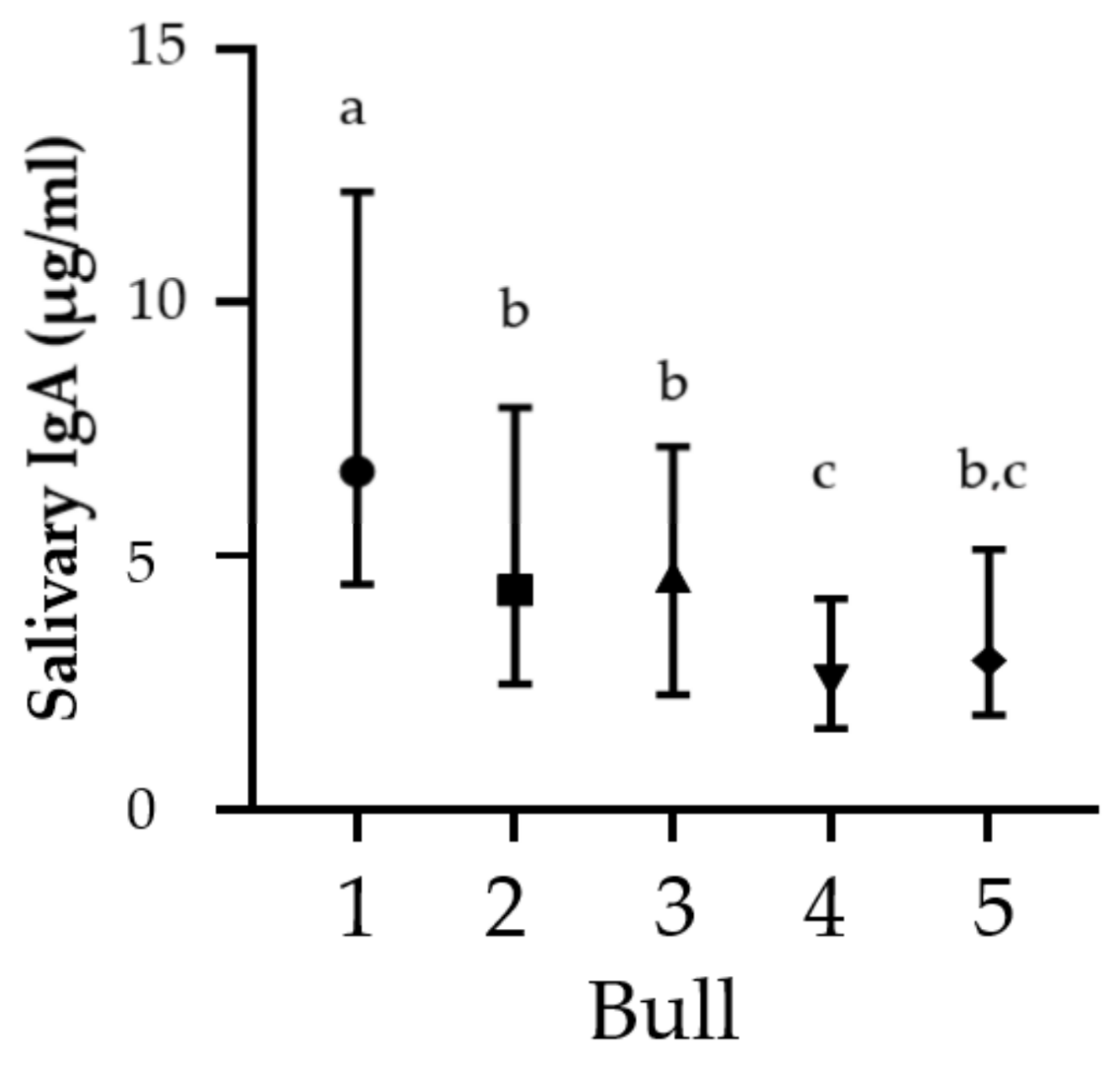
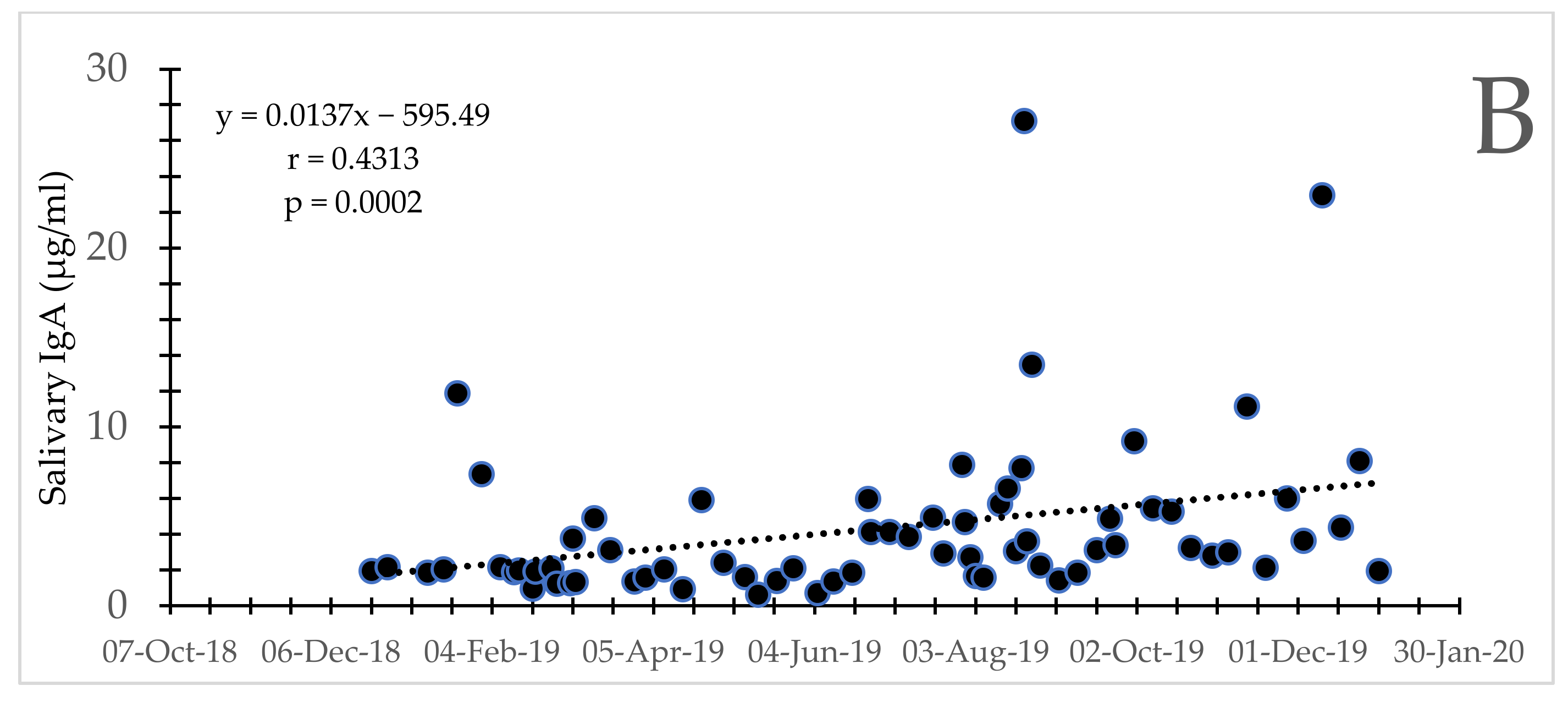
3.2. Secretory IgA Concentration
3.3. SIgA and Salivary Oxytocin
3.4. SIgA, Cortisol and Musth
3.5. SigA during Social and Solo Situations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacDougall, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “Stress” are not synonymous. Integr. Org. Biol. 2019, 1, obz017. [Google Scholar] [CrossRef]
- Dathe, H.H.; Kuckelkorn, B.; Minnemann, D. Salivary cortisol assessment for stress detection in the Asian elephant (Elephas maximus): A pilot study. Zoo Biol. 1992, 11, 285–289. [Google Scholar] [CrossRef]
- Palme, R.; Touma, C.; Arias, N.; Dominchin, M.F.; Lepschy, M. Steroid extraction: Get the best out of faecal samples. Wien Tierarztl Mon. 2013, 100, 238–246. [Google Scholar]
- Staley, M.; Conners, M.G.; Hall, K.; Miller, L.J. Linking stress and immunity: Immunoglobulin A as a non-invasive physiological biomarker in animal welfare studies. Horm. Behav. 2018, 102, 55–68. [Google Scholar] [CrossRef]
- Glaeser, S.S.; Edwards, K.L.; Wielebnowski, N.; Brown, J.L. Effects of physiological changes and social life events on adrenal glucocorticoid activity in female zoo-housed Asian elephants (Elephas maximus). PLoS ONE 2020, 15, e0241910. [Google Scholar] [CrossRef]
- Dalmau, A.; Ferret, A.; Chacon, G.; Manteca, X. Seasonal changes in fecal cortisol metabolites in Pyrenean chamois. J. Wildl. Manage. 2007, 71, 190–194. [Google Scholar] [CrossRef]
- Brown, J.L.; Lehnhardt, J. Serum and urinary hormones during pregnancy and the peri-and postpartum period in an Asian elephant (Elephas maximus). Zoo Biol. 1995, 14, 555–564. [Google Scholar] [CrossRef]
- Concannon, P.; Butler, W.; Hansel, W.; Knight, P.; Hamilton, J. Parturition and lactation in the bitch: Serum progesterone, cortisol and prolactin. Biol. Reprod. 1978, 19, 1113–1118. [Google Scholar] [CrossRef]
- Weingrill, T.; Gray, D.A.; Barrett, L.; Henzi, S.P. Fecal cortisol levels in free-ranging female chacma baboons: Relationship to dominance, reproductive state and environmental factors. Horm. Behav. 2004, 45, 259–269. [Google Scholar] [CrossRef]
- Brown, J.L.; Somerville, M.; Riddle, H.S.; Keele, M.; Duer, C.K.; Freeman, E.W. Comparative endocrinology of testicular, adrenal and thyroid function in captive Asian and African elephant bulls. Gen. Comp. Endocrinol. 2007, 151, 153–162. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Flávio, H.; Kristensen, M.L.; Walton-Rabideau, S.; Cooke, S.J.; Willmore, W.G.; Koed, A.; Aarestrup, K. Cortisol predicts migration timing and success in both Atlantic salmon and sea trout kelts. Sci. Rep. 2019, 9, 2422. [Google Scholar] [CrossRef]
- Edwards, K.L.; Miller, M.A.; Carlstead, K.; Brown, J.L. Relationships between housing and management factors and clinical health events in elephants in North American zoos. PLoS ONE 2019, 14, e0217774. [Google Scholar]
- Chrousos, G.P. Stressors, stress, and neuroendocrine integration of the adaptive response: The 1997 Hans Selye Memorial Lecture. Ann. N. Y. Acad. Sci. 1998, 851, 311–335. [Google Scholar]
- Edes, A.N.; Edwards, K.L.; Wolfe, B.A.; Brown, J.L.; Crews, D.E. Allostatic load indices with cholesterol and triglycerides predict disease and mortality risk in zoo-housed Western Lowland Gorillas (Gorilla gorilla gorilla). Biomark. Insights 2020, 15, 1177271920914585. [Google Scholar] [CrossRef]
- Edes, A.N.; Wolfe, B.A.; Crews, D.E. Evaluating allostatic load: A new approach to measuring long-term stress in wildlife. J. Zoo Wildl. Med. 2018, 49, 272–282. [Google Scholar] [CrossRef]
- Bansiddhi, P.; Brown, J.L.; Khonmee, J.; Norkaew, T.; Nganvongpanit, K.; Punyapornwithaya, V.; Angkawanish, T.; Somgird, C.; Thitaram, C. Management factors affecting adrenal glucocorticoid activity of tourist camp elephants in Thailand and implications for elephant welfare. PLoS ONE 2019, 14, e0221537. [Google Scholar] [CrossRef]
- Vanitha, V.; Thiyagesan, K.; Baskaran, N. Social life of captive Asian elephants (Elephas maximus) in Southern India: Implications for elephant welfare. J. Appl. Anim. Welf. Sci. 2011, 14, 42–58. [Google Scholar] [CrossRef]
- Evans, K.E.; Harris, S. Adolescence in male African elephants, Loxodonta africana, and the importance of sociality. Anim. Behav. 2008, 76, 779–787. [Google Scholar] [CrossRef]
- Lee, P.C.; Moss, C.J. African elephant play, competence and social complexity. Anim. Behav. Cogn. 2014, 1, 144–156. [Google Scholar] [CrossRef]
- De Silva, S.; Wittemyer, G. A comparison of social organization in Asian elephants and African savannah elephants. Int. J. Primatol. 2012, 33, 1125–1141. [Google Scholar] [CrossRef]
- Joshi, R. Tusker’s social bonds in Rajaji. Hystrix 2015, 26, 41–45. [Google Scholar] [CrossRef]
- Srinivasaiah, N.; Kumar, V.; Vaidyanathan, S.; Sukumar, R.; Sinha, A. All-male groups in Asian elephants: A novel, adaptive social strategy in increasingly anthropogenic landscapes of Southern India. Sci. Rep. 2019, 9, 8678. [Google Scholar]
- Rees, P.A. The sizes of elephant groups in zoos: Implications for elephant welfare. J. Appl. Anim. Welf. Sci. 2009, 12, 44–60. [Google Scholar] [CrossRef]
- Saragusty, J.; Hermes, R.; Göritz, F.; Schmitt, D.L.; Hildebrandt, T.B. Skewed birth sex ratio and premature mortality in elephants. Anim. Reprod. Sci. 2009, 115, 247–254. [Google Scholar] [CrossRef]
- Prado-Oviedo, N.A.; Bonaparte-Saller, M.K.; Malloy, E.J.; Meehan, C.L.; Mench, J.A.; Carlstead, K.; Brown, J.L. Evaluation of demographics and social life events of Asian (Elephas maximus) and African elephants (Loxodonta africana) in North American zoos. PLoS ONE 2016, 11, e0154750. [Google Scholar] [CrossRef]
- AZA. African Elephant Population Analysis and Breeding and Transfer Plan; Association of Zoos and Aquariums: Silver Spring, MD, 2020. [Google Scholar]
- AZA. Asian Elephant Population Analysis and Breeding and Transfer Plan; Association of Zoos and Aquariums: Silver Spring, MD, 2020. [Google Scholar]
- Ganswindt, A.; Rasmussen, H.B.; Heistermann, M.; Hodges, J.K. The sexually active states of free-ranging male African elephants (Loxodonta africana): Defining musth and non-musth using endocrinology, physical signals, and behavior. Horm. Behav. 2005, 47, 83–91. [Google Scholar] [CrossRef]
- Hartley, M.; Wood, A.; Yon, L. Facilitating the social behaviour of bull elephants in zoos. Int. Zoo Yearb. 2019, 53, 62–77. [Google Scholar] [CrossRef]
- Poole, J.H. Announcing intent: The aggressive state of musth in African elephants. Anim. Behav. 1989, 37, 140–152. [Google Scholar] [CrossRef]
- Readyhough, T.; Joseph, S.; Davis, M.; Moresco, A.; Schreier, A.L. Impacts of socialization on bull Asian elephants (Elephas maximus) stereotypical behavior. J. Zool. Bot. Gard. 2022, 3, 113–130. [Google Scholar] [CrossRef]
- Schreier, A.L.; Readyhough, T.S.; Moresco, A.; Davis, M.; Joseph, S. Social dynamics of a newly integrated bachelor group of Asian elephants (Elephas maximus). J. Appl. Anim. Welf. Sci. 2021, 1–18. [Google Scholar] [CrossRef]
- Schmid, J.; Heistermann, M.; Ganslosser, U.; Hodges, J. Introduction of foreign female Asian elephants (Elephas maximus) into an existing group: Behavioural reactions and changes in cortisol levels. Anim. Welf. 2001, 10, 357–372. [Google Scholar]
- Laws, N.; Ganswindt, A.; Heistermann, M.; Harris, M.; Harris, S.; Sherwin, C. A case study: Fecal corticosteroid and behavior as indicators of welfare during relocation of an Asian elephant. J. Appl. Anim. Welf. Sci. 2007, 10, 349–358. [Google Scholar] [CrossRef]
- Hansen, I.S.; Baeten, D.L.; den Dunnen, J. The inflammatory function of human IgA. Cell. Mol. Life Sci. 2019, 76, 1041–1055. [Google Scholar]
- Skandakumar, S.; Stodulski, G.; Hau, J. Salivary IgA: A possible stress marker in dogs. Anim. Welf. 1995, 4, 339–350. [Google Scholar] [CrossRef]
- Kosaruk, W.; Brown, J.L.; Plangsangmas, T.; Towiboon, P.; Punyapornwithaya, V.; Silva-Fletcher, A.; Thitaram, C.; Khonmee, J.; Edwards, K.L.; Somgird, C. Effect of tourist activities on fecal and salivary glucocorticoids and Immunoglobulin A in female captive Asian elephants in Thailand. Animals 2020, 10, 1928. [Google Scholar] [CrossRef]
- Plangsangmas, T.; Brown, J.L.; Thitaram, C.; Silva-Fletcher, A.; Edwards, K.L.; Punyapornwithaya, V.; Towiboon, P.; Somgird, C. Circadian Rhythm of Salivary Immunoglobulin A and Associations with Cortisol as A Stress Biomarker in Captive Asian Elephants (Elephas maximus). Animals 2020, 10, 157. [Google Scholar] [CrossRef]
- Richard, P.; Moos, F.; Freund-Mercier, M. Central effects of oxytocin. Physiol. Rev. 1991, 71, 331–370. [Google Scholar] [CrossRef]
- Tabak, J.; Gonzalez-Iglesias, A.E.; Toporikova, N.; Bertram, R.; Freeman, M.E. Variations in the response of pituitary lactotrophs to oxytocin during the rat estrous cycle. Endocrinology 2010, 151, 1806–1813. [Google Scholar] [CrossRef][Green Version]
- Prado, N.A.; Keady, M.; Oestmann, A.; Steinbeiser, C.M.; Brown, J.L. Hyperprolactinemic African elephant (Loxodonta africana) females exhibit elevated dopamine, oxytocin and serotonin concentrations compared to normal cycling and noncycling, low prolactin elephants. Biol. Reprod. 2019, 100, 1549–1560. [Google Scholar] [CrossRef]
- Brown, J.L.; Paris, S.; Prado-Oviedo, N.A.; Meehan, C.L.; Hogan, J.N.; Morfeld, K.A.; Carlstead, K. Reproductive health assessment of female elephants in North American zoos and association of husbandry practices with reproductive dysfunction in African elephants (Loxodonta africana). PLoS ONE 2016, 11, e0145673. [Google Scholar] [CrossRef]
- Ganswindt, A.; Heistermann, M.; Hodges, K. Physical, physiological, and behavioral correlates of musth in captive African elephants (Loxodonta africana). Physiol. Biochem. Zool. 2005, 78, 505–514. [Google Scholar] [CrossRef]
- Edwards, K.L.; Bansiddhi, P.; Paris, S.; Galloway, M.; Brown, J.L. The development of an immunoassay to measure immunoglobulin A in Asian elephant feces, saliva, urine and serum as a potential biomarker of well-being. Conserv. Physiol. 2019, 7, coy077. [Google Scholar] [CrossRef]
- Glaeser, S.S.; Edwards, K.L.; Paris, S.; Scarlata, C.; Lee, B.; Wielebnowski, N.; Finnell, S.; Somgird, C.; Brown, J.L. Characterization of longitudinal testosterone, cortisol, and musth in male Asian elephants (Elephas maximus), eEffects of aging, and adrenal responses to social changes and health events. Animals 2022, 12, 1332. [Google Scholar] [CrossRef]
- Bechert, U.; Hixon, S.; Schmitt, D. Diurnal variation in serum concentrations of cortisol in captive African (Loxodonta africana) and Asian (Elephas maximus) elephants. Zoo Biol. 2021, 40, 458–471. [Google Scholar]
- Guhad, F.; Hau, J. Salivary IgA as a marker of social stress in rats. Neurosci. Lett. 1996, 216, 137–140. [Google Scholar] [CrossRef]
- Lantz, E.L.; Lonsdorf, E.V.; Heintz, M.R.; Murray, C.M.; Lipende, I.; Travis, D.A.; Santymire, R.M. Non-invasive quantification of immunoglobulin A in chimpanzees (Pan troglodytes schweinfurthii) at Gombe National Park, Tanzania. Am. J. Primatol. 2018, 1, e22558. [Google Scholar] [CrossRef]
- Palm, A.-K.E.; Wattle, O.; Lundström, T.; Wattrang, E. Secretory immunoglobulin A and immunoglobulin G in horse saliva. Vet. Immunol. Immunopathol. 2016, 180, 59–65. [Google Scholar] [CrossRef]
- Bielsky, I.F.; Young, L.J. Oxytocin, vasopressin, and social recognition in mammals. Peptides 2004, 25, 1565–1574. [Google Scholar] [CrossRef]
- Smith, A.S.; Ågmo, A.; Birnie, A.K.; French, J.A. Manipulation of the oxytocin system alters social behavior and attraction in pair-bonding primates, Callithrix penicillata. Horm. Behav. 2010, 57, 255–262. [Google Scholar] [CrossRef]
- Veissier, I.; Boissy, A. Stress and welfare: Two complementary concepts that are intrinsically related to the animal’s point of view. Physiol. Behav. 2007, 92, 429–433. [Google Scholar] [CrossRef]
- Romero, L.M. Physiological stress in ecology: Lessons from biomedical research. Trends Ecol. Evol. 2004, 19, 249–255. [Google Scholar]
- Zeier, H.; Brauchli, P.; Joller-Jemelka, H.I. Effects of work demands on immunoglobulin A and cortisol in air traffic controllers. Biol. Psychol. 1996, 42, 413–423. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bull | Age at the Start (yr) | Time in Musth During Study (%) | Before Introductions 6 August 2018 to 11 February 2019 | During Introductions 12 February 2019 to 15 July 2019 | After Introductions 6 August 2019 to 21 December 2019 | |||
|---|---|---|---|---|---|---|---|---|
| Serum | Saliva | Serum | Saliva | Serum | Saliva | |||
| 1 | 49 | 21% | 20 | 28 | 22 | 23 | 13 | 12 |
| 2 | 14 | 39% | 27 | 28 | 21 | 19 | 19 | 14 |
| 3 | 11 | 20% | 30 | 32 | 24 | 27 | 22 | 27 |
| 4 | 10 | 0% * | 20 | 8 | 11 | 28 | 10 | 28 |
| 5 | 9 | 0% * | 16 | 7 | 4 | 29 | 16 | 27 |
| Total samples | 113 | 103 | 126 | 126 | 108 | 108 | ||
| Secretory IgA (μg/mL) | Salivary Oxytocin (pg/mL) | Cortisol (ng/mL) | |
|---|---|---|---|
| n | 355 | 356 | 282 |
| Median | 4.10 | 60.44 | 1.00 |
| Range | 0–27.11 | 2.71–279.90 | 0–26.36 |
| Interquartile range | 2.20–6.80 | 43.65–85.38 | 0.41–1.97 |
| Secretory IgA (μg/mL) | Salivary Oxytocin (pg/mL) | Cortisol (ng/mL) | Social † | |
|---|---|---|---|---|
| Secretory IgA (μg/mL) | 1.00 | |||
| Salivary oxytocin (pg/mL) | 0.28 *** | 1.00 | ||
| Cortisol (ng/mL) | 0.01 | −015 * | 1.00 | |
| Social † | −0.22 *** | −0.13 * | −0.11 | 1.00 |
| Secretory IgA (μg/mL) | Salivary Oxytocin (pg/mL) | Cortisol (ng/mL) | Testosterone (ng/mL) | |
|---|---|---|---|---|
| Secretory IgA (μg/mL) | 1.00 | |||
| Salivary oxytocin (pg/mL) | 0.28 *** | 1.00 | ||
| Cortisol (ng/mL) | 0.01 | −0.15 * | 1.00 | |
| Testosterone (ng/mL) | 0.15 * | −0.22 ** | 0.59 *** | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moresco, A.; Prado, N.; Davis, M.; Schreier, A.L.; Readyhough, T.S.; Joseph, S.; Gray, C.; Brown, J.L. Immunoglobulin A and Physiologic Correlates of Well-Being in Asian Elephants. J. Zool. Bot. Gard. 2022, 3, 677-687. https://doi.org/10.3390/jzbg3040050
Moresco A, Prado N, Davis M, Schreier AL, Readyhough TS, Joseph S, Gray C, Brown JL. Immunoglobulin A and Physiologic Correlates of Well-Being in Asian Elephants. Journal of Zoological and Botanical Gardens. 2022; 3(4):677-687. https://doi.org/10.3390/jzbg3040050
Chicago/Turabian StyleMoresco, Anneke, Natalia Prado, Maura Davis, Amy L. Schreier, Taylor S. Readyhough, Sharon Joseph, Charlie Gray, and Janine L. Brown. 2022. "Immunoglobulin A and Physiologic Correlates of Well-Being in Asian Elephants" Journal of Zoological and Botanical Gardens 3, no. 4: 677-687. https://doi.org/10.3390/jzbg3040050
APA StyleMoresco, A., Prado, N., Davis, M., Schreier, A. L., Readyhough, T. S., Joseph, S., Gray, C., & Brown, J. L. (2022). Immunoglobulin A and Physiologic Correlates of Well-Being in Asian Elephants. Journal of Zoological and Botanical Gardens, 3(4), 677-687. https://doi.org/10.3390/jzbg3040050