
Evaluating Environmental Enrichment Methods in Three Zoo-Housed Varanidae Lizard Species

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Individuals and Housing
2.2. Testing Protocol
2.3. Data Collection
2.4. Data Preparation
2.5. Statistical Analysis
2.5.1. Model 1: Effect of Enrichment on Exploratory Behaviour
2.5.2. Model 2: Effect of Enrichment Type on Engagement Time and Longevity
2.5.3. Model 3: Effect of Food- vs. Scent-Based Enrichment on Engagement Longevity
3. Results
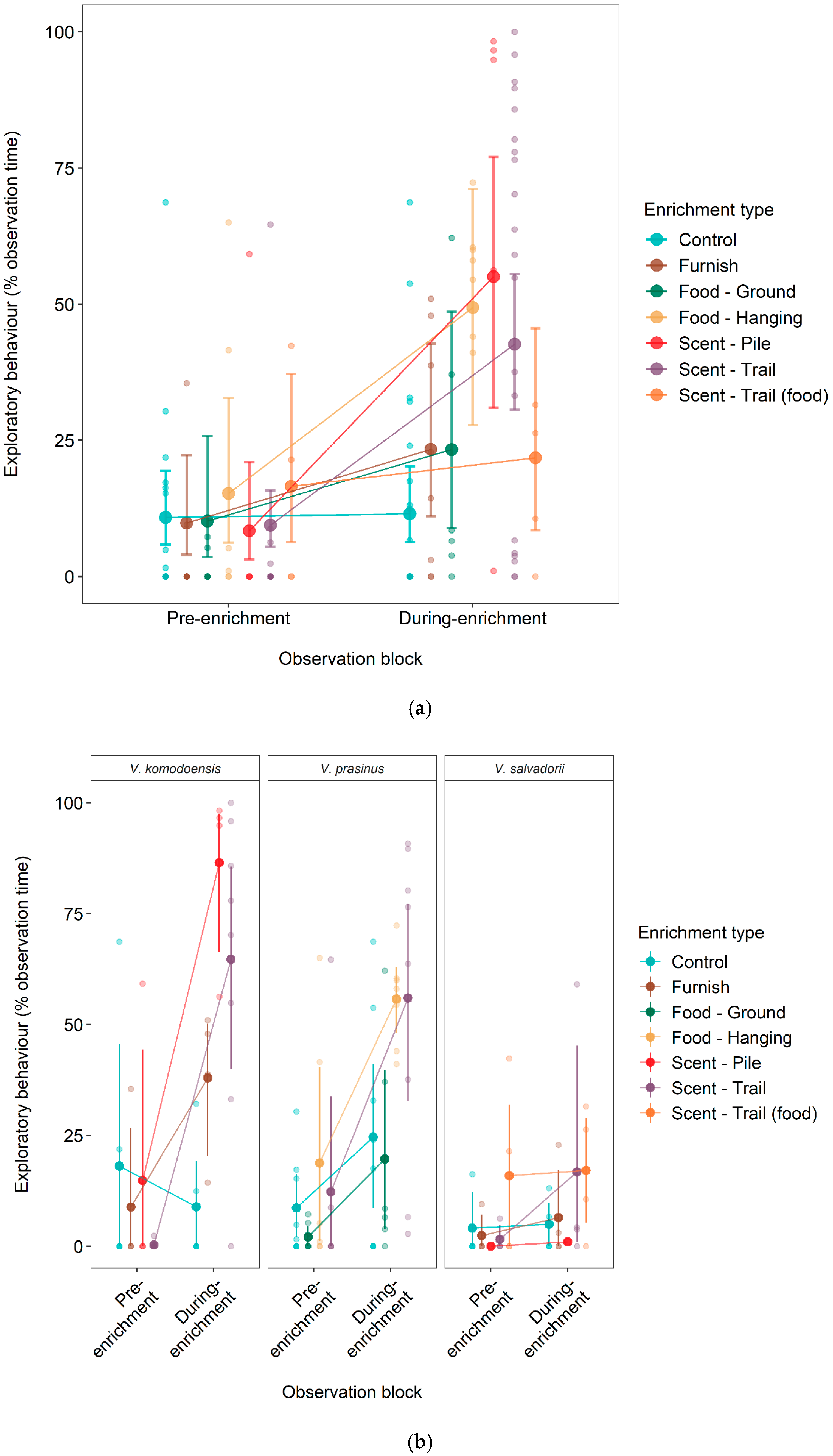
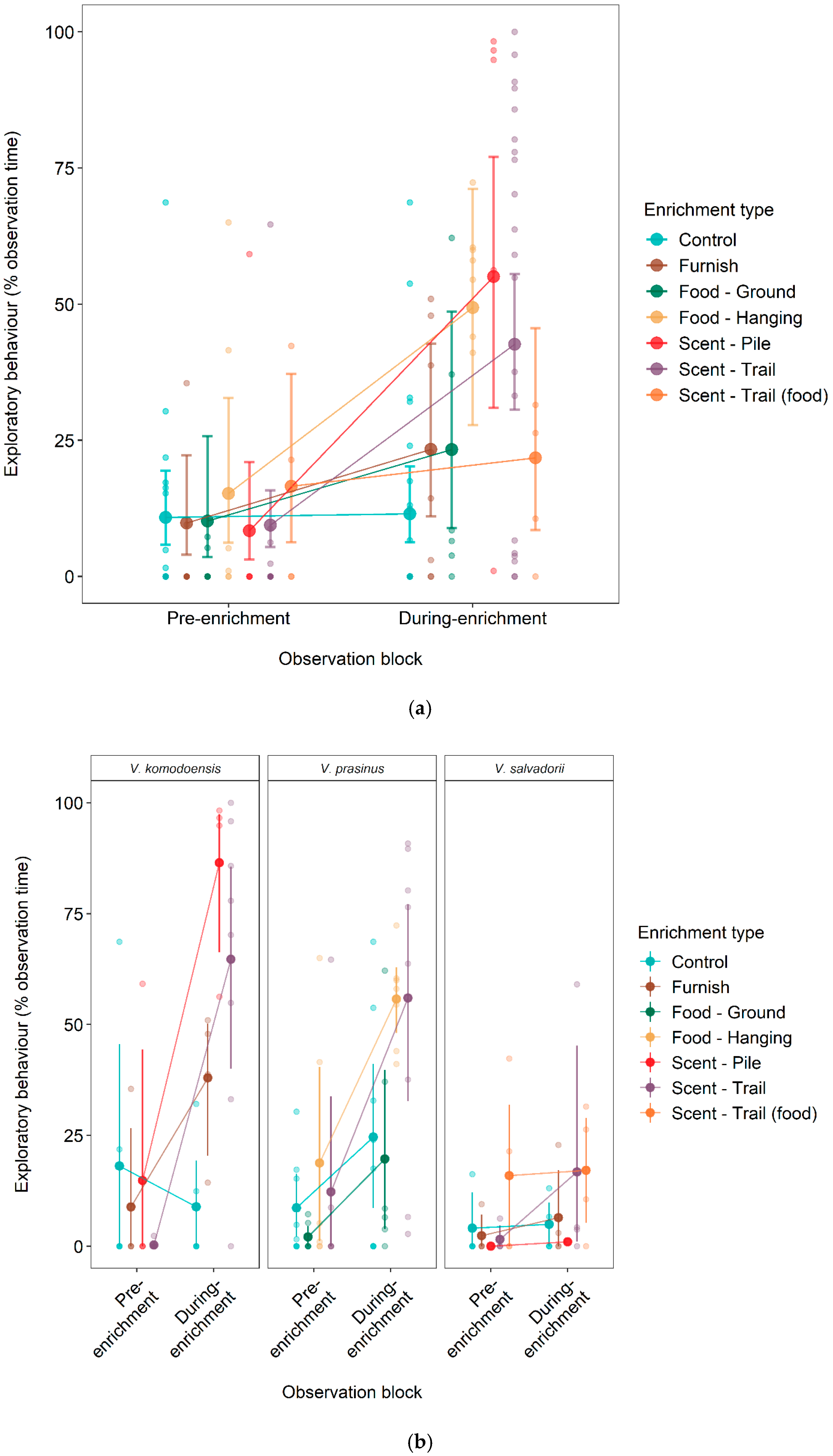
3.1. Effect of Enrichment on Exploratory Behaviour
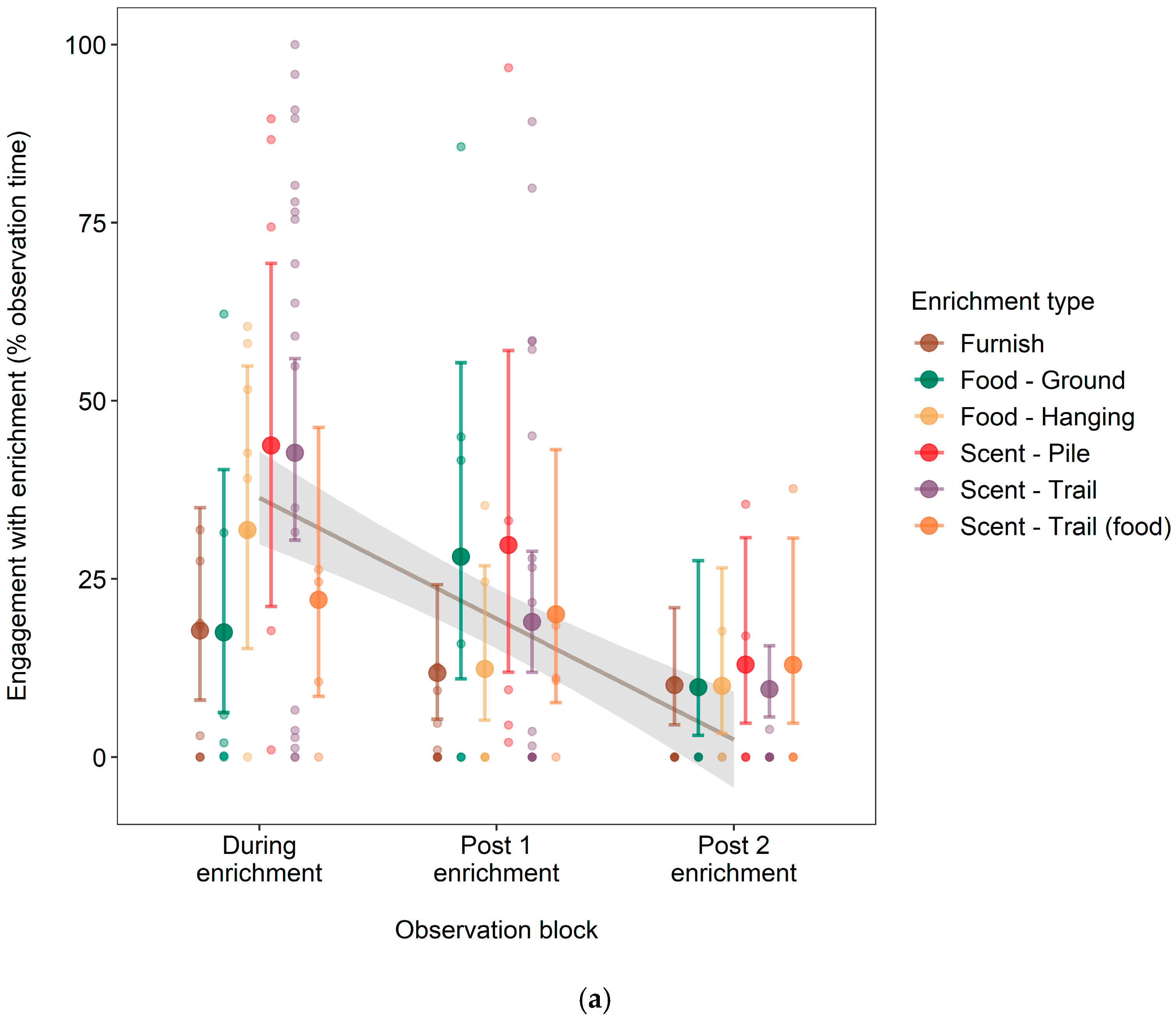
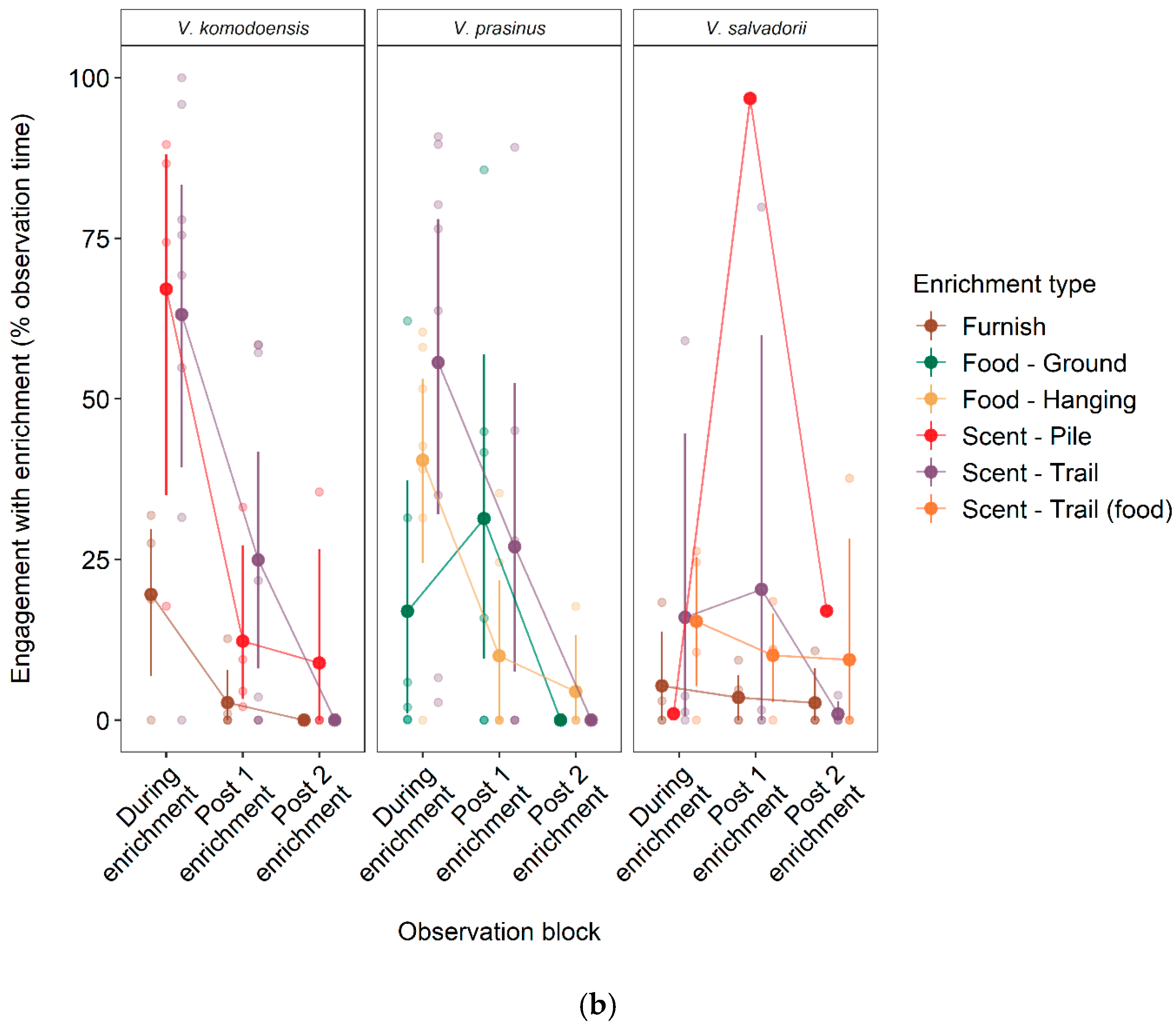
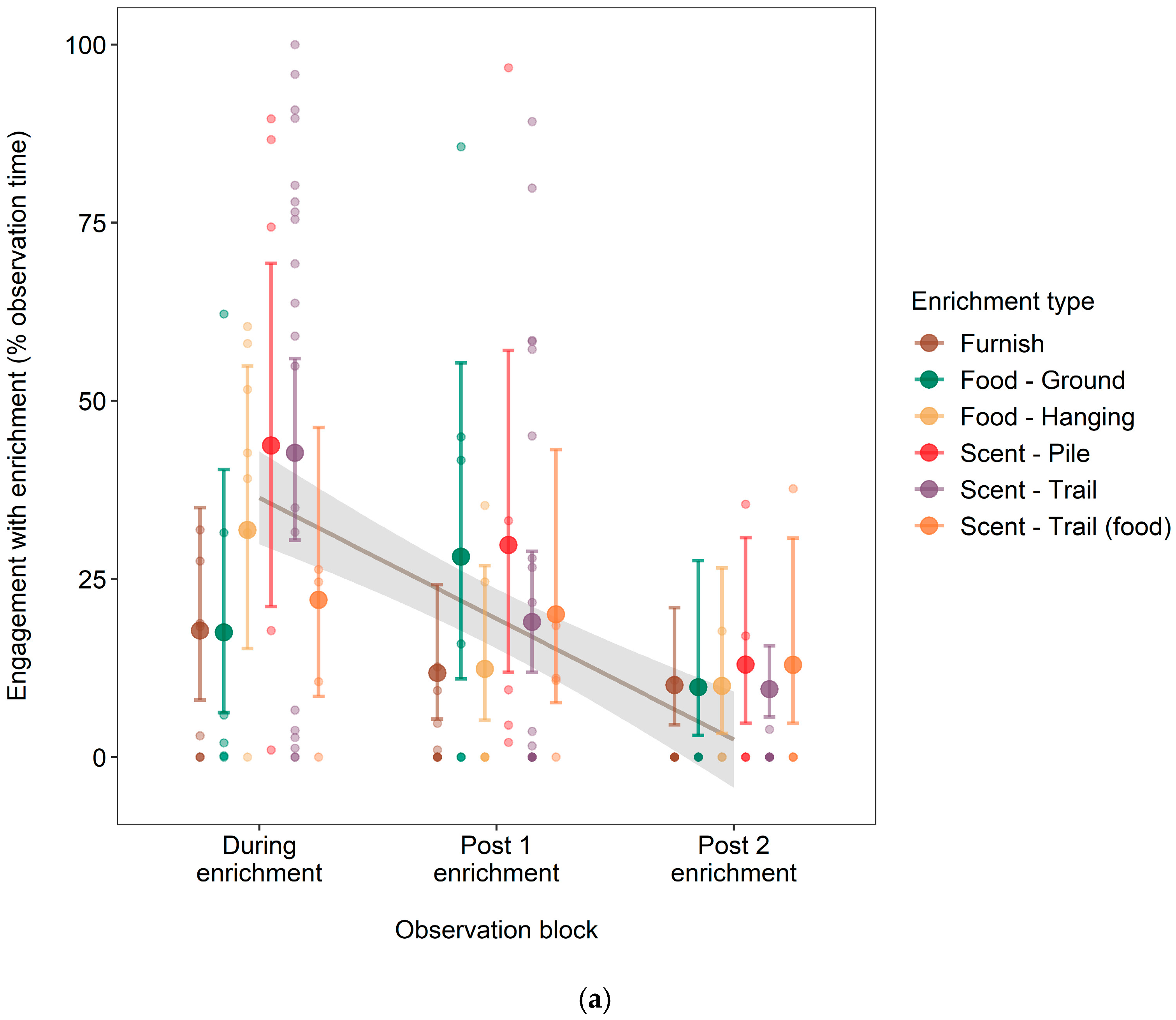
3.2. Effect of Enrichment Type on Engagement Time and Longevity
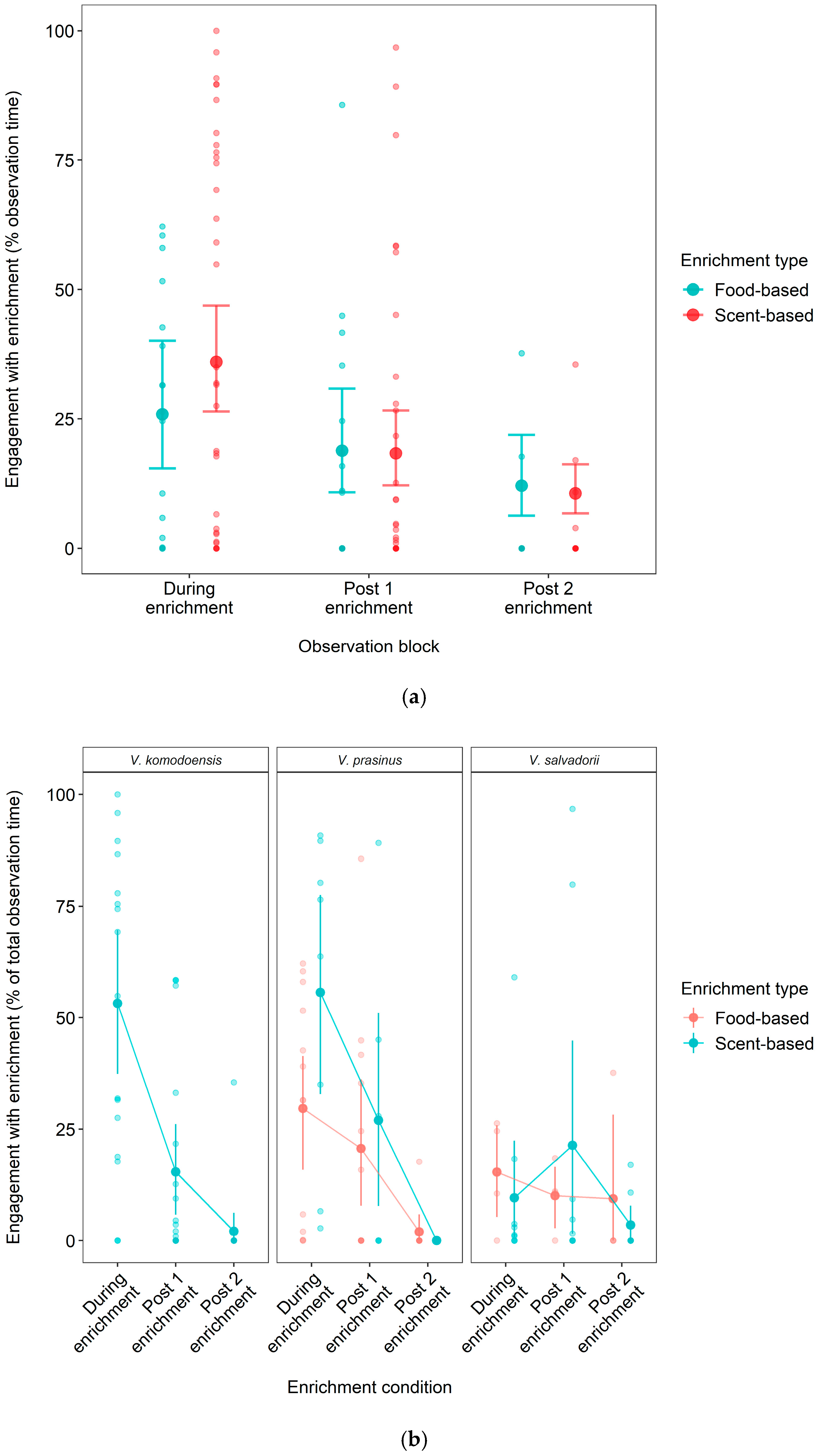
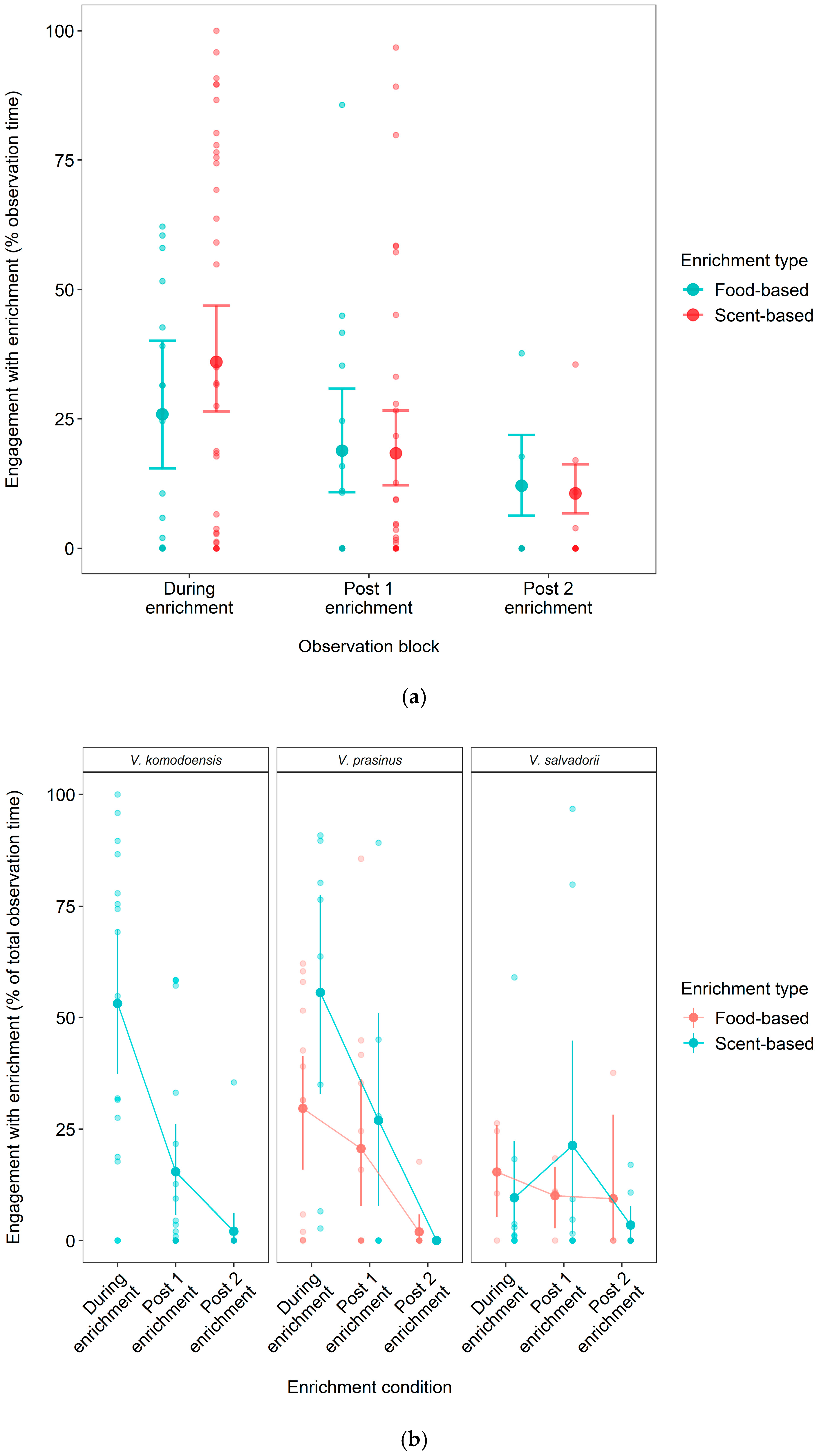
3.3. Effect of Food- vs. Scent-Based Enrichment on Engagement Longevity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bashaw, M.J.; Gibson, M.D.; Schowe, D.M.; Kucher, A.S. Does enrichment improve reptile welfare? Leopard geckos (Eublepharis macularius) respond to five types of environmental enrichment. Appl. Anim. Behav. Sci. 2016, 184, 150–160. [Google Scholar] [CrossRef]
- Arbuckle, K. Folklore husbandry and a philosophical model for the design of captive management regimes. Herpetol. Rev. 2013, 44, 448–452. [Google Scholar]
- Januszczak, I.S.; Bryant, Z.; Tapley, B.; Gill, I.; Harding, L.; Michaels, C.J. Is behavioural enrichment always a success? Comparing food presentation strategies in an insectivorous lizard (Plica plica). Appl. Anim. Behav. Sci. 2016, 183, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Horn, H.-G.; Visser, G.J. Review of reproduction of monitor lizards Varanus spp. in captivity II. Int. Zoo Yearb. 1997, 35, 227–246. [Google Scholar] [CrossRef]
- Horn, H.-G.; Visser, G.J. Review of reproduction of Monitor lizards Varanm spp. in captivity. Int. Zoo Yearb. 1989, 28, 140–150. [Google Scholar] [CrossRef]
- Mendyk, R.W. Life expectancy and longevity of varanid lizards (Reptilia:Squamata:Varanidae) in North American zoos. Zoo Biol. 2015, 34, 139–152. [Google Scholar] [CrossRef]
- Mendyk, R.W.; Newton, A.L.; Baumer, M. A Retrospective Study of Mortality in Varanid Lizards (Reptilia:Squamata:Varanidae) at the Bronx Zoo: Implications for Husbandry and Reproductive Management in Zoos. Zoo Biol. 2013, 32, 152–162. [Google Scholar] [CrossRef]
- Burghardt, G.M. Environmental enrichment and cognitive complexity in reptiles and amphibians: Concepts, review, and implications for captive populations. Appl. Anim. Behav. Sci. 2013, 147, 286–298. [Google Scholar] [CrossRef]
- Kuczaj, S.; Lacinak, T.; Fad, O.; Trone, M.; Solangi, M.; Ramos, J. Keeping environmental enrichment enriching. Int. J. Comp. Psychol. 2002, 15, 127–137. [Google Scholar]
- Tarou, L.R.; Bashaw, M.J. Maximizing the effectiveness of environmental enrichment: Suggestions from the experimental analysis of behavior. Appl. Anim. Behav. Sci. 2007, 102, 189–204. [Google Scholar] [CrossRef]
- Michaels, C.J.; Downie, J.R.; Campbell-Palmer, R. The importance of enrichment for advancing amphibian welfare and conservation goals: A review of a neglected topic. Amphib. Reptile Conserv. 2014, 8, 7–23. [Google Scholar]
- Rosier, R.L.; Langkilde, T. Does environmental enrichment really matter? A case study using the eastern fence lizard, Sceloporus undulatus. Appl. Anim. Behav. Sci. 2011, 131, 71–76. [Google Scholar] [CrossRef]
- Almli, L.M.; Burghardt, G.M. Environmental Enrichment Alters the Behavioral Profile of Ratsnakes (Elaphe). J. Appl. Anim. Welf. Sci. 2006, 9, 85–109. [Google Scholar] [CrossRef]
- Burghardt, G.M.; Ward, B.; Rosscoe, R. Problem of reptile play: Environmental enrichment and play behavior in a captive Nile soft-shelled turtle, Trionyx triunguis. Zoo Biol. 1996, 15, 223–238. [Google Scholar] [CrossRef]
- Doody, J.; Burghardt, G.; Dinets, V. Breaking the Social-Non-social Dichotomy: A Role for Reptiles in Vertebrate Social Behavior Research? Ethology 2013, 119, 95–103. [Google Scholar] [CrossRef]
- Manrod, J.; Hartdegen, R.; Burghardt, G. Rapid solving of a problem apparatus by juvenile black-throated monitor lizards (Varanus albigularis albigularis). Anim. Cogn. 2008, 11, 267–273. [Google Scholar] [CrossRef]
- Wilkinson, S.L. Reptile wellness management. Vet. Clin. Exot. Anim. Pract. 2015, 18, 281–304. [Google Scholar] [CrossRef]
- Bennett, D. Monitor Lizards: Natural History, Biology & Husbandry; Chimaira Bucnhandelsgesellschaft: Frankfurt, Germany, 1998. [Google Scholar]
- McBrayer, L.D.; McBrayer, L.B.; Miles, D.B. (Eds.) Lizard Ecology; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Pianka, E.R.; Pianka, E.R.; Vitt, L.J. Lizards: Windows to the Evolution of Diversity; University of California Press: Berkeley, CA, USA, 2003; Volume 5. [Google Scholar]
- Hurme, K.; Gonzalez, K.; Halvorsen, M.; Foster, B.; Moore, D.; Chepko-Sade, B.D. Environmental Enrichment for Dendrobatid Frogs. J. Appl. Anim. Welf. Sci. 2003, 6, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Young, R.J. Environmental Enrichment for Captive Animals; John Wiley & Sons: Chichester, UK, 2013. [Google Scholar]
- Warwick, C.; Arena, P.; Lindley, S.; Jessop, M.; Steedman, C. Assessing reptile welfare using behavioural criteria. Practice 2013, 35, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, G.; Whittaker, M.; Coe, J. Prototyping Naturalistic Enrichment Features: A Case Study. In Proceedings of the Seventh International Conference on Environmental Enrichment, New York, NY, USA, 31 July–5 August 2005; p. 60. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smithson, M.; Verkuilen, J. A Better Lemon Squeezer? Maximum-Likelihood Regression with BetaDistributed Dependent Variables. Psychol. Methods 2006, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology With R; Springer: New York, NY, USA, 2009; Volume 574. [Google Scholar]
- Douma, J.C.; Weedon, J.T. Analysing continuous proportions in ecology and evolution: A practical introduction to beta and Dirichlet regression. Methods Ecol. Evol. 2019, 10, 1412–1430. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. Introduction to Biostatistics; Dover Publications: Mineola, NY, USA, 2009. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. 2021. Available online: r-project.org (accessed on 31 October 2021).
- Lenth, R.V.; Buerkner, P.; Herve, M.; Love, J.; Riebl, H.; Singmann, H. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. 2021. Available online: uni-muenster.de (accessed on 31 October 2021).
- Londoño, C.; Bartolomé, A.; Carazo, P.; Font, E. Chemosensory enrichment as a simple and effective way to improve the welfare of captive lizards. Ethology 2018, 124, 674–683. [Google Scholar] [CrossRef]
- Therrien, C.L.; Gaster, L.; Cunningham-Smith, P.; Manire, C.A. Experimental evaluation of environmental enrichment of sea turtles. Zoo Biol. 2007, 26, 407–416. [Google Scholar] [CrossRef]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits; Bradford Books: Cambridge, MA, USA, 2005. [Google Scholar]
- Murphy, J.B.; Ciofi, C.; de La Panouse, C.; Walsh, T. Komodo Dragons: Biology and Conservation; Smithsonian Institution: Washington, DC, USA, 2015. [Google Scholar]
- Camina, Á.; Salinas, N.; Cuevas, J. Husbandry and breeding of the crocodile monitor Varanus salvadorii Peters & Doria, 1878 in captivity. Biawak 2013, 7, 56–62. [Google Scholar]
- Mason, R.T. Reptilian pheromones. Biol. Reptil. 1992, 18, 114–228. [Google Scholar]
- Ord, T.J.; Martins, E.P. Tracing the origins of signal diversity in anole lizards: Phylogenetic approaches to inferring the evolution of complex behaviour. Anim. Behav. 2006, 71, 1411–1429. [Google Scholar] [CrossRef]
- Auffenberg, W. The Behavioral Ecology of the Komodo Monitor; University Press of Florida: Gainesville, FL, USA, 1981. [Google Scholar]
- Smith, K.K. Morphology and function of the tongue and hyoid apparatus in Varanus (varanidae, lacertilia). J. Morphol. 1986, 187, 261–287. [Google Scholar] [CrossRef] [PubMed]
- Cogger, H. Reptiles and Amphibians of Australia; AH and AW Reed: Dunedin, New Zealand, 1975. [Google Scholar]
- Ziegler, T.; Schmitz, A.; Koch, A.; Böhme, W. A review of the subgenus Euprepiosaurus of Varanus (Squamata: Varanidae): Morphological and molecular phylogeny, distribution and zoogeography, with an identification key for the members of the V. indicus and the V. prasinus species groups. Zootaxa 2007, 1472, e28. [Google Scholar] [CrossRef]
- Murphy, J.; Mendyk, R.; Miller, K.; Augustine, L. Tales of Monitor Lizard Tails and Other Perspectives. Herpetol. Rev. 2019, 50, 178–201. [Google Scholar]
- Gaalema, D. Visual Discrimination and Reversal Learning in Rough-Necked Monitor Lizards (Varanus rudicollis). J. Comp. Psychol. 2011, 125, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Pepin, D.J. Natural History of Monitor Lizards (Family Varanidae) with Evidence from Phylogeny, Ecology, Life History and Morphology; Washington University: St. Louis, MO, USA, 2011. [Google Scholar]
- Shyne, A. Meta-analytic review of the effects of enrichment on stereotypic behavior in zoo mammals. Zoo Biol. 2006, 25, 317–337. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enrichment Condition | Description | Species Sampled |
|---|---|---|
| Control | Keeper entered enclosure as per normal husbandry routine for two minutes. | V. komodoensis |
| V. prasinus | ||
| V. salvadorii | ||
| Furnishings | Bedding and enclosure furniture from four mammalian exhibits: Congo buffalo (Syncerus caffer nanus) bedding; red river hog (Potamochoerus porcus) browse logs; white-faced saki monkey (Pithecia pithecia) enclosure logs; mixed bedding from Bovidae species. | V. komodoensis |
| V. salvadorii | ||
| Food (ground) | Hollowed log feeders filled with black crickets (Gryllus sp.) on enclosure floor. | V. prasinus |
| Food (suspended) | Hollowed log feeders filled with black crickets (Gryllus sp.) suspended on enclosure furniture. | V. prasinus |
| Scent (trail—food) | Blended food items (quail eggs, quail feathers, chicken eggs) spread throughout the enclosure. Food items (quail meat, day-old chickens) were also placed along the scent trail. | V. salvadorii |
| Scent (trail) | Liquids spread throughout the enclosure (blood–water solution, fish defrosting water, blended pinkie mice, blended quail eggs and feathers). | V. komodoensis |
| V. prasinus | ||
| V. salvadorii | ||
| Scent (pile) | A blood–water solution spread throughout the exhibit (within leaf/litter, on logs, buried in substrate surface, as a frozen solution on ground). | V. komodoensis |
| Behaviour | Description |
|---|---|
| Bask | Individual stationary underneath a heat/UV lamp for a minimum of five seconds. |
| Rest | Individual stationary (not under a heat/UV lamp) for a minimum of five seconds. |
| Explore | Relaxed interest/awareness in proximate or novel objects, relaxed visual explorations. Calm chemical sampling of surrounding, e.g., smelling or tasting objects or air (tongue-flicking). Individual moves more than half a body length from its starting position. |
| Feed | Consumption of food items, including holding food in mouth, chewing and swallowing. Feeding was considered finished after swallowing had stopped. |
| Social | Touching, vocalising and/or signalling to a conspecific. |
| Enrichment engagement (interest) | Rapid chemical sampling of surrounding, e.g., smelling or tasting objects or air (tongue-flicking). Individual moves (more than half a body length from its starting position) directly towards and/or stares directly at enrichment item. |
| Enrichment engagement (use) | Direct manipulation of enrichment item, including attempts to reach the item and/or active following of scent trails. Where live food was presented, included chasing food items. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Waterman, J.O.; McNally, R.; Harrold, D.; Cook, M.; Garcia, G.; Fidgett, A.L.; Holmes, L. Evaluating Environmental Enrichment Methods in Three Zoo-Housed Varanidae Lizard Species. J. Zool. Bot. Gard. 2021, 2, 716-727. https://doi.org/10.3390/jzbg2040051
Waterman JO, McNally R, Harrold D, Cook M, Garcia G, Fidgett AL, Holmes L. Evaluating Environmental Enrichment Methods in Three Zoo-Housed Varanidae Lizard Species. Journal of Zoological and Botanical Gardens. 2021; 2(4):716-727. https://doi.org/10.3390/jzbg2040051
Chicago/Turabian StyleWaterman, James O., Rachel McNally, Daniel Harrold, Matthew Cook, Gerardo Garcia, Andrea L. Fidgett, and Lisa Holmes. 2021. "Evaluating Environmental Enrichment Methods in Three Zoo-Housed Varanidae Lizard Species" Journal of Zoological and Botanical Gardens 2, no. 4: 716-727. https://doi.org/10.3390/jzbg2040051
APA StyleWaterman, J. O., McNally, R., Harrold, D., Cook, M., Garcia, G., Fidgett, A. L., & Holmes, L. (2021). Evaluating Environmental Enrichment Methods in Three Zoo-Housed Varanidae Lizard Species. Journal of Zoological and Botanical Gardens, 2(4), 716-727. https://doi.org/10.3390/jzbg2040051