
Mastitis: What It Is, Current Diagnostics, and the Potential of Metabolomics to Identify New Predictive Biomarkers




Abstract
1. Introduction
2. Disease Etiology and Pathogenesis
2.1. Disease Etiology and Traditional Bacterial Infections
2.1.1. Staphylococcus aureus
2.1.2. Escherichia coli
2.1.3. Streptococcus uberis
2.1.4. Klebsiella spp.
2.1.5. Opportunistic Pathogens
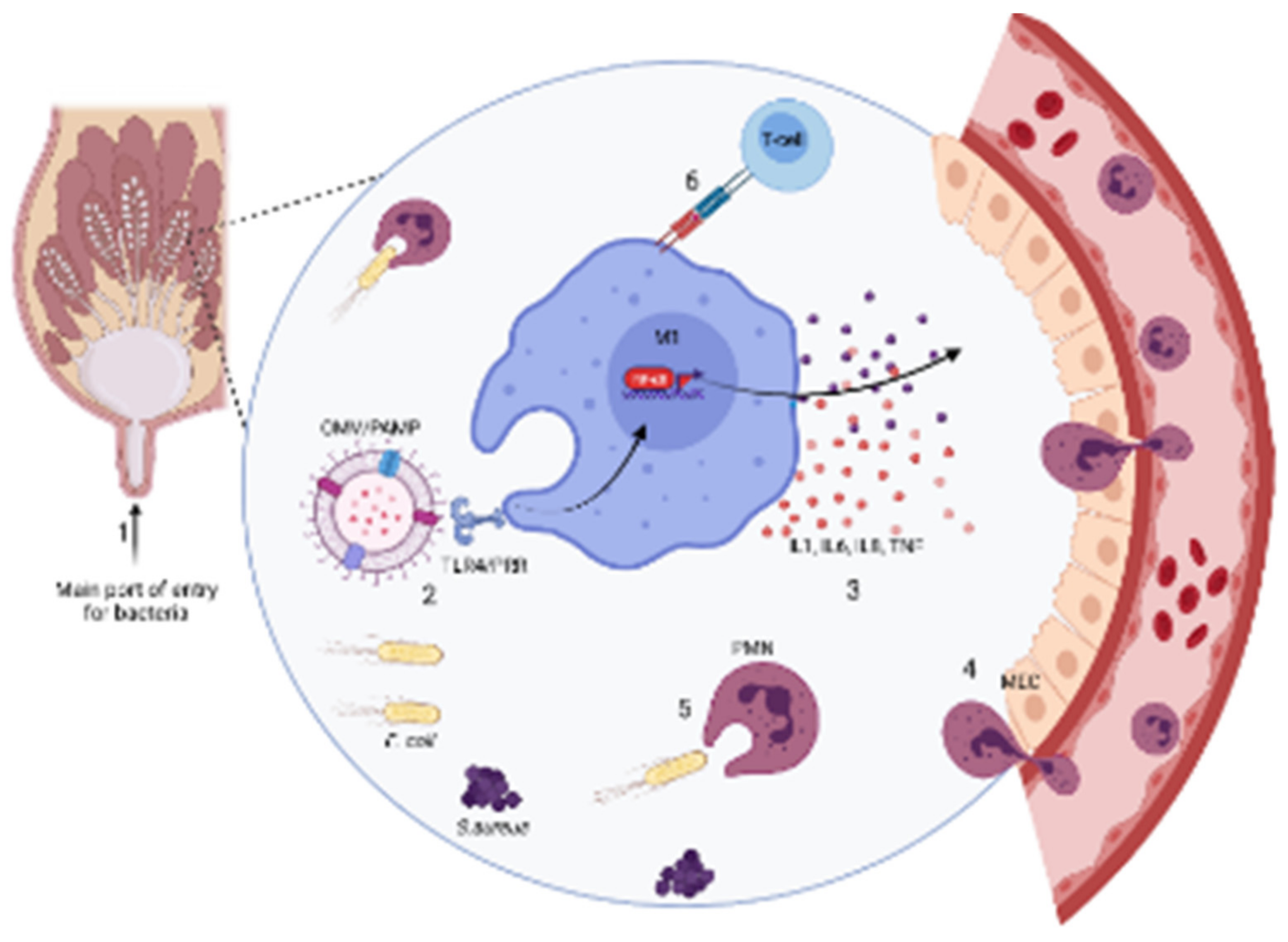
2.2. Pathogenesis
2.2.1. Clinical Mastitis
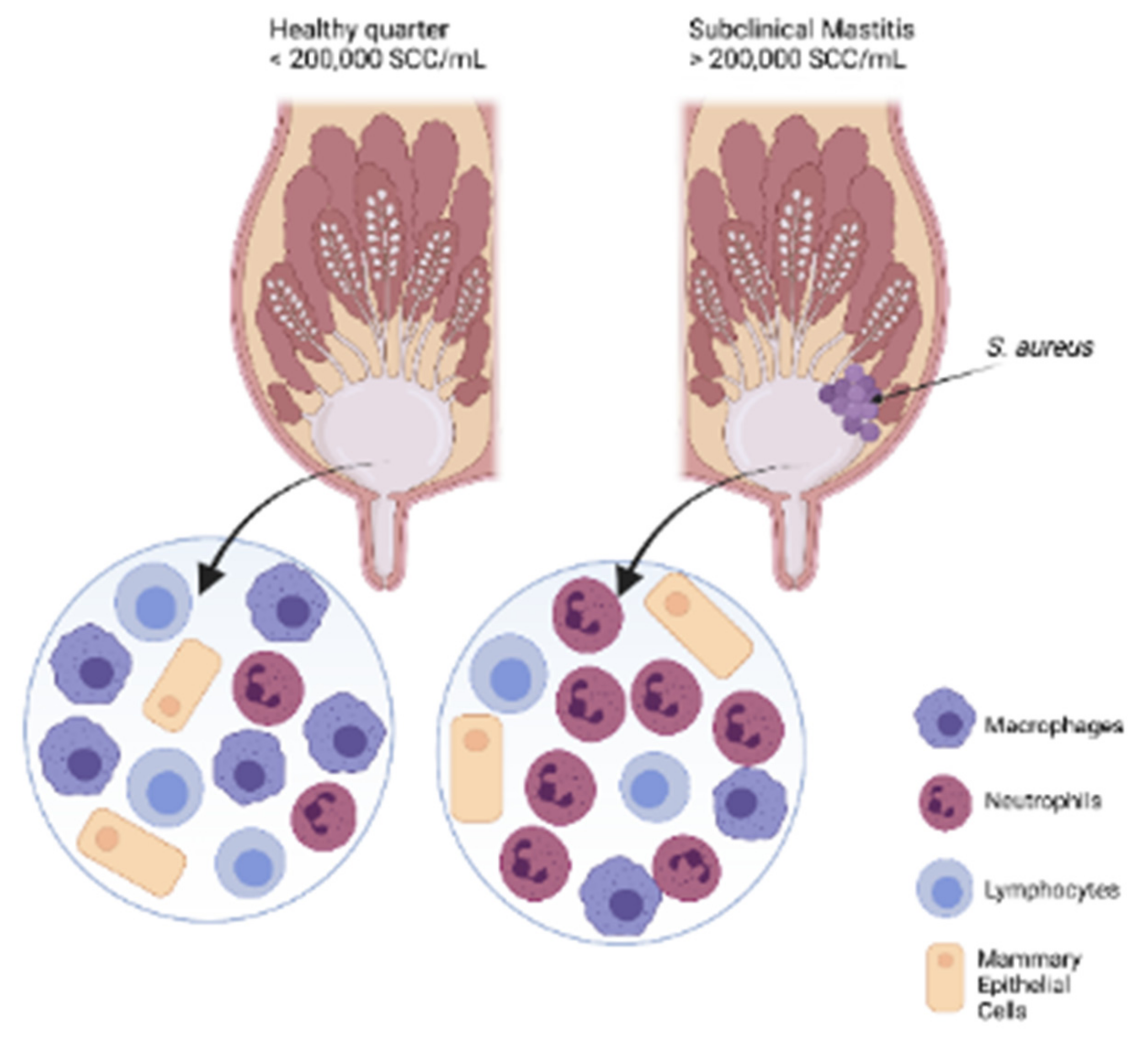
2.2.2. Subclinical Mastitis
3. Role of the Dry Period in the Life of Cows
3.1. Anatomical and Physiological Changes of the Mammary Gland
3.2. Susceptability to Intramammary Infections during the Dry Period
4. Impact of Mammary Gland Infections on Dairy Herds
4.1. Mastitis—A Threat to the Dairy Industry Profitability
4.2. Mastitis in Relation to Other Diseases
5. Current Diagnostic Approaches
5.1. Laboratory Techniques
5.1.1. Somatic Cell Count
5.1.2. Bacterial Culturing
5.1.3. Polymerase Chain Reaction (PCR)—Based Methods
5.2. Cow-Side Tests
5.2.1. California Mastitis Test
5.2.2. Electrical Conductivity Test
5.2.3. On-Farm Culture
6. Comparison of Mastitis Tests and Future Approaches
7. Metabolomics Investigation of Mastitis
7.1. Application of Metabolomics in Periparturient Diseases
7.2. Application of Metabolomics for Mastitis Biomarker Discovery
8. Predictive Biomarkers: Opportunity for the Dairy Industry
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thompson-Crispi, K.A.; Miglior, F.; Mallard, B.A. Incidence Rates of Clinical Mastitis among Canadian Holsteins Classified as High, Average, or Low Immune Responders. Clin. Vaccine Immunol. 2013, 20, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Reyher, K.K.; Dohoo, I.R. Diagnosing Intramammary Infections: Evaluation of Composite Milk Samples to Detect Intramammary Infections. J. Dairy Sci. 2011, 94, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Leslie, K.E. Evaluation of Changes in Somatic Cell Counts as Indicators of New Intramammary Infections. Prev. Vet. Med. 1991, 10, 225–237. [Google Scholar] [CrossRef]
- Ruegg, P.L.; Petersson-Wolfe, C.S. Mastitis in Dairy Cows. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, ix–x. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadi, M.; Haine, D.; Kelton, D.F.; Barkema, H.W.; Hogeveen, H.; Keefe, G.P.; Dufour, S. Herd-Level Mastitis-Associated Costs on Canadian Dairy Farms. Front. Vet. Sci. 2018, 5, 100. [Google Scholar] [CrossRef] [PubMed]
- Culling and Replacement Rates in Dairy Herds in Canada; Canadian Dairy Information Centre (CDIC). Available online: https://agriculture.canada.ca/en/canadas-agriculture-sectors/animal-industry/canadian-dairy-information-centre/dairy-statistics-and-market-information/dairy-animal-genetics/culling-and-replacement-rates-dairy-herds-canada (accessed on 30 June 2021).
- Busanello, M.; Rossi, R.S.; Cassoli, L.D.; Pantoja, J.C.F.; Machado, P.F. Estimation of Prevalence and Incidence of Subclinical Mastitis in a Large Population of Brazilian Dairy Herds. J. Dairy Sci. 2017, 100, 6545–6553. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, J.; Piepers, S.; Supré, K.; De Vliegher, S. Pathogen-Specific Incidence Rate of Clinical Mastitis in Flemish Dairy Herds, Severity, and Association with Herd Hygiene. J. Dairy Sci. 2014, 97, 6926–6934. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.L. Etiological Agents of Bovine Mastitis. Vet. Microbiol. 1988, 16, 41–66. [Google Scholar] [CrossRef]
- Zadoks, R.N.; Middleton, J.R.; McDougall, S.; Katholm, J.; Schukken, Y.H. Molecular Epidemiology of Mastitis Pathogens of Dairy Cattle and Comparative Relevance to Humans. J. Mammary Gland. Biol. Neoplasia 2011, 16, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Moroni, P.; Nydam, D.V.; Ospina, P.A.; Scillieri-Smith, J.C.; Virkler, P.D.; Watters, R.D.; Welcome, F.L.; Zurakowski, M.J.; Ducharme, N.G.; Yeager, A.E. Diseases of the Teats and Udder. In Rebhun’s Diseases of Dairy Cattle; Elsevier: Amsterdam, The Netherlands, 2018; pp. 389–465. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Günther, J.; Fitzpatrick, J.; Fontaine, M.C.; Goetze, L.; Holst, O.; Leigh, J.; Petzl, W.; Schuberth, H.-J.; Sipka, A.; et al. Host-Response Patterns of Intramammary Infections in Dairy Cows. Vet. Immunol. Immunopathol. 2011, 144, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Dego, O.K.; van Dijk, J.E.; Nederbragt, H. Factors Involved in the Early Pathogenesis of Bovine Staphylococcus Aureus Mastitis with Emphasis on Bacterial Adhesion and Invasion. A Review. Vet. Q. 2002, 24, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Perssonwaller, K.; Bengtsson, B.; Lindberg, A.; Nyman, A.; Ericssonunnerstad, H. Incidence of Mastitis and Bacterial Findings at Clinical Mastitis in Swedish Primiparous Cows—Influence of Breed and Stage of Lactation. Vet. Microbiol. 2009, 134, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Heikkilä, A.-M.; Liski, E.; Pyörälä, S.; Taponen, S. Pathogen-Specific Production Losses in Bovine Mastitis. J. Dairy Sci. 2018, 101, 9493–9504. [Google Scholar] [CrossRef] [PubMed]
- Lara-Zárate, L.; López-Meza, J.E.; Ochoa-Zarzosa, A. Staphylococcus Aureus Inhibits Nuclear Factor Kappa B Activation Mediated by Prolactin in Bovine Mammary Epithelial Cells. Microb. Pathog. 2011, 51, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, A.E.J.; van den Borne, B.H.P.; Wall, S.K.; Wellnitz, O.; Bruckmaier, R.M.; Spadavecchia, C. Experimentally Induced Subclinical Mastitis: Are Lipopolysaccharide and Lipoteichoic Acid Eliciting Similar Pain Responses? Acta Vet. Scand. 2017, 59, 40. [Google Scholar] [CrossRef]
- Barkema, H.W.; Schukken, Y.H.; Zadoks, R.N. Invited Review: The Role of Cow, Pathogen, and Treatment Regimen in the Therapeutic Success of Bovine Staphylococcus Aureus Mastitis. J. Dairy Sci. 2006, 89, 1877–1895. [Google Scholar] [CrossRef]
- Pyörälä, S.; Hovinen, M.; Simojoki, H.; Fitzpatrick, J.; Eckersall, P.D.; Orro, T. Acute Phase Proteins in Milk in Naturally Acquired Bovine Mastitis Caused by Different Pathogens. Vet. Rec. 2011, 168, 535. [Google Scholar] [CrossRef]
- Hogan, J.; Larry Smith, K. Coliform Mastitis. Vet. Res. 2003, 34, 507–519. [Google Scholar] [CrossRef]
- Eckel, E.F.; Ametaj, B.N. Invited Review: Role of Bacterial Endotoxins in the Etiopathogenesis of Periparturient Diseases of Transition Dairy Cows. J. Dairy Sci. 2016, 99, 5967–5990. [Google Scholar] [CrossRef]
- Leigh, J.A. Streptococcus Uberis: A Permanent Barrier to the Control of Bovine Mastitis? Vet. J. 1999, 157, 225–238. [Google Scholar] [CrossRef]
- Bradley, A.J.; Leach, K.A.; Breen, J.E.; Green, L.E.; Green, M.J. Survey of the Incidence and Aetiology of Mastitis on Dairy Farms in England and Wales. Vet. Rec. 2007, 160, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Rato, M.G.; Nerlich, A.; Bergmann, R.; Bexiga, R.; Nunes, S.F.; Vilela, C.L.; Santos-Sanches, I.; Chhatwal, G.S. Virulence Gene Pool Detected in Bovine Group C Streptococcus Dysgalactiae Subsp. Dysgalactiae Isolates by Use of a Group A S. Pyogenes Virulence Microarray. J. Clin. Microbiol. 2011, 49, 2470–2479. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.; Hulland, C.; Ruegg, P.L. Characterization of Clinical Mastitis Occurring in Cows on 50 Large Dairy Herds in Wisconsin. J. Dairy Sci. 2013, 96, 7538–7549. [Google Scholar] [CrossRef] [PubMed]
- Levison, L.J.; Miller-Cushon, E.K.; Tucker, A.L.; Bergeron, R.; Leslie, K.E.; Barkema, H.W.; DeVries, T.J. Incidence Rate of Pathogen-Specific Clinical Mastitis on Conventional and Organic Canadian Dairy Farms. J. Dairy Sci. 2016, 99, 1341–1350. [Google Scholar] [CrossRef] [PubMed]
- Gröhn, Y.T.; Erb, H.N.; McCulloch, C.E.; Saloniemi, H.S. Epidemiology of Metabolic Disorders in Dairy Cattle: Association Among Host Characteristics, Disease, and Production. J. Dairy Sci. 1989, 72, 1876–1885. [Google Scholar] [CrossRef]
- Schönborn, S.; Wente, N.; Paduch, J.-H.; Krömker, V. In Vitro Ability of Mastitis Causing Pathogens to Form Biofilms. J. Dairy Res. 2017, 84, 198–201. [Google Scholar] [CrossRef]
- Massé, J.; Dufour, S.; Archambault, M. Characterization of Klebsiella Isolates Obtained from Clinical Mastitis Cases in Dairy Cattle. J. Dairy Sci. 2020, 103, 3392–3400. [Google Scholar] [CrossRef]
- Tremblay, Y.D.N.; Lamarche, D.; Chever, P.; Haine, D.; Messier, S.; Jacques, M. Characterization of the Ability of Coagulase-Negative Staphylococci Isolated from the Milk of Canadian Farms to Form Biofilms. J. Dairy Sci. 2013, 96, 234–246. [Google Scholar] [CrossRef]
- Reyher, K.K.; Dohoo, I.R.; Scholl, D.T.; Keefe, G.P. Evaluation of Minor Pathogen Intramammary Infection, Susceptibility Parameters, and Somatic Cell Counts on the Development of New Intramammary Infections with Major Mastitis Pathogens. J. Dairy Sci. 2012, 95, 3766–3780. [Google Scholar] [CrossRef]
- Sargeant, J.M.; Scott, H.M.; Leslie, K.E.; Ireland, M.J.; Bashiri, A. Clinical Mastitis in Dairy Cattle in Ontario: Frequency of Occurrence and Bacteriological Isolates. Can. Vet. J. 1998, 39, 33–38. [Google Scholar]
- Barkema, H.W.; Schukken, Y.H.; Lam, T.J.G.M.; Beiboer, M.L.; Wilmink, H.; Benedictus, G.; Brand, A. Incidence of Clinical Mastitis in Dairy Herds Grouped in Three Categories by Bulk Milk Somatic Cell Counts. J. Dairy Sci. 1998, 81, 411–419. [Google Scholar] [CrossRef]
- Roberson, J.R.; Warnick, L.D.; Moore, G. Mild to Moderate Clinical Mastitis: Efficacy of Intramammary Amoxicillin, Frequent Milk-Out, a Combined Intramammary Amoxicillin, and Frequent Milk-Out Treatment Versus No Treatment. J. Dairy Sci. 2004, 87, 583–592. [Google Scholar] [CrossRef]
- Reksen, O.; Sølverød, L.; Branscum, A.J.; Østerås, O. Relationships Between Milk Culture Results and Treatment for Clinical Mastitis or Culling in Norwegian Dairy Cattle. J. Dairy Sci. 2006, 89, 2928–2937. [Google Scholar] [CrossRef]
- McDougall, S. Prevalence of Clinical Mastitis in 38 Waikato Dairy Herds in Early Lactation. N. Z. Vet. J. 1999, 47, 143–149. [Google Scholar] [CrossRef]
- Kopali, A.; Shoshi, N.; Koleci, X. Prevalence of Subclinical Mastitis in Dairy Cows: A Case Study of the Livestock Complex, Tirana (Albania). Res. Opin. Anim. Vet. Sci. 2011, 593–596. [Google Scholar]
- He, W.; Ma, S.; Lei, L.; He, J.; Li, X.; Tao, J.; Wang, X.; Song, S.; Wang, Y.; Wang, Y.; et al. Prevalence, Etiology, and Economic Impact of Clinical Mastitis on Large Dairy Farms in China. Vet. Microbiol. 2020, 242, 108570. [Google Scholar] [CrossRef] [PubMed]
- Breen, J.E.; Green, M.J.; Bradley, A.J. Quarter and Cow Risk Factors Associated with the Occurrence of Clinical Mastitis in Dairy Cows in the United Kingdom. J. Dairy Sci. 2009, 92, 2551–2561. [Google Scholar] [CrossRef]
- Sordillo, L.M. Mammary Gland Immunobiology and Resistance to Mastitis. Vet. Clin. N. Am.-Food Anim. Pract. 2018, 34, 507–523. [Google Scholar] [CrossRef]
- Rainard, P.; Riollet, C. Innate Immunity of the Bovine Mammary Gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Leopold, S.J.; Cranendonk, D.R.; van der Poll, T. Host Innate Immune Responses to Sepsis. Virulence 2014, 5, 36–44. [Google Scholar] [CrossRef]
- Miyake, K. Innate Immune Sensing of Pathogens and Danger Signals by Cell Surface Toll-like Receptors. Semin. Immunol. 2007, 19, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Mani, V.; Weber, T.E.; Baumgard, L.H.; Gabler, N.K. Growth and Development Symposium: Endotoxin, Inflammation, and Intestinal Function in Livestock1,2. J. Anim. Sci. 2012, 90, 1452–1465. [Google Scholar] [CrossRef] [PubMed]
- Brenaut, P.; Lefèvre, L.; Rau, A.; Laloë, D.; Pisoni, G.; Moroni, P.; Bevilacqua, C.; Martin, P. Contribution of Mammary Epithelial Cells to the Immune Response during Early Stages of a Bacterial Infection to Staphylococcus Aureus. Vet. Res. 2014, 45, 16. [Google Scholar] [CrossRef]
- Zhao, X.; Lacasse, P. Mammary Tissue Damage during Bovine Mastitis: Causes and Control1. J. Anim. Sci. 2008, 86 (Suppl. 13), 57–65. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Streicher, K.L. Mammary Gland Immunity and Mastitis Susceptibility. J. Mammary Gland. Biol. Neoplasia 2002, 7, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Ezzat Alnakip, M.; Quintela-Baluja, M.; Böhme, K.; Fernández-No, I.; Caamaño-Antelo, S.; Calo-Mata, P.; Barros-Velázquez, J. The Immunology of Mammary Gland of Dairy Ruminants between Healthy and Inflammatory Conditions. J. Vet. Med. 2014, 2014, 659801. [Google Scholar] [CrossRef] [PubMed]
- Leslie, K.E.; Petersson-Wolfe, C.S. Assessment and Management of Pain in Dairy Cows with Clinical Mastitis. Vet. Clin. N. Am.-Food Anim. Pract. 2012, 28, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Roberson, J.R. Treatment of Clinical Mastitis. Vet. Clin. N. Am.-Food Anim. Pract. 2012, 28, 271–288. [Google Scholar] [CrossRef] [PubMed]
- Adkins, P.R.F.; Middleton, J.R. Methods for Diagnosing Mastitis. Vet. Clin. N. Am.-Food Anim. Pract. 2018, 34, 479–491. [Google Scholar] [CrossRef]
- Ruegg, P.L. Managing Mastitis and Producing Quality Milk. In Dairy Production Medicine; Risco, C.A., Retamal, P.M., Eds.; Wiley: Hoboken, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Suojala, L.; Kaartinen, L.; Pyörälä, S. Treatment for Bovine Escherichia Coli Mastitis—An Evidence-Based Approach. J. Vet. Pharmacol. Ther. 2013, 36, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Forsbäck, L.; Lindmark-Månsson, H.; Andrén, A.; Åkerstedt, M.; Svennersten-Sjaunja, K. Udder Quarter Milk Composition at Different Levels of Somatic Cell Count in Cow Composite Milk. Animal 2009, 3, 710–717. [Google Scholar] [CrossRef]
- Leitner, G.; Silanikove, N.; Merin, U. Estimate of Milk and Curd Yield Loss of Sheep and Goats with Intrammamary Infection and Its Relation to Somatic Cell Count. Small Rumin. Res. 2008, 74, 221–225. [Google Scholar] [CrossRef]
- Pitkälä, A.; Haveri, M.; Pyörälä, S.; Myllys, V.; Honkanen-Buzalski, T. Bovine Mastitis in Finland 2001—Prevalence, Distribution of Bacteria, and Antimicrobial Resistance. J. Dairy Sci. 2004, 87, 2433–2441. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Wilson, D.J.; Welcome, F.; Garrison-Tikofsky, L.; Gonzalez, R.N. Monitoring Udder Health and Milk Quality Using Somatic Cell Counts. Vet. Res. 2003, 34, 579–596. [Google Scholar] [CrossRef] [PubMed]
- Rhoda, D.A.; Pantoja, J.C.F. Using Mastitis Records and Somatic Cell Count Data. Vet. Clin. N. Am.-Food Anim. Pract. 2012, 28, 347–361. [Google Scholar] [CrossRef] [PubMed]
- Dingwell, R.T.; Kelton, D.F.; Leslie, K.E. Management of the Dry Cow in Control of Peripartum Disease and Mastitis. Vet. Clin. N. Am.-Food Anim. Pract. 2003, 19, 235–265. [Google Scholar] [CrossRef]
- Jones, G. Proper Dry Cow Management Critical for Mastitis Control Drying-Off. Va. Tech. 2009, 404–212, 1–6. [Google Scholar]
- Capuco, A.V.; Akers, R.M.; Smith, J.J. Mammary Growth in Holstein Cows During the Dry Period: Quantification of Nucleic Acids and Histology. J. Dairy Sci. 1997, 80, 477–487. [Google Scholar] [CrossRef]
- Kuhn, M.T.; Hutchison, J.L.; Norman, H.D. Minimum Days Dry to Maximize Milk Yield in Subsequent Lactation. Anim. Res. 2005, 54, 351–367. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Moyes, K.M. Factors Contributing to Immunosuppression in the Dairy Cow during the Periparturient Period. Jpn. J. Vet. Res. 2015, 63 (Suppl. 1), S15–S24. [Google Scholar]
- Dingwell, R.T.; Leslie, K.E.; Schukken, Y.H.; Sargeant, J.M.; Timms, L.L.; Duffield, T.F.; Keefe, G.P.; Kelton, D.F.; Lissemore, K.D.; Conklin, J. Association of Cow and Quarter-Level Factors at Drying-off with New Intramammary Infections during the Dry Period. Prev. Vet. Med. 2004, 63, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Butler, W.R.; Smith, R.D. Interrelationships Between Energy Balance and Postpartum Reproductive Function in Dairy Cattle. J. Dairy Sci. 1989, 72, 767–783. [Google Scholar] [CrossRef]
- Bell, A.W. Regulation of Organic Nutrient Metabolism during Transition from Late Pregnancy to Early Lactation. J. Anim. Sci. 1995, 73, 2804. [Google Scholar] [CrossRef] [PubMed]
- Ametaj, B. A New Understanding of the Causes of Fatty Liver in Dairy Cows. Adv. Dairy Technol. 2005, 17, 97–112. [Google Scholar]
- Von Keyserlingk, M.A.G.; Rushen, J.; de Passillé, A.M.; Weary, D.M. Invited Review: The Welfare of Dairy Cattle—Key Concepts and the Role of Science. J. Dairy Sci. 2009, 92, 4101–4111. [Google Scholar] [CrossRef]
- Lucy, M.C.; Jiang, H.; Kobayashi, Y. Changes in the Somatotrophic Axis Associated with the Initiation of Lactation. J. Dairy Sci. 2001, 84, E113–E119. [Google Scholar] [CrossRef]
- Ingvartsen, K.L. Feeding- and Management-Related Diseases in the Transition Cow. Anim. Feed. Sci. Technol. 2006, 126, 175–213. [Google Scholar] [CrossRef]
- De Vliegher, S.; Ohnstad, I.; Piepers, S. Management and Prevention of Mastitis: A Multifactorial Approach with a Focus on Milking, Bedding and Data-Management. J. Integr. Agric. 2018, 17, 1214–1233. [Google Scholar] [CrossRef]
- Sordillo, L.M. Factors Affecting Mammary Gland Immunity and Mastitis Susceptibility. Livest. Prod. Sci. 2005, 98, 89–99. [Google Scholar] [CrossRef]
- Contreras, G.A.; Rodríguez, J.M. Mastitis: Comparative Etiology and Epidemiology. J. Mammary Gland. Biol. Neoplasia 2011, 16, 339–356. [Google Scholar] [CrossRef] [PubMed]
- Hogan, J.S.; Smith, K.L.; Hoblet, K.H.; Todhunter, D.A.; Schoenberger, P.S.; Hueston, W.D.; Pritchard, D.E.; Bowman, G.L.; Heider, L.E.; Brockett, B.L. Bacterial Counts in Bedding Materials Used on Nine Commercial Dairies. J. Dairy Sci. 1989, 72, 250–258. [Google Scholar] [CrossRef]
- Hogan, J.S.; Bogacz, V.L.; Thompson, L.M.; Romig, S.; Schoenberger, P.S.; Weiss, W.P.; Smith, K.L. Bacterial Counts Associated with Sawdust and Recycled Manure Bedding Treated with Commercial Conditioners. J. Dairy Sci. 1999, 82, 1690–1695. [Google Scholar] [CrossRef]
- Ruegg, P.L. Management of Mastitis on Organic and Conventional Dairy Farms1. J. Anim. Sci. 2009, 87 (Suppl. 13), 43–55. [Google Scholar] [CrossRef]
- Bannerman, D.D.; Kauf, A.C.W.; Paape, M.J.; Springer, H.R.; Goff, J.P. Comparison of Holstein and Jersey Innate Immune Responses to Escherichia Coli Intramammary Infection. J. Dairy Sci. 2008, 91, 2225–2235. [Google Scholar] [CrossRef]
- Dezetter, C.; Bareille, N.; Billon, D.; Côrtes, C.; Lechartier, C.; Seegers, H. Changes in Animal Performance and Profitability of Holstein Dairy Operations after Introduction of Crossbreeding with Montbéliarde, Normande, and Scandinavian Red. J. Dairy Sci. 2017, 100, 8239–8264. [Google Scholar] [CrossRef] [PubMed]
- Heringstad, B.; Chang, Y.M.; Gianola, D.; Klemetsdal, G. Genetic Analysis of Clinical Mastitis, Milk Fever, Ketosis, and Retained Placenta in Three Lactations of Norwegian Red Cows. J. Dairy Sci. 2005, 88, 3273–3281. [Google Scholar] [CrossRef]
- Negussie, E.; Strandén, I.; Mäntysaari, E.A. Genetic Association of Clinical Mastitis with Test-Day Somatic Cell Score and Milk Yield During First Lactation of Finnish Ayrshire Cows. J. Dairy Sci. 2008, 91, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Lush, J.L. Inheritance of Susceptibility to Mastitis. J. Dairy Sci. 1950, 33, 121–125. [Google Scholar] [CrossRef]
- Shook, G.E. Selection for Disease Resistance. J. Dairy Sci. 1989, 72, 1349–1362. [Google Scholar] [CrossRef]
- De Haas, Y.; Ouweltjes, W.; ten Napel, J.; Windig, J.J.; de Jong, G. Alternative Somatic Cell Count Traits as Mastitis Indicators for Genetic Selection. J. Dairy Sci. 2008, 91, 2501–2511. [Google Scholar] [CrossRef] [PubMed]
- Urioste, J.I.; Franzén, J.; Windig, J.J.; Strandberg, E. Genetic Relationships among Mastitis and Alternative Somatic Cell Count Traits in the First 3 Lactations of Swedish Holsteins. J. Dairy Sci. 2012, 95, 3428–3434. [Google Scholar] [CrossRef] [PubMed]
- Narayana, S.G.; Miglior, F.; Naqvi, S.A.; Malchiodi, F.; Martin, P.; Barkema, H.W. Genetic Analysis of Subclinical Mastitis in Early Lactation of Heifers Using Both Linear and Threshold Models. J. Dairy Sci. 2018, 101, 11120–11131. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, D.; Bennewitz, J.; Stamer, E.; Junge, W.; Kalm, E.; Thaller, G. Genetic Analysis of Mastitis Data with Different Models. J. Dairy Sci. 2011, 94, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Svendsen, M.; Heringstad, B. Somatic Cell Count as an Indicator of Sub-Clinical Mastitis. Genetic Parameters and Correlations with Clinical Mastitis. Interbull Bull. 2006, 35, 12. [Google Scholar]
- Olde Riekerink, R.G.M.; Barkema, H.W.; Kelton, D.F.; Scholl, D.T. Incidence Rate of Clinical Mastitis on Canadian Dairy Farms. J. Dairy Sci. 2008, 91, 1366–1377. [Google Scholar] [CrossRef]
- Green, M.J.; Green, L.E.; Medley, G.F.; Schukken, Y.H.; Bradley, A.J. Influence of Dry Period Bacterial Intramammary Infection on Clinical Mastitis in Dairy Cows. J. Dairy Sci. 2002, 85, 2589–2599. [Google Scholar] [CrossRef]
- Bradley, A.; Breen, J.; Green, M. Management: Mastitis Pattern Analysis—A Fresh Look at the Analysis of Bovine Mastitis: Part 2—Clinical Mastitis Data. Livestock 2008, 13, 30–35. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Dunn, S.M.; Ametaj, B.N. Innate Immunity and Carbohydrate Metabolism Alterations Precede Occurrence of Subclinical Mastitis in Transition Dairy Cows. J. Anim. Sci. Technol. 2015, 57, 46. [Google Scholar] [CrossRef]
- Jamali, H.; Barkema, H.W.; Jacques, M.; Lavallée-Bourget, E.-M.; Malouin, F.; Saini, V.; Stryhn, H.; Dufour, S. Invited Review: Incidence, Risk Factors, and Effects of Clinical Mastitis Recurrence in Dairy Cows. J. Dairy Sci. 2018, 101, 4729–4746. [Google Scholar] [CrossRef]
- Dairy Cattle: Principles, Practices, Problems, Profits; Foley, R.C., Ed.; Lea & Febiger: Philadelphia, PA, USA, 1972. [Google Scholar]
- Hertl, J.A.; Schukken, Y.H.; Welcome, F.L.; Tauer, L.W.; Gröhn, Y.T. Effects of Pathogen-Specific Clinical Mastitis on Probability of Conception in Holstein Dairy Cows. J. Dairy Sci. 2014, 97, 6942–6954. [Google Scholar] [CrossRef]
- Halasa, T.; Nielen, M.; De Roos, A.P.W.; Van Hoorne, R.; de Jong, G.; Lam, T.J.G.M.; van Werven, T.; Hogeveen, H. Production Loss Due to New Subclinical Mastitis in Dutch Dairy Cows Estimated with a Test-Day Model. J. Dairy Sci. 2009, 92, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Botaro, B.G.; Cortinhas, C.S.; Dibbern, A.G.; e Silva, L.F.P.; Benites, N.R.; dos Santos, M.V. Staphylococcus Aureus Intramammary Infection Affects Milk Yield and SCC of Dairy Cows. Trop. Anim. Health Prod. 2015, 47, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, S. Monitoring Metabolic Health of Dairy Cattle in the Transition Period. J. Reprod. Dev. 2010, 56, S29–S35. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.L. Investigation of Mastitis Problems on Farms. Vet. Clin. N. Am.-Food Anim. Pract. 2003, 19, 47–73. [Google Scholar] [CrossRef]
- Hogeveen, H.; Huijps, K.; Lam, T. Economic Aspects of Mastitis: New Developments. N. Z. Vet. J. 2011, 59, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Halasa, T.; Huijps, K.; Østerås, O.; Hogeveen, H. Economic Effects of Bovine Mastitis and Mastitis Management: A Review. Vet. Q. 2007, 29, 18–31. [Google Scholar] [CrossRef]
- Brown, K.; Uwiera, R.R.E.; Kalmokoff, M.L.; Brooks, S.P.J.; Inglis, G.D. Antimicrobial Growth Promoter Use in Livestock: A Requirement to Understand Their Modes of Action to Develop Effective Alternatives. Int. J. Antimicrob. Agents 2017, 49, 12–24. [Google Scholar] [CrossRef]
- Oliver, S.P.; Murinda, S.E. Antimicrobial Resistance of Mastitis Pathogens. Vet. Clin. N. Am.-Food Anim. Pract. 2012, 28, 165–185. [Google Scholar] [CrossRef]
- Aga, D.S.; Lenczewski, M.; Snow, D.; Muurinen, J.; Sallach, J.B.; Wallace, J.S. Challenges in the Measurement of Antibiotics and in Evaluating Their Impacts in Agroecosystems: A Critical Review. J. Environ. Qual. 2016, 45, 407–419. [Google Scholar] [CrossRef]
- Pol, M.; Ruegg, P.L. Treatment Practices and Quantification of Antimicrobial Drug Usage in Conventional and Organic Dairy Farms in Wisconsin. J. Dairy Sci. 2007, 90, 249–261. [Google Scholar] [CrossRef]
- Maynou, G.; Migura-Garcia, L.; Chester-Jones, H.; Ziegler, D.; Bach, A.; Terré, M. Effects of Feeding Pasteurized Waste Milk to Dairy Calves on Phenotypes and Genotypes of Antimicrobial Resistance in Fecal Escherichia Coli Isolates before and after Weaning. J. Dairy Sci. 2017, 100, 7967–7979. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.V. Does the Use of Antimicrobial Agents in Veterinary Medicine and Animal Husbandry Select Antibiotic-Resistant Bacteria That Infect Man and Compromise Antimicrobial Chemotherapy? J. Antimicrob. Chemother. 1996, 38, 1–3. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oliver, S.P.; Jayarao, B.M.; Almeida, R.A. Foodborne Pathogens in Milk and the Dairy Farm Environment: Food Safety and Public Health Implications. Foodborne Pathog. Dis. 2005, 2, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.A.M.; Martins, V.C.; Cardoso, F.A.; Germano, J.; Rodrigues, M.; Duarte, C.; Bexiga, R.; Cardoso, S.; Freitas, P.P. Biosensors for On-Farm Diagnosis of Mastitis. Front. Bioeng. Biotechnol. 2019, 7, 186. [Google Scholar] [CrossRef]
- Otto, S.J.G.; Szkotnicki, J.; McElwain, C.; So, I.; Weese, J.S.; Prescott, J.F. Building the Antimicrobial Stewardship Leadership Plan for Animal Health in Canada (Workshop, Ottawa, 3–4 October 2017). Can. Vet. J. 2018, 59, 746–748. [Google Scholar] [PubMed]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Deng, Q.; Goldansaz, S.; Dunn, S.; Ametaj, B. Alterations of Innate Immunity Reactants in Transition Dairy Cows before Clinical Signs of Lameness. Animals 2015, 5, 717–747. [Google Scholar] [CrossRef]
- Zhang, G.; Hailemariam, D.; Dervishi, E.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Dairy Cows Affected by Ketosis Show Alterations in Innate Immunity and Lipid and Carbohydrate Metabolism during the Dry off Period and Postpartum. Res. Vet. Sci. 2016, 107, 246–256. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Dunn, S.M.; Ametaj, B.N. Occurrence of Retained Placenta Is Preceded by an Inflammatory State and Alterations of Energy Metabolism in Transition Dairy Cows. J. Anim. Sci. Biotechnol. 2016, 7, 26. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Hailemariam, D.; Goldansaz, S.A.; Deng, Q.; Dunn, S.M.; Ametaj, B.N. Alterations in Innate Immunity Reactants and Carbohydrate and Lipid Metabolism Precede Occurrence of Metritis in Transition Dairy Cows. Res. Vet. Sci. 2016, 104, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, F.J.; Doherty, M.L. Production Diseases of the Transition Cow. Vet. J. 2008, 176, 3–9. [Google Scholar] [CrossRef]
- Emmanuel, D.G.V.; Madsen, K.L.; Churchill, T.A.; Dunn, S.M.; Ametaj, B.N. Acidosis and Lipopolysaccharide from Escherichia Coli B:055 Cause Hyperpermeability of Rumen and Colon Tissues. J. Dairy Sci. 2007, 90, 5552–5557. [Google Scholar] [CrossRef] [PubMed]
- Zebeli, Q.; Dunn, S.M.; Ametaj, B.N. Perturbations of Plasma Metabolites Correlated with the Rise of Rumen Endotoxin in Dairy Cows Fed Diets Rich in Easily Degradable Carbohydrates. J. Dairy Sci. 2011, 94, 2374–2382. [Google Scholar] [CrossRef]
- Saleem, F.; Ametaj, B.N.; Bouatra, S.; Mandal, R.; Zebeli, Q.; Dunn, S.M.; Wishart, D.S. A Metabolomics Approach to Uncover the Effects of Grain Diets on Rumen Health in Dairy Cows. J. Dairy Sci. 2012, 95, 6606–6623. [Google Scholar] [CrossRef]
- Zwierzchowski, G.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Mass-Spec-Based Urinary Metabotyping around Parturition Identifies Screening Biomarkers for Subclinical Mastitis in Dairy Cows. Res. Vet. Sci. 2020, 129, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.F. The Effect of Nutritional Management of the Dairy Cow on Reproductive Efficiency. Anim. Reprod. Sci. 2006, 96, 282–296. [Google Scholar] [CrossRef] [PubMed]
- Lam, T.J.; Schukken, Y.H.; van Vliet, J.H.; Grommers, F.J.; Tielen, M.J.; Brand, A. Effect of Natural Infection with Minor Pathogens on Susceptibility to Natural Infection with Major Pathogens in the Bovine Mammary Gland. Am. J. Vet. Res. 1997, 58, 17–22. [Google Scholar] [PubMed]
- Zadoks, R.N.; Allore, H.G.; Barkema, H.W.; Sampimon, O.C.; Wellenberg, G.J.; Gröhn, Y.T.; Schukken, Y.H. Cow- and Quarter-Level Risk Factors for Streptococcus Uberis and Staphylococcus Aureus Mastitis. J. Dairy Sci. 2001, 84, 2649–2663. [Google Scholar] [CrossRef]
- De Vries, A. Economic Trade-Offs between Genetic Improvement and Longevity in Dairy Cattle. J. Dairy Sci. 2017, 100, 4184–4192. [Google Scholar] [CrossRef] [PubMed]
- Ahmadzadeh, A.; Frago, F.; Shafii, B.; Dalton, J.C.; Price, W.J.; McGuire, M.A. Effect of Clinical Mastitis and Other Diseases on Reproductive Performance of Holstein Cows. Anim. Reprod. Sci. 2009, 112, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Barker, A.R.; Schrick, F.N.; Lewis, M.J.; Dowlen, H.H.; Oliver, S.P. Influence of Clinical Mastitis During Early Lactation on Reproductive Performance of Jersey Cows. J. Dairy Sci. 1998, 81, 1285–1290. [Google Scholar] [CrossRef]
- Schrick, F.N.; Hockett, M.E.; Saxton, A.M.; Lewis, M.J.; Dowlen, H.H.; Oliver, S.P. Influence of Subclinical Mastitis During Early Lactation on Reproductive Parameters. J. Dairy Sci. 2001, 84, 1407–1412. [Google Scholar] [CrossRef]
- Fourichon, C.; Seegers, H.; Malher, X. Effect of Disease on Reproduction in the Dairy Cow: A Meta-Analysis. Theriogenology 2000, 53, 1729–1759. [Google Scholar] [CrossRef]
- Dolecheck, K.A.; García-Guerra, A.; Moraes, L.E. Quantifying the Effects of Mastitis on the Reproductive Performance of Dairy Cows: A Meta-Analysis. J. Dairy Sci. 2019, 102, 8454–8477. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.E.B.; Dyk, P.B.; Herdt, T.H.; Kaneene, J.B.; Miller, R.; Bucholtz, H.F.; Liesman, J.S.; Vandehaar, M.J.; Emery, R.S. Dry Cow Diet, Management, and Energy Balance as Risk Factors for Displaced Abomasum in High Producing Dairy Herds. J. Dairy Sci. 1998, 81, 132–139. [Google Scholar] [CrossRef]
- Moyes, K.M.; Larsen, T.; Friggens, N.C.; Drackley, J.K.; Ingvartsen, K.L. Identification of Potential Markers in Blood for the Development of Subclinical and Clinical Mastitis in Dairy Cattle at Parturition and during Early Lactation. J. Dairy Sci. 2009, 92, 5419–5428. [Google Scholar] [CrossRef] [PubMed]
- Zarrin, M.; Wellnitz, O.; van Dorland, H.A.; Bruckmaier, R.M. Induced Hyperketonemia Affects the Mammary Immune Response during Lipopolysaccharide Challenge in Dairy Cows. J. Dairy Sci. 2014, 97, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Wellnitz, O.; Bruckmaier, R.M. The Innate Immune Response of the Bovine Mammary Gland to Bacterial Infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Waldron, M.R.; Kulick, A.E.; Bell, A.W.; Overton, T.R. Acute Experimental Mastitis Is Not Causal Toward the Development of Energy-Related Metabolic Disorders in Early Postpartum Dairy Cows. J. Dairy Sci. 2006, 89, 596–610. [Google Scholar] [CrossRef]
- Vernay, M.C.M.B.; Wellnitz, O.; Kreipe, L.; van Dorland, H.A.; Bruckmaier, R.M. Local and Systemic Response to Intramammary Lipopolysaccharide Challenge during Long-Term Manipulated Plasma Glucose and Insulin Concentrations in Dairy Cows. J. Dairy Sci. 2012, 95, 2540–2549. [Google Scholar] [CrossRef]
- Waldron, M.R.; Nishida, T.; Nonnecke, B.J.; Overton, T.R. Effect of Lipopolysaccharide on Indices of Peripheral and Hepatic Metabolism in Lactating Cows. J. Dairy Sci. 2003, 86, 3447–3459. [Google Scholar] [CrossRef]
- Huszenicza, G.; Jánosi, S.; Gáspárdy, A.; Kulcsár, M. Endocrine Aspects in Pathogenesis of Mastitis in Postpartum Dairy Cows. Anim. Reprod. Sci. 2004, 82–83, 389–400. [Google Scholar] [CrossRef]
- Ametaj, B.N.; Zebeli, Q.; Iqbal, S. Nutrition, Microbiota, and Endotoxin-Related Diseases in Dairy Cows. Rev. Bras. Zootec. 2010, 39, 433–444. [Google Scholar] [CrossRef]
- Viguier, C.; Arora, S.; Gilmartin, N.; Welbeck, K.; O’Kennedy, R. Mastitis Detection: Current Trends and Future Perspectives. Trends Biotechnol. 2009, 27, 486–493. [Google Scholar] [CrossRef]
- Koeck, A.; Miglior, F.; Kelton, D.F.; Schenkel, F.S. Alternative Somatic Cell Count Traits to Improve Mastitis Resistance in Canadian Holsteins. J. Dairy Sci. 2012, 95, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Berry, E.A.; Hillerton, J.E. The Effect of Selective Dry Cow Treatment on New Intramammary Infections. J. Dairy Sci. 2002, 85, 112–121. [Google Scholar] [CrossRef]
- Beli, E. Evaluation Of Albanian Raw Milk Quality Situation By Using Somatic Cell Counts. J. Multidiscip. Eng. Sci. Technol. (JMEST) 2016, 3, 5971–5974. [Google Scholar]
- Vissio, C.; Dieser, S.A.; Agnelli, H.L.; Odierno, L.M.; Larriestra, A.J. Accuracy of the Composite Somatic Cell Count to Detect Intra-Mammary Infection in Dairy Cows Using Latent Class Analysis. Prev. Vet. Med. 2014, 113, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, P.L.; Pantoja, J.C.F. Understanding and Using Somatic Cell Counts to Improve Milk Quality. Ir. J. Agric. Food Res. 2013, 52, 101–117. [Google Scholar]
- Pilla, R.; Schwarz, D.; König, S.; Piccinini, R. Microscopic Differential Cell Counting to Identify Inflammatory Reactions in Dairy Cow Quarter Milk Samples. J. Dairy Sci. 2012, 95, 4410–4420. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, T. Potential for Broad Applications of Flow Cytometry and Fluorescence Techniques in Microbiological and Somatic Cell Analyses of Milk. Int. J. Food Microbiol. 2003, 85, 269–279. [Google Scholar] [CrossRef]
- Dufour, S.; Fréchette, A.; Barkema, H.W.; Mussell, A.; Scholl, D.T. Invited Review: Effect of Udder Health Management Practices on Herd Somatic Cell Count. J. Dairy Sci. 2011, 94, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Bauman, C.A.; Barkema, H.W.; Dubuc, J.; Keefe, G.P.; Kelton, D.F. Canadian National Dairy Study: Herd-Level Milk Quality. J. Dairy Sci. 2018, 101, 2679–2691. [Google Scholar] [CrossRef]
- Hernández-Castellano, L.; Wall, S.; Stephan, R.; Corti, S.; Bruckmaier, R. Milk Somatic Cell Count, Lactate Dehydrogenase Activity, and Immunoglobulin G Concentration Associated with Mastitis Caused by Different Pathogens: A Field Study. SAT 2017, 159, 283–290. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hope, A. Laboratory Handbook on Bovine Mastitis. Aust. Vet. J. 2000, 78, 488. [Google Scholar] [CrossRef]
- Murphy, S.C.; Martin, N.H.; Barbano, D.M.; Wiedmann, M. Influence of Raw Milk Quality on Processed Dairy Products: How Do Raw Milk Quality Test Results Relate to Product Quality and Yield? J. Dairy Sci. 2016, 99, 10128–10149. [Google Scholar] [CrossRef] [PubMed]
- Klaas, I.C.; Zadoks, R.N. An Update on Environmental Mastitis: Challenging Perceptions. Transbound. Emerg. Dis. 2018, 65, 166–185. [Google Scholar] [CrossRef]
- Diseases of the Mammary Gland. In Veterinary Medicine; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1904–2001. [CrossRef]
- Ashraf, A.; Imran, M. Diagnosis of Bovine Mastitis: From Laboratory to Farm. Trop. Anim. Health Prod. 2018, 50, 1193–1202. [Google Scholar] [CrossRef]
- Bexiga, R.; Koskinen, M.T.; Holopainen, J.; Carneiro, C.; Pereira, H.; Ellis, K.A.; Vilela, C.L. Diagnosis of Intramammary Infection in Samples Yielding Negative Results or Minor Pathogens in Conventional Bacterial Culturing. J. Dairy Res. 2011, 78, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Lui, C.; Cady, N.; Batt, C. Nucleic Acid-Based Detection of Bacterial Pathogens Using Integrated Microfluidic Platform Systems. Sensors 2009, 9, 3713–3744. [Google Scholar] [CrossRef]
- Perreten, V.; Endimiani, A.; Thomann, A.; Wipf, J.R.K.; Rossano, A.; Bodmer, M.; Raemy, A.; Sannes-Lowery, K.A.; Ecker, D.J.; Sampath, R.; et al. Evaluation of PCR Electrospray-Ionization Mass Spectrometry for Rapid Molecular Diagnosis of Bovine Mastitis. J. Dairy Sci. 2013, 96, 3611–3620. [Google Scholar] [CrossRef]
- Lange, C.C.; Brito, M.A.V.P.; Reis, D.R.L.; Machado, M.A.; Guimarães, A.S.; Azevedo, A.L.S.; Salles, É.B.; Alvim, M.C.T.; Silva, F.S.; Meurer, I.R. Species-Level Identification of Staphylococci Isolated from Bovine Mastitis in Brazil Using Partial 16S RRNA Sequencing. Vet. Microbiol. 2015, 176, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Cantekïn, Z.; Ergün, Y.; Doğruer, G.; Saribay, M.K.; Solmaz, H. Süt Sığırlarında Sub-Klinik Mastitisin Tanısında Kültür ve PCR Yöntemlerinin Karşılaştırılması. Kafkas Univ. Vet. Fak. Derg. 2015, 21, 277–282. [Google Scholar] [CrossRef]
- Graber, H.U.; Casey, M.G.; Naskova, J.; Steiner, A.; Schaeren, W. Development of a Highly Sensitive and Specific Assay to Detect Staphylococcus Aureus in Bovine Mastitic Milk. J. Dairy Sci. 2007, 90, 4661–4669. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, P.; Zhou, S.; Zhang, L. Loop-Mediated Isothermal Amplification (LAMP): A Novel Rapid Detection Platform for Pathogens. Microb. Pathog. 2017, 107, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Anis, E.; Hawkins, I.K.; Ilha, M.R.S.; Woldemeskel, M.W.; Saliki, J.T.; Wilkes, R.P. Evaluation of Targeted Next-Generation Sequencing for Detection of Bovine Pathogens in Clinical Samples. J. Clin. Microbiol. 2018, 56, e00399-18. [Google Scholar] [CrossRef]
- Gonçalves, J.L.; Tomazi, T.; Barreiro, J.R.; de Campos Braga, P.A.; Ferreira, C.R.; Araújo Junior, J.P.; Eberlin, M.N.; dos Santos, M.V. Identification of Corynebacterium Spp. Isolated from Bovine Intramammary Infections by Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry. Vet. Microbiol. 2014, 173, 147–151. [Google Scholar] [CrossRef]
- Cameron, M.; Barkema, H.W.; De Buck, J.; De Vliegher, S.; Chaffer, M.; Lewis, J.; Keefe, G.P. Identification of Bovine-Associated Coagulase-Negative Staphylococci by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry Using a Direct Transfer Protocol. J. Dairy Sci. 2017, 100, 2137–2147. [Google Scholar] [CrossRef]
- Barreiro, J.R.; Gonçalves, J.L.; Braga, P.A.C.; Dibbern, A.G.; Eberlin, M.N.; Veiga dos Santos, M. Non-Culture-Based Identification of Mastitis-Causing Bacteria by MALDI-TOF Mass Spectrometry. J. Dairy Sci. 2017, 100, 2928–2934. [Google Scholar] [CrossRef]
- Schalm, O.W.; Noorlander, D.O. Experiments and Observations Leading to Development of the California Mastitis Test. J. Am. Vet. Med. Assoc. 1957, 130, 199–204. [Google Scholar]
- Sargeant, J.M.; Leslie, K.E.; Shirley, J.E.; Pulkrabek, B.J.; Lim, G.H. Sensitivity and Specificity of Somatic Cell Count and California Mastitis Test for Identifying Intramammary Infection in Early Lactation. J. Dairy Sci. 2001, 84, 2018–2024. [Google Scholar] [CrossRef]
- Gordon, W.A.; Morris, H.A.; Packard, V. Methods to Detect Abnormal Milk—A Review. J. Food Prot. 1980, 43, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Pyörälä, S. Indicators of Inflammation in the Diagnosis of Mastitis. Vet. Res. 2003, 34, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Hovinen, M.; Pyörälä, S. Invited Review: Udder Health of Dairy Cows in Automatic Milking. J. Dairy Sci. 2011, 94, 547–562. [Google Scholar] [CrossRef] [PubMed]
- John, A.J.; Garcia, S.C.; Kerrisk, K.L.; Freeman, M.J.; Islam, M.R.; Clark, C.E.F. Short Communication: The Diurnal Intake and Behavior of Dairy Cows When Access to a Feed of Consistent Nutritive Value Is Restricted. J. Dairy Sci. 2017, 100, 9279–9284. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.; Clark, C.E.F.; Lyons, N.A.; Thomson, P.C.; Kerrisk, K.L.; García, S.C. Early Detection of Clinical Mastitis from Electrical Conductivity Data in an Automatic Milking System. Anim. Prod. Sci. 2017, 57, 1226. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Blum, J.W. Oxytocin Release and Milk Removal in Ruminants. J. Dairy Sci. 1998, 81, 939–949. [Google Scholar] [CrossRef]
- Lehmann, M.; Wall, S.K.; Wellnitz, O.; Bruckmaier, R.M. Changes in Milk L-Lactate, Lactate Dehydrogenase, Serum Albumin, and IgG during Milk Ejection and Their Association with Somatic Cell Count. J. Dairy Sci. 2015, 82, 129–134. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Weiss, D.; Wiedemann, M.; Schmitz, S.; Wendl, G. Changes of Physicochemical Indicators during Mastitis and the Effects of Milk Ejection on Their Sensitivity. J. Dairy Sci. 2004, 71, 316–321. [Google Scholar] [CrossRef]
- Bansal, B.K.; Hamann, J.; Grabowski, N.T.; Singh, K.B. Variation in the Composition of Selected Milk Fraction Samples from Healthy and Mastitic Quarters, and Its Significance for Mastitis Diagnosis. J. Dairy Sci. 2005, 72, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.; Bruckmaier, R.M.; Thomson, P.C.; House, J.; García, S.C. Suitability of Somatic Cell Count, Electrical Conductivity, and Lactate Dehydrogenase Activity in Foremilk before versus after Alveolar Milk Ejection for Mastitis Detection. J. Dairy Sci. 2019, 102, 9200–9212. [Google Scholar] [CrossRef]
- Royster, E.; Godden, S.; Goulart, D.; Dahlke, A.; Rapnicki, P.; Timmerman, J. Evaluation of the Minnesota Easy Culture System II Bi-Plate and Tri-Plate for Identification of Common Mastitis Pathogens in Milk. J. Dairy Sci. 2014, 97, 3648–3659. [Google Scholar] [CrossRef] [PubMed]
- Dohoo, I.R.; Smith, J.; Andersen, S.; Kelton, D.F.; Godden, S. Diagnosing Intramammary Infections: Evaluation of Definitions Based on a Single Milk Sample. J. Dairy Sci. 2011, 94, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Kandeel, S.A.; Megahed, A.A.; Ebeid, M.H.; Constable, P.D. Ability of Milk PH to Predict Subclinical Mastitis and Intramammary Infection in Quarters from Lactating Dairy Cattle. J. Dairy Sci. 2019, 102, 1417–1427. [Google Scholar] [CrossRef]
- Boyd-Moss, M.; Baratchi, S.; Di Venere, M.; Khoshmanesh, K. Self-Contained Microfluidic Systems: A Review. Lab Chip 2016, 16, 3177–3192. [Google Scholar] [CrossRef]
- Sang, S.; Zhang, W.; Zhao, Y. Review on the Design Art of Biosensors. In State of the Art in Biosensors-General Aspects; Rinken, T., Ed.; InTech: London, UK, 2013. [Google Scholar] [CrossRef]
- Ruegg, P.L. A 100-Year Review: Mastitis Detection, Management, and Prevention. J. Dairy Sci. 2017, 100, 10381–10397. [Google Scholar] [CrossRef] [PubMed]
- Luedecke, L.O.; Forster, T.L.; Ashworth, U.S. Relationship Between California Mastitis Test Reaction and Leucocyte Count, Catalase Activity, and A-Esterase Activity of Milk from Opposite Quarters. J. Dairy Sci. 1967, 50, 1592–1596. [Google Scholar] [CrossRef]
- Lam, T.; Olde Riekerink, R.; Sampimon, O.; Smith, H. Mastitis Diagnostics and Performance Monitoring: A Practical Approach. Ir. Vet. J. 2009, 62, S34. [Google Scholar] [CrossRef] [PubMed]
- Dingwell, R.T.; Leslie, K.E.; Schukken, Y.H.; Sargeant, J.M.; Timms, L.L. Evaluation of the California Mastitis Test to Detect an Intramammary Infection with a Major Pathogen in Early Lactation Dairy Cows. Can. Vet. J. 2003, 44, 413–415. [Google Scholar] [PubMed]
- Rossi, R.S.; Amarante, A.F.; Correia, L.B.N.; Guerra, S.T.; Nobrega, D.B.; Latosinski, G.S.; Rossi, B.F.; Rall, V.L.M.; Pantoja, J.C.F. Diagnostic Accuracy of Somaticell, California Mastitis Test, and Microbiological Examination of Composite Milk to Detect Streptococcus Agalactiae Intramammary Infections. J. Dairy Sci. 2018, 101, 10220–10229. [Google Scholar] [CrossRef] [PubMed]
- Labohm, R.; Götz, E.; Luhofer, G.; Hess, R.G.; Bostedt, H. Factors Influencing the Somatic Milk-Cell-Count in Dairy Cows. 1. Influence of Bacteriological Findings, Stage and Number of Lactation. Milchwissenschaft 1998, 53, 63–66. [Google Scholar]
- Hillerton, E. Detecting Mastitis Cow-Side. In Proceedings of the National Mastitis Council Annual Meeting Proceedings (2000), Atlanta, GA, USA, 13–16 February 2000; pp. 48–53. [Google Scholar]
- Leslie, K.; Barratt, K.; Petersson, C.; Bashiri, A. An Evaluation of the Portascc® Test as a Measure of Udder Health Status Dairy Cows (an Excerpt From a Technical Report). Viewed 2015, 20, 2060–2061. [Google Scholar]
- Ferronatto, J.A.; Ferronatto, T.C.; Schneider, M.; Pessoa, L.F.; Blagitz, M.G.; Heinemann, M.B.; Della Libera, A.M.M.P.; Souza, F.N. Diagnosing Mastitis in Early Lactation: Use of Somaticell®, California Mastitis Test and Somatic Cell Count. Ital. J. Anim. Sci. 2018, 17, 723–729. [Google Scholar] [CrossRef]
- Olde Riekerink, R.G.M.; Barkema, H.W.; Veenstra, W.; Berg, F.E.; Stryhn, H.; Zadoks, R.N. Somatic Cell Count During and Between Milkings. J. Dairy Sci. 2007, 90, 3733–3741. [Google Scholar] [CrossRef] [PubMed]
- Petzer, I.-M.; Karzis, J.; Donkin, E.F.; Webb, E.C.; Etter, E.M.C. Somatic Cell Count Thresholds in Composite and Quarter Milk Samples as Indicator of Bovine Intramammary Infection Status. Onderstepoort J. Vet. Res. 2017, 84, a1269. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Odhiambo, J.F.; Farooq, U.; Lam, T.; Dunn, S.M.; Ametaj, B.N. Intravaginal Probiotics Modulated Metabolic Status and Improved Milk Production and Composition of Transition Dairy Cows. J. Anim. Sci. 2016, 94, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Jiminez, J.A.; Uwiera, T.C.; Abbott, D.W.; Uwiera, R.R.E.; Inglis, G.D. Butyrate Supplementation at High Concentrations Alters Enteric Bacterial Communities and Reduces Intestinal Inflammation in Mice Infected with Citrobacter Rodentium. mSphere 2017, 2, e00243-17. [Google Scholar] [CrossRef]
- Tarsillo, B.; Priefer, R. Proteobiotics as a New Antimicrobial Therapy. Microb. Pathog. 2020, 142, 104093. [Google Scholar] [CrossRef]
- Chakraborty, S.; Dhama, K.; Tiwari, R.; Iqbal Yatoo, M.; Khurana, S.K.; Khandia, R.; Munjal, A.; Munuswamy, P.; Kumar, M.A.; Singh, M.; et al. Technological Interventions and Advances in the Diagnosis of Intramammary Infections in Animals with Emphasis on Bovine Population—A Review. Vet. Q. 2019, 39, 76–94. [Google Scholar] [CrossRef] [PubMed]
- Dobson, H.; Smith, R.; Royal, M.; Knight, C.; Sheldon, I. The High-Producing Dairy Cow and Its Reproductive Performance. Reprod. Domest. Anim. 2007, 42, 17–23. [Google Scholar] [CrossRef]
- Sundrum, A. Metabolic Disorders in the Transition Period Indicate That the Dairy Cows’ Ability to Adapt Is Overstressed. Animals 2015, 5, 978–1020. [Google Scholar] [CrossRef]
- Drackley, J.K. Biology of Dairy Cows During the Transition Period: The Final Frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Raphael, W. Significance of Metabolic Stress, Lipid Mobilization, and Inflammation on Transition Cow Disorders. Vet. Clin. N. Am.-Food Anim. Pract. 2013, 29, 267–278. [Google Scholar] [CrossRef] [PubMed]
- David Baird, G. Primary Ketosis in the High-Producing Dairy Cow: Clinical and Subclinical Disorders, Treatment, Prevention, and Outlook. J. Dairy Sci. 1982, 65, 1–10. [Google Scholar] [CrossRef]
- Brunner, N.; Groeger, S.; Canelas Raposo, J.; Bruckmaier, R.M.; Gross, J.J. Prevalence of Subclinical Ketosis and Production Diseases in Dairy Cows in Central and South America, Africa, Asia, Australia, New Zealand, and Eastern Europe1. Transl. Anim. Sci. 2019, 3, 84–92. [Google Scholar] [CrossRef]
- Serkova, N.J.; Niemann, C.U. Pattern Recognition and Biomarker Validation Using Quantitative 1 H-NMR-Based Metabolomics. Expert Rev. Mol. Diagn. 2006, 6, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Dervishi, E.; Ametaj, B.N. Milk Fever in Dairy Cows Is Preceded by Activation of Innate Immunity and Alterations in Carbohydrate Metabolism Prior to Disease Occurrence. Res. Vet. Sci. 2018, 117, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Thammavongsa, V.; Kim, H.K.; Missiakas, D.; Schneewind, O. Staphylococcal Manipulation of Host Immune Responses. Nat. Rev. Microbiol. 2015, 13, 529–543. [Google Scholar] [CrossRef]
- Do Vale, A.; Cabanes, D.; Sousa, S. Bacterial Toxins as Pathogen Weapons Against Phagocytes. Front. Microbiol. 2016, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Loughman, J.A.; Hunstad, D.A. Attenuation of Human Neutrophil Migration and Function by Uropathogenic Bacteria. Microbes Infect. 2011, 13, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Lau, M.E.; Loughman, J.A.; Hunstad, D.A. YbcL of Uropathogenic Escherichia Coli Suppresses Transepithelial Neutrophil Migration. Infect. Immun. 2012, 80, 4123–4132. [Google Scholar] [CrossRef] [PubMed]
- Hailemariam, D.; Mandal, R.; Saleem, F.; Dunn, S.M.; Wishart, D.S.; Ametaj, B.N. Identification of Predictive Biomarkers of Disease State in Transition Dairy Cows. J. Dairy Sci. 2014, 97, 2680–2693. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.Z.; Shen, L.H.; Jiang, J.; Huang, Y.X.; Bai, L.P.; Yu, S.M.; Yao, X.P.; Ren, Z.H.; Yang, Y.X.; Cao, S.Z. Plasma Metabolite Changes in Dairy Cows during Parturition Identified Using Untargeted Metabolomics. J. Dairy Sci. 2019, 102, 4639–4650. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, C.; Xia, C.; Zhang, H.; Sun, L.; Gao, Y. Plasma Metabolic Profiling of Dairy Cows Affected with Clinical Ketosis Using LC/MS Technology. Vet. Q. 2014, 34, 152–158. [Google Scholar] [CrossRef]
- Sun, L.W.; Zhang, H.Y.; Wu, L.; Shu, S.; Xia, C.; Xu, C.; Zheng, J.S. 1H-Nuclear Magnetic Resonance-Based Plasma Metabolic Profiling of Dairy Cows with Clinical and Subclinical Ketosis. J. Dairy Sci. 2014, 97, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shu, S.; Xia, C.; Wang, P.; Sun, Y.; Xu, C.; Li, C. Mass Spectral Analysis of Urine Proteomic Profiles of Dairy Cows Suffering from Clinical Ketosis. Vet. Q. 2015, 35, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Deng, Q.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. DI/LC-MS/MS-Based Metabolic Profiling for Identification of Early Predictive Serum Biomarkers of Metritis in Transition Dairy Cows. J. Agric. Food Chem. 2017, 65, 8510–8521. [Google Scholar] [CrossRef]
- Zhang, G.; Dervishi, E.; Dunn, S.M.; Mandal, R.; Liu, P.; Han, B.; Wishart, D.S.; Ametaj, B.N. Metabotyping Reveals Distinct Metabolic Alterations in Ketotic Cows and Identifies Early Predictive Serum Biomarkers for the Risk of Disease. Metabolomics 2017, 13, 43. [Google Scholar] [CrossRef]
- Klein, M.S.; Almstetter, M.F.; Schlamberger, G.; Nürnberger, N.; Dettmer, K.; Oefner, P.J.; Meyer, H.H.D.; Wiedemann, S.; Gronwald, W. Nuclear Magnetic Resonance and Mass Spectrometry-Based Milk Metabolomics in Dairy Cows during Early and Late Lactation. J. Dairy Sci. 2010, 93, 1539–1550. [Google Scholar] [CrossRef]
- Klein, M.S.; Buttchereit, N.; Miemczyk, S.P.; Immervoll, A.-K.; Louis, C.; Wiedemann, S.; Junge, W.; Thaller, G.; Oefner, P.J.; Gronwald, W. NMR Metabolomic Analysis of Dairy Cows Reveals Milk Glycerophosphocholine to Phosphocholine Ratio as Prognostic Biomarker for Risk of Ketosis. J. Proteome Res. 2012, 11, 1373–1381. [Google Scholar] [CrossRef]
- Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. A Multi-Platform Metabolomics Approach Identifies Urinary Metabolite Signatures That Differentiate Ketotic From Healthy Dairy Cows. Front. Vet. Sci. 2021, 8, 595983. [Google Scholar] [CrossRef]
- Zheng, J.; Sun, L.; Shu, S.; Zhu, K.; Xu, C.; Wang, J.; Wang, H. Nuclear Magnetic Resonance-Based Serum Metabolic Profiling of Dairy Cows with Footrot. J. Vet. Med. Sci. 2016, 78, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Serum Metabolic Fingerprinting of Pre-Lameness Dairy Cows by GC–MS Reveals Typical Profiles That Can Identify Susceptible Cows. J. Proteom. 2020, 213, 103620. [Google Scholar] [CrossRef] [PubMed]
- Haxhiaj, K.; Li, Z.; Johnson, M.; Dunn, S.M.; Wishart, D.S.; Ametaj, B.N. Blood Metabolomic Phenotyping of Dry Cows Could Predict the High Milk Somatic Cells in Early Lactation—Preliminary Results. Dairy 2022, 3, 5. [Google Scholar] [CrossRef]
- Zhang, G.; Dervishi, E.; Zwierzchowski, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Urinary Metabolomics around Parturition Identifies Metabolite Alterations in Dairy Cows Affected Postpartum by Lameness: Preliminary Study. Dairy 2020, 1, 2. [Google Scholar] [CrossRef]
- Zwierzchowski, G.; Zhang, G.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. Milk Metabotyping Identifies Metabolite Alterations in the Whole Raw Milk of Dairy Cows with Lameness. J. Agric. Food Chem. 2020, 68, 4507–4514. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, Å.; Persson Waller, K.; Svennersten-Sjaunja, K.; Haugen, J.-E.; Lundby, F.; Lind, O. Detection of Mastitic Milk Using a Gas-Sensor Array System (Electronic Nose). Int. Dairy J. 2005, 15, 1193–1201. [Google Scholar] [CrossRef]
- Hettinga, K.A.; van Valenberg, H.J.F.; Lam, T.J.G.M.; van Hooijdonk, A.C.M. Detection of Mastitis Pathogens by Analysis of Volatile Bacterial Metabolites. J. Dairy Sci. 2008, 91, 3834–3839. [Google Scholar] [CrossRef]
- Hettinga, K.A.; van Valenberg, H.J.F.; Lam, T.J.G.M.; van Hooijdonk, A.C.M. The Origin of the Volatile Metabolites Found in Mastitis Milk. Vet. Microbiol. 2009, 137, 384–387. [Google Scholar] [CrossRef]
- Sundekilde, U.K.; Poulsen, N.A.; Larsen, L.B.; Bertram, H.C. Nuclear Magnetic Resonance Metabonomics Reveals Strong Association between Milk Metabolites and Somatic Cell Count in Bovine Milk. J. Dairy Sci. 2013, 96, 290–299. [Google Scholar] [CrossRef]
- Ryman, V.E.; Pighetti, G.M.; Lippolis, J.D.; Gandy, J.C.; Applegate, C.M.; Sordillo, L.M. Quantification of Bovine Oxylipids during Intramammary Streptococcus Uberis Infection. Prostaglandins Other Lipid Mediat. 2015, 121, 207–217. [Google Scholar] [CrossRef]
- Giri, S.N.; Chen, Z.; Carroll, E.J.; Mueller, R.; Schiedt, M.J.; Panico, L. Role of Prostaglandins in Pathogenesis of Bovine Mastitis Induced by Escherichia Coli Endotoxin. Am. J. Vet. Res. 1984, 45, 586–591. [Google Scholar] [PubMed]
- Atroshi, F.; Työppönen, J.; Sankari, S.; Kangasniemi, R.; Parantainen, J. Possible Roles of Vitamin E and Glutathione Metabolism in Bovine Mastitis. Int. J. Vitam. Nutr. Res. 1987, 57, 37–43. [Google Scholar] [PubMed]
- Aitken, S.L.; Corl, C.M.; Sordillo, L.M. Immunopathology of Mastitis: Insights into Disease Recognition and Resolution. J. Mammary Gland. Biol. Neoplasia 2011, 16, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.C.; Mudaliar, M.; Tassi, R.; McNeilly, T.N.; Burchmore, R.; Burgess, K.; Herzyk, P.; Zadoks, R.N.; Eckersall, P.D. Mastitomics, the Integrated Omics of Bovine Milk in an Experimental Model of Streptococcus Uberis Mastitis: 3. Untargeted Metabolomics. Mol. BioSyst. 2016, 12, 2762–2769. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Eckmann, L. How Bile Acids Confer Gut Mucosal Protection against Bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 4333–4334. [Google Scholar] [CrossRef]
- Calmus, Y.; Poupon, R. Shaping Macrophages Function and Innate Immunity by Bile Acids: Mechanisms and Implication in Cholestatic Liver Diseases. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xie, W.; Krasowski, M.D. PXR: A Xenobiotic Receptor of Diverse Function Implicated in Pharmacogenetics. Pharmacogenomics 2008, 9, 1695–1709. [Google Scholar] [CrossRef]
- Sipka, S.; Bruckner, G. The Immunomodulatory Role of Bile Acids. Int. Arch. Allergy Immunol. 2014, 165, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC–MS Metabolomics Identifies Metabolite Alterations That Precede Subclinical Mastitis in the Blood of Transition Dairy Cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Zandkarimi, F.; Vanegas, J.; Fern, X.; Maier, C.S.; Bobe, G. Metabotypes with Elevated Protein and Lipid Catabolism and Inflammation Precede Clinical Mastitis in Prepartal Transition Dairy Cows. J. Dairy Sci. 2018, 101, 5531–5548. [Google Scholar] [CrossRef] [PubMed]
- Ten Bruggencate, S.J.; Bovee-Oudenhoven, I.M.; Feitsma, A.L.; van Hoffen, E.; Schoterman, M.H. Functional Role and Mechanisms of Sialyllactose and Other Sialylated Milk Oligosaccharides. Nutr. Rev. 2014, 72, 377–389. [Google Scholar] [CrossRef]
- Nakamura, T.; Kawase, H.; Kimura, K.; Watanabe, Y.; Ohtani, M.; Arai, I.; Urashima, T. Concentrations of Sialyloligosaccharides in Bovine Colostrum and Milk during the Prepartum and Early Lactation. J. Dairy Sci. 2003, 86, 1315–1320. [Google Scholar] [CrossRef]
- Hamann, J.; Krömker, V. Potential of Specific Milk Composition Variables for Cow Health Management. Livest. Prod. Sci. 1997, 48, 201–208. [Google Scholar] [CrossRef]
- Davis, S.R.; Farr, V.C.; Prosser, C.G.; Nicholas, G.D.; Turner, S.-A.; Lee, J.; Hart, A.L. Milk L-Lactate Concentration Is Increased during Mastitis. J. Dairy Res. 2004, 71, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Welderufael, B.G.; Løvendahl, P.; de Koning, D.-J.; Janss, L.L.G.; Fikse, W.F. Genome-Wide Association Study for Susceptibility to and Recoverability From Mastitis in Danish Holstein Cows. Front. Genet. 2018, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Gröhn, Y.T.; Wilson, D.J.; González, R.N.; Hertl, J.A.; Schulte, H.; Bennett, G.; Schukken, Y.H. Effect of Pathogen-Specific Clinical Mastitis on Milk Yield in Dairy Cows. J. Dairy Sci. 2004, 87, 3358–3374. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Contagious | Environmental | Opportunistic |
|---|---|---|
| Staphylococcus aureus | Escherichia coli | Non-aureus Staphylococci: |
| Streptococcus agalactiae | Streptococcus uberis | Staphylococcus simulans |
| Corynebacterium bovis | Streptococcus dysgalactiae | Staphylococcus chromogens |
| Mycoplasma spp | Klebsiella spp | |
| Corynebacterium pyogenes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haxhiaj, K.; Wishart, D.S.; Ametaj, B.N. Mastitis: What It Is, Current Diagnostics, and the Potential of Metabolomics to Identify New Predictive Biomarkers. Dairy 2022, 3, 722-746. https://doi.org/10.3390/dairy3040050
Haxhiaj K, Wishart DS, Ametaj BN. Mastitis: What It Is, Current Diagnostics, and the Potential of Metabolomics to Identify New Predictive Biomarkers. Dairy. 2022; 3(4):722-746. https://doi.org/10.3390/dairy3040050
Chicago/Turabian StyleHaxhiaj, Klevis, David S. Wishart, and Burim N. Ametaj. 2022. "Mastitis: What It Is, Current Diagnostics, and the Potential of Metabolomics to Identify New Predictive Biomarkers" Dairy 3, no. 4: 722-746. https://doi.org/10.3390/dairy3040050
APA StyleHaxhiaj, K., Wishart, D. S., & Ametaj, B. N. (2022). Mastitis: What It Is, Current Diagnostics, and the Potential of Metabolomics to Identify New Predictive Biomarkers. Dairy, 3(4), 722-746. https://doi.org/10.3390/dairy3040050