
Investigation of Acetoin Biosynthesis by Bacillus subtilis ACA-DC 1176 Growing on Crude Glycerol in Flask and Bioreactor Trials




Abstract
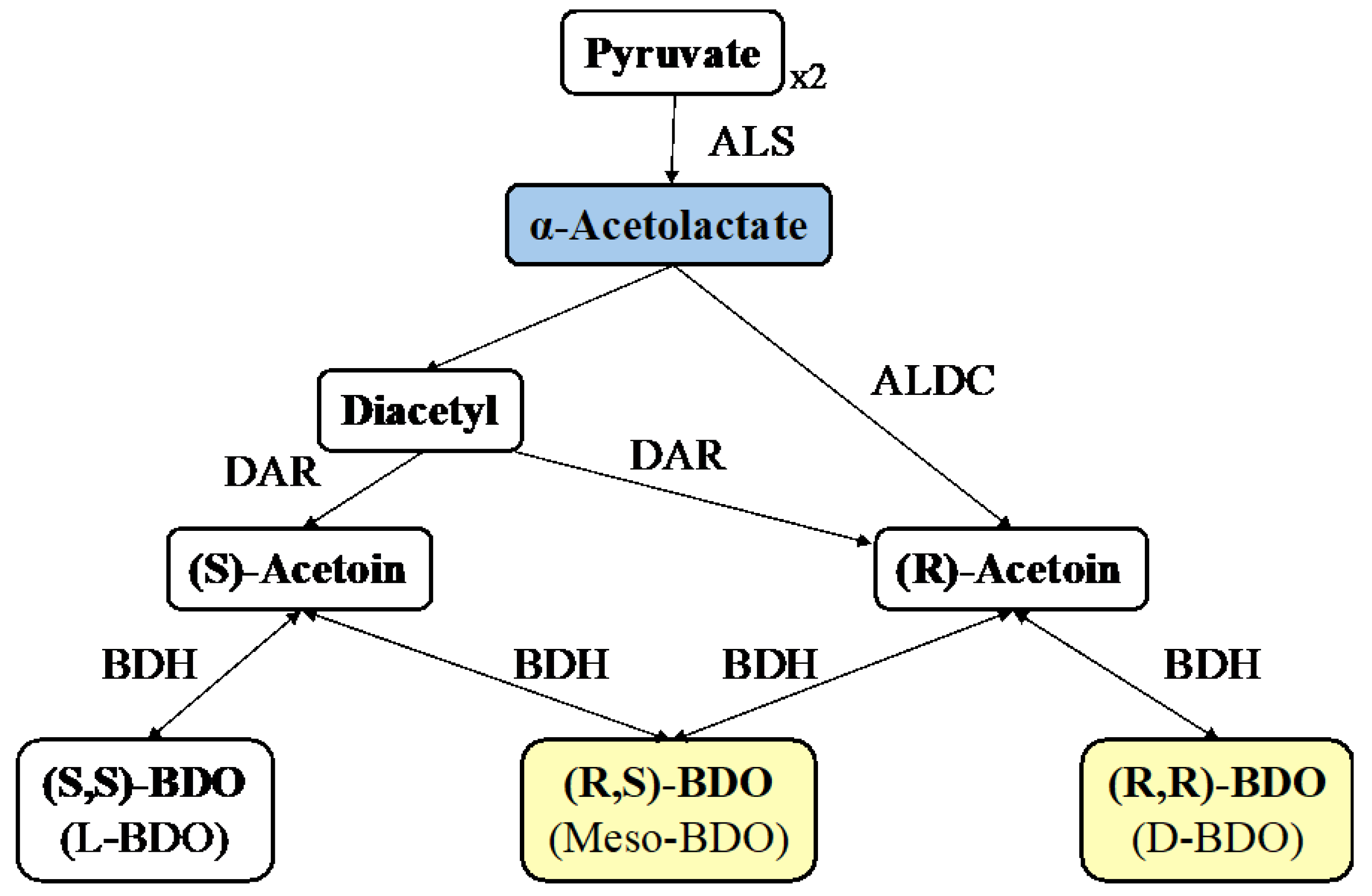
1. Introduction
2. Materials and Methods
2.1. Microorganism and Media
2.2. Cultures of B. subtilis
2.2.1. Preculture
2.2.2. Shake-Flask and Duran Bottle Cultures
2.2.3. Bioreactor Culture
2.3. Analytical Methods
2.3.1. Determination of Physicochemical Characteristics of Raw Materials
2.3.2. Growth Determination
2.3.3. Substrate and Metabolites Quantitative Determination
2.3.4. Estimation of kLa Value
2.4. Data Analysis
3. Results
3.1. Initial Trials on Acetoin Production Feasibility
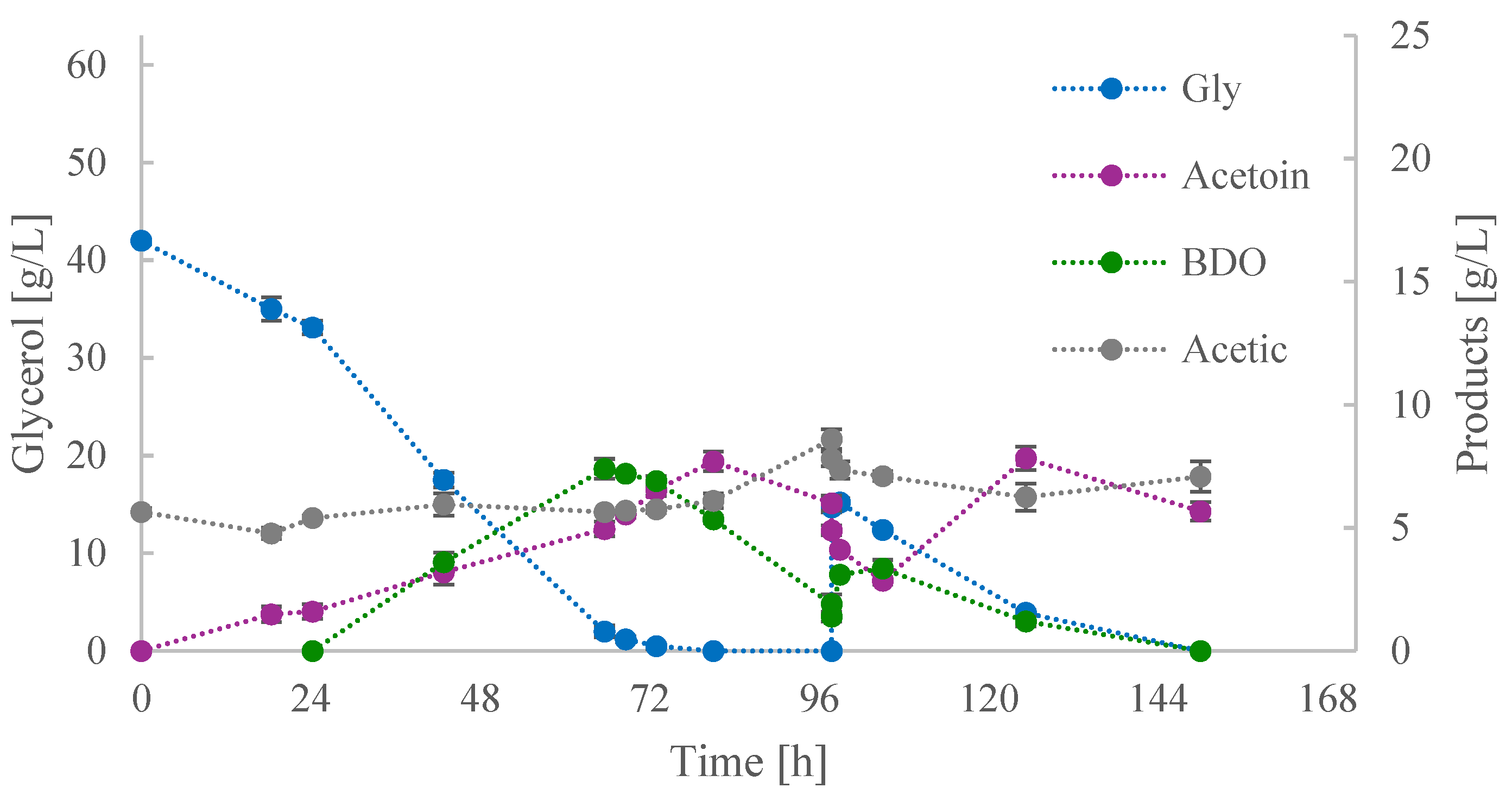
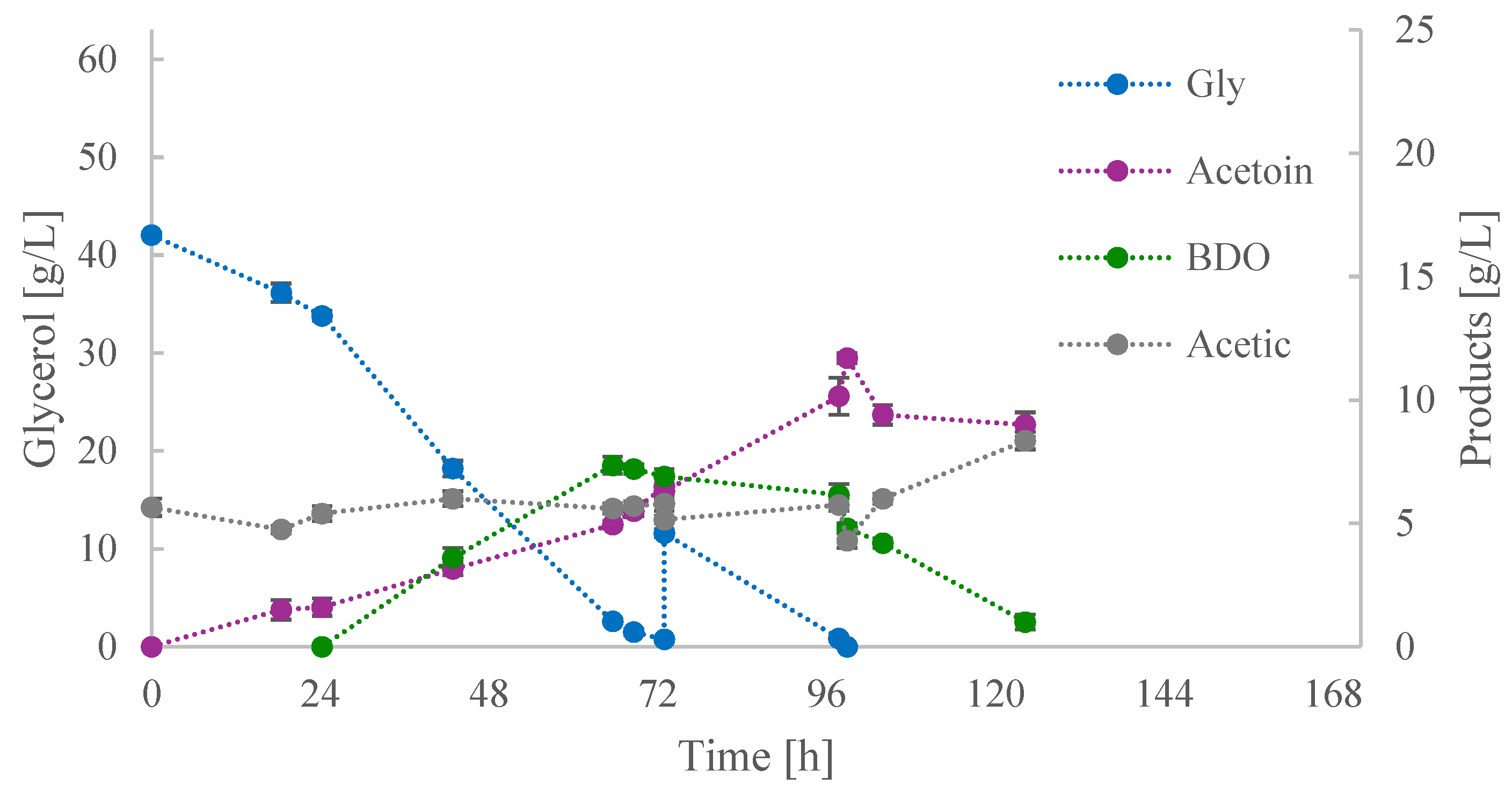
3.2. Investigation of Acetoin Production by B. subtilis ACA-DC 1176 in Shake-Flask Cultures
3.2.1. Culture Medium and Temperature
3.2.2. Feeding Strategy
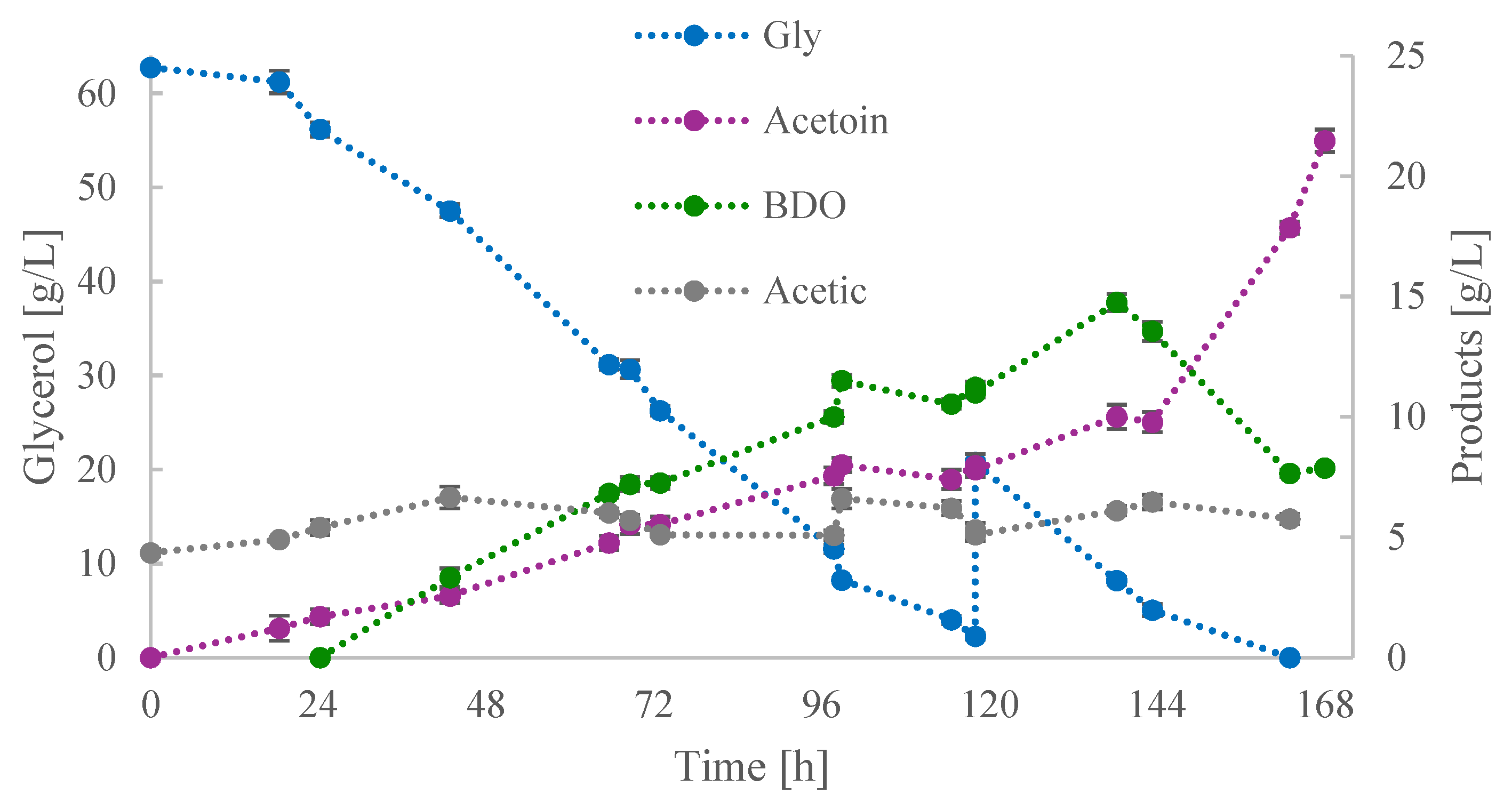
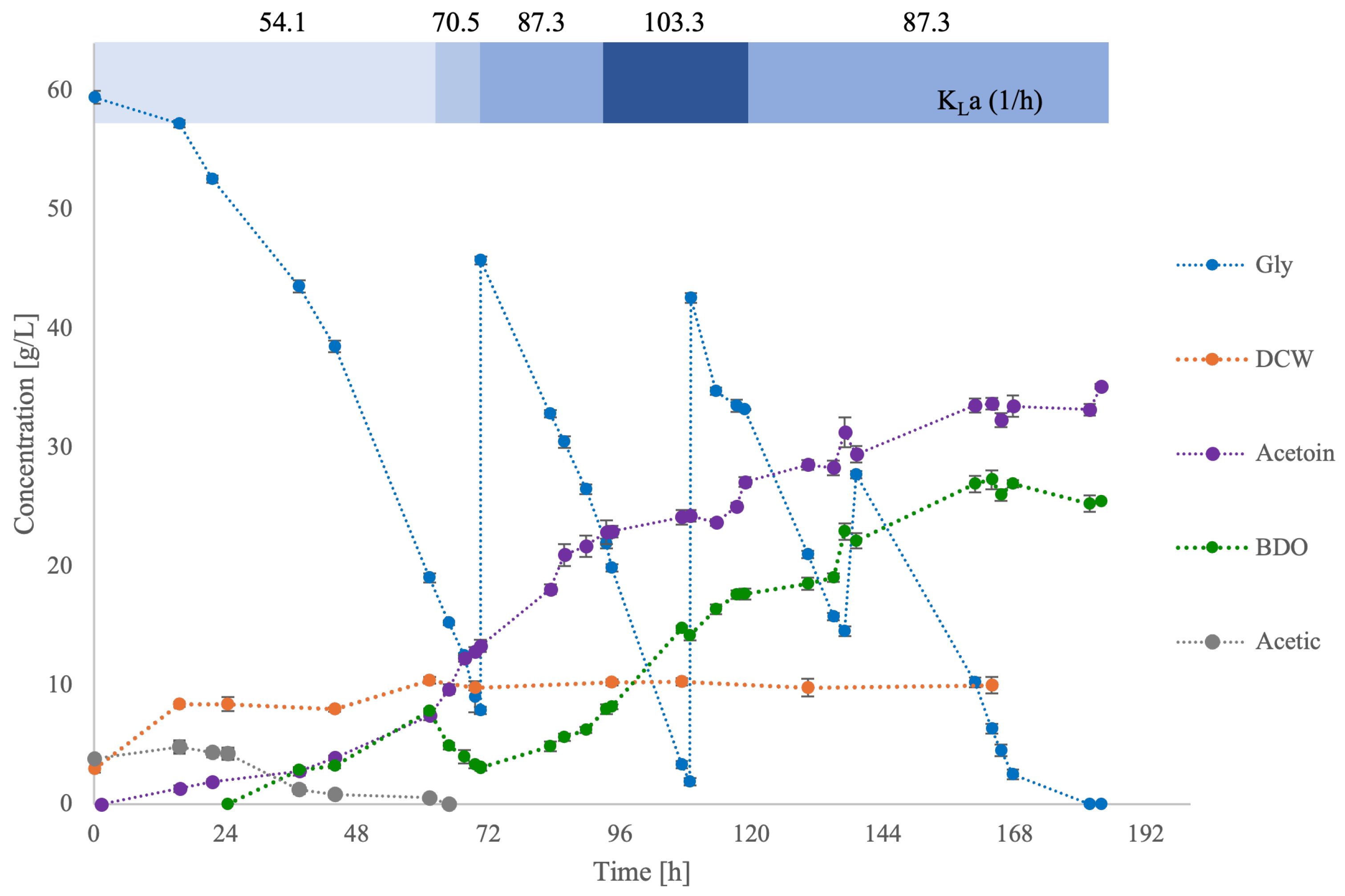
3.3. Scale-Up in Bioreactor Culture
4. Concluding Remarks
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Koutinas, A.A.; Vlysidis, A.; Pleissner, D.; Kopsahelis, N.; Garcia, I.L.; Kookos, I.K.; Lin, C.S.K. Valorization of industrial waste and by-product streams via fermentation for the production of chemicals and biopolymers. Chem. Soc. Rev. 2014, 8, 2587–2627. [Google Scholar] [CrossRef] [PubMed]
- Maina, S.; Kachrimanidou, V.; Koutinas, A.A. A roadmap towards a circular and sustainable bioeconomy through waste valorization. Curr. Opin. Green Sustain. Chem. 2017, 8, 18–23. [Google Scholar] [CrossRef]
- Vasilakis, G.; Karayannis, D.; Massouras, T.; Politis, I.; Papanikolaou, S. Biotechnological conversions of Mizithra second cheese whey by wild-type non-conventional yeast strains: Production of yeast cell biomass, single-cell oil and polysaccharides. Appl. Sci. 2022, 12, 11471. [Google Scholar] [CrossRef]
- The Brainy Insights. Biodiesel Market Size, Share & Trends Analysis Report by Feedstock (Vegetable Oils, Animal Fats), by Application (Fuel, Power Generation), by Region (Europe, APAC), and Segment Forecasts, 2022–2030. Available online: https://www.researchandmarkets.com/reports/4375442/biodiesel-market-size-share-and-trends-analysis?srsltid=AfmBOoqb4ZMkLeq4ksLwxTvW2m4iEQ94cng7BwLI3VMNg_4tevNoaxKH (accessed on 10 July 2024).
- Papanikolaou, S.; Aggelis, G. Microbial products from wastes and residues. FEMS Microbiol. Lett. 2020, 367, fnaa156. [Google Scholar] [CrossRef]
- Zhang, C.; Sharma, S.; Ma, C.; Zeng, A.P. Strain evolution and novel downstream processing with integrated catalysis enable highly efficient coproduction of 1,3-propanediol and organic acid esters from crude glycerol. Biotechnol. Bioeng. 2022, 119, 1450–1466. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Papanikolaou, S. Effect of impurities in biodiesel-derived waste glycerol on the performance and feasibility of biotechnological processes. Appl. Microbiol. Biotechnol. 2012, 95, 13–27. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Rontou, M.; Belka, A.; Athenaki, M.; Gardeli, C.; Mallouchos, A.; Kalantzi, O.; Koutinas, A.A.; Kookos, I.K.; Zeng, A.P.; et al. Conversion of biodiesel-derived glycerol into biotechnological products of industrial significance by yeast and fungal strains. Eng. Life Sci. 2016, 17, 262–281. [Google Scholar] [CrossRef]
- Karayannis, D.; Papanikolaou, S.; Vatistas, C.; Paris, C.; Chevalot, I. Yeast lipid produced through glycerol conversions and its use for enzymatic synthesis of amino acid-based biosurfactants. Int. J. Mol. Sci. 2023, 24, 714. [Google Scholar] [CrossRef]
- Wang, Z.; Zhuge, J.; Fang, H.; Prior, B.A. Glycerol production by microbial fermentation: A review. Biotechnol. Adv. 2001, 19, 201–223. [Google Scholar] [CrossRef]
- Vastaroucha, E.S.; Maina, S.; Michou, S.; Kalantzi, O.; Pateraki, C.; Koutinas, A.A.; Papanikolaou, S. Bioconversions of biodiesel-derived glycerol into sugar alcohols by newly isolated wild-type Yarrowia lipolytica strains. Reaction. 2021, 2, 499–513. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; U.S. Department of Energy Office of Scientific and Technical Information: Oak Ridge, TN, USA, 2004. [Google Scholar]
- Market Research. Global Acetoin Market Research Report 2023. Available online: https://www.marketresearch.com/Maia-Research-v4212/Global-Acetoin-Research-Competitive-Status-34497228/ (accessed on 10 July 2024).
- Petrov, K.; Petrova, P. Current Advances in Microbial Production of Acetoin and 2,3-Butanediol by Bacillus spp. Fermentation 2021, 7, 307. [Google Scholar] [CrossRef]
- Al-Auda, Z.; Li, X.; Hohn, K.L. Dehydrogenation of 2,3-butanediol to acetoin using copper catalysts. Ind. Eng. Chem. Res. 2022, 61, 3530–3538. [Google Scholar] [CrossRef]
- Forlani, G.; Mantelli, M.; Nielsen, E. Biochemical evidence for multiple acetoin-forming enzymes in cultured plant cells. Phytochemistry 1999, 50, 255–262. [Google Scholar] [CrossRef]
- Romano, P.; Suzzi, G. Origin and production of acetoin during wine yeast fermentation. Appl. Environ. Microbiol. 1996, 62, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Lu, J.R. Strategies for enhancing fermentative production of acetoin: A review. Biotechnol. Adv. 2014, 32, 492–503. [Google Scholar] [CrossRef]
- MacFaddin, J.F. Biochemical Tests for Identification of Medical Bacteria; Williams and Wilkins: Baltimore, MD, USA, 1980. [Google Scholar]
- Wang, Q.; Zhang, X.; Ren, K.; Han, R.; Lu, R.; Bao, T.; Pan, X.; Yang, T.; Xu, M.; Rao, Z. Acetoin production from lignocellulosic biomass hydrolysates with a modular metabolic engineering system in Bacillus subtilis. Biotechnol. Biofuels Bioprod. 2022, 15, 87. [Google Scholar] [CrossRef]
- Celinska, E.; Grajek, W. Biotechnological production of 2,3-butanediol--current state and prospects. Biotechnol. Adv. 2009, 27, 715–725. [Google Scholar] [CrossRef]
- Chen, C.; Wei, D.; Shi, J.; Wang, M.; Hao, J. Mechanism of 2,3-butanediol stereoisomer formation in Klebsiella pneumoniae. Appl. Microbiol. Biotechnol. 2014, 98, 4603–4613. [Google Scholar] [CrossRef]
- Karayannis, D.; Aggelou, N.; Vasilakis, G.; Charisteidis, I.; Litinas, A.; Papanikolaou, S. A non-aseptic bioprocess for production and recovery of 2,3-butanediol via conversion of crude glycerol and corn steep liquor at pilot-scale. Carbon Resour. Convers. 2024, in press. [Google Scholar] [CrossRef]
- Maina, S.; Mallouchos, A.; Nychas, G.J.E.; Freire, D.M.; de Castro, A.M.; Papanikolaou, S.; Kookos, I.K.; Koutinas, A. Bioprocess development for (2R, 3R)-butanediol and acetoin production using very high polarity cane sugar and sugarcane molasses by a Bacillus amyloliquefaciens strain. J. Chem. Technol. Biotechnol. 2019, 94, 2167–2177. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Yang, T.; Zhang, J.; Xu, M.; Li, H.; Xu, Z.; Rao, Z. Mutation breeding of acetoin high producing Bacillus subtilis blocked in 2,3-butanediol dehydrogenase. World J. Microbiol. Biotechnol. 2013, 29, 1783–1789. [Google Scholar] [CrossRef] [PubMed]
- Tsigoriyna, L.; Petrova, P.; Petrov, K. High production of acetoin from glycerol by Bacillus subtilis 35. Appl. Microbiol. Biotechnol. 2022, 107, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.J.; Liu, P.H.; Qin, J.Y.; Xu, P. Statistical optimization of medium components for enhanced acetoin production from molasses and soybean meal hydrolysate. Appl. Microbiol. Biotechnol. 2007, 75, 61–68. [Google Scholar] [CrossRef]
- Dai, J.Y.; Cheng, L.; He, Q.F.; Xiu, Z.L. High acetoin production by a newly isolated marine Bacillus subtilis strain with low requirement of oxygen supply. Process Biochem. 2015, 50, 1730–1734. [Google Scholar] [CrossRef]
- Gingichashvili, S.; Duanis-Assaf, D.; Shemesh, M.; Featherstone, J.D.B.; Feuerstein, O.; Steinberg, D. Bacillus subtilis Biofilm Development—A Computerized Study of Morphology and Kinetics. Front. Microbiol. 2017, 8, 2072. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Hobley, L.; Ostrowski, A.; Rao, F.V.; Bromley, K.M.; Porter, M.; Prescott, A.R.; MacPhee, C.E.; van Aalten, D.M.; Stanley-Wall, N.R. BslA is a self-assembling bacterial hydrophobin that coats the Bacillus subtilis biofilm. Proc. Natl. Acad. Sci. USA 2013, 110, 13600–13605. [Google Scholar] [CrossRef]
- Kobayashi, K.; Iwano, M. BslA (YuaB) forms a hydrophobic layer on the surface of Bacillus subtilis biofilms. Mol. Microbiol. 2012, 85, 51–66. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef]
- Arnaouteli, S.; Bamford, N.C.; Stanley-Wall, N.R.; Kovács, A.T. Bacillus subtilis biofilm formation and social interactions. Nat. Rev. Microbiol. 2021, 19, 600–614. [Google Scholar] [CrossRef]
- Costerton, J.W.; Lewandowski, Z.; Caldwell, D.E.; Korber, D.R.; Lappin-Scott, H.M. Microbial biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Liu, C.; Fang, H.; Zhang, D. Bacillus subtilis: A universal cell factory for industry, agriculture, biomaterials and medicine. Microb. Cell Fact. 2020, 19, 173. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.; Zhang, Y.; Zhou, B.; Qin, Z.; Wu, J.; Wang, Q.; Yin, Y. Effects of Bacillus subtilis on carbon components and microbial functional metabolism during cow manure-straw composting. Bioresour. Technol. 2020, 303, 122868. [Google Scholar] [CrossRef]
- Aroniada, M.; Maina, S.; Koutinas, A.; Kookos, I.K. Estimation of volumetric mass transfer coefficient (kLa)—Review of classical approaches and contribution of a novel methodology. Biochem. Eng. J. 2020, 155, 107458. [Google Scholar] [CrossRef]
- ISO 8968-1; Milk Determination of Nitrogen Content–Part 1: Kjeldahl Method. First Edition IDF 20-1; ISO: Geneva, Switzerland, 2011. Available online: https://www.iso.org/standard/35120.html (accessed on 10 June 2024).
- Lee, Y.H.; Tsao, G.T. Dissolved oxygen electrodes. Adv. Biochem. Eng. 1979, 13, 35–86. [Google Scholar]
- Karayannis, D.; Vasilakis, G.; Charisteidis, I.; Litinas, A.; Manolopoulou, E.; Tsakalidou, E.; Papanikolaou, S. Screening of mew industrially important bacterial strains for 1,3-propanediol, 2,3-butanediol and ethanol production through biodiesel-derived glycerol fermentations. Microorganisms 2023, 11, 1424. [Google Scholar] [CrossRef]
- Metsoviti, M.; Zeng, A.P.; Koutinas, A.A.; Papanikolaou, S. Enhanced 1,3-propanediol production by a newly isolated Citrobacter freundii strain cultivated on biodiesel-derived waste glycerol through sterile and non-sterile bioprocesses. J. Biotechnol. 2013, 163, 408–418. [Google Scholar] [CrossRef]
- Maina, S.; Prabhu, A.A.; Vivek, N.; Vlysidis, A.; Koutinas, A.; Kumar, V. Prospects on bio-based 2,3-butanediol and acetoin production: Recent progress and advances. Biotechnol. Adv. 2022, 54, 107783. [Google Scholar] [CrossRef]
- Maina, S.; Schneider, R.; Alexandri, M.; Papapostolou, H.; Nychas, G.J.; Koutinas, A.; Venus, J. Volumetric oxygen transfer coefficient as fermentation control parameter to manipulate the production of either acetoin or D-2,3-butanediol using bakery waste. Bioresour. Technol. 2021, 335, 125155. [Google Scholar] [CrossRef]
- Danilova, I.; Sharipova, M. The practical potential of Bacilli and their enzymes for industrial production. Front. Microbiol. 2020, 11, 1782. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxygen Regime | Carbon Source | B. subtilis ACA-DC 1176 | B. subtilis ACA-DC 1225 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Acetoin (g/L) | BDO (g/L) | DCW (g/L) | SubC (%) | Time (h) | Acetoin (g/L) | BDO (g/L) | DCW (g/L) | SubC (%) | Time (h) | ||
| Aerobiosis | |||||||||||
| Glucose | 3.8 | 3.3 | 3.4 | 100 | 45 | 4.9 | 2.5 | 3.8 | 100 | 48 | |
| Glycerol | 3.5 | 1.2 | 5.7 | 100 | 44 | 2.5 | 1.3 | 5.8 | 100 | 55 | |
| Crude glycerol | 2.6 | 1.3 | 5.0 | 100 | 45 | 2.4 | 0.8 | 4.8 | 100 | 70 | |
| Anaerobiosis | |||||||||||
| Glycerol | - | - | 1.1 | 12 | - | Neg | - | 3.8 | 50 | 70 | |
| Crude glycerol | Neg | - | 1.1 | 15 | 70 | - | - | Neg | - | - | |
| No. | Gly Type | Nitrogen | Temp | Gly0 | Acetoin | BDO | DCW | YACTN/Gly | ACTN/BDO | ConRate | Time | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | ComOrg | CSL | (°C) | (g/L) | (g/L) | (g/L) | (g/L) | (g/g) | (g/g) | (g/L/h) | (h) | ||
| 1 | Pu. | 100% | 100% | - | 30 | 43.3 | 9.0 | 4.4 | 12.7 | 0.21 | 9.0 | 0.50 | 83 |
| 2 | 67.0 | 17.5 | 6.1 | 14.3 | 0.27 | 6.3 | 0.54 | 116 | |||||
| 3 | Pu. | 100% | 50% | 50% | 30 | 40.6 | 11.2 | 4.5 | 6.9 | 0.28 | 7.8 | 0.57 | 93 |
| 4 | 61.9 | 5.6 | 13.8 | 5.1 | 0.12 | 0.4 | 0.49 | 135 | |||||
| 5 | Pu. | 50% | 50% | - | 30 | 41.0 | 2.0 | 2.2 | 5.2 | Neg | 1.1 | 0.27 | 157 |
| 6 | 62.5 | 1.9 | 0.9 | 4.2 | Neg | 1.5 | 0.37 | 184 | |||||
| 7 | Cr. | 100% | 100% | - | 30 | 38.3 | 8.5 | 4.2 | 10.3 | 0.22 | 8.0 | 0.51 | 81 |
| 8 | 60.1 | 10.1 | 6.9 | 11.0 | 0.17 | 7.1 | 0.56 | 128 | |||||
| 9 | Cr. | 100% | 50% | 50% | 30 | 42.8 | 8.0 | 3.9 | 13.1 | 0.19 | 8.0 | 0.54 | 85 |
| 10 | 62.4 | 5.8 | 6.7 | 14.0 | 0.11 | 0.9 | 0.48 | 136 | |||||
| 11 | Cr. | 100% | 50% | 50% | 37 | 44.0 | 11.2 | 6.5 | 7.4 | 0.25 | 1.0 | 0.49 | 90 |
| 12 | 63.9 | 12.5 | 7.5 | 9.6 | 0.20 | 1.5 | 0.49 | 120 | |||||
| 13 | Cr. | 50% | 50% | - | 37 | 41.0 | 1.7 | 3.3 | 4.4 | Neg | 0.9 | 0.31 | 145 |
| 14 | 62.5 | 2.0 | 3.7 | 5.6 | Neg | 0.5 | 0.42 | 150 | |||||
| Time Periods (h) | kLa (1/h) | ConRate (g/L/h) | ACTN (g/L) | BDO (g/L) | DCW (g/L) | ACTN/BDO (g/g) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Δt | t | Δ | Total | Δ | Total | |||||
| 1 | 0–70.5 | 64.7 | 54.1 | 0.80 | +7.4 | 7.4 | +7.8 | 7.8 | 10.4 | 0.94 |
| 70.5 | 70.5 | 1.20 | +5.9 | 13.3 | −4.7 | 3.1 | 10.6 | 4.29 | ||
| 2 | 70.5–108.7 | 94.5 | 87.3 | 1.00 | +9.8 | 23.1 | +4.9 | 8.0 | 10.4 | 2.22 |
| 108.7 | 103.3 | 1.30 | +1.4 | 24.5 | +6.2 | 14.2 | 10.5 | 1.73 | ||
| 3 | 108.7–137 | 130.2 | 103.3 | 0.90 | +3.4 | 27.9 | +3.5 | 17.7 | 10.0 | 1.58 |
| 137 | 87.3 | 1.00 | +4.2 | 32.1 | +5.2 | 22.9 | 10.4 | 1.40 | ||
| 4 | 137–183.7 | 167.7 | 87.3 | 0.80 | +2.2 | 34.3 | +4 | 26.9 | 10.9 | 1.28 |
| 183.7 | 0.20 | +1.7 | 36.0 | −1.4 | 25.5 | 10.5 | 1.41 | |||
| Sum | 183.7 | 0.88 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karayannis, D.; Moraiti, E.; Kalantzi, O.; Papanikolaou, S. Investigation of Acetoin Biosynthesis by Bacillus subtilis ACA-DC 1176 Growing on Crude Glycerol in Flask and Bioreactor Trials. Reactions 2024, 5, 664-679. https://doi.org/10.3390/reactions5040034
Karayannis D, Moraiti E, Kalantzi O, Papanikolaou S. Investigation of Acetoin Biosynthesis by Bacillus subtilis ACA-DC 1176 Growing on Crude Glycerol in Flask and Bioreactor Trials. Reactions. 2024; 5(4):664-679. https://doi.org/10.3390/reactions5040034
Chicago/Turabian StyleKarayannis, Dimitris, Eleni Moraiti, Ourania Kalantzi, and Seraphim Papanikolaou. 2024. "Investigation of Acetoin Biosynthesis by Bacillus subtilis ACA-DC 1176 Growing on Crude Glycerol in Flask and Bioreactor Trials" Reactions 5, no. 4: 664-679. https://doi.org/10.3390/reactions5040034
APA StyleKarayannis, D., Moraiti, E., Kalantzi, O., & Papanikolaou, S. (2024). Investigation of Acetoin Biosynthesis by Bacillus subtilis ACA-DC 1176 Growing on Crude Glycerol in Flask and Bioreactor Trials. Reactions, 5(4), 664-679. https://doi.org/10.3390/reactions5040034