Abstract
The COVID-19 pandemic, caused by SARS-CoV-2, is a deadly disease affecting millions due to the non-availability of drugs and vaccines. The majority of COVID-19 drugs have been repurposed based on antiviral, immunomodulatory, and antibiotic potential. The pathogenesis and advanced complications with infection involve the immune-inflammatory cascade. Therefore, a therapeutic strategy could reduce infectivity, inflammation, and immune modulation. In recent years, modulating the endocannabinoid system, particularly activation of the cannabinoid type 2 (CB2) receptor is a promising therapeutic target for modulation of immune-inflammatory responses. JWH133, a selective, full functional agonist of the CB2 receptor, has been extensively studied for its potent anti-inflammatory, antiviral, and immunomodulatory properties. JWH133 modulates numerous signaling pathways and inhibits inflammatory mediators, including cytokines, chemokines, adhesion molecules, prostanoids, and eicosanoids. In this study, we propose that JWH133 could be a promising candidate for targeting infection, immunity, and inflammation in COVID-19, due to its pharmacological and molecular mechanisms in numerous preclinical efficacy and safety studies, along with its immunomodulatory, anti-inflammatory, organoprotective, and antiviral properties. Thus, JWH133 should be investigated in preclinical and clinical studies for its potential as an agent or adjuvant with other agents for its effect on viremia, infectivity, immune modulation, resolution of inflammation, reduction in severity, and progression of complications in COVID-19. JWH133 is devoid of psychotropic effects due to CB2 receptor selectivity, has negligible toxicity, good bioavailability and druggable properties, including pharmacokinetic and physicochemical effects. We believe that JWH133 could be a promising drug and may inspire further studies for an evidence-based approach against COVID-19.
1. Introduction
Coronavirus disease-2019 (COVID-19), a pandemic and public health emergency caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), is a deadly disease that is affecting millions of people all over the world because of the non-availability of specific drugs or vaccines [1]. Currently, numerous efforts are underway to discover and develop preventive and therapeutic agents for SARS-CoV-2 infections [2]. Despite availability of the vaccines for COVID-19, identifying candidate drugs which could be effective for therapeutic management of COVID-19 is crucial. The majority of the drugs used in COVID-19 have been repurposed based on antiviral, antibiotic, anti-inflammatory, or immunomodulatory activities [3]. Considering the emergence of COVID-19-related mortality, effective medications are needed to improve patient prognosis and to stem the spread of the virus [3]. Among the numerous therapeutic avenues to be explored, the endocannabinoid system (ECS), which physiologically regulates innate and adaptive immunity, inflammation, pain, and oxidative stress [4] represents an important strategy for therapeutic targeting of hyperimmune-inflammatory responses during COVID-19.
The ECS typically consists of two receptors, cannabinoid receptor type 1 (CB1R) and 2 (CB2R), their endogenous ligands (endocannabinoids) and metabolic enzymes, as well as nonclassical targets of cannabinoids (e.g., transient receptor potential (TRP) channels and peroxisome proliferator-activated receptors) that are major players in the immune system and control a wide variety of diseases involving immune-inflammatory states [5]. The ECS is one of the newest drug targets receiving attention and has an excellent reputation due to the emergence of many successful drugs in the clinic in the past few years [6,7,8]. In the ECS, the CB2R is a G-protein-coupled receptor (GPCR) that, upon activation, regulates immune responses and inflammatory pathways; therefore, CB2R agonists have received enormous interest for possible therapeutic applications owing to their beneficial immunomodulatory, anti-inflammatory, and antioxidant roles, with the absence of psychotropic effects attributable to CB1R activation [9,10].
To date, numerous cannabinoid ligands have been classified as classical, non-classical, aminoalkylindoles, and eicosanoids that have been synthesized. Among the numerous CB2R ligands, JWH133, which was first synthesized by Huffman et al. (2010), has received enormous attention in experimental studies investigating CB2R-dependent pharmacological mechanisms and therapeutic potential [11]. Since its synthesis, it has been shown to be one of the most studied CB2R full functional agonist that exhibits high affinity and approximately 200-fold more selectivity towards CB2R than CB1R. This emerging ligand shows a wide range of therapeutic effects, including cardioprotective, hepatoprotective, neuroprotective, nephroprotective, anticonvulsive, antipsychotic, anticancer, anti-oxidant, anti-inflammatory, immunomodulatory, and antiviral, mediating selective activation of CB2R mimicking as full agonist.
Since the emergence of COVID-19, several drugs, including remdesivir, lopinavir, ritonavir, interferon-β, ribavirin, chloroquine/hydroxychloroquine, azithromycin, tocilizumab, and ivermectin, have appeared as promising therapeutics for COVID-19 [12]. From a pharmacological perspective, these drugs have the potential to either block the virus from entering host cells or prevent viral replication, and attenuate hyperimmune and hyperinflammatory states to prevent the disease progression and complications [3]. The utilization of these drugs in COVID-19 is mostly empirical, based on clinical experience of their therapeutic benefits in the management of previous SARS, Middle East respiratory syndrome, and Ebola virus epidemics.
In principle, immune responses and the resultant inflammatory process are imperative for the abolition of viremia, but this may significantly influence pathogenesis of SARS-CoV-2 and contribute to the signs and symptoms of COVID-19 [13]. In SARS-CoV infections, the use of antiviral agents alone is insufficient to prevent a cytokine storm and related complications in critically ill patients because immune dysregulation with hyperinflammatory conditions lead to complications, worsening, and poor prognosis rather than control of viremia [14]. To reduce morbidity and mortality, it is important to repurpose old drugs and to identify novel agents capable of attenuating a cytokine storm [15].
Current research efforts are ongoing across pharmaceutical, biotechnological, and academic studies for the discovery of novel drugs, as well as vaccines for SARS-CoV-2 [15]. COVID-19 pathogenesis involves hyperfunctioning of immunoregulatory cells (B cells, natural killer (NK) cells, CD4+ T cells, and CD8+ T cells) in defensive responses to the pathogen [16]. Subsequently, a hyperimmune status induces hyperinflammatory conditions by overproduction and release of cytokines, termed a cytokine storm, which determines the intensity of symptoms, mortality rate, progression, and worsening of the disease, mainly the pulmonary system, and causes extrapulmonary complications and multi-organ failure [17]. COVID-19 pathogenesis and complications involve the immune-inflammatory cascade; therefore, the available approaches emphasize this cascade to reduce inflammation and immune modulation [13,16].
Considering the pharmacological effects, molecular mechanisms, and therapeutic potential of JWH133, we reasonably hypothesize that JWH133 could be useful in COVID-19 because of its notable immunomodulatory, anti-inflammatory, and antiviral properties. Recently, CB2R has been suggested as a potential therapeutic target for regulating the immune-inflammatory axis in COVID-19 [18]. Our proposition is to scientifically contemplate the therapeutic perspective and prospect of JWH133 on infection, immunity, and inflammation with a potential use in COVID-19 to curb severity and progression, as well as poor prognosis. In this article, we discuss the possible prophylactic and treatment mechanisms of JWH133 in COVID-19. Much of the information presented is based on data derived from previously published studies reporting the immunomodulatory, anti-inflammatory, and antimicrobial properties of JWH133.
The roles of cannabinoids are well explored for their antiviral, anti-inflammatory, and immunomodulatory properties [8,19,20] and gaining attention for their candidature for potential in COVID-19 [21]. It has become apparent that agents that have antiviral, anti-inflammatory and immunomodulatory properties altogether could be important in context of COVID-19 to target the trinity of infection, inflammation and immunity. Many of the compounds showed targeting of SARS-CoV-2 using bioinformatics tools, such as in silico analysis, molecular docking, or molecular farming to enhance the production of recombinant proteins including vaccines and antibodies [22,23]. To tackle SARS-CoV-2, the identification of viral protease appears a striking therapeutic target to limit the replication of SARS-CoV-2 and many of the compounds are being investigated for their potential to target replication by inhibiting viral components, such as Mpro (3CLpro), PLpro and spike proteins [22,23]. Identifying candidate compounds, that have selectivity against viral components and prevent viral entry, as well as improve immunity and attenuate inflammatory factors in host cells, could be more important in context to SARS-CoV-2 infections. In the present article scientifically contemplates the therapeutic prospects of JWH133 in SARS-CoV-2 infection.
In this review, we perform molecular docking studies on JWH133 for the viral and host targets and found that Mpro appear to be a one of the important targets, we also elaborated the potential of JWH133 in SARS-CoV-2 infection integrating with previous findings, particularly regarding its immunomodulatory, anti-inflammatory, and antiviral properties.
2. Molecular Docking of JWH133 for its Activity on Mpro
Molecular docking is a powerful technique used to check the binding orientation of ligand into the active site of the target protein. The crystal structure of SARS-CoV-2 main protease (SARS-CoV-2 Mpro was retrieved from Protein Data Bank (PDB—available at http://www.rcsb.org) using the PDB code: 6LU7 [24], (Berman et al., 2015). Dock Prep tool of UCSF Chimera program was used to prepare receptor molecule [25]. During preparation, binding ligand, hetatoms, and the solvents were removed while the hydrogen atoms were added to the structure. The structure of JWH133 was searched and retrieved from PubChem database (CID: 6918505) [26]. The ligand structure was prepared in chimera by adding hydrogen atoms and charges.
The ligand binding residues in the structure of SARS-CoV-2 Mpro was designated to dock JWH133 using the Autodock Vina chimera plugin [27]. The best-docked ligand pose was selected for further analysis. Energy minimization of the docked complex was performed in Chimera using the energy minimization program [25]. A protein-ligand complex was processed and optimized in the free maestro program to refine molecular interactions [28] (Schrödinger, 2018). The molecular 2D interaction image was also generated using the ligand-receptor interaction module of the maestro package (Schrödinger, 2018). The non-covalent interactions were calculated at cutoff radius of 2.50 Å.
Docking of JWH133 into the active site of SARS-CoV-2 Mpro generated several binding poses. The best binding pose with docking energy −6.0 Kcal/mol was selected for molar interaction analysis. Molecular interaction analysis results revealed that JWH133 formed hydrophobic contact with Cys44, Met49, Pro52, Tyr54, Phe140, Leu141, and Met 165 of the target protein (Figure 1). His41, Asn142, Ser144, His163, His164, His172, and Gln189 were involved in polar contacts with target protein (Figure 1). These residues are the key residues, which play an important role in ligand binding. In a recent study, the importance of similar binding pattern of doxycycline, minocycline, lopinavir, oseltamivir, and ritonavir with SARS-CoV-2 Mpro have been highlighted [23,29].
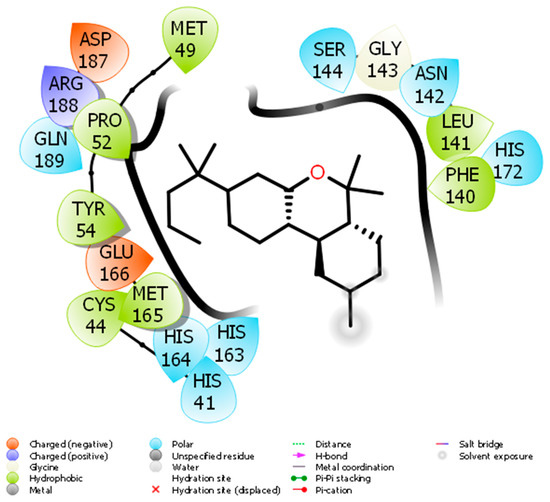
Figure 1.
The molecular interaction between SARS-CoV-2 Mpro and JWH133.
Based on the role of CB2R in immune-inflammatory mechanisms, the antiviral and agonist properties of JWH133 on CB2R, we hypothesized that JWH133 may be a potentially novel candidate to limit the severity and progression of COVID-19 by modulating infection, immunity, and inflammation. A scheme of the effect of JWH133 mediating CB2 receptor activation on the infection, inflammation, and immunity in context of SARS-CoV-2 is presented in Figure 2.
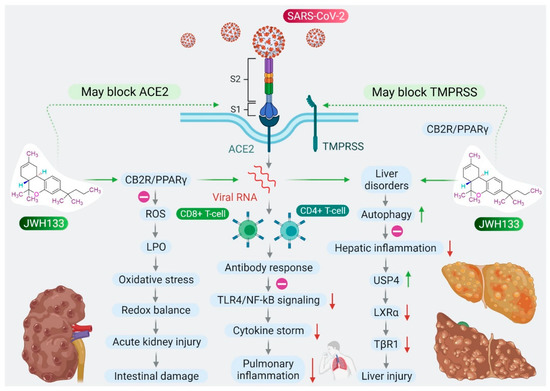
Figure 2.
Proposed scheme of potential and mechanisms of JWH133 on immunity, infection, and inflammation against SARS-CoV-2.
3. CB2 Receptors Mediated Anti-Inflammatory Activity of JWH133
The clinical manifestations of SARS-CoV-2 infections range from mild to severe, with widespread participation of the lungs, beginning from pneumonia to acute respiratory distress syndrome (ARDS), as well as acute injury to the liver, heart, intestine, coagulopathy, thrombosis, and neurological manifestations that may lead to sepsis and multi-organ failure with poor prognosis [30,31]. Widespread alveolar damage, along with progressive lung dysfunction, leads to respiratory failure that may cause fatalities [32]. Fatalities are higher in elderly people with cardiometabolic diseases, cancer, patients who are immunocompromised, or with comorbidities of diabetes or cardiometabolic diseases [33]. COVID-19 also causes interstitial lymphopenia, lymphocyte infiltration, and T cell hyperactivation in the lungs and blood [30,31].
CB2Rs are largely expressed in macrophages and participate in the inflammatory process mainly by regulating proinflammatory factors, including cytokines, chemokines, adhesion molecules, and the polarization of macrophages, a key regulator of the M1/M2 pathway of inflammation [34,35]. Activation of CB2R produces anti-inflammatory action by inhibiting leukocyte recruitment, reducing the synthesis and release of proinflammatory cytokines, such as interleukin (IL)-6, IL-18, monocyte chemoattractant protein 1, and reactive oxygen species (ROS) [7]. CB2R primarily couples with Gi/o proteins upon activation, resulting in inhibition of adenylyl cyclase agonism, further activating the 5′ AMP-activated protein kinase (AMPK) pathways that result in reduced anabolic reactions, which, in turn, promote oxidative phosphorylation and exert anti-inflammatory effects [7,36]. Several studies have demonstrated the anti-inflammatory activities of JWH133 in inflammatory models, including lipopolysaccharide (LPS)-induced macrophages, monocytes, and eosinophils by inhibiting proinflammatory cytokines, inflammatory enzymes, such as inducible nitric oxide synthase (iNOS) and cyclooxygenase (COX)-2, and production of nitric oxide (NO) and prostaglandin E2 [37,38,39].
Patients with COVID-19 mainly present with acute respiratory distress causing acute lung injuries characterized by neutrophil infiltration, vasculitis, and secretion of proinflammatory cytokines, particularly a massive increase in IL-6, which is related to the severity of the disease pathology, poor prognosis, and death [40,41]. Elevated IL-6 levels have also been demonstrated to contribute to acute lung injury (ALI) in murine models [42], similar to those observed in patients with ARDS and COVID-19; thus, inhibition of IL-6 appears to mitigate ALI [42,43]. A few of the potent inhibitors of IL-6 are tocilizumab and sarilumab; these drugs have gained attention in the inhibition of the cytokine storm in COVID-19, but possess numerous adverse effects, such as liver damage, thrombocytopenia, leukopenia, serious infections, gastrointestinal perforations, hypertension, skin reactions, and anaphylaxis [44]. Macrophages present in the human lung express CB2R, which, upon activation, significantly inhibits LPS-induced production of vascular endothelial growth factor-A and C, angiopoietins, and IL-6 secretion [45]. In addition to IL-6, the NOD-like receptor protein 3 (NLRP3) inflammasome is a mediator of the cytokine storm, and, thereby, clinical and pathological manifestations of patients infected with COVID-19 [46]. Recently, JWH133 has been found to exert protective effects in experimental models of ALI by activating CB2R [37,38]. JWH133 significantly inhibits proinflammatory cytokines, including IL-6, and improves levels of antioxidants, mediating the inhibition of inflammasomes [39], the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway [37] and the mitogen-activated protein kinase (MAPK)/c-Jun N-terminal kinase (JNK) and nuclear factor-kappa B (NF-κB) pathways [38].
Some patients that have recovered from COVID-19 are reported to have progressive post-infection consequences with persistent lung dysfunction and fibrosis, a life-threatening disease [47]. Pulmonary fibrosis begins with microinjury, resulting in inflammation and over-activation of repair mechanisms following activation of fibroblasts. CB2R present in fibroblasts plays a role in fibrosis, and many studies have demonstrated that activating CB2R exerts anti-inflammatory and antifibrotic effects [48,49,50]. Notably, JWH133, via activation of CB2R, prevents lung fibrosis and reduces fibroblast proliferation, along with suppression of autoantibodies [48]. By activating CB2R, JWH133 also inhibits hyperemia, hyperplasia of type II pneumocytes, interstitial fibrosis and salvaged lungs, reduced fibrotic markers, collagen deposition, decreased levels of the profibrotic cytokine transforming growth factor (TGF)-β1, and mitigated activation of the TGF-β1/mothers against decapentaplegic homolog 2 pathway [49,50].
Additionally, JWH133, mediating CB2R-dependent anti-inflammatory action mitigates neurogenic pulmonary edema developed following subarachnoid hemorrhage, as evidenced by lung permeability, leukocyte trafficking, and preserved tight junctions [51]. Based on the therapeutic and preventive effects of JWH133 in experimental models of ALI, drug-induced lung injuries, inflammation, and fibrosis, as well as airway hyper-responsiveness and cough centers, it is conceivable to speculate that JWH133 may have the potential to curb ALI in COVID-19. It may also limit late-onset pulmonary fibrosis in recovered patients or may be useful in patients with compromised pulmonary function. However, further proof of concept studies is needed for conclusive evidence.
In extrapulmonary manifestations of COVID-19, cardiac injury also occurs in patients with a critical illness [52]. Patients with cardiovascular disorders, such as ischemic heart disease, hypertension, and hyperlipidemia are also at a greater risk of disease severity and death [52]. Systemic infections and inflammation may cause acute thrombosis by activating platelets, vasoconstriction of the coronary artery, hypoxemia, enhanced sympathetic tone, altered heart rate, coagulation pattern, and impaired endothelium [53]. JWH133, by activating CB2R, suppresses rostral ventrolateral medulla neuroinflammation associated with hypertension by reducing blood pressure, heart rate, renal sympathetic nerve activity, and proinflammatory cytokines in spontaneously hypertensive rats [54]. JWH133 has been shown to be cardioprotective in acute myocardial injury in numerous experimental models; the protective effects are mediated by CB2R activation and inhibition of inflammasome activation [39], downregulation of receptor interacting protein 1 (RIP1)/RIP3/mixed lineage kinase domain like pseudokinase (MLKL)-mediated necroptosis [55], inhibition of cardiomyocyte hypertrophy through AMPK-endothelial NOS signaling [56], increasing extracellular signal-regulated kinase 1/2 (ERK1/2) phosphorylation and inhibiting mitochondrial permeability transition pore opening [57], suppression of Na+/Ca2+ exchanger current [58], and upregulation of integrins CD18/CD11b (Mac-1) on human neutrophils in tumor necrosis factor (TNF)-α-induced chemotaxis [59]. JWH133 also exerts vasodilatory and positive inotropic effects in spontaneously beating Langendorff-perfused rat hearts [60]. Additionally, JWH133 mitigates antipsychotic drug-induced cardiotoxicity by enhancing RIP3, MLKL, and the phosphorylation of MLKL [61], suppressing proinflammatory mediators and fibrotic markers, and improving survival rate [62].
In patients with COVID-19, cardiovascular complications are aggravated by the high incidence of venous and arterial thrombosis and coagulopathy involving platelet activation, the formation of platelet-monocyte aggregates, complement activation, increases in lipoproteins, endothelial dysfunction, stasis, hypoxia, and overexpression of tissue factors following a cytokine storm or capillary leak syndrome, in reference to thrombosis [63,64]. CB2Rs are widely expressed in hematopoietic and endothelial cells and regulate endothelial inflammation, chemotaxis, adhesion of inflammatory cells to the stimulated endothelium, and the resultant secretion of proinflammatory molecules [65,66]. The activation of CB2R attenuates inflammatory responses, including activation of endothelial cells, adhesion, and migration of immune cells, a common accompaniment of atherosclerosis and restenosis [67]. JWH133 inhibits inflammation and vascular remodeling by attenuating cell proliferation, intima and media formation, macrophage infiltration, and reducing numbers of nuclei and proliferating cells in the intima [68]. Reduced CB2R levels are found in asymptomatic patients with atherosclerosis [69]. JWH133 decreases matrix metallopeptidase 9 (MMP-9) levels in the aortic root, plaque formation in the carotid artery, and with human neutrophils, reduces TNF-α-induced ERK1/2 phosphorylation [69]. Additionally, JWH133 protects against neuroinflammation by activating CB2R enhanced trans endothelial resistance and tight junction proteins by inhibiting proinflammatory mediators, including adhesion molecules [70].
In patients with COVID-19, liver injury or dysfunction is a common issue due to the virus itself or other concurrent conditions, such as hepatotoxicity from the drugs, mainly antipyretics or immunomodulators used in COVID-19 management, or the presence and progression of chronic liver diseases, coexisting systemic inflammation, acute respiratory distress associated hypoxia, and multi-organ failure [71]. SARS-CoV-2 virus causes liver injury via many methods, including cytopathic effects via angiotensin-converting enzyme 2 (ACE2) receptors and immune-mediated hyperinflammatory state caused by cytokine storm. Numerous studies reported the incidence of impaired liver function ranging from 10.5% to 69% in patients with COVID-19 with rise in liver enzymes; alanine amino transaminase (ALT) and aspartate amino transaminase (AST), with a more specific increase in AST [3,5,11,12,13].
Patients with COVID-19 with a pre-existing liver disease or liver impairment are prone to show poorer prognosis. CB2R regulates innate immunity and is a critical mediator in liver diseases by exerting anti-inflammatory and antifibrogenic effects [72,73]. Polymorphisms in the CB2R gene following liver dysfunction in obese children suggest the role and importance of CB2R in liver diseases [74]. Numerous studies demonstrated hepatoprotective effects of JWH133 against acute liver injury or failure [75,76,77], septic liver [78], liver ischemia-reperfusion (I/R) injury [79], liver fibrosis [72,73], steatosis [75], ascites, and peritonitis [80] mediating activation of CB2R.
Hepatoprotective mechanisms include inhibition of hepatic inflammation by inducing autophagy [81], activating heme oxygenase-1, promoting an M1 to M2 shift in macrophages, and regulating microRNAs targeting Toll-like receptor 4 (TLR4) [76], inhibiting CD4+ T cell recruitment in I/R-induced injury in the liver [82], suppressing proinflammatory effects of IL-17 and its production by Th17 lymphocytes mediating a signal transducer and activator of transcription 5 (STAT5)-dependent mechanism, along with restoring IL-22 production [73], inhibiting COX, NOS, and vascular endothelial growth factor [83] reducing inflammatory cell infiltration, lipid peroxidation, restoring oxidant/antioxidant balance and levels of proinflammatory mediators [79], and the promotion of liver regeneration. JWH133 also ameliorates portal hypertension, the severity of portosystemic collaterals and mesenteric angiogenesis, intrahepatic angiogenesis, and fibrosis in cirrhotic rats [83,84].
Furthermore, JWH133 also showed to attenuate ascites and peritonitis, and inhibits inflammation and oxidative stress in cirrhosis [80]. JWH133 showed protective effects in an experimental model of acute liver injury by inhibiting ubiquitin-specific peptidase 4 (USP4), deubiquitylating TGF-β receptor 1 (TβRI), downstream of CB2 microRNA 27b, which has been identified as an inhibitor of USP4 and TβRI in hepatocytes [85].
In patients with COVID-19, acute kidney injury includes direct virus-induced tubular or glomerular injury, as well as sepsis-associated injury or thrombotic disease as complications in a significant number of severely affected patients [86]. Interestingly, JWH133 has been shown to be protective in experimental models of acute kidney injury by inhibiting pro-inflammatory cytokines, chemokines, and apoptosis [87], and salvaging kidneys [88] mediated by activation of CB2R.
Intestinal inflammation and diarrhea also occur as a complication in patients with COVID-19 due to SARS-CoV-2-mediated reduction in mucosal angiotensin-converting enzyme 2 following entry, resulting in elevated angiotensin levels and increased TNF-α and tryptophan deficiency [89]. Cannabinoid ligands have roles in inflammation, secretion, and motility, as the ECS regulates the physiology and pathophysiology of the intestine, including motility, secretion, integrity, and immunity, as well as satiety and emesis [90]. Normally, neutrophils are not present in the intestinal mucosa, but during acute inflammation they quickly infiltrate the mucosa to control the pathogen or combat inflammation [91]. If the inflammation does not resolve quickly, neutrophil infiltration leads to massive damage to the intestine. Numerous studies have demonstrated that the cannabinoid system plays an important role in intestinal inflammation induced by the synthesis or release of proinflammatory cytokines following overactivation of immune cells [92,93,94,95].
The role of CB2R has been well demonstrated in intestinal inflammation, pain, and immunity [92]. JWH133 attenuates intestinal inflammation by enhancing apoptosis of activated T cells, decreasing the numbers of activated T cells, and inhibiting the induction of neutrophils, mast cells, and NK cells at the sites of inflammation [93]. Additionally, JWH133 also corrects motility impairment in LPS-induced septic ileus by decreasing myoelectrical activity and preventing delay of gastrointestinal transit, along with inhibition of inflammation [94]. JWH133, by activating CB2R in the enteric nervous system, attenuates LPS-induced increases in intestinal contractility [95], neurogenic intestinal inflammation [96], and suggests uses in individuals experiencing diarrhea-predominant inflammatory bowel [97]. CB2R activation also attenuates intestinal ischemia-reperfusion injury by inhibiting proinflammatory cytokines and restoring the oxidant/antioxidant balance.
Patients with COVID-19 also have stroke as a complication, and as a common accompaniment with atherosclerosis, hypertension, and atrial fibrillation [98]. The pathogenesis of stroke involves endothelial dysfunction, hypercoagulopathy, microvascular thrombosis, vasculitis, hypoxia, hemodynamic and cardiac dysfunction, and systemic inflammation following a cytokine storm [98]. In a thrombin-induced in vitro and in vivo rat model and collagenase-induced germinal matrix hemorrhage in rats, JWH133 ameliorates neuroinflammation, brain edema, neuronal degeneration, microglial accumulation, and levels of phosphorylated ERK proteins [99], and protects the blood-brain barrier by reducing extravasation, activities of MMP-9 and -12, and the number of microglia in rats [100].
Additionally, COVID-19 may influence mental well-being and adversely impact immune functioning [101]. Psychosocial issues, such as stress, anxiety, and depression are believed to increase susceptibility to viral upper respiratory infections [102]. Psychological distress is linked to immune-inflammatory responses and suggests that psycho-neuroimmunity is important in COVID-19 infection [102]. JWH133 has been shown to be beneficial in relieving stress, anxiety, and depression [103,104]. Stress exposure causes excitotoxicity and neuroinflammation, which contributes to stress-related neuropathology’s, such as depression. Clinically, approximately 30% of patients with stroke develop post-stroke depression. JWH133 shows antidepressant, antistress, and anxiolytic activity in post-stroke depression induced by chronic unpredictable mild stress followed by middle cerebral artery occlusion in rats [103], and in a battery of behavior and stress models [104]. Mechanistically, the pharmacological effects are mediated by CB2R-dependent inhibition of proinflammatory cytokines and inflammatory mediators [104], interactions with the cholinergic system [105] and upregulation of serotonergic receptors, such as 5-hydroxytryptamine receptor 2A (5-HT2A) [106].
Additionally, JWH133 exhibits an antiallodynic effect in a neuropathic pain model induced by retrovirus infection by suppressing neuroinflammation, macrophage activation, and T-cell infiltration via blocking the Janus kinase/STAT3 pathway [107]. JWH133 acts as an analgesic, as it exhibits antihyperalgesic and antinociceptive actions, mainly by mitigating synthesis of proinflammatory molecules and the inhibition of nociception induced by oxidative stress-induced TRPA1 activation, inhibition of vascular permeability and migration of neutrophils, exhibiting systemic and peripheral analgesic-dependent effects on the opioid system [108].
4. CB2 Receptors Mediated Immunomodulatory Activity of JWH133
CB2R is significantly expressed in immunoregulatory cells, including macrophages, B and T cells, and upon activation leads to the subsequent inhibition of cyclic adenosine monophosphate production [109]. CB2R regulates the immune system by controlling immune cell activation through the modulation of T helper cells [110], attenuation of proinflammatory cytokines [111], and NF-κB-mediated apoptosis [112] and found useful in immune-related diseases [113]. CB2R activation has also been shown to mediate immunosuppressive activities of mesenchymal stem cells in immunocompromised conditions [114].
A recent study has demonstrated that JWH133, in combination with dexamethasone, is effective in immune thrombocytopenia purpura (ITP), an autoimmune disease characterized by antibodies against platelets [114]. The combination is effective in mesenchymal stem cells, multipotent cells that have significant roles in immunomodulation and suppress proliferation and activation of both T- and B-lymphocytes, ameliorate apoptotic cell death via B-cell lymphoma 2 signaling, and reinstate the immunomodulatory properties of mesenchymal stem cells derived from patients with ITP [114]. Recently, dexamethasone has been reported to be effective in patients with COVID-19. Thus, JWH133 may reduce the dose of dexamethasone and its adverse effects, along with maintaining its therapeutic effects due to the synergistic combination of dexamethasone and JWH133 [114].
JWH133 prevents the secretion of IL-12p40 and enhances secretion of IL-10 in LPS- or Theiler’s virus-activated macrophages, mediating activation of the CB2R-dependent ERK1/2 MAPK pathway [115]. IL-10 and IL-12 both regulate priming of Th1 or Th2 cells in immune responses. IL-12 plays a significant role in innate and adaptive immunity, and differentiates the immune system towards a Th-1 protective response against viral infections. IL-10 plays a role in maintaining the balance of appropriate macrophage responses to LPS by curbing the synthesis and release of IL-12. CB2R activation in cells belonging to macrophage lineages inhibits the induction of a Th-1 immune response, affecting the required immunity to counter a pathogen or inflammatory state [115].
5. CB2 Receptors Mediated Effects of JWH133 on Acute Lung Injury and Airway Activity
ALI in experimental models is akin to acute respiratory distress in COVID-19. ALI is caused by infections, pneumonia, sepsis, acid aspiration, toxic inhalation, and xenobiotics, which are the major causes of a cytokine storm. CB2R stimulation plays a significant role in protecting the lungs in numerous models of ALI, including cecal ligation puncture-induced septic lung injury [116], I/R-induced lung injury and LPS-induced lung injury [37], paraquat-induced ALI [38], and LPS-induced ALI [117]. JWH133, by activating CB2R, shows potent anti-inflammatory effects in LPS-induced ALI mice by reducing leukocyte migration, vascular permeability, and reducing levels of cytokines, chemokines, and adhesion molecules in the lungs and blood, along with salvaging the lungs [117].
JWH133 has been shown to protect against ALI by inhibiting proinflammatory cytokines, MAPKs, and NF-κB activation via activating CB2R [38], as shown in Figure 3. It also reduces neutrophil infiltration and edema, improving histology of the lungs along with PaO2 in arterial blood [38]. JWH133 has also been found to protect against I/R-induced ALI by reducing levels of cytokines, lipid peroxidation, neutrophil infiltration, lung edema, and improving anti-oxidant and lung histology, along with the PaO2/FiO2 ratio, mediated by the CB2R-dependent PI3K/Akt pathway [37], as shown in Figure 3. Furthermore, JWH133 exerts an antitussive effect in chronic cough by inhibiting activation of sensory nerves in guinea pig and vagus nerves in humans, and suppresses the cough reflex mediated by CB2R activation [118]. In addition to antitussive activity, JWH133 also mitigates bronchoconstriction via inhibition of pre-junctional neurotransmission, neurogenic airway inflammation, and hyper-reactivity [118,119], as shown in Figure 3.
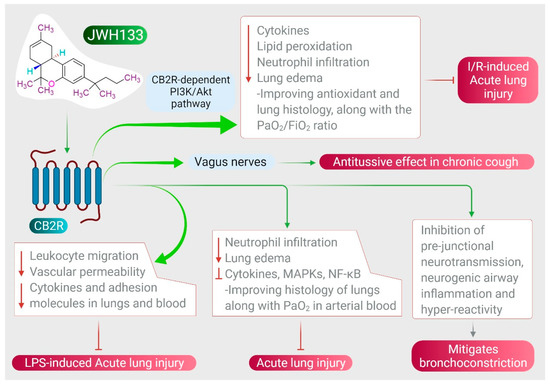
Figure 3.
Effect of JWH133 on acute lung injury and airway activity.
Furthermore, at the doses at which cannabinoids produce bronchodilation, JWH133 does not elicit respiratory depression at the central level. CB2R expressed on eosinophils plays a role in lung inflammation mediated by the generation of NO and prostaglandin-E2 [120]. CB2R is involved in antigen processing, immune cell differentiation, and macrophage migration, which have all been shown to play a role in airway immunomodulation [121,122].
7. CB2 Receptors Mediated Protective Effects of JWH133 in Organ Injuries and Sepsis
Uncontrolled infection and increased inflammatory mediators might cause a systemic inflammatory response and sepsis. CB2R-selective cannabinoids exert potent immunomodulatory and anti-inflammatory effects in the brain, pancreas, intestine, liver, heart, and kidney [34,35]. Activation of CB2R attenuates inflammatory states and oxidative stress in the liver [75,76,77], lungs [50], heart [39], kidney [87], intestine [93], brain [135], and in sepsis [78] by inhibiting inflammatory cell recruitment, proinflammatory cytokines, and increasing levels of anti-inflammatory cytokines.
In a polymicrobial sepsis model in rats [78], JWH133 shows protective effects on brain, lung, liver and, heart, mediated by CB2R activation [78]. JWH133 decreases proinflammatory cytokines and increases the anti-inflammatory cytokine IL-10 [78]. Sepsis is associated with neuronal damage and cognitive impairment, with the participation of proinflammatory cytokines and oxidative/nitrosative stress [78]. Deregulated immunity and an imbalance between the proinflammatory and anti-inflammatory systems results in multi-organ dysfunction and failure, and consequently may cause death. Acute central nervous system (CNS) injury perturbs the homeostasis of the CNS and immune system and enhances patient susceptibility to infections [136]. JWH133 shows neuroprotective effects in LPS-induced neuroinflammation and endotoxemia by mitigating levels of proinflammatory cytokines, adhesion molecules (vascular cell adhesion protein 1 and E-selectin), and oxidative/nitrosative stress [135]. Based on the role of JWH133 in ameliorating sepsis, JWH133 appears to be a potent candidate for limiting COVID-19 progression and post-infection sequelae, including its impact on the multi-organ system.
Furthermore, JWH133 also shows ROS or free radical scavenging and Fe+2 chelating activity against free radicals in numerous in vitro assays, including 2,2-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid, 2,2-diphenyl-1-picryl-hydrazyl-hydrate, ferric reducing antioxidant power, and oxygen radical absorbance capacity, with chelating and reducing power [137,138], promoting mitochondrial biogenesis [139] and improving endogenous antioxidants in vivo in many tissues. JWH133 inhibits oxidative stress, which initiates and contributes to numerous pathways, including inflammasome activation, nuclear factor erythroid 2-related factor 2 (Nrf2)/Kelch-like ECH-associated protein 1 (Keap1), TLR4/high mobility group box 1, MAPK, and sirtuin/PPAR gamma coactivator 1-α (PGC1-α) pathways, leading to the release of inflammatory mediators and cytokines that sustain inflammation, and involving metabolic reprogramming of innate immune cells [77,84,99]. Taken together, JWH133 has been shown to modulate the majority of the signaling pathways that contribute to redox immune-inflammatory signaling those results in organoprotective effects. In addition to the lungs, COVID-19 affects almost all organ systems, including the heart, brain, liver, kidney, intestine, and coagulation system. Thus, the organoprotective effects demonstrated in the in vivo experimental models are encouraging for speculation of the therapeutic benefits of JWH133.
8. Limitations on the Proposed Therapeutic Applications of JWH133
In present manuscript, the possible therapeutic role of JWH133 in COVID-19 has been proposed based on the previously reported potent pharmacological activity of JWH133 against infection, inflammation, and immunity in experimental models of human diseases involving dysregulated immune-inflammatory and redox homeostasis. Many authors proposed the hypotheses that CB2R, an important constituent in endocannabinoid system may play role in maintaining immune system and targeting inflammation and infection [18,21,140]. A recent report suggests that CB2R may play role in targeting the trinity of infection, inflammation and immune dysregulation [18,21]. Given the possible role of CB2R activation in attenuating inflammation, viral replication and favorably modulating immune systems, it has been speculated that JWH133 endowed with CB2 selective agonist property and showing affinity to Mpro may be a candidate for further investigation for its possible use in management of COVID-19. There are reports of long-term complications in some patients even after recovery from COVID-19. Thus, given the tissue protective effects and effect on numerous tissue remodeling effects, JWH133 could be a candidate to be investigated for possible use in combating the long-term complications in COVID-19. Taking into consideration the safety of JWH133 and efficacy in various disease models in experimental studies, JWH133 may be a valuable agent to be investigated further in COVID-19. The inhibitory activity on the proteases and other molecular targets should be assessed for specificity, affinity, dose-response, and kinetics in experimental studies. The binding of these compounds limits the availability of the substrate, modifies configuration of active sites, and prevents dimerization, viral entry and, viral replication. The available reports clearly demonstrate that the progression and complications of COVID-19 involves cytokine storm, therefore, cannabinoids activating CB2R may inhibit cytokine storm, coupled with their additional organ-protective effects. However, until now there is no clear evidence available on the antiviral activity of JWH133 on SARS-CoV-2. There are no data available in experimental studies whether JWH133 can protect against COVID-19 or may be useful in treatment of COVID-19. There is paucity of preclinical and clinical data on infection, inflammation, and immunity in context to COVID-19. The recent availability of animal models could be important in evaluating its preclinical efficacy. However, there is lack of clinical data and rigorous pharmacokinetics in humans. Thus, the preclinical evaluation including duration of use and dose to be explored, the safety and interaction with concomitant drugs, as well as the heterogeneity of the target population should be considered before the possible use of JWH133 in therapeutics. Nonetheless, given the preclinical studies on anti-inflammatory and immunomodulatory properties, there are opportunities for further studies to investigate the possible use in COVID-19. Considering the safety in numerous preclinical studies, further proof of the concept preclinical and translational clinical studies is encouraged to determine the clinical usage and pharmaceutical development of JWH133.
9. Conclusions
Thus, including the immunomodulatory, anti-inflammatory, and antiviral properties of JWH133 and integrating its pharmacological and molecular mechanisms, JWH133 could be a promising therapeutic candidate for COVID-19. The potent anti-inflammatory activity involves multiple pathways, including inhibition of proinflammatory cytokines, chemokines, and adhesion molecules, along with suppression of macrophage infiltration and neutrophil-endothelial cell interactions that inhibit a cytokine storm, which is a major reason for death in patients with COVID-19. JWH133 has potential as an immunomodulatory, as well as a potent anti-oxidant, in improving host cellular immunity against infection; its ability to interfere with virus replication, along with its antibacterial activity, may further help in controlling symptoms and worsening of the disease, secondary infections, complications, progression, and resultant death.
JWH133 appears non-toxic in experimental studies with no abuse potential and possesses numerous characteristics that make it an attractive therapeutic candidate to explore immunomodulatory, anti-inflammatory, and antiviral activities within the context of COVID-19. Furthermore, the drug likeliness properties, pharmacological actions, and molecular mechanisms provide a rationale for the evaluation of JWH133 as a plausible therapeutic candidate against COVID-19. However, it is important to highlight that none of the above studies have demonstrated the effect of JWH133 in COVID-19, due to the lack of a preclinical COVID-19 infected animal model to perform preclinical evaluations and to distinguish whether candidate compounds may become effective drugs. Nevertheless, previous studies have shown efficacy in limiting infection, inflammation, and immunity, which reasonably suggests JWH133 may be a potential candidate for further evaluation in COVID-19.
Author Contributions
S.O.: conceptualized, wrote, and edited the manuscript, and performed the literature survey and. N.K.J.: ideated the scheme, performed artwork and drafted the manuscript. V.D.D.: performed in silico studies. S.K.J., M.F.N.M., C.S., P.K.G., S.O., A.D., and K.K.K.: edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no specific external funding. The author, Shreesh Ojha acknowledge the research grant support from United Arab Emirates University, United Arab Emirates on researches related to study the pharmacological potential of compounds targeting cannabinoid receptors.
Institutional Review Board Statement
This is not applicable for the present manuscript.
Informed Consent Statement
This is not applicable for the present manuscript.
Data Availability Statement
This is not applicable for the present manuscript. The data reviewed herein is from previously published manuscripts and well referred appropriately. The in silico analysis was performed only in the present manuscript by the authors. It will be available upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 5-HT2A | 5-hydroxytryptamine receptor 2A |
| ACE-2 | Angiotensin-converting enzyme 2 |
| ALI | Acute lung injury |
| ALT | Alanine amino transaminase |
| ARDS | Acute respiratory distress syndrome |
| AST | Aspartate amino transaminase |
| CB1R | Cannabinoid receptor type 1 |
| CB2R | Cannabinoid receptor type 1 |
| CNS | Central nervous system |
| COVID-19 | Coronavirus disease-2019 |
| COX-2 | Cyclooxygenase-2 |
| ECS | Endocannabinoid system |
| ERK1/2 | Extracellular signal-regulated kinase ½ |
| GPCR | G-protein-coupled receptor |
| HCV | Chronic hepatitis C |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| ITP | Immune thrombocytopenia purpura |
| LPS | Lipopolysaccharide |
| MAPK/JNK | Mitogen-activated protein kinase/c-Jun N-terminal kinase |
| MLKL | Mixed lineage kinase domain like pseudo kinase |
| NF-κB | Nuclear factor-kappa B |
| NK cells | Natural killer cells |
| NLRP# | NOD-like receptor protein 3 |
| NO | Nitric oxide |
| NRF2/Keap1 | Nuclear factor erythroid 2-related factor 2/Kelch-like ECH-associated protein |
| PGCV1-α | PPAR gamma coactivator 1-α |
| PI3K/Akt | Phosphoinositide 3-kinase/protein kinase B |
| RIP | Receptor interacting protein |
| RSV | Respiratory syncytial virus |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| STAT-5 | Signal transducer and activator of transcription 5 |
| TGF | Transforming growth factor |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor |
| TRP | Transient receptor potential |
| USP4 | Ubiquitin-specific peptidase 4 |
References
- Oa, A.; Je, A. Social determinants of health: The role of effective communication in the COVID-19 pandemic in developing countries. Glob. Health Action 2020, 13, 1788263. [Google Scholar]
- Altay, O.; Mohammadi, E.; Lam, S.; Turkez, H.; Boren, J.; Nielsen, J.; Uhlen, M.; Mardinoglu, A. Current Status of COVID-19 Therapies and Drug Repositioning Applications. iScience 2020, 23, 101303. [Google Scholar] [CrossRef]
- Shiyong, F.; Dian, X.; Yanming, W.; Lianqi, L.; Xinbo, Z.; Wu, Z. Research progress on repositioning drugs and specific therapeutic drugs for SARS-CoV-2. Future Med. Chem. 2020, 0158. [Google Scholar] [CrossRef]
- Chiurchiu, V.; Battistini, L.; Maccarrone, M. Endocannabinoid signalling in innate and adaptive immunity. Immunology 2015, 144, 352–364. [Google Scholar] [CrossRef]
- Oláh, A.; Szekanecz, Z.; Bíró, T. Targeting Cannabinoid Signaling in the Immune System: “High”-ly Exciting Questions, Possibilities, and Challenges. Front. Immunol. 2017, 8, 1487. [Google Scholar] [CrossRef] [Green Version]
- Almogi-Hazan, O.; Or, R. Cannabis, the Endocannabinoid System and Immunity-the Journey from the Bedside to the Bench and Back. Int. J. Mol. Sci. 2020, 21, 4448. [Google Scholar] [CrossRef]
- Stasiulewicz, A.; Znajdek, K.; Grudzień, M.; Pawiński, T.; Sulkowska, A. Guide to Targeting the Endocannabinoid System in Drug Design. Int. J. Mol. Sci. 2020, 21, 2778. [Google Scholar] [CrossRef] [PubMed]
- Argenziano, M.; Tortora, C.; Bellini, G.; Di Paola, A.; Punzo, F.; Rossi, F. The Endocannabinoid System in Pediatric Inflammatory and Immune Diseases. Int. J. Mol. Sci. 2019, 20, 5875. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Xiao, F.; Yuan, Q.; Zhang, J.; Zhan, J.; Zhang, Z. Cannabinoid Receptor 2: A Potential Novel Therapeutic Target for Sepsis? Acta Clin. Belg. 2019, 74, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.T.; Greaves, D.R.; Iqbal, A.J. The Impact of Cannabinoid Receptor 2 Deficiency on Neutrophil Recruitment and Inflammation. DNA Cell Biol. 2019, 38, 1025–1029. [Google Scholar] [CrossRef]
- Huffman, J.W.; Hepburn, S.A.; Lyutenko, N.; Thompson, A.L.; Wiley, J.L.; Selley, D.E.; Martin, B.R. 1-Bromo-3-(1′,1′-dimethylalkyl)-1-deoxy-Δ(8)-tetrahydrocannabinols: New selective ligands for the cannabinoid CB(2) receptor. Bioorganic Med. Chem. 2010, 18, 7809–7815. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Wang, L.; Kuo, H.-C.D.; Shannar, A.; Peter, R.; Chou, P.J. An Update on Current Therapeutic Drugs Treating COVID-19. Curr. Pharmacol. Rep. 2020, 6, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Song, J.W.; Zhang, C.; Fan, X.; Meng, F.P.; Xu, Z.; Xia, P.; Cao, W.J.; Yang, T.; Dai, X.P.; Wang, S.Y.; et al. Immunological and inflammatory profiles in mild and severe cases of COVID-19. Nat. Commun. 2020, 11, 3410. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fehr, A.R.; Vijay, R.; Mack, M.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Dysregulated Type I Interferon and Inflammatory Monocyte-Macrophage Responses Cause Lethal Pneumonia in SARS-Cova-Infected Mice. Cell Host Microbe 2016, 19, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Lythgoe, M.P.; Middleton, P. Ongoing Clinical Trials for the Management of the COVID-19 Pandemic. Trends Pharmacol. Sci. 2020, 41, 363–382. [Google Scholar] [CrossRef]
- Allegra, A.; Di Gioacchino, M.; Tonacci, A.; Musolino, C.; Gangemi, S. Immunopathology of SARS-CoV-2 Infection: Immune Cells and Mediators, Prognostic Factors, and Immune-Therapeutic Implications. Int. J. Mol. Sci. 2020, 21, 4782. [Google Scholar] [CrossRef]
- Henderson, L.A.; Canna, S.W.; Schulert, G.S.; Volpi, S.; Lee, P.Y.; Kernan, K.F.; Caricchio, R.; Mahmud, S.; Hazen, M.M.; Halyabar, O.; et al. On the Alert for Cytokine Storm: Immunopathology in COVID-19. Arthritis Rheumatol. 2020, 72, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Tortora, C.; Argenziano, M.; Di Paola, A.; Punzo, F. Cannabinoid Receptor Type 2: A Possible Target in SARS-CoV-2 (CoV-19) Infection? Int. J. Mol. Sci. 2020, 21, 3809. [Google Scholar] [CrossRef]
- Peyravian, N.; Deo, S.; Daunert, S.; Jimenez, J.J. Cannabidiol as a Novel Therapeutic for Immune Modulation. ImmunoTargets Ther. 2020, 9, 131–140. [Google Scholar] [CrossRef]
- Rizzo, M.D.; Henriquez, J.E.; Blevins, L.K.; Bach, A.; Crawford, R.B.; Kaminski, N.E. Targeting Cannabinoid Receptor 2 on Peripheral Leukocytes to Attenuate Inflammatory Mechanisms Implicated in HIV-Associated Neurocognitive Disorder. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2020, 15, 780–793. [Google Scholar] [CrossRef]
- Nagoor Meeran, M.F.; Sharma, C.; Goyal, S.N.; Kumar, S.; Ojha, S. CB2 receptor-selective agonists as candidates for targeting infection, inflammation, and immunity in SARS-CoV-2 infections. Drug Dev. Res. 2021, 82, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, S.; Azhar, E.I.; Kamal, M.A.; Bajrai, L.H.; Dubey, A.; Jha, K.; Yadava, U.; Kang, S.G.; Dwivedi, V.D. SARS-CoV-2 M pro inhibitors: Identification of anti-SARS-CoV-2 M pro compounds from FDA approved drugs. J. Biomol. Struct. Dyn. 2020, 1–16. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Lee, K.E.; Dwivedi, V.D.; Kang, S.G. Computational insights into tetracyclines as inhibitors against SARS-CoV-2 M pro via combinatorial molecular simulation calculations. Life Sci. 2020, 257, 118080. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of M pro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Rostkowski, M.; Olsson, M.H.; Søndergaard, C.R.; Jensen, J.H. Graphical analysis of pH-dependent properties of proteins predicted using PROPKA. BMC Struct. Biol. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Muralidharan, N.; Sakthivel, R.; Velmurugan, D.; Gromiha, M.M. Computational studies of drug repurposing and synergism of lopinavir, oseltamivir and ritonavir binding with SARS-CoV-2 protease against COVID-19. J. Biomol. Struct. Dyn. 2020, 39, 2673–2678. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, e26. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Hiyoshi, A.; Montgomery, S. COVID-19 case-fatality rate and demographic and socioeconomic influencers: Worldwide spatial regression analysis based on country-level data. BMJ Open 2020, 10, e043560. [Google Scholar] [CrossRef] [PubMed]
- Kapellos, T.S.; Taylor, L.; Feuerborn, A.; Valaris, S.; Hussain, M.T.; Rainger, G.E.; Greaves, D.R.; Iqbal, A.J. Cannabinoid receptor 2 deficiency exacerbates inflammation and neutrophil recruitment. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 6154–6167. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, C.; Blanchet, M.R.; Laviolette, M.; Flamand, N. The CB 2 receptor and its role as a regulator of inflammation. Cellular Mol. Life Sci. CMLS 2016, 73, 4449–4470. [Google Scholar] [CrossRef] [Green Version]
- van Niekerk, G.; Mabin, T.; Engelbrecht, A.M. Anti-inflammatory mechanisms of cannabinoids: An immunometabolic perspective. Inflammopharmacology 2019, 27, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Li, X.; Cheng, Y.; Ke, B.; Wang, R. Activation of cannabinoid receptor type 2 reduces lung ischemia reperfusion injury through PI3K/Akt pathway. Int. J. Clin. Exp. Pathol. 2019, 12, 4096–4105. [Google Scholar] [PubMed]
- Liu, Z.; Wang, Y.; Zhao, H.; Zheng, Q.; Xiao, L.; Zhao, M. CB2 receptor activation ameliorates the proinflammatory activity in acute lung injury induced by paraquat. BioMed Res. Int. 2014, 2014, 971750. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Jin, G.; Zhang, J.; Wei, W. Selective Activation of Cannabinoid Receptor 2 Attenuates Myocardial Infarction via Suppressing NLRP3 Inflammasome. Inflammation 2019, 42, 904–914. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microbe 2020, 27, 992–1000. [Google Scholar] [CrossRef]
- Han, H.; Ma, Q.; Li, C.; Liu, R.; Zhao, L.; Wang, W.; Zhang, P.; Liu, X.; Gao, G.; Liu, F.; et al. Profiling serum cytokines in COVID-19 patients reveals IL-6 and IL-10 are disease severity predictors. Emerg. Microbes Infect. 2020, 9, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.L.; Sammani, S.; Kempf, C.; Saadat, L.; Letsiou, E.; Wang, T.; Moreno-Vinasco, L.; Rizzo, A.N.; Fortman, J.D.; Garcia, J.G. Pleiotropic effects of interleukin-6 in a “two-hit” murine model of acute respiratory distress syndrome. Pulm. Circ. 2014, 4, 280–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelaia, C.; Tinello, C.; Vatrella, A.; De Sarro, G.; Pelaia, G. Lung under attack by COVID-19-induced cytokine storm: Pathogenic mechanisms and therapeutic implications. Ther. Adv. Respir. Dis. 2020, 14, 1753466620933508. [Google Scholar] [CrossRef]
- Zhang, S.; Li, L.; Shen, A.; Chen, Y.; Qi, Z. Rational Use of Tocilizumab in the Treatment of Novel Coronavirus Pneumonia. Clin. Drug Investig. 2020, 40, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Staiano, R.I.; Loffredo, S.; Borriello, F.; Iannotti, F.A.; Piscitelli, F.; Orlando, P.; Secondo, A.; Granata, F.; Lepore, M.T.; Fiorelli, A.; et al. Human lung-resident macrophages express CB1 and CB2 receptors whose activation inhibits the release of angiogenic and lymphangiogenic factors. J. Leukoc. Biol. 2016, 99, 531–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; MacAry, P.A.; Ng, L.F.P. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef]
- Fang, Y.; Zhou, J.; Ding, X.; Ling, G.; Yu, S. Pulmonary fibrosis in critical ill patients recovered from COVID-19 pneumonia: Preliminary experience. Am. J. Emerg. Med. 2020, 38, 2134–2138. [Google Scholar] [CrossRef]
- Servettaz, A.; Kavian, N.; Nicco, C.; Deveaux, V.; Chéreau, C.; Wang, A.; Zimmer, A.; Lotersztajn, S.; Weill, B.; Batteux, F. Targeting the cannabinoid pathway limits the development of fibrosis and autoimmunity in a mouse model of systemic sclerosis. Am. J. Pathol. 2010, 177, 187–196. [Google Scholar] [CrossRef]
- Fu, Q.; Zheng, Y.; Dong, X.; Wang, L.; Jiang, C.G. Activation of cannabinoid receptor type 2 by JWH133 alleviates bleomycin-induced pulmonary fibrosis in mice. Oncotarget 2017, 8, 103486–103498. [Google Scholar] [CrossRef] [Green Version]
- Wawryk-Gawda, E.; Chłapek, K.; Zarobkiewicz, M.K.; Lis-Sochocka, M.; Chylińska-Wrzos, P.; Boguszewska-Czubara, A.; Sławiński, M.A.; Franczak, A.; Jodłowska-Jędrych, B.; Biała, G. CB2R agonist prevents nicotine induced lung fibrosis. Exp. Lung Res. 2018, 44, 344–351. [Google Scholar] [CrossRef]
- Fujii, M.; Sherchan, P.; Soejima, Y.; Doycheva, D.; Zhao, D.; Zhang, J.H. Cannabinoid Receptor Type 2 Agonist Attenuates Acute Neurogenic Pulmonary Edema by Preventing Neutrophil Migration after Subarachnoid Hemorrhage in Rats. Acta Neurochir. Suppl. 2016, 121, 135–139. [Google Scholar]
- Krittanawong, C.; Kumar, A.; Hahn, J.; Wang, Z.; Zhang, H.J.; Sun, T.; Bozkurt, B.; Ballantyne, C.M.; Virani, S.S.; Halperin, J.L.; et al. Cardiovascular risk and complications associated with COVID-19. Am. J. Cardiovasc. Dis. 2020, 10, 479–489. [Google Scholar] [PubMed]
- Capone, V.; Cuomo, V.; Esposito, R.; Canonico, M.E.; Ilardi, F.; Prastaro, M.; Esposito, G.; Santoro, C. Epidemiology, prognosis and clinical manifestation of cardiovascular disease in COVID-19. Expert Rev. Cardiovasc. Ther. 2020, 18, 531–539. [Google Scholar] [CrossRef]
- Shi, H.K.; Guo, H.C.; Liu, H.Y.; Zhang, Z.L.; Hu, M.Y.; Zhang, Y.; Li, Q. Cannabinoid type 2 receptor agonist JWH133 decreases blood pressure of spontaneously hypertensive rats through relieving inflammation in the rostral ventrolateral medulla of the brain. J. Hypertens. 2020, 38, 886–895. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, D.; Dong, X.; Zhu, R.; Ye, Y.; Li, L.; Jiang, Y. Pharmacological activation of CB2 receptor protects against ethanol-induced myocardial injury related to RIP1/RIP3/MLKL-mediated necroptosis. Mol. Cell. Biochem. 2020, 474, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Akinwumi, B.C.; Shao, Z.; Anderson, H.D. Ligand activation of cannabinoid receptors attenuates hypertrophy of neonatal rat cardiomyocytes. J. Cardiovasc. Pharmacol. 2014, 64, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Guo, H.C.; Maslov, L.N.; Qiao, X.W.; Zhou, J.J.; Zhang, Y. Mitochondrial permeability transition pore plays a role in the cardioprotection of CB2 receptor against ischemia-reperfusion injury. Can. J. Physiol. Pharmacol. 2014, 92, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Cui, N.; Du, Y.; Ma, H.; Zhang, Y. Anandamide reduces intracellular Ca2+ concentration through suppression of Na+/Ca2+ exchanger current in rat cardiac myocytes. PLoS ONE 2013, 8, e006386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecucco, F.; Lenglet, S.; Braunersreuther, V.; Burger, F.; Pelli, G.; Bertolotto, M.; Mach, F.; Steffens, S. CB(2) cannabinoid receptor activation is cardioprotective in a mouse model of ischemia/reperfusion. J. Mol. Cell. Cardiol. 2009, 46, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.A.; Abesser, M.; Karcher, J.; Laser, M.; Kunos, G. Coronary vasodilator effects of endogenous cannabinoids in vasopressin-preconstricted unpaced rat isolated hearts. J. Cardiovasc. Pharmacol. 2005, 46, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, Z.; Zhou, Y.; Wang, J.; Lin, X.; Dong, X.; Liu, X.; Jiang, J.; Jiang, Y.; Li, L. Quetiapine induces myocardial necroptotic cell death through bidirectional regulation of cannabinoid receptors. Toxicol. Lett. 2019, 313, 77–90. [Google Scholar] [CrossRef]
- Li, L.; Dong, X.; Tu, C.; Li, X.; Peng, Z.; Zhou, Y.; Zhang, D.; Jiang, J.; Burke, A.; Zhao, Z.; et al. Opposite effects of cannabinoid CB 1 and CB 2 receptors on antipsychotic clozapine-induced cardiotoxicity. Br. J. Pharmacol. 2019, 176, 890–905. [Google Scholar] [CrossRef] [PubMed]
- Gąsecka, A.; Borovac, J.A.; Guerreiro, R.A.; Giustozzi, M.; Parker, W.; Caldeira, D.; Chiva-Blanch, G. Thrombotic Complications in Patients with COVID-19: Pathophysiological Mechanisms, Diagnosis, and Treatment. Cardiovasc. Drugs Ther. 2020, 35, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tecson, K.M.; McCullough, P.A. Endothelial dysfunction contributes to COVID-19-associated vascular inflammation and coagulopathy. Rev. Cardiovasc. Med. 2020, 21, 315–319. [Google Scholar] [CrossRef]
- Rajesh, M.; Mukhopadhyay, P.; Bátkai, S.; Haskó, G.; Liaudet, L.; Huffman, J.W.; Csiszar, A.; Ungvari, Z.; Mackie, K.; Chatterjee, S.; et al. CB2-receptor stimulation attenuates TNF-alpha-induced human endothelial cell activation, transendothelial migration of monocytes, and monocyte-endothelial adhesion. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2210–H2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netherland, C.D.; Pickle, T.G.; Bales, A.; Thewke, D.P. Cannabinoid receptor type 2 (CB2) deficiency alters atherosclerotic lesion formation in hyperlipidemic Ldlr-null mice. Atherosclerosis 2010, 213, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yuan, Z.; Liu, Y.; Xue, J.; Tian, Y.; Liu, W.; Zhang, W.; Shen, Y.; Xu, W.; Liang, X.; et al. Activation of cannabinoid CB2 receptor ameliorates atherosclerosis associated with suppression of adhesion molecules. J. Cardiovasc. Pharmacol. 2010, 55, 292–298. [Google Scholar] [CrossRef]
- Molica, F.; Matter, C.M.; Burger, F.; Pelli, G.; Lenglet, S.; Zimmer, A.; Pacher, P.; Steffens, S. Cannabinoid receptor CB2 protects against balloon-induced neointima formation. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H1064–H1074. [Google Scholar] [CrossRef] [Green Version]
- Montecucco, F.; Di Marzo, V.; da Silva, R.F.; Vuilleumier, N.; Capettini, L.; Lenglet, S.; Pagano, S.; Piscitelli, F.; Quintao, S.; Bertolotto, M.; et al. The activation of the cannabinoid receptor type 2 reduces neutrophilic protease-mediated vulnerability in atherosclerotic plaques. Eur. Heart J. 2012, 33, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, S.H.; Haskó, J.; Skuba, A.; Fan, S.; Dykstra, H.; McCormick, R.; Reichenbach, N.; Krizbai, I.; Mahadevan, A.; Zhang, M.; et al. Activation of cannabinoid receptor 2 attenuates leukocyte-endothelial cell interactions and blood-brain barrier dysfunction under inflammatory conditions. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 4004–4016. [Google Scholar] [CrossRef]
- Feng, G.; Zheng, K.I.; Yan, Q.Q.; Rios, R.S.; Targher, G.; Byrne, C.D.; Poucke, S.V.; Liu, W.Y.; Zheng, M.H. COVID-19 and Liver Dysfunction: Current Insights and Emergent Therapeutic Strategies. J. Clin. Transl. Hepatol. 2020, 8, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Luque, J.; Ros, J.; Fernández-Varo, G.; Tugues, S.; Morales-Ruiz, M.; Alvarez, C.E.; Friedman, S.L.; Arroyo, V.; Jiménez, W. Regression of fibrosis after chronic stimulation of cannabinoid CB2 receptor in cirrhotic rats. J. Pharmacol. Exp. Ther. 2008, 324, 475–483. [Google Scholar] [CrossRef] [Green Version]
- Guillot, A.; Hamdaoui, N.; Bizy, A.; Zoltani, K.; Souktani, R.; Zafrani, E.S.; Mallat, A.; Lotersztajn, S.; Lafdil, F. Cannabinoid receptor 2 counteracts interleukin-17-induced immune and fibrogenic responses in mouse liver. Hepatology 2014, 59, 296–306. [Google Scholar] [CrossRef]
- Rossi, F.; Bellini, G.; Nobili, B.; Maione, S.; Perrone, L.; del Giudice, E.M. Association of the cannabinoid receptor 2 (CB2) Gln63Arg polymorphism with indices of liver damage in obese children: An alternative way to highlight the CB2 hepatoprotective properties. Hepatology 2011, 54, 1102–1103. [Google Scholar] [CrossRef] [PubMed]
- Denaës, T.; Lodder, J.; Chobert, M.N.; Ruiz, I.; Pawlotsky, J.M.; Lotersztajn, S.; Teixeira-Clerc, F. The Cannabinoid Receptor 2 Protects Against Alcoholic Liver Disease Via a Macrophage Autophagy-Dependent Pathway. Sci. Rep. 2016, 6, 28806. [Google Scholar] [CrossRef] [Green Version]
- Tomar, S.; Zumbrun, E.E.; Nagarkatti, M.; Nagarkatti, P.S. Protective role of cannabinoid receptor 2 activation in galactosamine/lipopolysaccharide-induced acute liver failure through regulation of macrophage polarization and microRNAs. J. Pharmacol. Exp. Ther. 2015, 353, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louvet, A.; Teixeira-Clerc, F.; Chobert, M.N.; Deveaux, V.; Pavoine, C.; Zimmer, A.; Pecker, F.; Mallat, A.; Lotersztajn, S. Cannabinoid CB2 receptors protect against alcoholic liver disease by regulating Kupffer cell polarization in mice. Hepatology. 2011, 54, 1217–1226. [Google Scholar] [CrossRef]
- Çakır, M.; Tekin, S.; Okan, A.; Çakan, P.; Doğanyiğit, Z. The ameliorating effect of cannabinoid type 2 receptor activation on brain, lung, liver and heart damage in cecal ligation and puncture-induced sepsis model in rats. Int. Immunopharmacol. 2020, 78, 105978. [Google Scholar] [CrossRef]
- Bátkai, S.; Osei-Hyiaman, D.; Pan, H.; El-Assal, O.; Rajesh, M.; Mukhopadhyay, P.; Hong, F.; Harvey-White, J.; Jafri, A.; Haskó, G.; et al. Cannabinoid-2 receptor mediates protection against hepatic ischemia/reperfusion injury. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2007, 21, 1788–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.Y.; Hsieh, S.L.; Lee, P.C.; Yeh, Y.C.; Lee, K.C.; Hsieh, Y.C.; Wang, Y.W.; Lee, T.Y.; Huang, Y.H.; Chan, C.C.; et al. Long-term cannabinoid type 2 receptor agonist therapy decreases bacterial translocation in rats with cirrhosis and ascites. J. Hepatol. 2014, 61, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Hashiesh, H.M.; Sharma, C.; Goyal, S.N.; Jha, N.K.; Ojha, S. Pharmacological Properties, Therapeutic Potential and Molecular Mechanisms of JWH133, a CB2 Receptor-Selective Agonist. Front Pharm. 2021, 12, 702675. [Google Scholar] [CrossRef]
- Reifart, J.; Rentsch, M.; Mende, K.; Coletti, R.; Sobocan, M.; Thasler, W.E.; Khandoga, A. Modulating CD4+ T cell migration in the postischemic liver: Hepatic stellate cells as new therapeutic target? Transplantation 2015, 99, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Wang, S.S.; Hsin, I.F.; Chang, C.C.; Lee, F.Y.; Lin, H.C.; Chuang, C.L.; Lee, J.Y.; Hsieh, H.G.; Lee, S.D. Cannabinoid receptor 2 agonist ameliorates mesenteric angiogenesis and portosystemic collaterals in cirrhotic rats. Hepatology 2012, 56, 248–258. [Google Scholar] [CrossRef]
- Steib, C.J.; Gmelin, L.; Pfeiler, S.; Schewe, J.; Brand, S.; Göke, B.; Gerbes, A.L. Functional relevance of the cannabinoid receptor 2-heme oxygenase pathway: A novel target for the attenuation of portal hypertension. Life Sci. 2013, 93, 543–551. [Google Scholar] [CrossRef]
- Wu, H.M.; Kim, T.H.; Kim, A.; Koo, J.H.; Joo, M.S.; Kim, S.G. Liver X Receptor α-Induced Cannabinoid Receptor 2 Inhibits Ubiquitin-Specific Peptidase 4 Through miR-27b, Protecting Hepatocytes From TGF-β. Hepatol. Commun. 2019, 3, 1373–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farouk, S.S.; Fiaccadori, E.; Cravedi, P.; Campbell, K.N. COVID-19 and the kidney: What we think we know so far and what we don’t. J. Nephrol. 2020, 33, 1213–1218. [Google Scholar] [CrossRef] [PubMed]
- Çakır, M.; Tekin, S.; Doğanyiğit, Z.; Çakan, P.; Kaymak, E. The protective effect of cannabinoid type 2 receptor activation on renal ischemia-reperfusion injury. Mol. Cell. Biochem. 2019, 462, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Feizi, A.; Jafari, M.R.; Hamedivafa, F.; Tabrizian, P.; Djahanguiri, B. The preventive effect of cannabinoids on reperfusion-induced ischemia of mouse kidney. Exp. Toxicol. Pathol. Official J. Ges. Toxikol. Pathol. 2008, 60, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Royce, S.G.; Lubel, J.S. Letter: Intestinal inflammation, COVID-19 and gastrointestinal ACE2-exploring RAS inhibitors. Aliment. Pharmacol. Ther. 2020, 52, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Smid, S.D. Gastrointestinal endocannabinoid system: Multifaceted roles in the healthy and inflamed intestine. Clinical Exp. Pharmacol. Physiol. 2008, 35, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Neish, A.S. Acute inflammation: Endogenous cannabinoids mellow the harsh proinflammatory environment. J. Clin. Investig. 2018, 128, 3750–3751. [Google Scholar] [CrossRef]
- Izzo, A.A. The cannabinoid CB(2) receptor: A good friend in the gut. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2007, 19, 704–708. [Google Scholar] [CrossRef]
- Singh, U.P.; Singh, N.P.; Singh, B.; Price, R.L.; Nagarkatti, M.; Nagarkatti, P.S. Cannabinoid receptor-2 (CB2) agonist ameliorates colitis in IL-10(-/-) mice by attenuating the activation of T cells and promoting their apoptosis. Toxicol. Appl. Pharmacol. 2012, 258, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Li, Y.N.; Ni, J.B.; Chen, C.J.; Lv, S.; Chai, S.Y.; Wu, R.H.; Yüce, B.; Storr, M. Involvement of cannabinoid-1 and cannabinoid-2 receptors in septic ileus. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2010, 22, 350–388. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.; Mouihate, A.; Mackie, K.; Keenan, C.M.; Buckley, N.E.; Davison, J.S.; Patel, K.D.; Pittman, Q.J.; Sharkey, K.A. Cannabinoid CB2 receptors in the enteric nervous system modulate gastrointestinal contractility in lipopolysaccharide-treated rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimball, E.S.; Schneider, C.R.; Wallace, N.H.; Hornby, P.J. Agonists of cannabinoid receptor 1 and 2 inhibit experimental colitis induced by oil of mustard and by dextran sulfate sodium. Am. J. Physiol. Gastrointest. and Liver Physiol. 2006, 291, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Kimball, E.S.; Wallace, N.H.; Schneider, C.R.; D’Andrea, M.R.; Hornby, P.J. Small intestinal cannabinoid receptor changes following a single colonic insult with oil of mustard in mice. Front. Pharmacol. 2010, 1, 132. [Google Scholar] [CrossRef] [Green Version]
- Ojo, A.S.; Balogun, S.A.; Idowu, A.O. Acute Ischemic Stroke in COVID-19: Putative Mechanisms, Clinical Characteristics, and Management. Neurol. Res. Int. 2020, 2020, 7397480. [Google Scholar] [CrossRef]
- Tang, J.; Tao, Y.; Tan, L.; Yang, L.; Niu, Y.; Chen, Q.; Yang, Y.; Feng, H.; Chen, Z.; Zhu, G. Cannabinoid receptor 2 attenuates microglial accumulation and brain injury following germinal matrix hemorrhage via ERK dephosphorylation in vivo and in vitro. Neuropharmacology 2015, 95, 424–433. [Google Scholar] [CrossRef]
- Li, L.; Tao, Y.; Tang, J.; Chen, Q.; Yang, Y.; Feng, Z.; Chen, Y.; Yang, L.; Yang, Y.; Zhu, G.; et al. A Cannabinoid Receptor 2 Agonist Prevents Thrombin-Induced Blood-Brain Barrier Damage via the Inhibition of Microglial Activation and Matrix Metalloproteinase Expression in Rats. Transl. Stroke Res. 2015, 6, 467–477. [Google Scholar] [CrossRef]
- Rajkumar, R.P. COVID-19 and mental health: A review of the existing literature. Asian J. Psychiatry 2020, 52, 102066. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.; Zachariae, R.; Bovbjerg, D.H. Influence of psychological stress on upper respiratory infection-a meta-analysis of prospective studies. Psychosom. Med. 2010, 72, 823–832. [Google Scholar] [CrossRef]
- Wang, S.; Sun, H.; Liu, S.; Wang, T.; Guan, J.; Jia, J. Role of hypothalamic cannabinoid receptors in post-stroke depression in rats. Brain Res. Bull. 2016, 121, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Zoppi, S.; Madrigal, J.L.; Caso, J.R.; García-Gutiérrez, M.S.; Manzanares, J.; Leza, J.C.; García-Bueno, B. Regulatory role of the cannabinoid CB2 receptor in stress-induced neuroinflammation in mice. Br. J. Pharmacol. 2014, 171, 2814–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruk-Slomka, M.; Michalak, A.; Biala, G. Antidepressant-like effects of the cannabinoid receptor ligands in the forced swimming test in mice: Mechanism of action and possible interactions with cholinergic system. Behav. Brain Res. 2015, 284, 24–36. [Google Scholar] [CrossRef]
- Franklin, J.M.; Vasiljevik, T.; Prisinzano, T.E.; Carrasco, G.A. Cannabinoid 2 receptor- and beta Arrestin 2-dependent upregulation of serotonin 2A receptors. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2013, 23, 760–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, W.S.; Chauhan, P.; Hu, S.; Prasad, S.; Lokensgard, J.R. Antiallodynic Effects of Cannabinoid Receptor 2 (CB 2 R) Agonists on Retrovirus Infection-Induced Neuropathic Pain. Pain Res. Manag. 2019, 2019, 1260353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, A.A.; Silva, R.O.; Nicolau, L.A.; de Brito, T.V.; de Sousa, D.P.; Barbosa, A.L.; de Freitas, R.M.; Lopes, L.D.; Medeiros, J.R.; Ferreira, P.M. Physio-pharmacological Investigations About the Anti-inflammatory and Antinociceptive Efficacy of (+)-Limonene Epoxide. Inflammation 2017, 40, 511–522. [Google Scholar] [CrossRef]
- Munro, S.; Thomas, K.L.; Abu-Shaar, M. Molecular characterization of a peripheral receptor for cannabinoids. Nature 1993, 365, 61–65. [Google Scholar] [CrossRef]
- Yuan, M.; Kiertscher, S.M.; Cheng, Q.; Zoumalan, R.; Tashkin, D.P.; Roth, M.D. Delta 9-Tetrahydrocannabinol regulates Th1/Th2 cytokine balance in activated human T cells. J. Neuroimmunol. 2002, 133, 124–131. [Google Scholar] [CrossRef]
- Klein, T.W.; Lane, B.; Newton, C.A.; Friedman, H. The cannabinoid system and cytokine network. Proc. Soc. Exp. Biol. Med. 2000, 225, 1–8. [Google Scholar] [CrossRef]
- Do, Y.; McKallip, R.J.; Nagarkatti, M.; Nagarkatti, P.S. Activation through cannabinoid receptors 1 and 2 on dendritic cells triggers NF-kappaB-dependent apoptosis: Novel role for endogenous and exogenous cannabinoids in immunoregulation. J. Immunol. 2004, 173, 2373–2382. [Google Scholar] [CrossRef] [Green Version]
- Leleu-Chavain, N.; Desreumaux, P.; Chavatte, P.; Millet, R. Therapeutical potential of CB2 receptors in immune-related diseases. Curr. Mol. Pharmacol. 2013, 6, 183–203. [Google Scholar] [CrossRef]
- Rossi, F.; Tortora, C.; Palumbo, G.; Punzo, F.; Argenziano, M.; Casale, M.; Di Paola, A.; Locatelli, F.; Perrotta, S. CB2 Receptor Stimulation and Dexamethasone Restore the Anti-Inflammatory and Immune-Regulatory Properties of Mesenchymal Stromal Cells of Children with Immune Thrombocytopenia. Int. J. Mol. Sci. 2019, 20, 1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, F.; Mestre, L.; Docagne, F.; Guaza, C. Activation of cannabinoid CB2 receptor negatively regulates IL-12p40 production in murine macrophages: Role of IL-10 and ERK1/2 kinase signaling. Br. J. Pharmacol. 2005, 145, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, A.P.; Yuan, Q.H.; Zhang, B.; Yang, L.; He, Q.W.; Chen, K.; Liu, Q.S.; Li, Z.; Zhan, J. Cannabinoid Receptor 2 Activation Alleviates Septic Lung Injury by Promoting Autophagy via Inhibition of Inflammatory Mediator Release. Cell. Signal. 2020, 69, 109556. [Google Scholar] [CrossRef] [PubMed]
- Costola-de-Souza, C.; Ribeiro, A.; Ferraz-de-Paula, V.; Calefi, A.S.; Aloia, T.P.; Gimenes-Júnior, J.A.; de Almeida, V.I.; Pinheiro, M.L.; Palermo-Neto, J. Monoacylglycerol lipase (MAGL) inhibition attenuates acute lung injury in mice. PLoS ONE 2013, 8, e77706. [Google Scholar] [CrossRef]
- Patel, H.J.; Birrell, M.A.; Crispino, N.; Hele, D.J.; Venkatesan, P.; Barnes, P.J.; Yacoub, M.H.; Belvisi, M.G. Inhibition of guinea-pig and human sensory nerve activity and the cough reflex in guinea-pigs by cannabinoid (CB2) receptor activation. Br. J. Pharmacol. 2003, 140, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Bozkurt, T.E.; Kaya, Y.; Durlu-Kandilci, N.T.; Onder, S.; Sahin-Erdemli, I. The effect of cannabinoids on dinitrofluorobenzene-induced experimental asthma in mice. Respir. Physiol. Neurobiol. 2016, 231, 7–13. [Google Scholar] [CrossRef]
- Oka, S.; Ikeda, S.; Kishimoto, S.; Gokoh, M.; Yanagimoto, S.; Waku, K.; Sugiura, T. 2-arachidonoylglycerol, an endogenous cannabinoid receptor ligand, induces the migration of EoL-1 human eosinophilic leukemia cells and human peripheral blood eosinophils. J. Leukoc. Biol. 2004, 76, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- McCoy, K.L.; Matveyeva, M.; Carlisle, S.J.; Cabral, G.A. Cannabinoid inhibition of the processing of intact lysozyme by macrophages: Evidence for CB2 receptor participation. J. Pharmacol. Exp. Ther. 1999, 289, 1620–1625. [Google Scholar]
- Carayon, P.; Marchand, J.; Dussossoy, D.; Derocq, J.M.; Jbilo, O.; Bord, A.; Bouaboula, M.; Galiègue, S.; Mondière, P.; Pénarier, G.; et al. Modulation and functional involvement of CB2 peripheral cannabinoid receptors during B-cell differentiation. Blood 1998, 92, 3605–3615. [Google Scholar] [CrossRef]
- Karmaus, P.W.; Chen, W.; Kaplan, B.L.; Kaminski, N.E. Δ9-tetrahydrocannabinol suppresses cytotoxic T lymphocyte function independent of CB1 and CB 2, disrupting early activation events. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2012, 7, 843–855. [Google Scholar] [CrossRef] [Green Version]
- Buchweitz, J.P.; Karmaus, P.W.; Williams, K.J.; Harkema, J.R.; Kaminski, N.E. Targeted deletion of cannabinoid receptors CB1 and CB2 produced enhanced inflammatory responses to influenza A/PR/8/34 in the absence and presence of Delta9-tetrahydrocannabinol. J. Leukoc. Biol. 2008, 83, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Tahamtan, A.; Samieipoor, Y.; Nayeri, F.S.; Rahbarimanesh, A.A.; Izadi, A.; Rashidi-Nezhad, A.; Tavakoli-Yaraki, M.; Farahmand, M.; Bont, L.; Shokri, F.; et al. Effects of Cannabinoid Receptor Type 2 in Respiratory Syncytial Virus Infection in Human Subjects and Mice. Virulence 2018, 9, 217–230. [Google Scholar] [CrossRef] [Green Version]
- Coppola, N.; Zampino, R.; Bellini, G.; Macera, M.; Marrone, A.; Pisaturo, M.; Boemio, A.; Nobili, B.; Pasquale, G.; Maione, S.; et al. Association between a polymorphism in cannabinoid receptor 2 and severe necroinflammation in patients with chronic hepatitis C. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2014, 12, 334–340. [Google Scholar] [CrossRef]
- Rossi, F.; Mancusi, S.; Bellini, G.; Roberti, D.; Punzo, F.; Vetrella, S.; Matarese, S.M.; Nobili, B.; Maione, S.; Perrotta, S. CNR2 functional variant (Q63R) influences childhood immune thrombocytopenic purpura. Haematologica 2011, 96, 1883–1885. [Google Scholar] [CrossRef] [Green Version]
- Sagnelli, C.; Uberti-Foppa, C.; Hasson, H.; Bellini, G.; Minichini, C.; Salpietro, S.; Messina, E.; Barbanotti, D.; Merli, M.; Punzo, F.; et al. Cannabinoid receptor 2–63 RR variant is independently associated with severe necroinflammation in HIV/HCV coinfected patients. PLoS ONE 2017, 12, e0181890. [Google Scholar] [CrossRef] [Green Version]
- Murikinati, S.; Jüttler, E.; Keinert, T.; Ridder, D.A.; Muhammad, S.; Waibler, Z.; Ledent, C.; Zimmer, A.; Kalinke, U.; Schwaninger, M. Activation of cannabinoid 2 receptors protects against cerebral ischemia by inhibiting neutrophil recruitment. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 788–798. [Google Scholar] [CrossRef] [Green Version]
- Fraga, D.; Raborn, E.S.; Ferreira, G.A.; Cabral, G.A. Cannabinoids inhibit migration of microglial-like cells to the HIV protein Tat. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2011, 6, 566–577. [Google Scholar] [CrossRef]
- Rock, R.B.; Gekker, G.; Hu, S.; Sheng, W.S.; Cabral, G.A.; Martin, B.R.; Peterson, P.K. WIN55,212–2-mediated inhibition of HIV-1 expression in microglial cells: Involvement of cannabinoid receptors. J. Neuroimmune Pharmacol. Off. J. Soc. on NeuroImmune Pharmacol. 2007, 2, 178–183. [Google Scholar] [CrossRef]
- Peterson, P.K.; Gekker, G.; Hu, S.; Cabral, G.; Lokensgard, J.R. Cannabinoids and morphine differentially affect HIV-1 expression in CD4(+) lymphocyte and microglial cell cultures. J. Neuroimmunol. 2004, 147, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Costantino, C.M.; Gupta, A.; Yewdall, A.W.; Dale, B.M.; Devi, L.A.; Chen, B.K. Cannabinoid Receptor 2-mediated Attenuation of CXCR4-tropic HIV Infection in Primary CD4+ T Cells. PLoS ONE 2012, 7, e33961. [Google Scholar]
- Hu, S.; Sheng, W.S.; Rock, R.B. Immunomodulatory properties of kappa opioids and synthetic cannabinoids in HIV-1 neuropathogenesis. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2011, 6, 528–539. [Google Scholar] [CrossRef]
- Gamal, M.; Moawad, J.; Rashed, L.; El-Eraky, W.; Saleh, D.; Lehmann, C.; Sharawy, N. Evaluation of the effects of Eserine and JWH-133 on brain dysfunction associated with experimental endotoxemia. J. Neuroimmunol. 2015, 281, 9–16. [Google Scholar] [CrossRef]
- Zhou, J.; Burkovskiy, I.; Yang, H.; Sardinha, J.; Lehmann, C. CB2 and GPR55 Receptors as Therapeutic Targets for Systemic Immune Dysregulation. Front. Pharmacol. 2016, 7, 264. [Google Scholar] [CrossRef] [Green Version]
- Plastina, P.; Apriantini, A.; Meijerink, J.; Witkamp, R.; Gabriele, B.; Fazio, A. In Vitro Anti-Inflammatory and Radical Scavenging Properties of Chinotto (Citrus myrtifolia Raf.). Essent. Oils. Nutr. 2018, 10, 783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Hsouna, A.; Gargouri, M.; Dhifi, W.; Ben Saad, R.; Sayahi, N.; Mnif, W.; Saibi, W. Potential anti-inflammatory and antioxidant effects of Citrus aurantium essential oil against carbon tetrachloride-mediated hepatotoxicity: A biochemical, molecular and histopathological changes in adult rats. Environ. Toxicol. 2019, 34, 388–400. [Google Scholar] [CrossRef]
- Lone, J.; Yun, J.W. Monoterpene limonene induces brown fat-like phenotype in 3T3-L1 white adipocytes. Life Sci. 2016, 153, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Pesce, M.; Seguella, L.; Sanseverino, W.; Lu, J.; Corpetti, C.; Sarnelli, G. The potential of cannabidiol in the COVID-19 pandemic. Br. J. Pharmacol. 2020, 177, 4967–4970. [Google Scholar] [CrossRef]
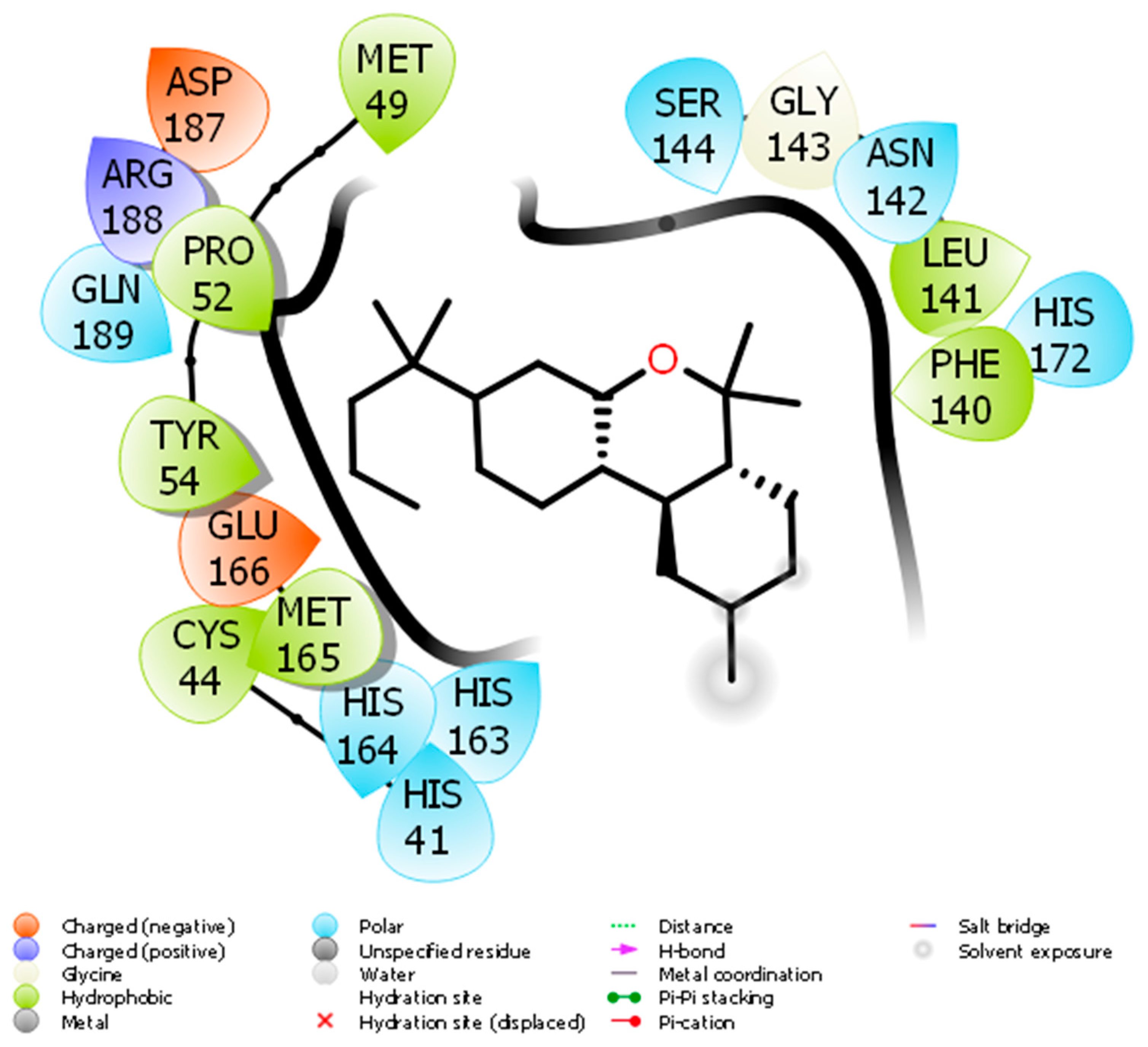
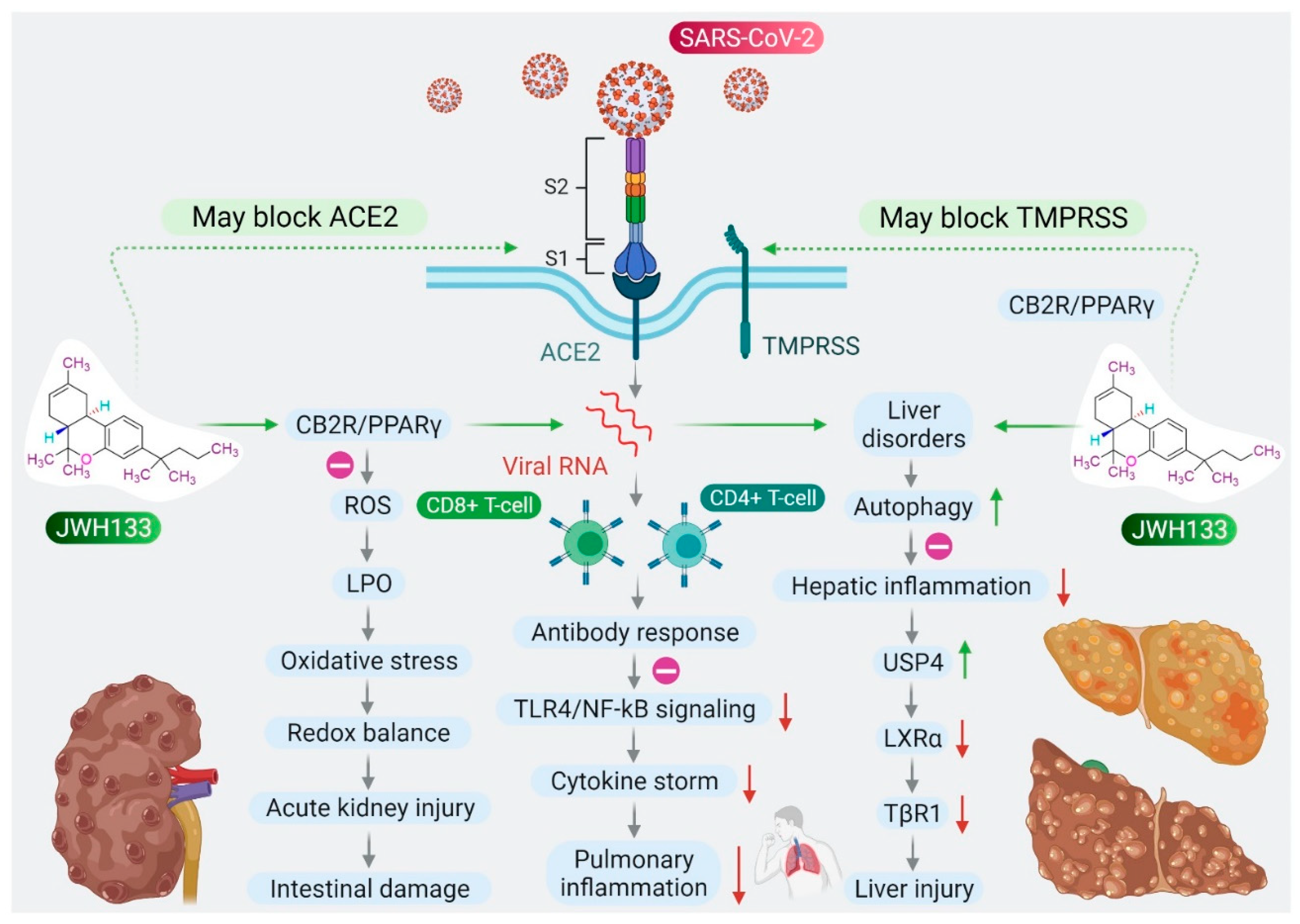
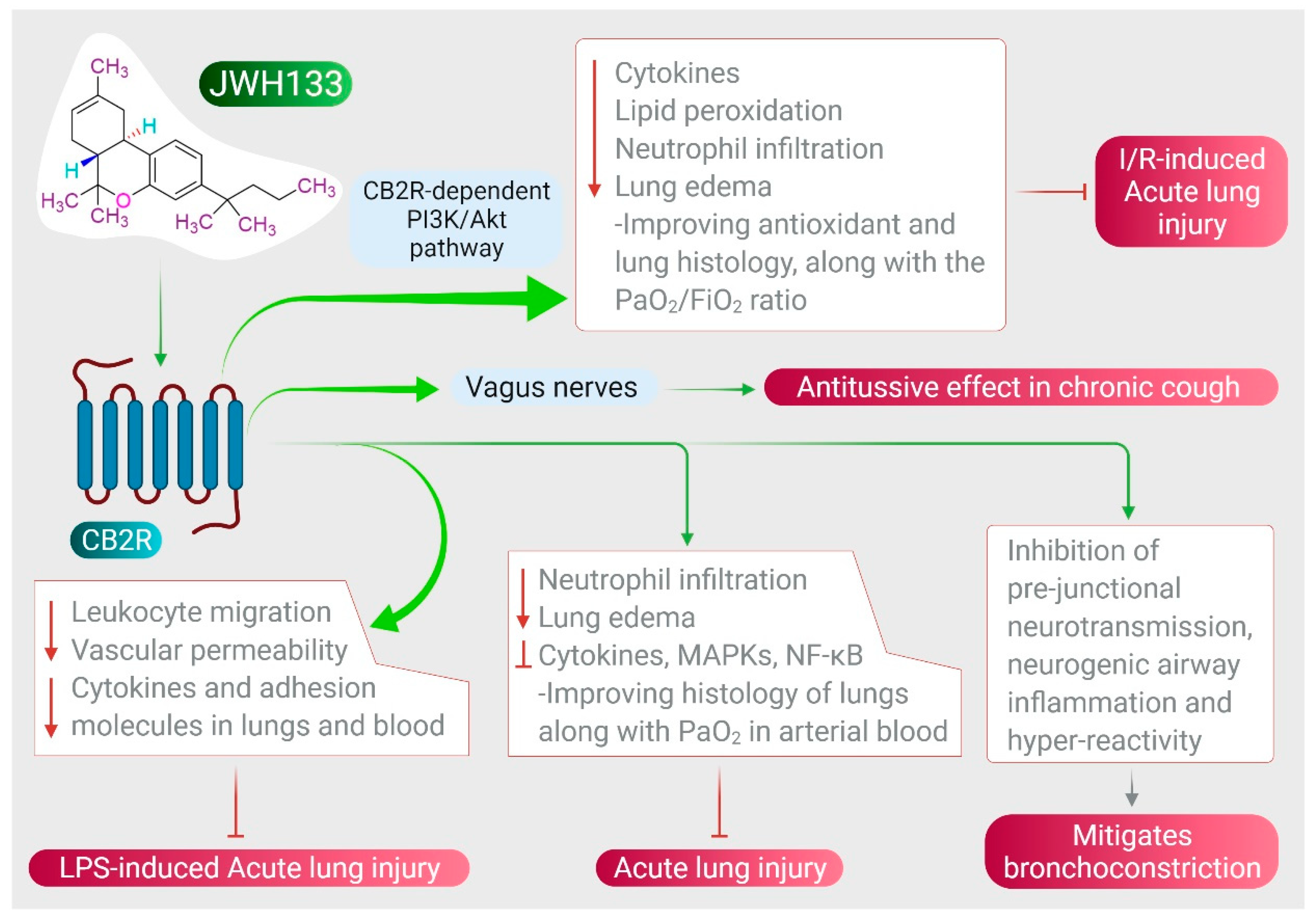
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).