Receptor Regulation in Taste: Can Diet Influence How We Perceive Foods?


Abstract
1. Taste and Taste Buds
2. Receptor Regulation
3. Evidence for Receptor Regulation in the Perception of Taste
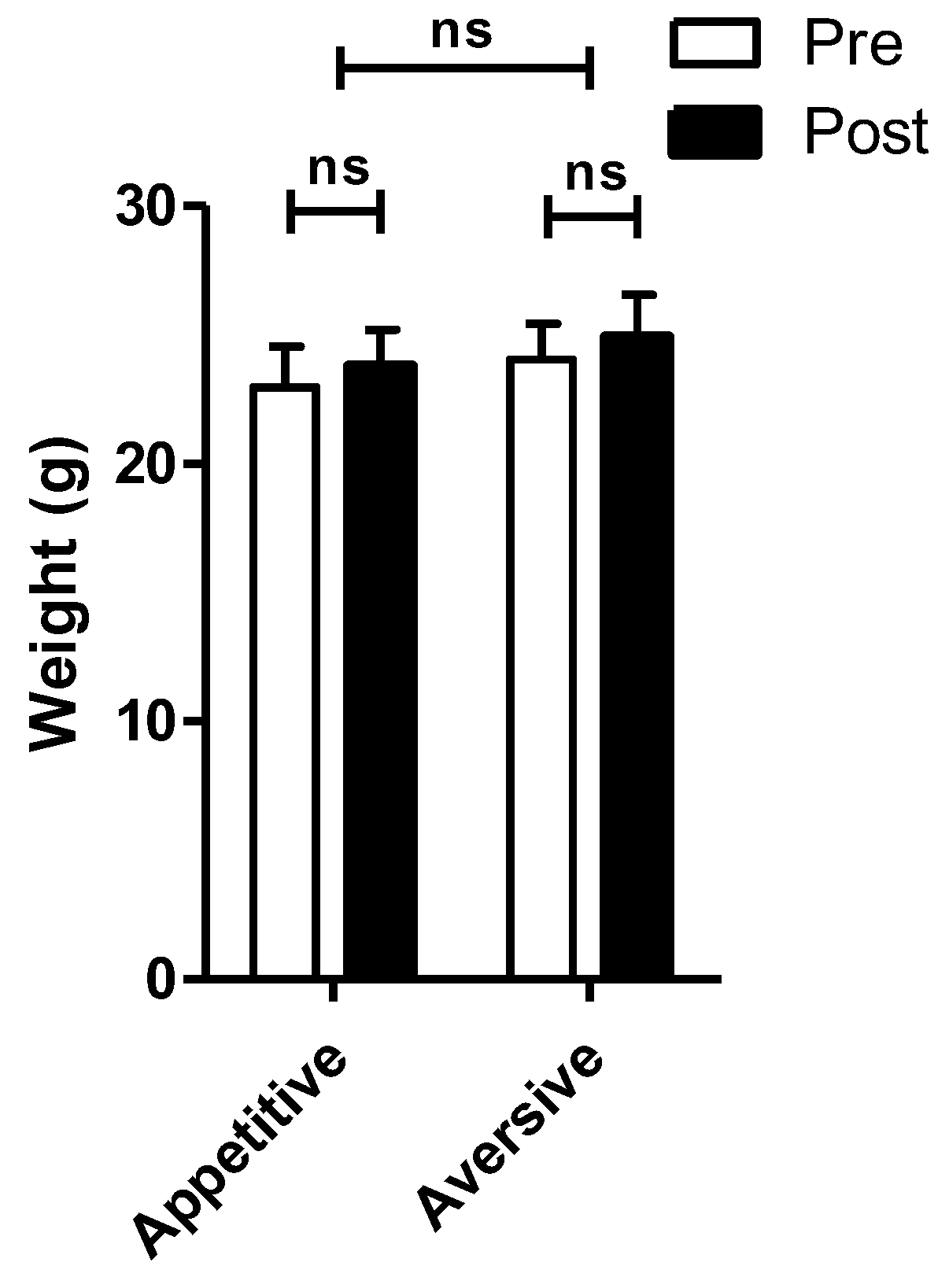
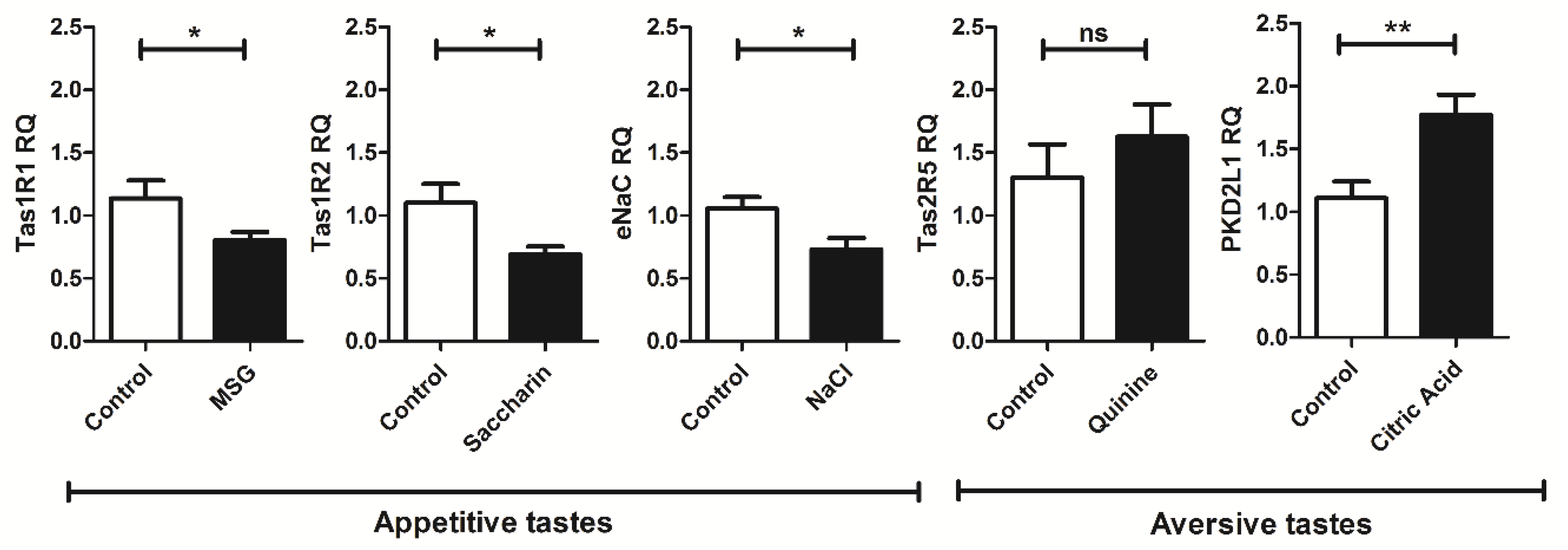
4. Exposure to Appetitive but Not Aversive Taste Stimuli Decreases mRNA Expression of Cogent Receptors
5. Implications for Chemosensory Perception and Food Choice
6. Materials and Methods
6.1. Animals
6.2. Dietary Treatment
6.3. Taste Bud Isolation
6.4. qRT–PCR
6.5. Data Analysis
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Gene | Direction | Sequence (5′ to 3′) |
|---|---|---|
| Β-actin | Forward | CACCCTGTGCTGC |
| Reverse | GCACGATTTCCCT | |
| PLCβ2 | Forward | GAGCAAATCGCCA |
| Reverse | CCTTGTCTGTGGT | |
| Tas1R1 | Forward | GTGGAATGGACCTGAATGGAC |
| Reverse | AGCAGCAGTGGTGGGAAC | |
| Tas1R2 | Forward | AAGCATCGCCTCCTACTCC |
| Reverse | GGCTGGCAACTCTTAGAACAC | |
| Tas2R5 | Forward | GAATCATAGAAACAGGACCTCG |
| Reverse | CTTTACAAAGGCTTGCTTTAGC | |
| PKD2L1 | Forward | TGCGCCTGAGGAAAGAGCGG |
| Reverse | AAGCGGCCGCTAGGACGGATTATACA | |
| ENaC | Forward | GGCAGCCCACCGAGGAGGA |
| Reverse | GCCACAGCACCGCCCAGAA | |
| Gustducin | Forward | TGTGCCAAATGAACAAGACGTTCT |
| Reverse | CTGATCTCTGGCCACCTACATCAA | |
| TRPM5 | Forward | GTCTGGAATCACAGGCCAAC |
| Reverse | GGTGATGTGCCCCAAAAACT | |
| Protein Kinase C | Forward | TGAATCCTCAGTGGAATGAGT |
| Reverse | GGTTGCTTTCTGTCTTCTGAA |
References
- Running, C.A.; Craig, B.A.; Mattes, R.D. Oleogustus: The unique taste of fat. Chem. Senses 2015, 40, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Vandenbeuch, A.; Clapp, T.R.; Kinnamon, S.C. Amiloride-sensitive channels in type I fungiform taste cells in mouse. BMC Neurosci. 2008, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Heck, G.L.; Mierson, S.; DeSimone, J.A. Salt taste transduction occurs through an amiloride-sensitive sodium transport pathway. Science 1984, 223, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, J.; Kuhn, C.; Oka, Y.; Yarmolinsky, D.A.; Hummler, E.; Ryba, N.J.; Zuker, C.S. The cells and peripheral representation of sodium taste in mice. Nature 2010, 464, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Tomchik, S.M.; Berg, S.; Kim, J.W.; Chaudhari, N.; Roper, S.D. Breadth of Tuning and Taste Coding in Mammalian Taste Buds. J. Neurosci. 2007, 27, 10840–10848. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.; Hoon, M.A.; Chandrashekar, J.; Zhang, Y.; Ryba, N.J.; Zuker, C.S. Mammalian sweet taste receptors. Cell 2001, 106, 381–390. [Google Scholar] [CrossRef]
- Di Pizio, A.; Levit, A.; Slutzki, M.; Behrens, M.; Karaman, R.; Niv, M.Y. Comparing Class A GPCRs to bitter taste receptors: Structural motifs, ligand interactions and agonist-to-antagonist ratios. Methods Cell Biol. 2016, 132, 401–427. [Google Scholar] [PubMed]
- Zhao, G.Q.; Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Erlenbach, I.; Ryba, N.J.; Zuker, C.S. The receptors for mammalian sweet and umami taste. Cell 2003, 115, 255–266. [Google Scholar] [CrossRef]
- Kuhn, C.; Meyerhof, W. Oligomerization of sweet and bitter taste receptors. Methods Cell Biol. 2013, 117, 229–242. [Google Scholar] [PubMed]
- Huang, Y.A.; Maruyama, Y.; Stimac, R.; Roper, S.D. Presynaptic (Type III) cells in mouse taste buds sense sour (acid) taste. J. Physiol. 2008, 586, 2903–2912. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Inada, H.; Kubota, M.; Zhuang, H.; Tominaga, M.; Matsunami, H. Transient receptor potential family members PKD1L3 and PKD2L1 form a candidate sour taste receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 12569–12574. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.L.; Chen, X.; Hoon, M.A.; Chandrashekar, J.; Guo, W.; Tränkner, D.; Ryba, N.J.; Zuker, C.S. The cells and logic for mammalian sour taste detection. Nature 2006, 442, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, S.; Yang, R.; Ishimaru, Y.; Matsunami, H.; Sévigny, J.; Kinnamon, J.C.; Finger, T.E. The candidate sour taste receptor, PKD2L1, is expressed by type III taste cells in the mouse. Chem. Senses 2008, 33, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Horio, N.; Yoshida, R.; Yasumatsu, K.; Yanagawa, Y.; Ishimaru, Y.; Matsunami, H.; Ninomiya, Y. Sour taste responses in mice lacking PKD channels. PLoS ONE 2011, 6, e20007. [Google Scholar] [CrossRef] [PubMed]
- Shankaran, H.; Wiley, H.S.; Resat, H. Receptor downregulation and desensitization enhance the information processing ability of signalling receptors. BMC Syst. Biol. 2007, 1, 48. [Google Scholar] [CrossRef] [PubMed]
- Knutson, V.P.; Ronnett, G.V.; Lane, M.D. Rapid, reversible internalization of cell surface insulin receptors. Correlation with insulin-induced down-regulation. J. Biol. Chem. 1983, 258, 12139–12142. [Google Scholar] [PubMed]
- Gavin, J.R.; Roth, J.; Neville, D.M.; De Meyts, P.; Buell, D.N. Insulin-dependent regulation of insulin receptor concentrations: A direct demonstration in cell culture. Proc. Natl. Acad. Sci. USA 1974, 71, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Kahn, C.R.; Neville, D.M.; Roth, J. Insulin-receptor interaction in the obese-hyperglycemic mouse a model of insulin resistance. J. Biol. Chem. 1973, 248, 244–250. [Google Scholar] [PubMed]
- Kahn, C.R.; Neville, D.M.; Gorden, P.; Freychet, P.; Roth, J. Insulin receptor defect in insulin resistance: Studies in the obese-hyperglycemic mouse. Biochem. Biophys. Res. Commun. 1972, 48, 135–142. [Google Scholar] [CrossRef]
- Soll, A.H.; Kahn, C.R.; Neville, D.M. Insulin binding to liver plasm membranes in the obese hyperglycemic (ob/ob) mouse. Demonstration of a decreased number of functionally normal receptors. J. Biol. Chem. 1975, 250, 4702–4707. [Google Scholar] [PubMed]
- Petersen, B.; Beckner, S.; Blecher, M. Hormone receptors 7. Characteristics of insulin receptors in a new line of cloned neonatal rat hepatocytes. Biochim. Biophys. Acta (BBA) Gen. Subj. 1978, 542, 470–485. [Google Scholar] [CrossRef]
- Bar, R.S.; Gorden, P.H.; Roth, J.E.; Kahn, C.R.; De Meyts, P.I. Fluctuations in the affinity and concentration of insulin receptors on circulating monocytes of obese patients: Effects of starvation, refeeding, and dieting. J. Clin. Investig. 1976, 58, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Cidlowski, N.B.; Cidlowski, J.A. Regulation of Glucocorticoid Receptors by Glucocorticoids in Cultured HeLa S3 Cells. Endocrinology 1981, 109, 1975–1982. [Google Scholar] [CrossRef] [PubMed]
- Schlechte, J.A.; Ginsberg, B.H.; Sherman, B.M. Regulation of the glucocorticoid receptor in human lymphocytes. J. Steroid Biochem. 1982, 16, 69–74. [Google Scholar] [CrossRef]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.E.; Lee, G.H.; Zhang, Y.; Kern, P.A. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.L.; Perez, E.; He, Y.J.; Dawson, R.; Millard, W.J. Leptin resistance is associated with hypothalamic leptin receptor mRNA and protein downregulation. Metabolism 2000, 49, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Kebabian, J.W.; Zatz, M.; Romero, J.A.; Axelrod, J. Rapid changes in rat pineal beta-adrenergic receptor: Alterations in l-(3H) alprenolol binding and adenylate cyclase. Proc. Natl. Acad. Sci. USA 1975, 72, 3735–3739. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, R.J.; Mullikin, D.; Wood, C.L.; Gore, T.B.; Mukherjee, C.H. Regulation of prostaglandin receptors by prostaglandins and guanine nucleotides in frog erythrocytes. J. Biol. Chem. 1977, 252, 5295–5303. [Google Scholar] [PubMed]
- Ren, X.; Zhou, L.; Terwilliger, R.; Newton, S.; De Araujo, I.E. Sweet taste signaling functions as a hypothalamic glucose sensor. Front. Integr. Neurosci. 2009, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Nakagawa, Y.; Ma, J.; Li, L.; Hamano, K.; Akimoto, T.; Kojima, I. Expression of the glucose-sensing receptor TAS1R3 in pancreatic islet: Changes in the expression levels in various nutritional and metabolic states. Endocr. J. 2014, 61, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.R.; Hussain, T.; Azari, E.K.; Steiner, J.L.; Ayala, J.E.; Pratley, R.E.; Kyriazis, G.A. Disruption of the sugar-sensing receptor TAS1R2 attenuates metabolic derangements associated with diet-induced obesity. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E688–E698. [Google Scholar] [CrossRef] [PubMed]
- Perea-Martinez, I.; Nagai, T.; Chaudhari, N. Functional Cell Types in Taste Buds Have Distinct Longevities. PLoS ONE 2013, 8, e53399. [Google Scholar] [CrossRef] [PubMed]
- Bertino, M.; Beauchamp, G.K.; Engelman, K. Long-term reduction in dietary sodium alters the taste of salt. Am. J. Clin. Nutr. 1982, 36, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Bertino, M.; Beauchamp, G.K.; Engelman, K. Increasing dietary salt alters salt taste preference. Physiol. Behav. 1986, 38, 203–213. [Google Scholar] [CrossRef]
- Pangborn, R.M.; Pecore, S.D. Taste perception of sodium chloride in relation to dietary intake of salt. Am. J. Clin. Nutr. 1982, 35, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M.; Nattress, L.; Flammer, L.J.; Beauchamp, G.K. Reduced dietary intake of simple sugars alters perceived sweet taste intensity but not perceived pleasantness. Am. J. Clin. Nutr. 2016, 103, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Noel, C.A.; Sugrue, M.; Dando, R. Participants with pharmacologically impaired taste function seek out more intense, higher calorie stimuli. Appetite 2017, 117, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.E.; Keast, R.S.J. Recent fat intake modulates fat taste sensitivity in lean and overweight subjects. Int. J. Obes. 2012, 36, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Noel, C.A.; Finlayson, G.; Dando, R. Prolonged Exposure to Monosodium Glutamate in Healthy Young Adults Decreases Perceived Umami Taste and Diminishes Appetite for Savory Foods. J. Nutr. 2018, 148, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Tepper, B.J.; White, E.A.; Koelliker, Y.; Lanzara, C.; d’Adamo, P.; Gasparini, P. Genetic variation in taste sensitivity to 6-n-propylthiouracil and its relationship to taste perception and food selection. Ann. N. Y. Acad. Sci. 2009, 1170, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Feeney, E.; O’Brien, S.; Scannell, A.; Markey, A.; Gibney, E.R. Genetic variation in taste perception: Does it have a role in healthy eating? Proc. Nutr. Soc. 2011, 70, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Passilly-Degrace, P.; Gaillard, D.; Merlin, J.-F.; Chevrot, M.; Besnard, P. The Lipid-Sensor Candidates CD36 and GPR120 Are Differentially Regulated by Dietary Lipids in Mouse Taste Buds: Impact on Spontaneous Fat Preference. PLoS ONE 2011, 6, e24014. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Zhou, L.H.; Ban, X.; Liu, D.X.; Jiang, W.; Liu, X.M. Decreased expression of CD36 in circumvallate taste buds of high-fat diet induced obese rats. Acta Histochem. 2011, 113, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Maliphol, A.B.; Garth, D.J.; Medler, K.F. Diet-Induced Obesity Reduces the Responsiveness of the Peripheral Taste Receptor Cells. PLoS ONE 2013, 8, e79403. [Google Scholar] [CrossRef] [PubMed]
- Drewnowski, A.; Brunzell, J.D.; Sande, K.; Iverius, P.H.; Greenwood, M.R. Sweet tooth reconsidered: Taste responsiveness in human obesity. Physiol. Behav. 1985, 35, 617–622. [Google Scholar] [CrossRef]
- Shi, D.; Nikodijević, O.; Jacobson, K.A.; Daly, J.W. Chronic Caffeine Alters the Density of Adenosine, Adrenergic, Cholinergic, GABA, and Serotonin Receptors and Calcium Channels in Mouse Brain. Cell. Mol. Neurobiol. 1993, 13, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, R.; Ninomiya, Y. Taste information derived from TAS1R-expressing taste cells in mice. Biochem. J. 2016, 473, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Oka, Y.; Butnaru, M.; von Buchholtz, L.; Ryba, N.J.; Zuker, C.S. High salt recruits aversive taste pathways. Nature 2013, 494, 472. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.J.; Cowart, B.J.; Beauchamp, G.K. The Development of Salty Taste Acceptance Is Related to Dietary Experience in Human Infants: A Prospective Study. Am. J. Clin. Nutr. 2012, 95, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Grassin-Delyle, S.; Abrial, C.; Fayad-Kobeissi, S.; Brollo, M.; Faisy, C.; Alvarez, J.C.; Devillier, P. The expression and relaxant effect of bitter taste receptors in human bronchi. Respir. Res. 2013, 14, 134. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, J.; Mueller, K.L.; Hoon, M.A.; Adler, E.; Feng, L.; Guo, W.; Ryba, N.J. T2Rs function as bitter taste receptors. Cell 2000, 100, 703–711. [Google Scholar] [CrossRef]
- Lipchock, S.V.; Mennella, J.A.; Spielman, A.I.; Reed, D.R. Human bitter perception correlates with bitter receptor messenger RNA expression in taste cells. Am. J. Clin. Nutr. 2013, 98, 1136–1143. [Google Scholar] [CrossRef] [PubMed]
- Lipchock, S.V.; Spielman, A.I.; Mennella, J.A.; Mansfield, C.J.; Hwang, L.D.; Douglas, J.E.; Reed, D.R. Caffeine bitterness is related to daily caffeine intake and bitter receptor mRNA abundance in human taste tissue. Perception 2017, 46, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Shigemura, N.; Shirosaki, S.; Sanematsu, K.; Yoshida, R.; Ninomiya, Y. Genetic and molecular basis of individual differences in human umami taste perception. PLoS ONE 2009, 4, e6717. [Google Scholar] [CrossRef] [PubMed]
- Beeler, J.A.; McCutcheon, J.E.; Cao, Z.F.; Murakami, M.; Alexander, E.; Roitman, M.F.; Zhuang, X. Taste uncoupled from nutrition fails to sustain the reinforcing properties of food. Eur. J. Neurosci. 2012, 36, 2533–2546. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Dvoryanchikov, G.; Pereira, E.; Chaudhari, N.; Roper, S.D. Breadth of tuning in taste afferent neurons varies with stimulus strength. Nat. Commun. 2015, 6, 8171. [Google Scholar] [CrossRef] [PubMed]
- Boughter, J.D.; Raghow, S.; Nelson, T.M.; Munger, S.D. Inbred mouse strains C57BL/6J and DBA/2J vary in sensitivity to a subset of bitter stimuli. BMC Genet. 2005, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Dando, R.; Roper, S.D. Acetylcholine is released from taste cells, enhancing taste signalling. J. Physiol. 2012, 590, 3009–3017. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahbandi, A.A.; Choo, E.; Dando, R. Receptor Regulation in Taste: Can Diet Influence How We Perceive Foods? J 2018, 1, 106-115. https://doi.org/10.3390/j1010011
Shahbandi AA, Choo E, Dando R. Receptor Regulation in Taste: Can Diet Influence How We Perceive Foods? J. 2018; 1(1):106-115. https://doi.org/10.3390/j1010011
Chicago/Turabian StyleShahbandi, Ashkan A., Ezen Choo, and Robin Dando. 2018. "Receptor Regulation in Taste: Can Diet Influence How We Perceive Foods?" J 1, no. 1: 106-115. https://doi.org/10.3390/j1010011
APA StyleShahbandi, A. A., Choo, E., & Dando, R. (2018). Receptor Regulation in Taste: Can Diet Influence How We Perceive Foods? J, 1(1), 106-115. https://doi.org/10.3390/j1010011