
Isolation and Structural Characterization of Alginates from the Kelp Species Laminaria ochroleuca and Saccorhiza polyschides from the Atlantic Coast of Morocco

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Biomass Preparation
2.2. Extraction and Purification of Alginates
2.3. FT-IR Spectroscopy Analysis
2.4. Proton Nuclear Magnetic Resonance Spectroscopy (1H NMR)
2.5. Intrinsic Viscosity [η] and Molecular Weight (Mv)
3. Results and Discussion
3.1. Alginate Yield
3.2. FT-IR Spectroscopy
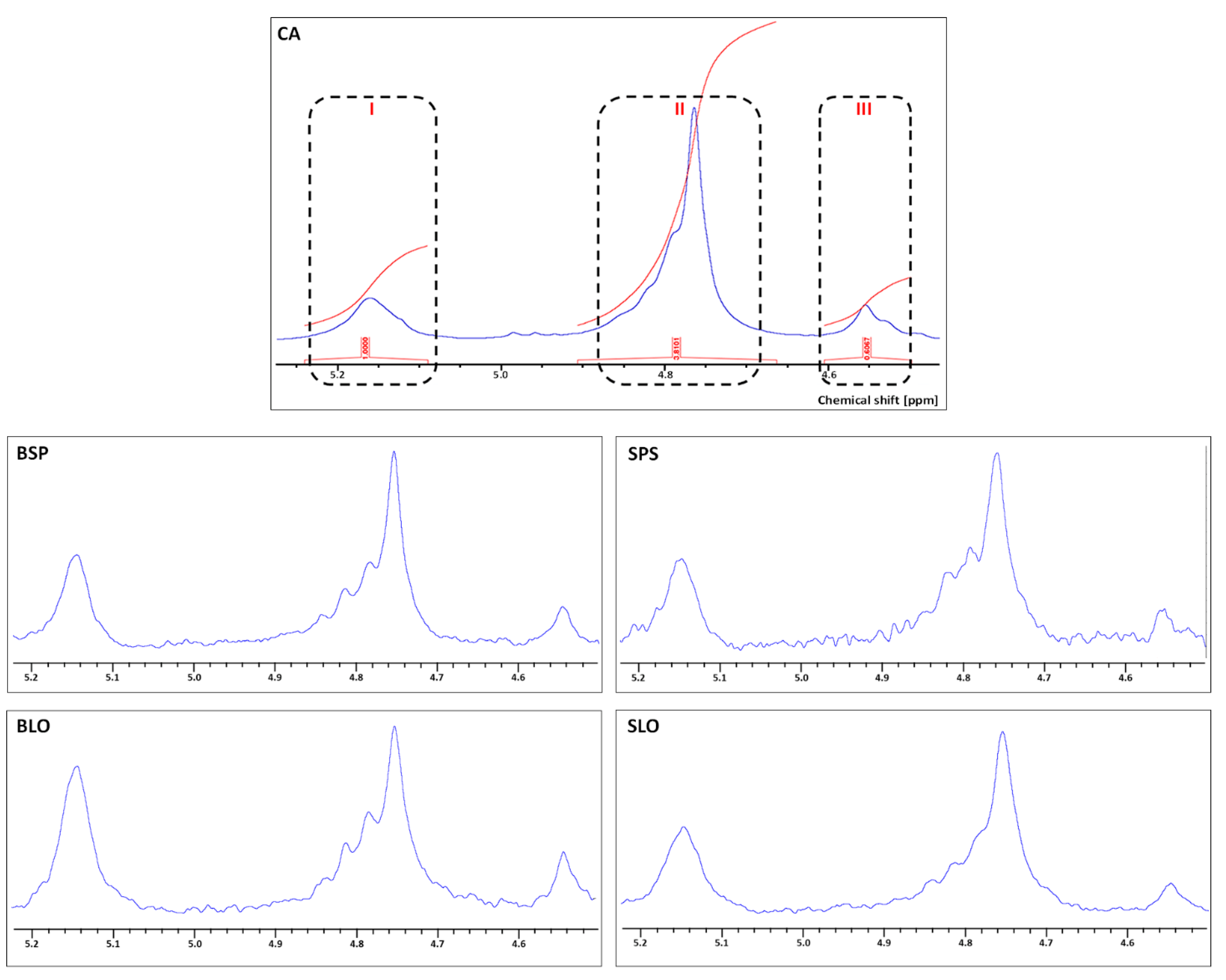
3.3. 1H NMR Spectroscopy Analysis
3.4. Viscosity and Molecular Weight
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Draget, K.I.; Moe, S.T.; Skjak-Bræk, G.; Smidsrød, O. Alginates. In Food Polysaccharides and Their Applications, 2nd ed.; Stephen, A.M., Phillips, G.O., Williams, P.A., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 289–334. [Google Scholar]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed]
- Morais, T.; Cotas, J.; Pacheco, D.; Pereira, L. Seaweeds compounds: An eco sustainable source of cosmetic ingredients. Cosmetics 2021, 8, 8. [Google Scholar] [CrossRef]
- Draget, K.I.; Skjåk-Braek, G.; Smidsrod, O.A. Alginate based new materials. Int. J. Biol. Macromol. 1997, 21, 47–55. [Google Scholar] [CrossRef]
- Peteiro, C. Alginate Production from Marine Macroalgae, with Emphasis on Kelp Farming. In Alginates and Their Biomedical Applications; Rehm, B., Moradali, M., Eds.; Springer: Singapore, 2018; Volume 11, pp. 27–66. [Google Scholar]
- Black, W.A.P. The seasonal variation in weight and chemical composition of the common British Laminariaceae. J. Mar. Biol. Assoc. UK 1950, 29, 45–72. [Google Scholar] [CrossRef]
- Jothisaraswathi, S.; Babu, B.; Rengasamy, R. Seasonal studies on alginate and its composition II: Turbinariaconoides (J.Ag.) Kütz. (Fucales, Phaeophyceae). J. Appl. Phycol. 2006, 18, 161–166. [Google Scholar] [CrossRef]
- Belattmania, Z.; Kaidi, S.; El Atouani, S.; Katif, C.; Bentiss, F.; Jama, C.; Reani, A.; Sabour, B.; Vasconcelos, V. Isolation and FTIR-ATR and 1H NMR Characterization of alginates from the main alginophyte species of the atlantic coast of Morocco. Molecules 2020, 25, 4335. [Google Scholar] [CrossRef]
- Venegas, M.; Matsuhiro, B.; Edding, M.E. Alginate composition of Lessonia trabeculate (Phaeophyta: Laminariales) growing in exposed and sheltered habitats. Bot. Mar. 1993, 36, 47–51. [Google Scholar] [CrossRef]
- Tønnesen, H.H.; Karlsen, J. Alginate in drug delivery systems. Drug. Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef]
- Chee, S.Y.; Wong, P.K.; Wong, C. Extraction and characterisation of alginate from brown seaweeds (Fucales, Phaeophyceae) collected from Port Dickson, Peninsular Malaysia. J. Appl. Phycol. 2010, 23, 191–196. [Google Scholar] [CrossRef]
- Calumpong, H.P.; Maypa, A.P.; Magbanua, M. Population and alginate yield and quality assessment of four Sargassum species in Negros Island, Central Philippines. Hydrobiologia 1999, 398, 211–215. [Google Scholar] [CrossRef]
- Torres, M.R.; Sousa, A.P.A.; Filho, E.A.T.S.; Melo, D.F.; Feitosa, J.P.A.; Paulab, R.C.M.; Limaa, M.G.S. Extraction and physicochemical characterization of Sargassum vulgare alginate from Brazil. Carbohydr. Res. 2007, 342, 2067–2074. [Google Scholar] [CrossRef] [PubMed]
- Westermeier, R.; Murúa, P.; Patiño, D.J.; Muñoz, L.; Ruiz, A.; Müller, D.G. Variations of chemical composition and energy content in natural and genetically defined cultivars of Macrocystis from Chile. J. Appl. Phycol. 2012, 24, 1191–1201. [Google Scholar] [CrossRef]
- Lorbeer, A.J.; Charoensiddhi, S.; Lahnstein, J.; Lars, C.; Franco, C.M.M.; Bulone, V.; Zhang, W. Sequential extraction and characterization of fucoidans and alginates from Ecklonia radiata, Macrocystis pyrifera, Durvillaea potatorum, and Seirococcusaxillaris. J. Appl. Phycol. 2017, 29, 1515–1526. [Google Scholar] [CrossRef]
- Honya, M.; Kinoshita, T.; Ishikawa, M.; Mori, H.; Nisizawa, K. Monthly determination of alginate, M/G ratio, mannitol, and minerals in cultivated Laminaria japonica. Nippon Suisan Gakk. 1993, 59, 295–299. [Google Scholar] [CrossRef]
- Manns, D.; Nielsen, M.M.; Bruhn, A.; Saake, B.; Meyer, A. Compositional variations of brown seaweeds Laminaria digitata and Saccharina latissima in Danish waters. J. Appl. Phycol. 2017, 29, 1493–1506. [Google Scholar] [CrossRef]
- Obluchinskaya, E.D.; Voskoboinikov, G.M.; Galynkin, V.A. Contents of alginic acid and fuccidan in Fucus algae of the Barents Sea. Appl. Biochem. Microbiol. 2002, 38, 186–188. [Google Scholar] [CrossRef]
- Kelly, B.J.; Brown, M.T. Variations in the alginate content and composition of Durvillaea antarctica and D. willana from southern New Zealand. J. Appl. Phycol. 2000, 12, 317–324. [Google Scholar] [CrossRef]
- Chandía, N.; Matsuhiro, B.; Vásquez, A.E. Alginic acids in Lessonia trabeculata: Characterization by formic acid hydrolysis and FT-IR spectroscopy. Carbohydr. Polym. 2001, 46, 81–87. [Google Scholar] [CrossRef]
- Mathlouthi, M.; Koenig, J.L. Vibrational Spectra of Carbohydrates. Adv. Carbohydr. Chem. Biochem. 1987, 44, 7–89. [Google Scholar]
- Papageorgiou, S.K.; Kouvelos, E.P.; Favvas, E.P.; Sapalidis, A.A.; Romanos, G.E.; Katsaros, F.K. Metal-carboxylate interactions in metal-alginate complexes studied with FTIR spectroscopy. Carbohydr. Res. 2010, 345, 469–473. [Google Scholar] [CrossRef]
- Rashedy, S.H.; Abd El Hafez, M.S.M.; Dar, M.A.; Cotas, J.; Pereira, L. Evaluation and characterization of alginate extracted from brown seaweed collected in the Red Sea. Appl. Sci. 2021, 11, 6290. [Google Scholar] [CrossRef]
- Davis, T.A.; Llanes, F.; Volesky, B.; Diaz-Pulido, G.; McCook, L.; Mucci, A. 1H-NMR study of Na alginates extracted from Sargassum spp. in relation to metal biosorption. Appl. Biochem. Biotechnol. 2003, 110, 75–90. [Google Scholar] [CrossRef]
- Leal, D.; Matsuhiro, B.; Rossi, M.; Caruso, F. FT-IR spectra of alginic acid block fractions in three species of brown seaweeds. Carbohydr. Res. 2008, 343, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Panikkar, R.; Brasch, D.J. Composition and block structure of alginates from New Zealand brown seaweeds. Carbohydr. Res. 1996, 293, 119–132. [Google Scholar] [CrossRef]
- Grasdalen, H.; Larsen, B.; Smidsrod, O.A. A p.m.r. study of the composition and sequence of uronate residues in alginates. Carbohydr. Res. 1979, 68, 23–31. [Google Scholar] [CrossRef]
- Stokke, B.T.; Draget, K.I.; Smidsrød, O.; Yuguchi, Y.; Urakawa, H.; Kajiwara, K. Small-angle X-ray scattering and rheological characterization of alginate gels. 1. Ca−Alginate Gels. Macromolecules 2000, 33, 1853–1863. [Google Scholar] [CrossRef]
- Smidsrød, O.; Draget, K.I. Chemistry and physical properties of alginates. Chemistry 1996, 14, 6–13. [Google Scholar]
- Hartmann, M.; Dentini, M.; Draget, K.I.; Skjåk-Bræk, G. Enzymatic modification of alginates with the mannuronan C-5epimerase AlgE4 enhances their solubility at low pH. Carbohydr. Polym. 2006, 63, 257–262. [Google Scholar] [CrossRef]
- Aarstad, O.; Heggset, E.B.; Pedersen, I.S.; Bjørnøy, S.H.; Syverud, K.; Strand, B.L. Mechanical properties of composite hydrogels of alginate and cellulose nanofibrils. Polymers 2017, 9, 378. [Google Scholar] [CrossRef]
- Murillo-Alvarez, J.I.; Hernández-Carmona, G. Monomer composition and sequence of sodium alginate extracted at pilot plant scale from three commercially important seaweeds from Mexico. J. Appl. Phycol. 2007, 19, 545–548. [Google Scholar] [CrossRef]
- Zheng, N.; Zhang, Y.; Fan, X.; Han, L. Effects of composition and structure of alginates on adsorption of divalent metals. Chin. J. Oceanol. Limnol. 1994, 12, 78–83. [Google Scholar]
- Indergaard, M.; Skjåk-Bræk, G.; Jensen, A. Studies on the influence of nutrients on the composition and structure of alginate in Laminaria saccharina (L.) Lamour. (Laminariales, Phaeophyceae). Bot. Mar. 1990, 33, 277–288. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- McKee, J.W.A.; Kavalieris, L.; Brasch, D.J.; Brown, M.T.; Melton, L.D. Alginate content and composition of Macrocystis pyrifera from New Zealand. J. Appl. Phycol. 1992, 4, 357–369. [Google Scholar] [CrossRef]
- Craigie, J.S.; Morris, E.R.; Rees, D.A.; Thom, D. Alginate block structure in phaeophyceae from Nova Scotia: Variation with species, environment and tissue-type. Carbohydr. Polym. 1984, 4, 237–252. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Haug, A.; Larsen, B.; Smidsrod, O.A. Uronic acid sequence in alginate from different sources. Carbohydr. Res. 1974, 32, 217–225. [Google Scholar] [CrossRef]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2010, 23, 321–335. [Google Scholar] [CrossRef]
- Kaidi, S.; Belattmania, Z.; Bentiss, F.; Jama, C.; Reani, A.; Sabour, B. Synthesis and characterization of silver nanoparticles using alginate from the brown seaweed Laminaria ochroleuca: Structural features and antibacterial activity. Biointerface Res. Appl. Chem. 2022, 12, 6046–6057. [Google Scholar]
- Mancini, F.; Montanari, L.; Peressini, D.; Fantozzi, P. Influence of alginate concentration and molecular weight on functional properties of mayonnaise. LWT Food Sci. Technol. 2002, 35, 517–525. [Google Scholar] [CrossRef]
- Draget, K.I.; Stokke, B.T.; Yuguchi, Y.; Urakawa, H.; Kajiwara, K. Small-angle X-ray scattering and rheological characterization of alginate gels. Alginic acid gels. Biomacromolecules 2003, 4, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Fourest, E.; Volesky, B. Alginate Properties and Heavy Metal Biosorption by Marine Algae. Appl. Biochem. Biotechnol. 1997, 67, 215–226. [Google Scholar] [CrossRef]
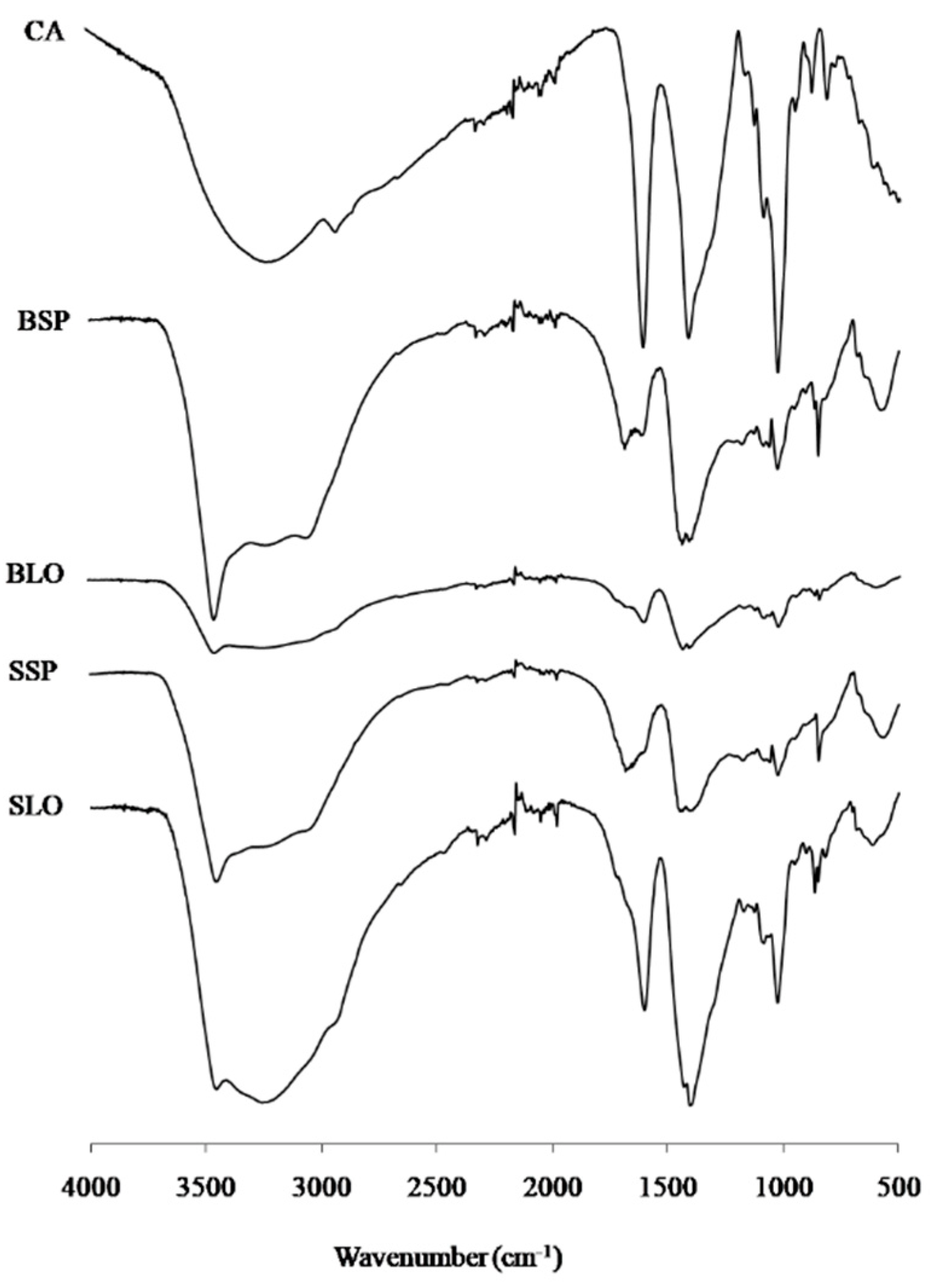

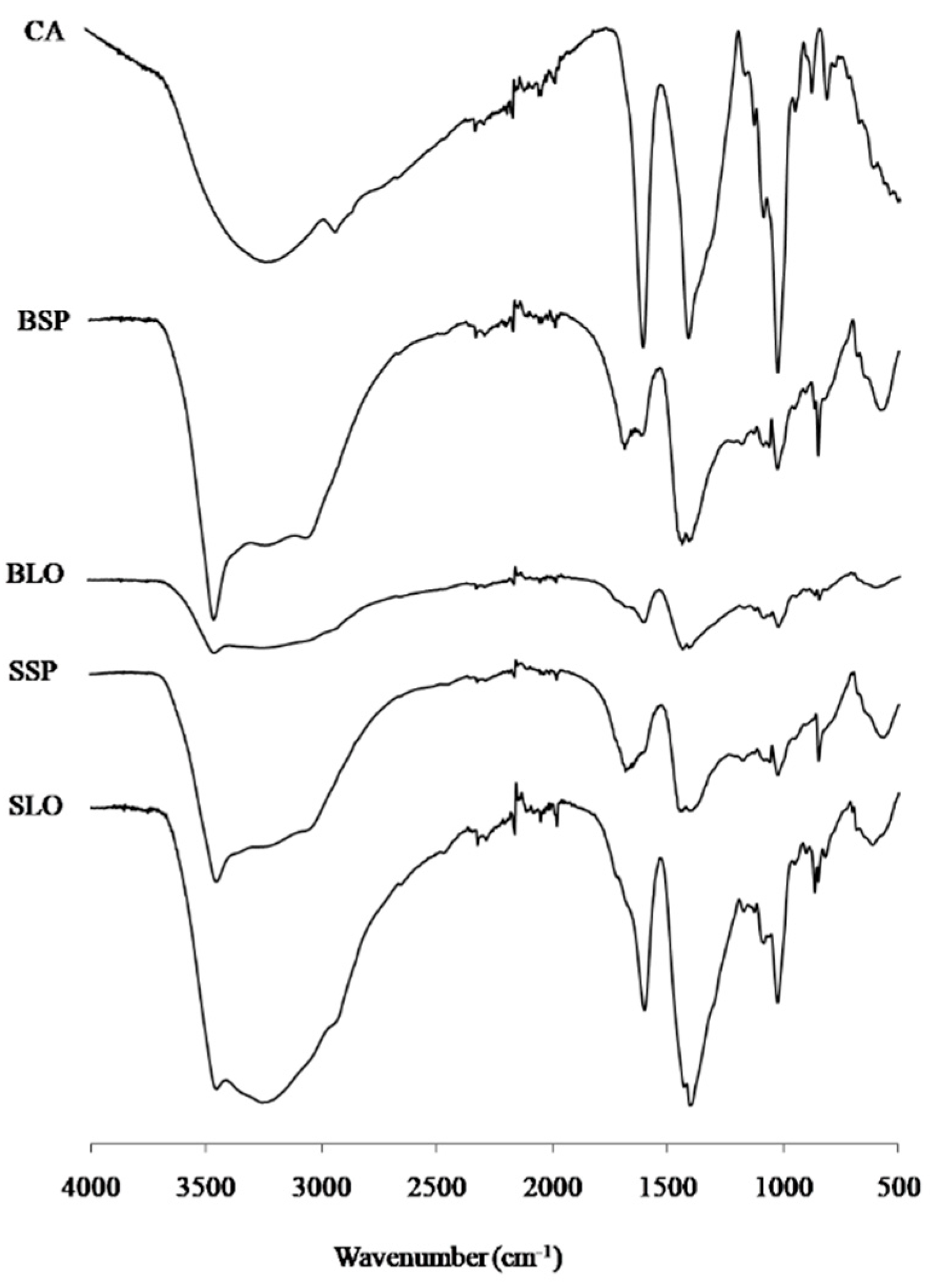{kind=link}
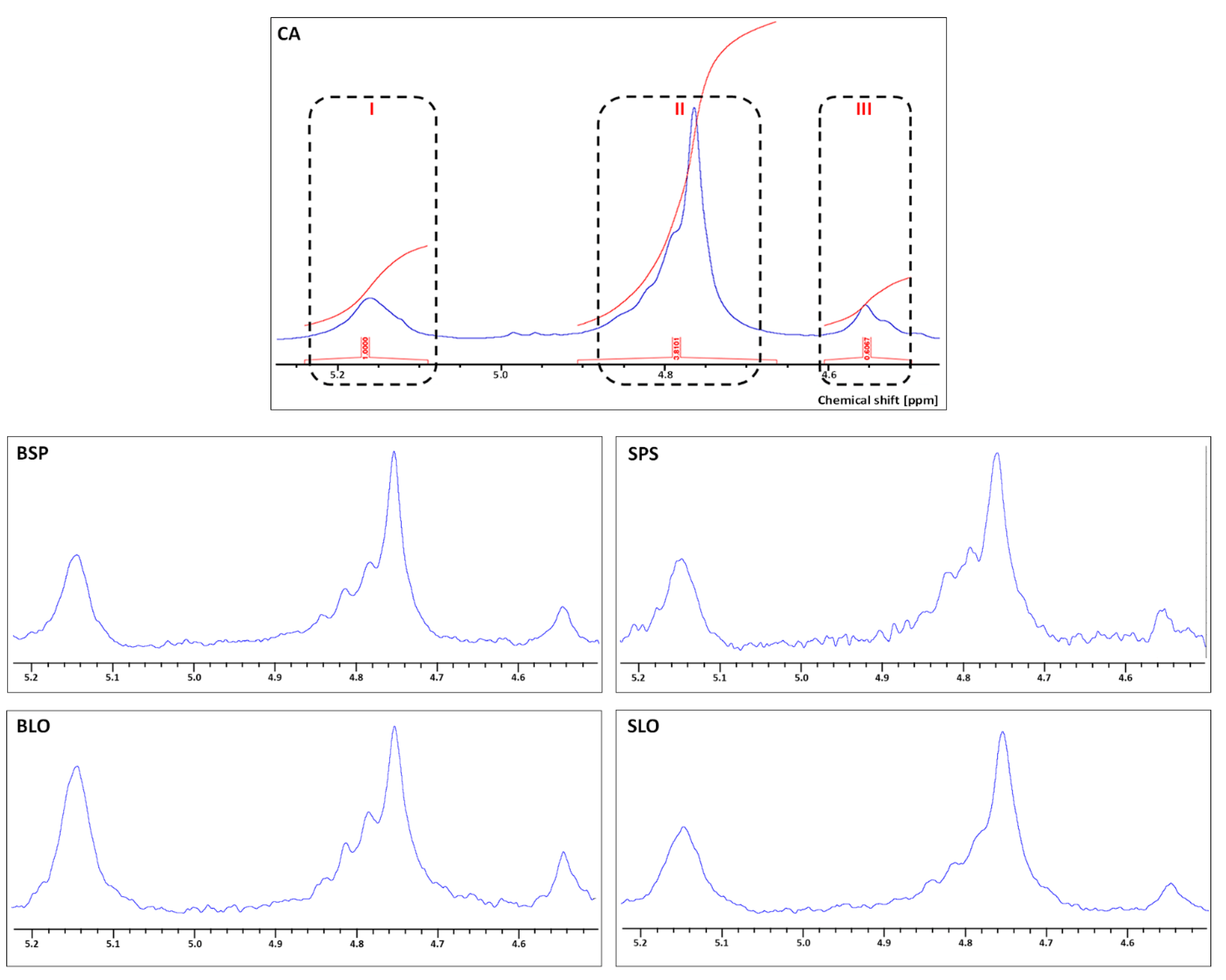{kind=link}
| Parameters | L. ochroleuca | S. polyschides | |
|---|---|---|---|
| Length (cm) | (blade) | 61.8 ± 2.8 | 76.18 ± 10 |
| (stipe) | 6.92 ± 0.5 | 3.16 ± 1.8 | |
| Fresh Weight (g) | (blade) | 227.6 ± 23.3 | 289.66 ± 85 |
| (stipe) | 6.75 ± 1 | 5.22 ± 7 | |
| Moisture (%) | (blade) | 5.7 ± 4 | 7.9 ± 0.6 |
| (stipe) | 4.37 ± 1.4 | 5 ± 0.9 | |
| Ash content (%) | (blade) | 0.50 ± 1.2 | 0.98 ± 2.4 |
| Density (2 m2) | 3.0 ± 1.0 | 3.0 ± 3.0 | |
| Fertility (%) | 64.1 ± 5 | 0 | |
| Kelp Species | Alginates Yield | References |
|---|---|---|
| Macrocystis pyrifera | 18–45% | [14] |
| Laminaria digitata | 16–36% | [17] |
| Laminaria hyperborea | 14–21% | [6] |
| Saccharina japonica | 17–25% | [16] |
| Saccharina latissima | 16–34% | [17] |
| Lessonia trabeculata | 13–29% | [20] |
| Durvillaea potatorum | 55% | [15] |
| Ascophyllum nodosum | 12–16% | [18] |
| Laminaria ochroleuca (blade) | 28.96 ± 0.20 | This study |
| Laminaria ochroleuca (stipe) | 49.80 ± 2.40 | |
| Saccorhiza polyschides (blade) | 20.19 ± 2.00 | |
| Saccorhiza polyschides (stipe) | 26.28 ± 0.08 |
| Kelp Species | Composition | Sequence | References | |||||
|---|---|---|---|---|---|---|---|---|
| FM | FG | M/G | FMM | FGG | FMG, GM | η | ||
| Macrocystis pyrifera | 0.62 | 0.38 | 1.63 | 0.42 | 0.18 | 0.40 | 0.85 | [32] |
| Laminaria hyperborean (blade) | 0.45 | 0.55 | 0.81 | 0.28 | 0.38 | 0.34 | 0.69 | [29] |
| Laminaria hyperborean (stipe) | 0.32 | 0.68 | 0.47 | 0.2 | 0.56 | 0.24 | 0.55 | [29] |
| Laminaria digitata | 0.61 | 0.39 | 1.56 | 0.41 | 0.15 | 0.30 | 0.63 | [27] |
| Laminaria ochroleuca | 0.72 | 0.28 | 2.52 | 0.5 | 0.06 | 0.44 | 1.09 | [8] |
| Laminaria brasiliensis (blade) | 0.47 | 0.53 | 0.89 | 0.27 | 0.33 | 0.40 | 0.80 | [1] |
| Laminaria brasiliensis (stipe) | 0.46 | 0.54 | 0.85 | 0.23 | 0.31 | 0.46 | 0.93 | [1] |
| Saccorhiza polyschides (blade) | 0.42 | 0.58 | 0.72 | 0.3 | 0.46 | 0.24 | 0.49 | [1] |
| Saccorhiza polyschides (stipe) | 0.51 | 0.49 | 1.04 | 0.36 | 0.34 | 0.30 | 0.60 | [1] |
| Saccharina japonica | 0.65 | 0.35 | 1.85 | 0.48 | 0.18 | 0.34 | 0.75 | [33] |
| Saccharina latissima (old blade) | 0.5 | 0.5 | 1.01 | 0.36 | 0.35 | 0.30 | 0.60 | [34] |
| Saccharina latissima (old stipe) | 0.56 | 0.44 | 1.27 | 0.39 | 0.27 | 0.34 | 0.69 | [34] |
| Saccharina longicruris | 0.41 | 0.59 | 0.69 | 0.07 | 0.25 | 0.68 | 1.41 | [35] |
| Lessonia nigrescens | 0.59 | 0.41 | 1.43 | 0.4 | 0.22 | 0.38 | 0.79 | [30] |
| Lessonia trabeculata (blade) | 0.38 | 0.62 | 0.61 | 0.21 | 0.47 | 0.30 | 0.64 | [9] |
| Ecklonia arborea | 0.52 | 0.48 | 1.08 | 0.37 | 0.33 | 0.30 | 0.60 | [32] |
| Ecklonia maxima | 0.55 | 0.45 | 1.22 | 0.32 | 0.22 | 0.64 | 1.29 | [1] |
| Durvillaea potatorum | 0.68 | 0.32 | 2.125 | 0.56 | 0.2 | 0.24 | 0.55 | [31] |
| Durvillaea willana | 0.72 | 0.28 | 2.57 | 0.51 | 0.07 | 0.42 | 1.04 | [26] |
| Durvillaea antarctica | 0.68 | 0.32 | 2.15 | 0.51 | 0.16 | 0.34 | 0.78 | [30] |
| Marginariella boryana | 0.44 | 0.56 | 0.79 | 0.28 | 0.4 | 0.32 | 0.65 | [26] |
| Ascophyllum nodosum | 0.61 | 0.39 | 1.56 | 0.46 | 0.23 | 0.32 | 0.67 | [28] |
| Saccorhiza polyschides | 0.63 | 0.37 | 1.73 | 0.38 | 0.11 | 0.50 | 1.09 | [8] |
| Laminaria ochroleuca (blade) | 0.47 | 0.53 | 0.89 | 0.09 | 0.16 | 0.75 | 1.51 | This study |
| Laminaria ochroleuca (stipe) | 0.58 | 0.42 | 1.38 | 0.24 | 0.08 | 0.68 | 1.39 | |
| Saccorhiza polyschides (blade) | 0.62 | 0.38 | 1.62 | 0.37 | 0.14 | 0.49 | 1.04 | |
| Saccorhiza polyschides (stipe) | 0.68 | 0.32 | 2.14 | 0.44 | 0.08 | 0.48 | 1.10 | |
| Commercial alginate (Sigma-Aldrich Na-Alginate) | 0.77 | 0.23 | 3.42 | 0.68 | 0.14 | 0.18 | 0.51 | |
| Kelp Species | [µ] (dL/g) | MW × 10−5 (g/mol) | References |
|---|---|---|---|
| Macrocystis pyrifera | 9.43 | 0.76 | [42] |
| Laminaria hyperborea (blade) | 5 | 1.55 | [43] |
| Laminaria hyperborea (stipe) | 5.2 | 1.6 | [43] |
| Ascophyllum nodosum | 2.8 | 1.32 | [44] |
| Laminaria japonica | 15.4 | 7.44 | [44] |
| Laminaria ochroleuca (blade) | 2.84 | 1.34 | This study |
| Laminaria ochroleuca (stipe) | 1.43 | 0.66 | |
| Saccorhiza polyschides (blade) | 1.56 | 0.73 | |
| Saccorhiza polyschides (stipe) | 1.14 | 0.53 | |
| Commercial alginate (Sigma-Aldrich Na-Alginate) | 1.83 | 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaidi, S.; Bentiss, F.; Jama, C.; Khaya, K.; Belattmania, Z.; Reani, A.; Sabour, B. Isolation and Structural Characterization of Alginates from the Kelp Species Laminaria ochroleuca and Saccorhiza polyschides from the Atlantic Coast of Morocco. Colloids Interfaces 2022, 6, 51. https://doi.org/10.3390/colloids6040051
Kaidi S, Bentiss F, Jama C, Khaya K, Belattmania Z, Reani A, Sabour B. Isolation and Structural Characterization of Alginates from the Kelp Species Laminaria ochroleuca and Saccorhiza polyschides from the Atlantic Coast of Morocco. Colloids and Interfaces. 2022; 6(4):51. https://doi.org/10.3390/colloids6040051
Chicago/Turabian StyleKaidi, Soukaina, Fouad Bentiss, Charafeddine Jama, Khaoula Khaya, Zahira Belattmania, Abdeltif Reani, and Brahim Sabour. 2022. "Isolation and Structural Characterization of Alginates from the Kelp Species Laminaria ochroleuca and Saccorhiza polyschides from the Atlantic Coast of Morocco" Colloids and Interfaces 6, no. 4: 51. https://doi.org/10.3390/colloids6040051
APA StyleKaidi, S., Bentiss, F., Jama, C., Khaya, K., Belattmania, Z., Reani, A., & Sabour, B. (2022). Isolation and Structural Characterization of Alginates from the Kelp Species Laminaria ochroleuca and Saccorhiza polyschides from the Atlantic Coast of Morocco. Colloids and Interfaces, 6(4), 51. https://doi.org/10.3390/colloids6040051