
Investigation of Spatiotemporal Patterns of Harbour Porpoise (Phocoena phocoena) Strandings in Swedish Waters for Improved Monitoring and Management


Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection and Study Area
2.2. Stranding Data, Necropsy Data, and Processing
2.3. Seasonality and Cluster Identification
3. Results
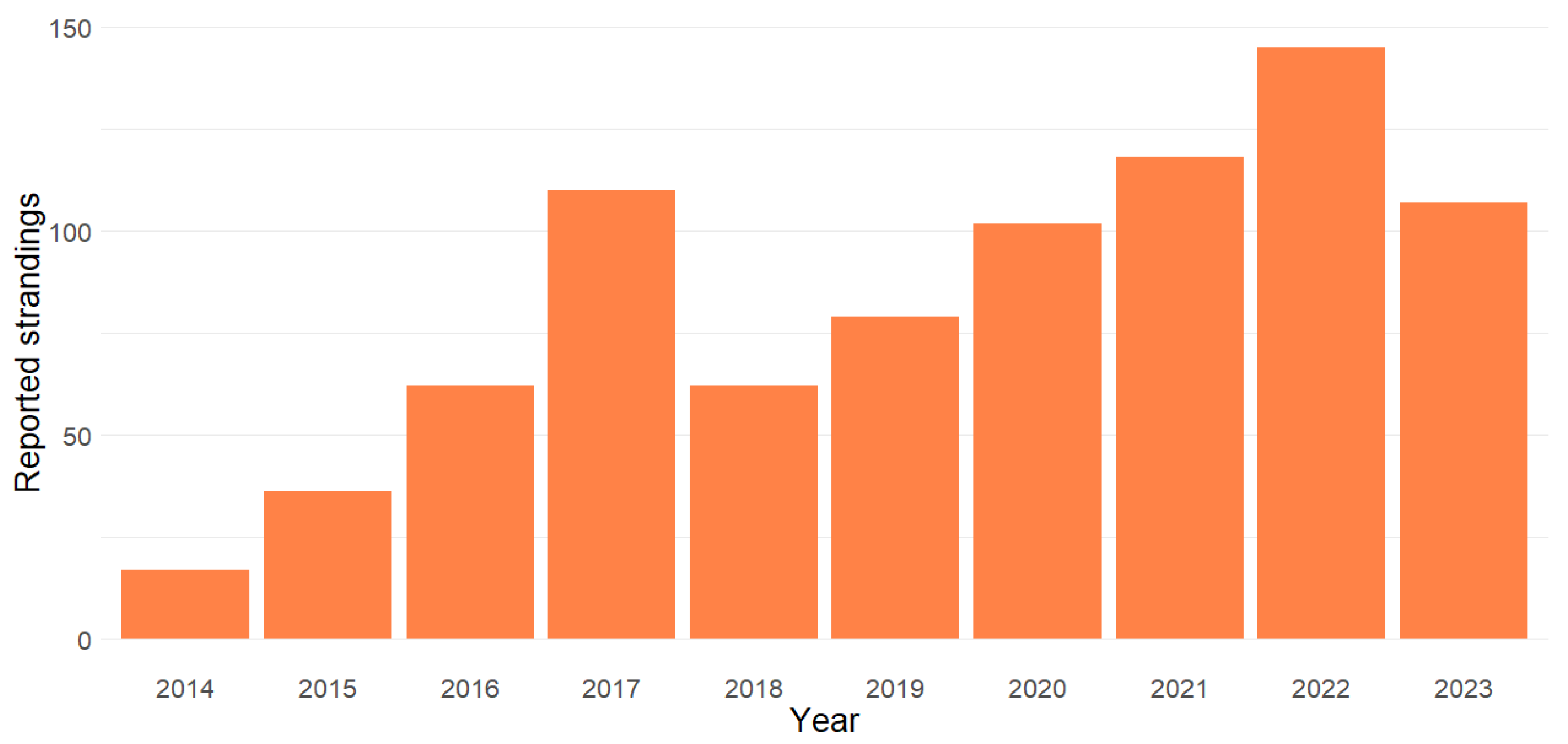
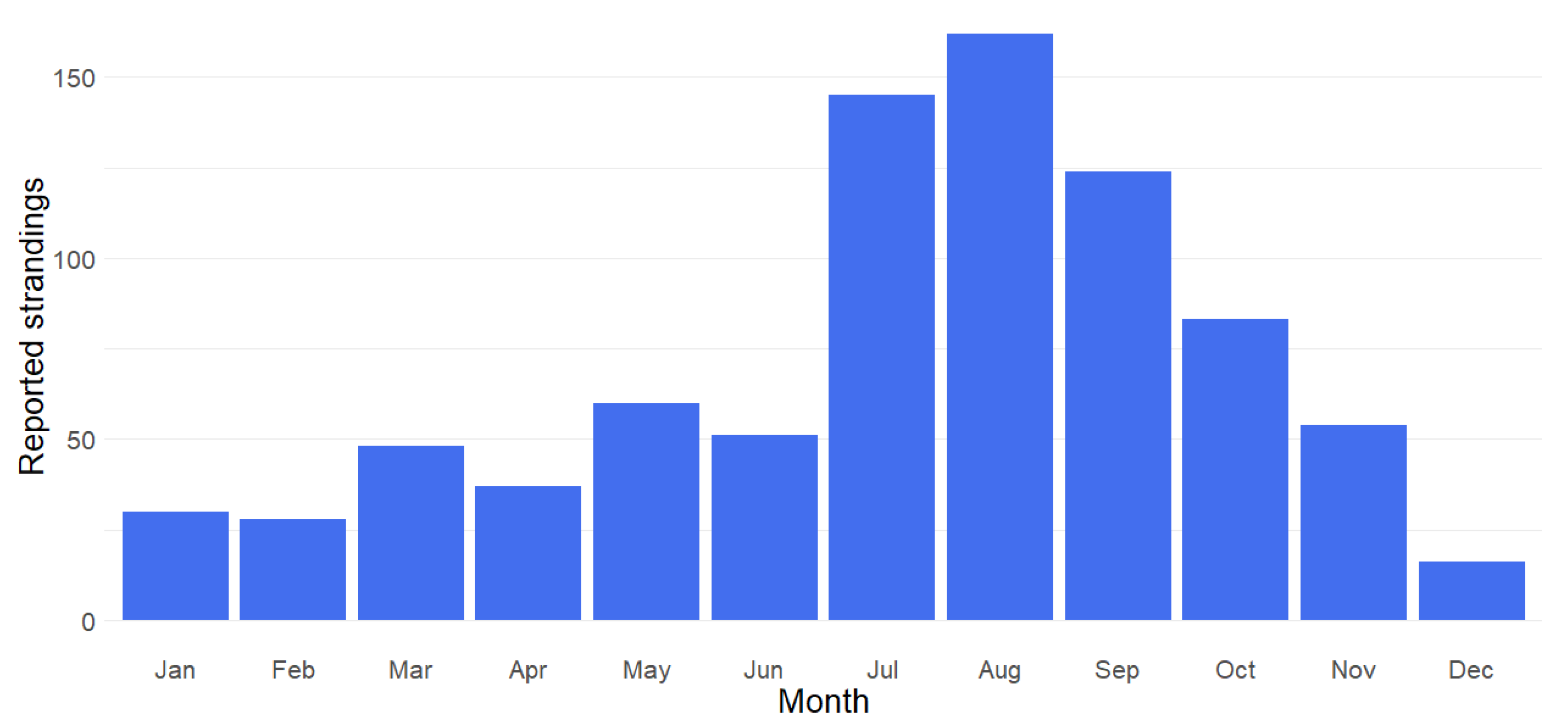
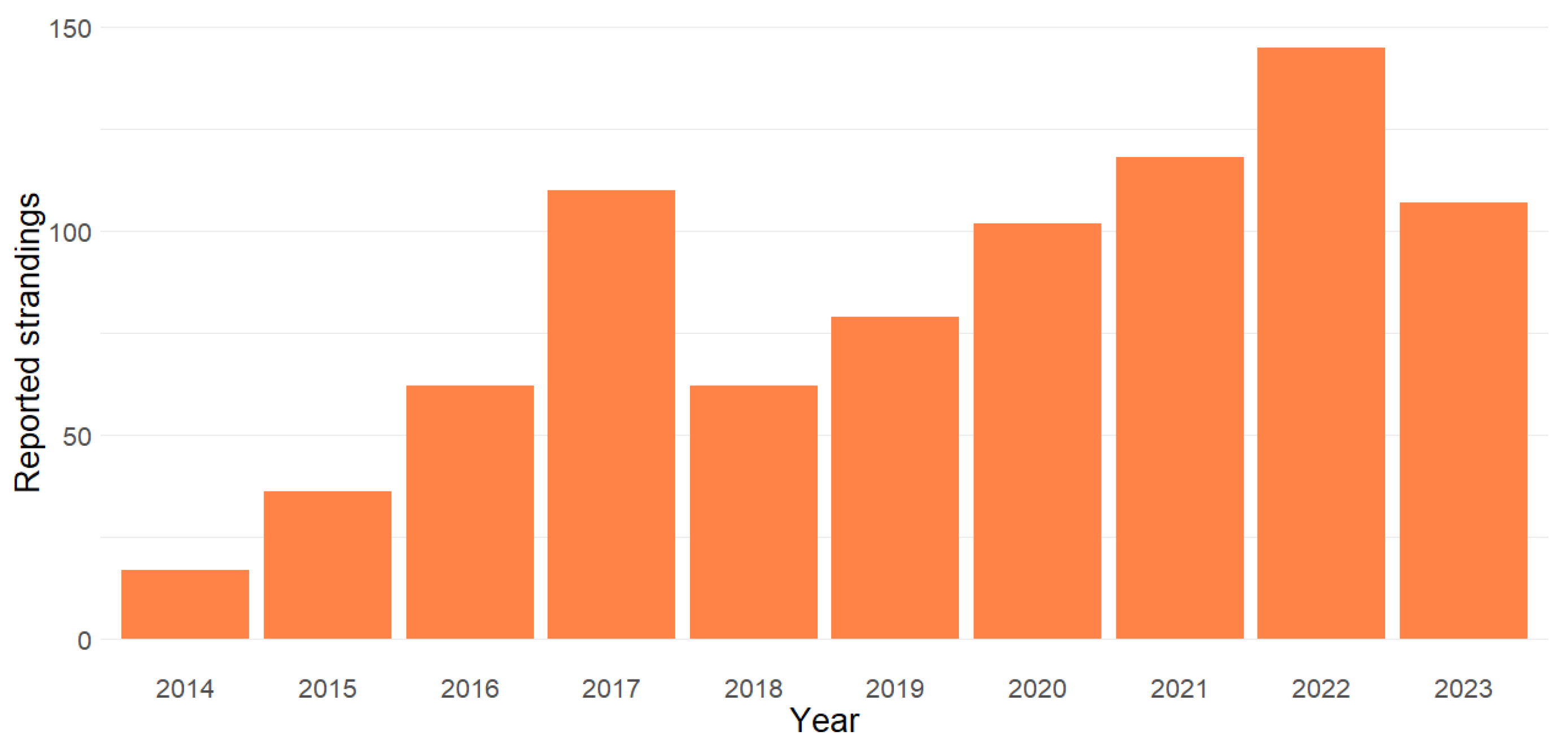
3.1. Seasonality and Temporal Trends in Stranding Reports
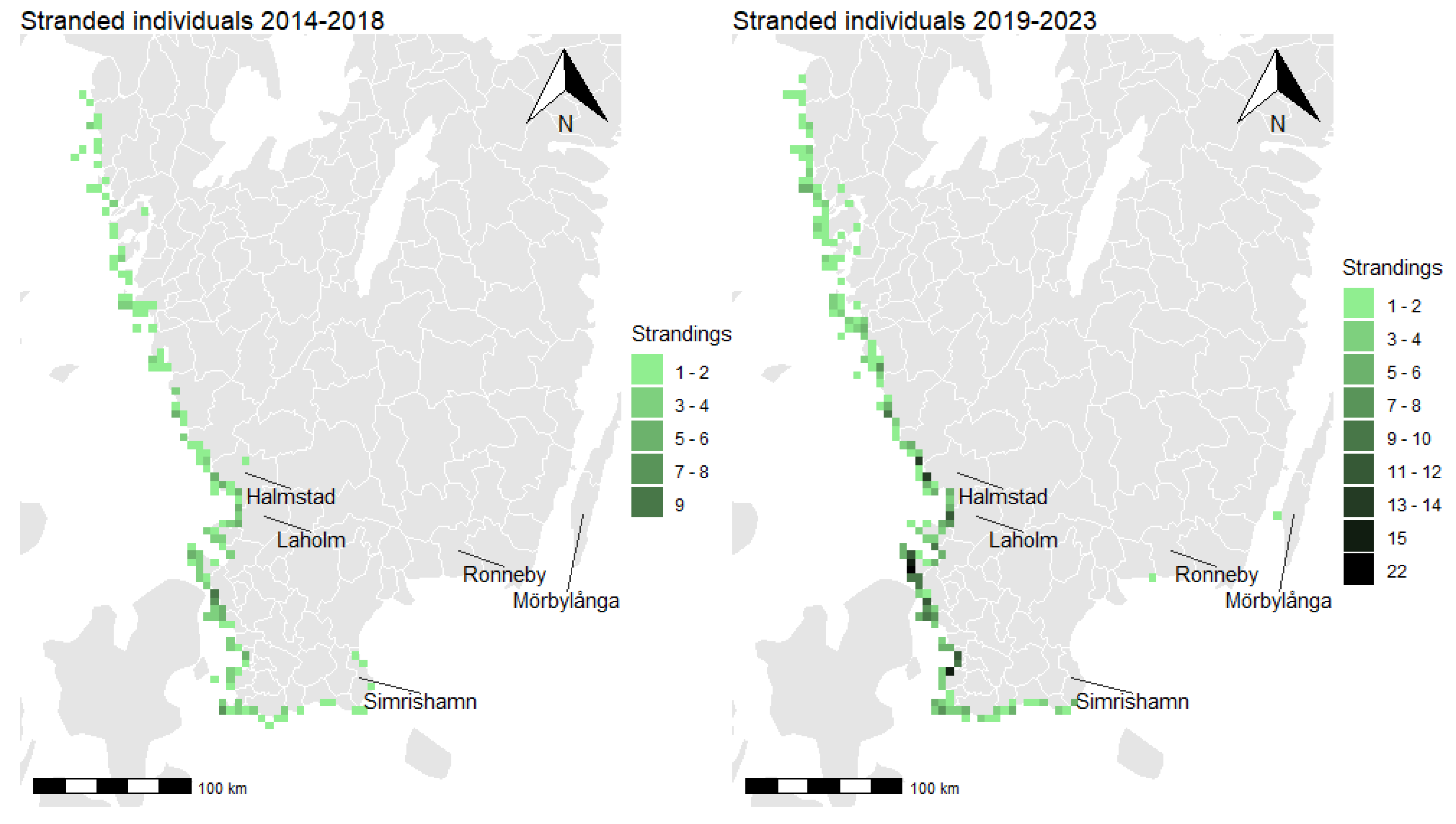
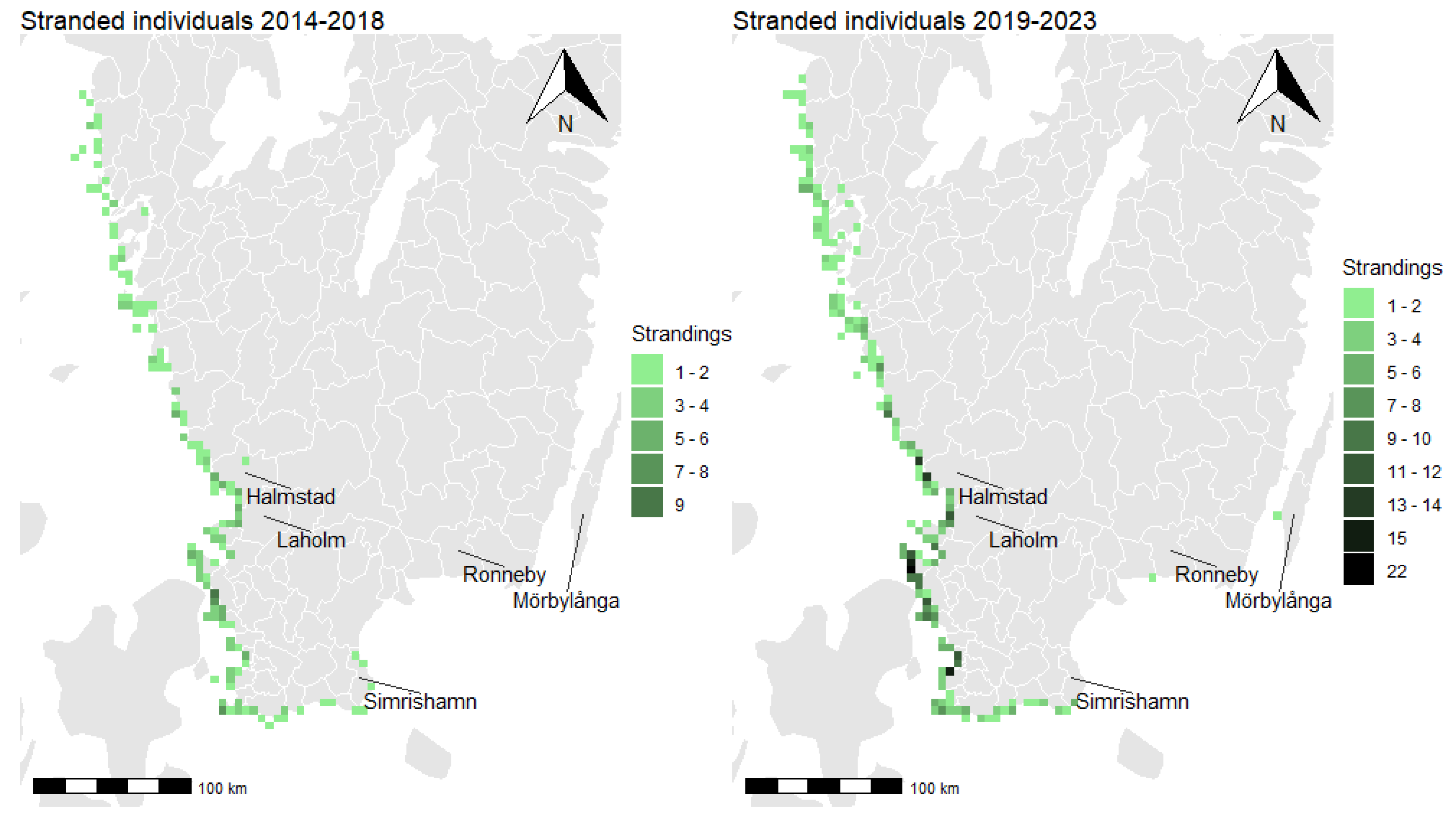
3.2. Distribution of Total Strandings
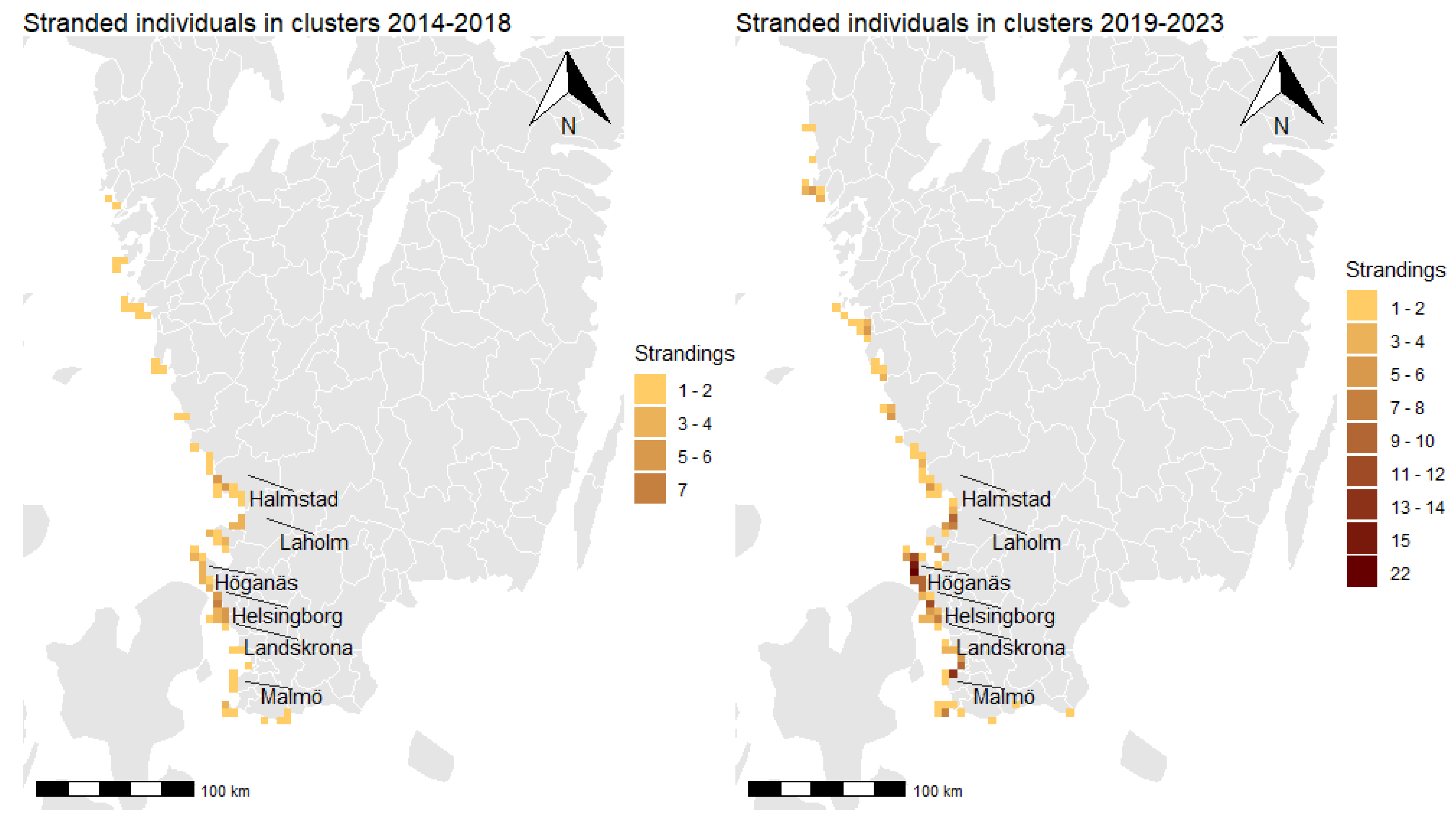
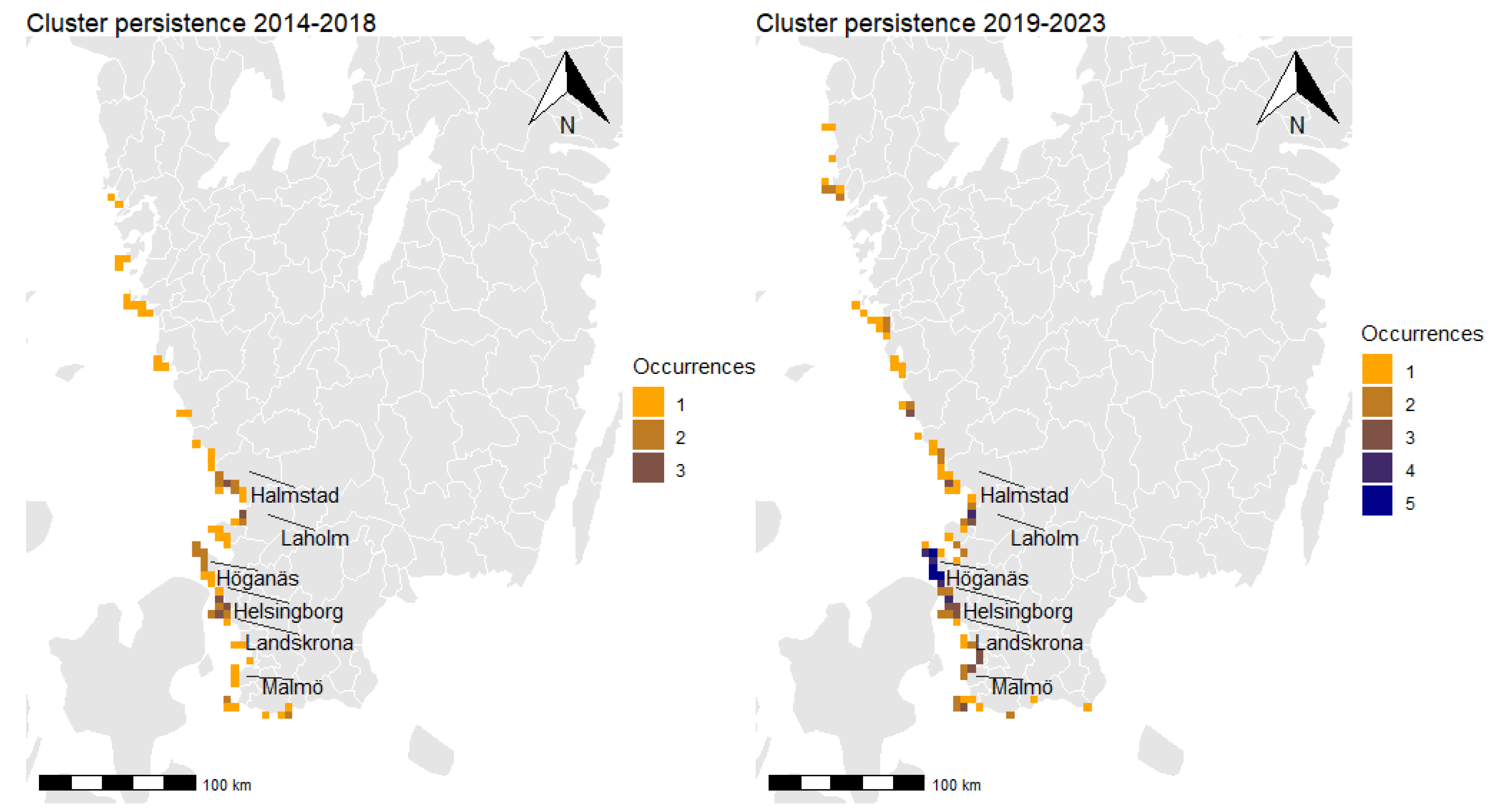
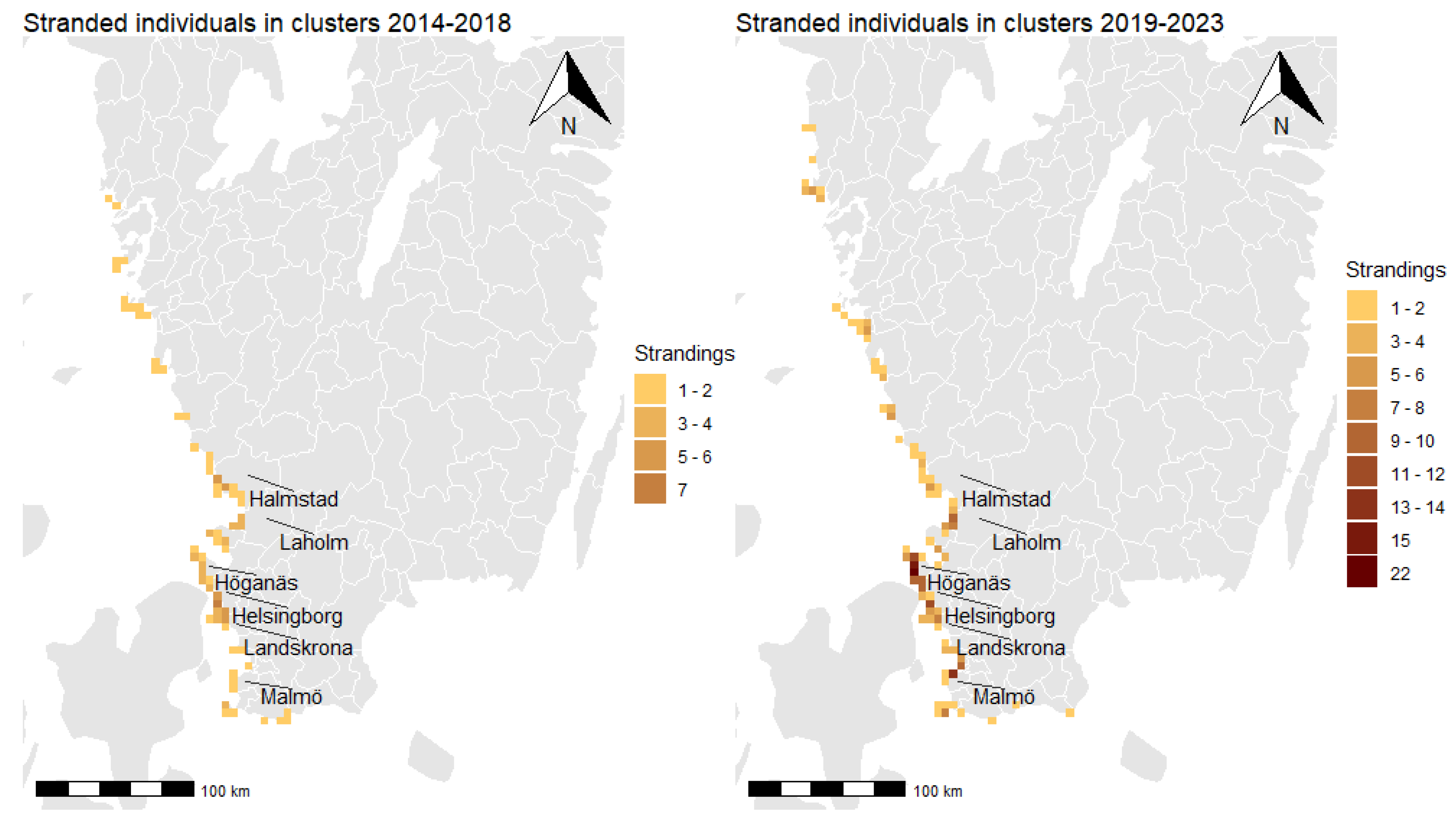
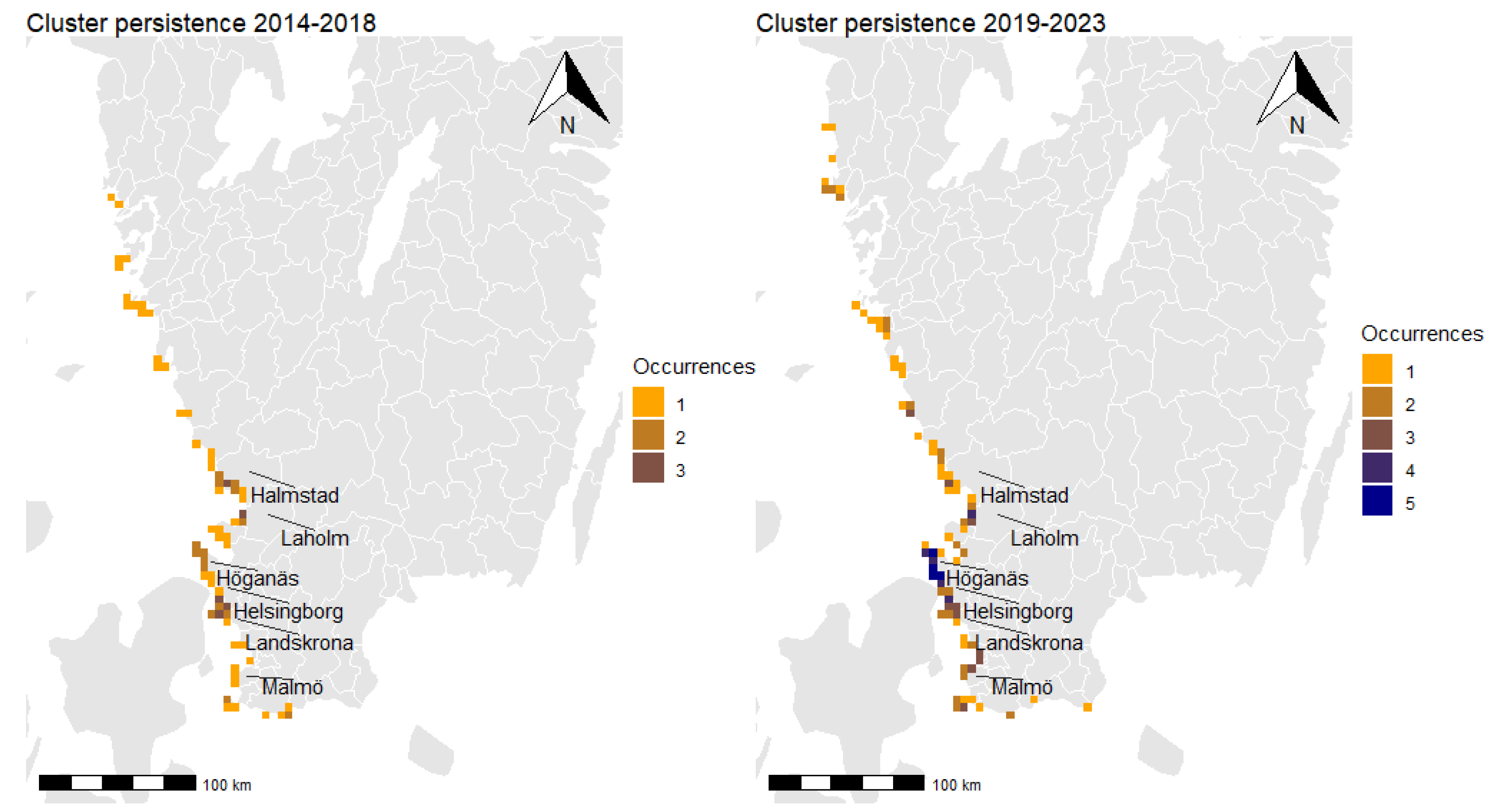
3.3. Spatial Clusters and Cluster Persistence
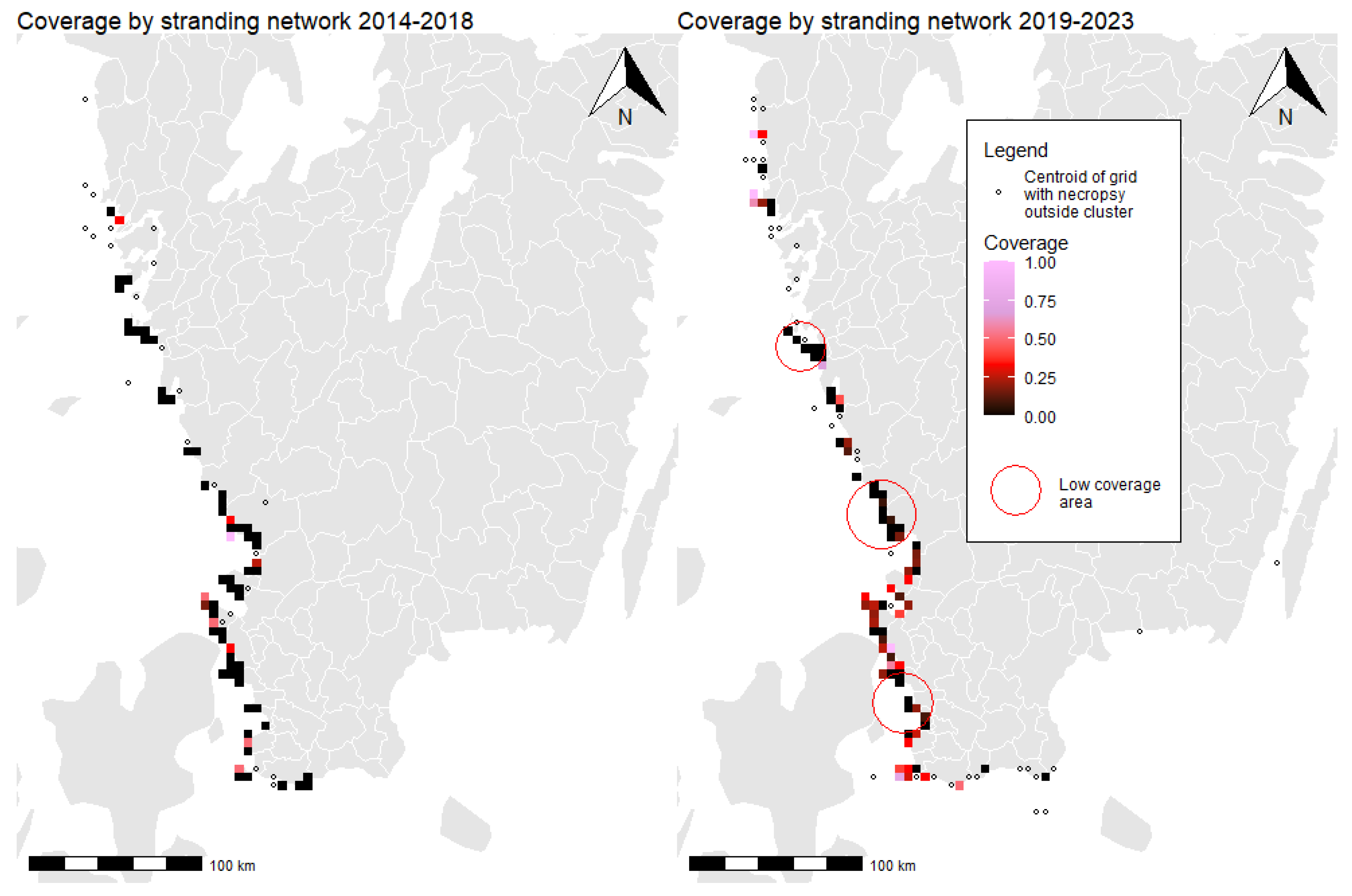
3.4. Stranding Network Coverage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neimanis, A.; Stavenow, J.; Ågren, E.O.; Wikström-Lassa, E.; Roos, A.M. Causes of Death and Pathological Findings in Stranded Harbour Porpoises (Phocoena phocoena) from Swedish Waters. Animals 2022, 12, 369. [Google Scholar] [CrossRef] [PubMed]
- Braulik, G.T.; Minton, G.; Amano, M.; Bjørge, A. Phocoena phocoena (amended version of 2020 assessment). IUCN Red List Threat. Species 2023, 1, e.T17027A247632759. [Google Scholar] [CrossRef]
- Sharpe, M.; Berggren, P. Phocoena phocoena (Europe assessment). IUCN Red List Threat. Species 2023, 1, e.T17027A219010660. Available online: https://www.iucnredlist.org/species/17027/219010660 (accessed on 16 January 2024).
- Wiemann, A.; Andersen, L.W.; Berggren, P.; Siebert, U.; Benke, H.; Teilmann, H.; Lockyer, C.; Pawliczka, I.; Skóra, K.; Roos, A.; et al. Mitochondrial Control Region and Microsatellite Analyses on Harbour Porpoise (Phocoena phocoena) Unravel Population Differentiation in the Baltic Sea and Adjacent Water. Conserv. Genet. 2010, 11, 195–211. [Google Scholar] [CrossRef]
- Galatius, A.; Kinze, C.; Teilmann, J. Population Structure of Harbour Porpoises in the Batlic Region: Evidence of Separation Based on Geometric Morphometric Comparisons. J. Mar. Biol. Assoc. U. K. 2012, 92, 1669–1676. [Google Scholar] [CrossRef]
- Carlström, J.; Carlén, I.; Dähne, M.; Hammond, P.S.; Koschinski, S.; Owen, K.; Sveegaard, S.; Tiedemann, R. Phocoena phocoena (Baltic Sea subpopulation). IUCN Red List Threat. Species 2023, 1, e.T17031A50370773. Available online: https://www.iucnredlist.org/species/17031/50370773 (accessed on 16 January 2024).
- Gilles, A.; Authier, M.; Ramirez-Martinez, N.C.; Araújo, H.; Blanchard, A.; Carlström, J.; Eira, C.; Dorémus, G.; Fernández-Maldonado, C.; Geelhoed, S.C.V.; et al. Estimates of Cetacean Abundance in European Atlantic Waters in Summer 2022 from the SCANS-IV Aerial and Shipboard Surveys. 2023; p. 64. Available online: https://www.tiho-hannover.de/itaw/scans-iv-survey (accessed on 16 January 2024).
- Amundin, M.; Carlström, J.; Thomas, L.; Carlén, I.; Koblitz, J.; Teilmann, J.; Tougaard, J.; Tregenza, N.; Wennerberg, D.; Loisa, O.; et al. Estimating the abundance of the critically endangered Baltic Proper harbour porpoise (Phocoena phocoena) population using passive acoustic monitoring. Ecol. Evol. 2022, 12, e8854. [Google Scholar] [CrossRef] [PubMed]
- Viquerat, S.; Herr, H.; Gilles, A.; Peschko, V.; Siebert, U.; Sveegaard, S.; Teilmann, J. Abundance of harbour porpoises (Phocoena phocoena) in the western Baltic, Belt Seas and Kattegat. Mar. Biol. 2014, 161, 745–754. [Google Scholar] [CrossRef]
- Hammond, P.S.; Lacey, C.; Gilles, A.; Viquerat, S.; Börjesson, P.; Herr, H.; Macelod, K.; Ridoux, V.; Santos, M.B.; Teilmann, J.; et al. Estimates of Cetacean Abundance in European Atlantic Waters in Summer 2016 from the SCANS-III Aerial and Shipboard Surveys. 2021. Available online: https://scans3.wp.st-andrews.ac.uk/files/2021/06/SCANS-III_design-based_estimates_final_report_revised_June_2021.pdf (accessed on 16 January 2024).
- Swedish Agency for Marine and Water Management. Åtgärdsprogram för Tumlare. 2021: 11. Available online: https://www.havochvatten.se/download/18.764c2cb917a6c5c85b03c106/1625648670494/rapport-2021-atgardsprogram-for-tumlare-v2.pdf (accessed on 29 November 2023).
- Berggren, P. Bycatches of the harbour porpoise (Phocoena phocoena) in the Swedish Skagerrak, Kattegat and Baltic Seas 1973–1993. Rep. Int. Whale. Comm. 1994, 15, 211–215. [Google Scholar]
- Vinther, M. Bycatches of harbour porpoises (Phocoena phocoena L.) in Danish set-net fisheries. J. Cetacean Res. Manag. 1999, 1, 123–135. [Google Scholar] [CrossRef]
- Berggren, P.; Wade, P.R.; Carlström, J.; Read, A.J. Potential limits to anthropogenic mortality for harbour porpoises in the Baltic region. Biol. Conserv. 2002, 103, 313–322. [Google Scholar] [CrossRef]
- Vinther, M.; Larsen, F. Updated estimates of harbour porpoise (Phocoena phocoena) bycatch in the Danish North Sea bottom-set gillnet fishery. J. Cetacean Res. Manag. 2004, 6, 12–24. [Google Scholar] [CrossRef]
- HELCOM. HELCOM Red List of Baltic Sea Species in Danger of Becoming Extinct; Baltic Sea Environment Proceedings No. 140; Helsinki Commission: Helsinki, Finland, 2013.
- Kindt-Larsen, L.; Glemarec, G.; Berg, C.W.; Königson, S.; Kroner, A.; Søgaard, M.; Lusseau, D. Knowing the fishery to know the bycatch: Bias-corrected estimates of harbour porpoise bycatch in gillnet fisheries. Proc. R. Soc. B 2023, 290, 20222570. [Google Scholar] [CrossRef]
- Glemarec, G.; Königson, S.; Kindt-Larsen, L. Bycatch in Baltic Sea Commercial Fisheries: High-Risk Areas and Evaluation of Measures to Reduce Bycatch. HELCOM ACTION. 2021. Available online: https://helcom.fi/wp-content/uploads/2021/11/Bycatch-in-Baltic-Sea-commercial-fisheries.pdf (accessed on 26 January 2024).
- Regulation 57/2011. Council Regulation (EU) No 57/2011 of 18 January 2011 Fixing for 2011 the Fishing Opportunities for Certain Fish Stocks and Groups of Fish Stocks, Applicable in EU Waters and, for EU Vessels, in Certain Non-EU Waters. Available online: http://data.europa.eu/eli/reg/2011/57/2011-08-04 (accessed on 17 January 2024).
- Swedish Agency of Marine and Water Management. Begränsat Fritidsfiske Efter Pigghaj Möjligt. Available online: https://www.havochvatten.se/arkiv/aktuellt/2023-06-02-begransat-fritidsfiske-efter-pigghaj-mojligt.html (accessed on 7 December 2023).
- Bryhn, A.C.; Bergek, S.; Bergström, U.; Casini, M.; Dahlgren, E.; Ek, C.; Hjelm, J.; Königson, S.; Ljungberg, P.; Lundström, K.; et al. Which factors can affect the productivity of cod stocks in the Baltic Sea, Kattegat and Skagerrak? Ocean Coast Manag. 2022, 223, 106154. [Google Scholar] [CrossRef]
- Siebert, U.; Gilles, A.; Lucke, K.; Ludwig, M.; Benke, H.; Kock, K.; Scheidat, M. A decade of harbour porpoise occurrence in German waters–Analyses of aerial surveys, incidental sightings and strandings. J. Sea Res. 2006, 56, 65–80. [Google Scholar] [CrossRef]
- Directive 92/43/EEC. Council Directive (EU) 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. Available online: http://data.europa.eu/eli/dir/1992/43/oj (accessed on 17 January 2024).
- OSPAR Commission. Convention for the Protection of the Marine Environment of the North-East Atlantic; OSPAR Commission: London, UK, 1992. [Google Scholar]
- HELCOM. Convention on the Protection of the Marine Environment of the Baltic Sea Area; Helsinki Commission: Helsinki, Finland, 1992.
- ASCOBANS. Agreement on the Conservation of Small Cetaceans of the Baltic, North East Atlantic, Irish and North Seas; ASCOBANS: Bonn, Germany, 1991. [Google Scholar]
- Moore, S.E. Marine Mammals as Ecosystem Sentinels. J. Mammal. 2008, 89, 534–540. [Google Scholar] [CrossRef]
- Bossart, G.D. Marine Mammals as Sentinel Species for Oceans and Human Health. Vet. Pathol. 2011, 48, 676–690. [Google Scholar] [CrossRef] [PubMed]
- Kiszka, J.; Heithaus, M.; Wirsing, A. Behavioural drivers of the ecological roles and importance of marine mammals. Mar. Ecol. Prog. 2015, 523, 267–281. [Google Scholar] [CrossRef]
- Peltier, H.; Dabin, W.; Daniel, P.; Van Canneyt, O.; Dorémus, G.; Huon, M.; Ridoux, V. The significance of stranding data as indicators of cetacean populations at sea: Modelling the drift of cetacean carcasses. Ecol. Indic. 2012, 18, 278–290. [Google Scholar] [CrossRef]
- Wund, S.; Méheust, E.; Dars, C.; Dabin, W.; Demaret, F.; Guichard, B.; Jauniaux, T.; Labrut, S.; Spitz, J.; Van Canneyt, O.; et al. Strengthening the health surveillance of marine mammals in the waters of metropolitan France by monitoring strandings. Front. Mar. Sci. 2023, 10, 1116819. [Google Scholar] [CrossRef]
- McGovern, B.; Gulloch, R.M.; O’Connell, M.; Berrow, S. Temporal and spatial trends in stranding records of cetaceans on the Irish coast, 2002–2014. J. Mar. Biol. Assoc. U. K. 2018, 98, 977–989. [Google Scholar] [CrossRef]
- IJsseldijk, L.L.; ten Doeschate, M.T.I.; Brownlow, A.; Davison, N.J.; Deaville, R.; Galatius, A.; Gilles, A.; Haelters, J.; Jepson, P.D.; Keijl, G.O.; et al. Spatiotemporal mortality and demographic trends in a small cetacean: Strandings to inform conservation management. Biol. Conserv. 2020, 249, 108733. [Google Scholar] [CrossRef]
- Rapportera Tumlare Och Sälar. Available online: https://marinadaggdjur.nrm.se/ (accessed on 31 October 2023).
- Vår Övervakning av Marina Däggdjur. Available online: https://www.sva.se/vilda-djur/marina-daggdjur/var-overvakning-av-marina-daggdjur/ (accessed on 7 November 2023).
- Open Data, Examinations of Cetaceans in Swedish Waters, 2020–2022. Available online: https://www.dataportal.se/en/datasets/59_2060/open-data-examinations-of-cetaceans-in-swedish-waters-2020-2022#ref=?p=1&q=marine%20mammal&s=2&t=20&f=&rt=dataset%24esterms_IndependentDataService%24esterms_ServedByDataService&c=false (accessed on 27 November 2023).
- Topografi 250 Nedladdning, Vector. Available online: https://geotorget.lantmateriet.se/geodataprodukter/topografi-250-nedladdning-vektor (accessed on 17 November 2023).
- Anselin, L. Local Indicators of Spatial Association–LISA. Geogr. Anal. 1995, 27, 93–115. [Google Scholar] [CrossRef]
- Bivand, R.; Wong, D. Comparing implementations of global and local indicators of spatial association. TEST 2018, 27, 718–748. [Google Scholar] [CrossRef]
- Wilcoxon, F. Individual Comparisons by Ranking Methods. Biometrics 1945, 1, 80–83. [Google Scholar] [CrossRef]
- Mann, H.B.; Whitney, D.R. On a Test of Whether one of Two Random Variables is Stochastically Larger than the Other. Ann. Math. Stat. 1947, 18, 50–60. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- South, A. rworldmap: A new R package for Mapping Global Data. R J. 2011, 3, 35–43. [Google Scholar] [CrossRef]
- Dunnington, D. ggspatial: Spatial Data Framework for ggplot2. 2023. Available online: https://paleolimbot.github.io/ggspatial/ (accessed on 15 January 2024).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 15 January 2024).
- Unger, B.; Nachtsheim, D.; Martínez, N.R.; Siebert, U.; Kyhn, L.A.; Balle, J.D.; Teilmann, J.; Calrström, J.; Owen, K.; Gilles, A. MiniSCANS-II: Aerial Survey for Harboir Porpoises in the Western Baltic Sea, Belt Sea, the Sound and Kattegat in 2020; Danish Environmental Protection Agency: Odense, Denmark; German Federal Agency for Nature: Bonn, Germany; Conservation and Swedish Agency for Marine and Water Management: Göteborg, Sweden, 2021; p. 28.
- Authier, M.; Peltier, H.; Dorémus, G.; Dabin, W.; Van Canneyt, O.; Ridoux, V. How much are strandings records affected by variation in reporting rates? A case study of small delphinids in the Bay of Biscay. Biodivers. Conserv. 2014, 23, 2591–2612. [Google Scholar] [CrossRef]
- Lockyer, C.; Kinze, C. Status, ecology and life history of harbour porpoise (Phocoena phocoena), in Danish waters. NAMMCO Sci. Publ. 2003, 5, 143–175. [Google Scholar] [CrossRef]
- Peltier, H.; Baagøe, H.J.; Camphuysen, K.C.J.; Czeck, R.; Dabin, W.; Daniel, P.; Deaville, R.; Haelters, J.; Jauniaux, T.; Jensen, L.F.; et al. The stranding anomaly as population indicator: The case of harbour porpoise Phocoena phocoena in North-Western Europe. PLoS ONE 2013, 8, e62180. [Google Scholar] [CrossRef]
- IJsseldijk, L.L.; Camphuysen, K.C.J.; Keijl, G.O.; Troost, G.; Aarts, G. Predicting harbour porpoise strandings based on near-shore sightings indicates elevated temporal mobility rates. Front. Mar. Sci. 2021, 8, 668038. [Google Scholar] [CrossRef]
- Vishnyakova, K.; Gol’din, P. Seasonality of strandings and bycatch of harbour porpoises in the Sea of Azov: The effects of fisheries, weather conditions, and life history. ICES Mar. Sci. 2014, 72, 981–991. [Google Scholar] [CrossRef]
- Börjesson, P.; Read, A.J. Variation in timing of conception between populations of the harbour porpoise. J. Mammal. 2003, 84, 948–955. [Google Scholar] [CrossRef]
- Woodley, T.H.; Read, A.J. Potential rates of increase of a harbour porpoise (Phocoena phocoena) population subjected to incidental mortality in commercial fisheries. Can. J. Fish. Aquat. Sci. 1991, 48, 2429–2435. [Google Scholar] [CrossRef]
- Sveegaard, S.; Teilmann, J.; Tougaard, J.; Dietz, R.; Mouritsen, K.N.; Desportes, G.; Siebert, U. High-density areas for harbor porpoises (Phocoena phocoena) identified by satellite tracking. Mar. Mamm. Sci. 2011, 27, 230–246. [Google Scholar] [CrossRef]
- Sveegaard, S.; Teilmann, J.; Berggren, P.; Mouritsen, K.; Gillespie, D.; Tougaard, J. Acoustic surveys confirm the high-density areas of harbour porpoises found by satellite tracking. ICES J. Mar. Sci. 2011, 68, 929–939. [Google Scholar] [CrossRef]
- Berrow, S.D.; Whooley, P.; O’Connell, M.; Wall, D. Irish Cetacean Review (2000–2009); Irish Whale and Dolphin Group: Kilrush, Ireland, 2016; p. 60. [Google Scholar]
- Teilmann, J.; Dietz, R.; Sveegaard, S. The Use of Marine Waters of Skåne nu Harbour Porpoises in Time and Space; Technical Report No. 236; Aarhus University, DCE—Danish Centre for Environment and Energy: Roskilde, Denmark, 2022; p. 76. [Google Scholar]
- Sveegaard, S. Spatial and Temporal Distribution of Harbour Porpoises in Relation to Their Prey. Ph.D. Thesis, Aarhus University, Aarhus, Denmark, 2011. [Google Scholar]
- Sveegaard, S.; Andreasen, H.; Mouritsen, K.; Jeppesen, J.P.; Teilmann, J.; Kinze, C. Correlation between the seasonal distribution of harbour porpoises and their prey in the Sound, Baltic Sea. Mar. Biol. 2012, 159, 1029–1037. [Google Scholar] [CrossRef]
- ASCOBANS/ACCOBAMS. Best Practice on Cetacean Post Mortem Investigation and Tissue Sampling. 2019. Available online: https://www.ascobans.org/en/document/best-practice-cetacean-post-mortem-investigation-and-tissue-sampling (accessed on 26 January 2024).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | 2023 | Total Reported Stranded Individuals | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | 18 (1) | 37 (1) | 63 (1) | 111 (1) | 64 (2) | 81 (2) | 104 (2) | 120 (2) | 147 (2) | 109 (2) | 854 (16) |
| January | 0 | 5 | 1 | 4 (1) | 2 | 3 | 2 | 3 | 9 | 2 | 31 (1) |
| February | 0 | 1 | 5 | 1 | 1 | 2 | 3 | 0 | 11 | 4 | 28 |
| March | 3 | 1 | 1 | 3 | 1 | 11 | 2 | 14 | 9 | 3 | 48 |
| April | 3 | 1 | 1 | 10 | 2 | 2 (1) | 5 (1) | 5 | 5 | 5 | 39 (2) |
| May | 0 | 5 (1) | 3 (1) | 5 | 1 | 2 | 3 | 14 | 24 | 5 | 62 (2) |
| June | 0 | 3 | 1 | 13 | 6 (1) | 2 | 4 | 10 (1) | 10 (1) | 5 | 54 (3) |
| July | 3 | 2 | 13 | 11 | 11 | 12 | 22 (1) | 16 | 31 | 26 (1) | 147 (2) |
| August | 5 | 3 | 11 | 25 | 11 | 11 | 31 | 22 | 22 | 21 | 162 |
| September | 2 (1) | 3 | 16 | 11 | 16 | 16 | 15 | 18 (1) | 10 (1) | 20 | 127 (3) |
| October | 0 | 4 | 2 | 16 | 8 (1) | 16 (1) | 7 | 9 | 10 | 13 | 85 (2) |
| November | 2 | 7 | 8 | 9 | 2 | 4 | 6 | 8 | 5 | 3 | 54 |
| December | 0 | 2 | 1 | 3 | 3 | 0 | 4 | 1 | 1 | 2 (1) | 17 (1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulfsson, V.; Kim, H.; Cervin, L.; Roos, A.; Neimanis, A. Investigation of Spatiotemporal Patterns of Harbour Porpoise (Phocoena phocoena) Strandings in Swedish Waters for Improved Monitoring and Management. Oceans 2024, 5, 166-180. https://doi.org/10.3390/oceans5020010
Ulfsson V, Kim H, Cervin L, Roos A, Neimanis A. Investigation of Spatiotemporal Patterns of Harbour Porpoise (Phocoena phocoena) Strandings in Swedish Waters for Improved Monitoring and Management. Oceans. 2024; 5(2):166-180. https://doi.org/10.3390/oceans5020010
Chicago/Turabian StyleUlfsson, Vigge, Hyeyoung Kim, Linnea Cervin, Anna Roos, and Aleksija Neimanis. 2024. "Investigation of Spatiotemporal Patterns of Harbour Porpoise (Phocoena phocoena) Strandings in Swedish Waters for Improved Monitoring and Management" Oceans 5, no. 2: 166-180. https://doi.org/10.3390/oceans5020010
APA StyleUlfsson, V., Kim, H., Cervin, L., Roos, A., & Neimanis, A. (2024). Investigation of Spatiotemporal Patterns of Harbour Porpoise (Phocoena phocoena) Strandings in Swedish Waters for Improved Monitoring and Management. Oceans, 5(2), 166-180. https://doi.org/10.3390/oceans5020010