
Species-Specific Responses of Bird Song Output in the Presence of Drones


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Study
2.2. Bioacoustic Analysis and Sound Localization
2.3. Statistical Analysis
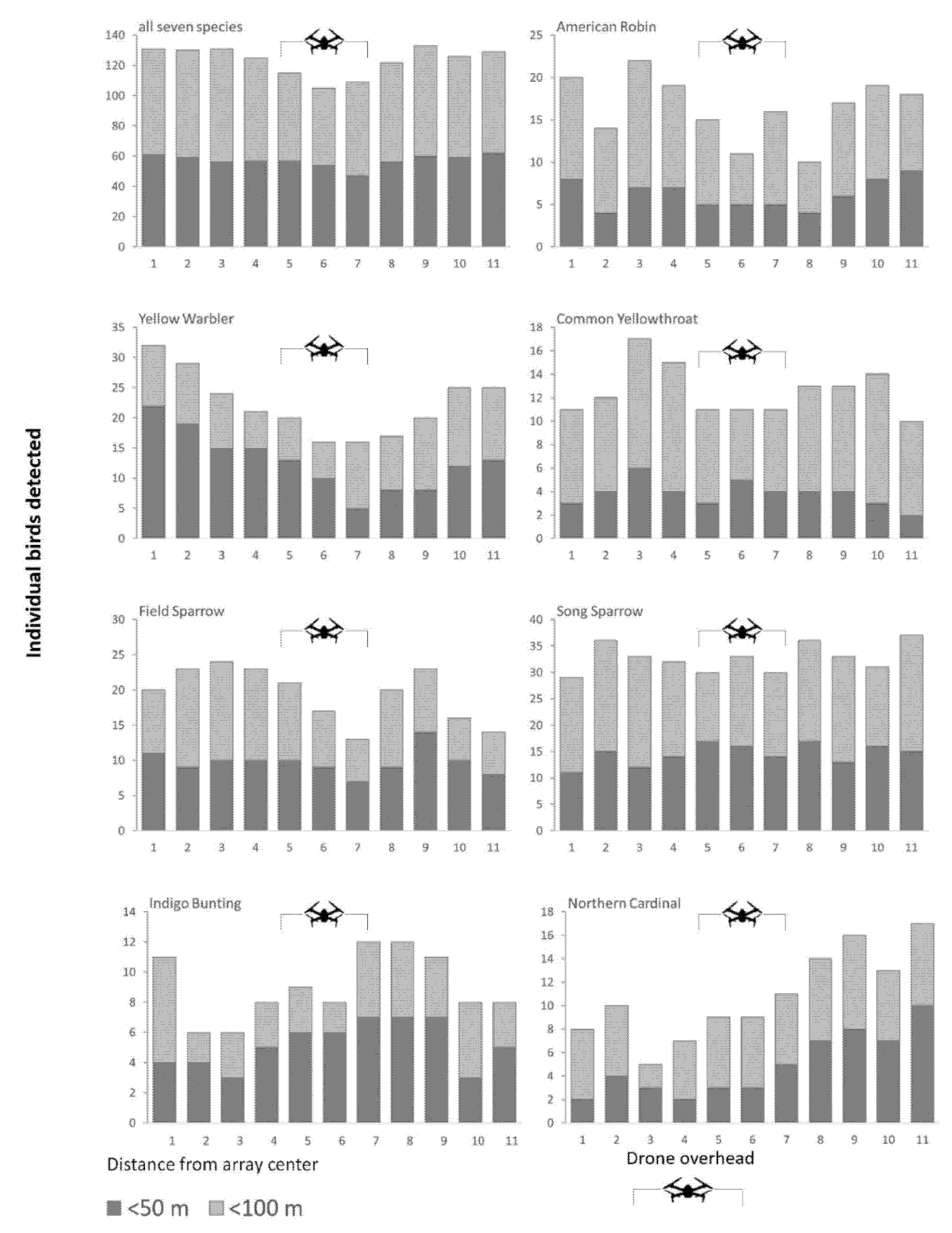
3. Results
3.1. Probability of Singing Bird Detection
3.2. Singing Rate
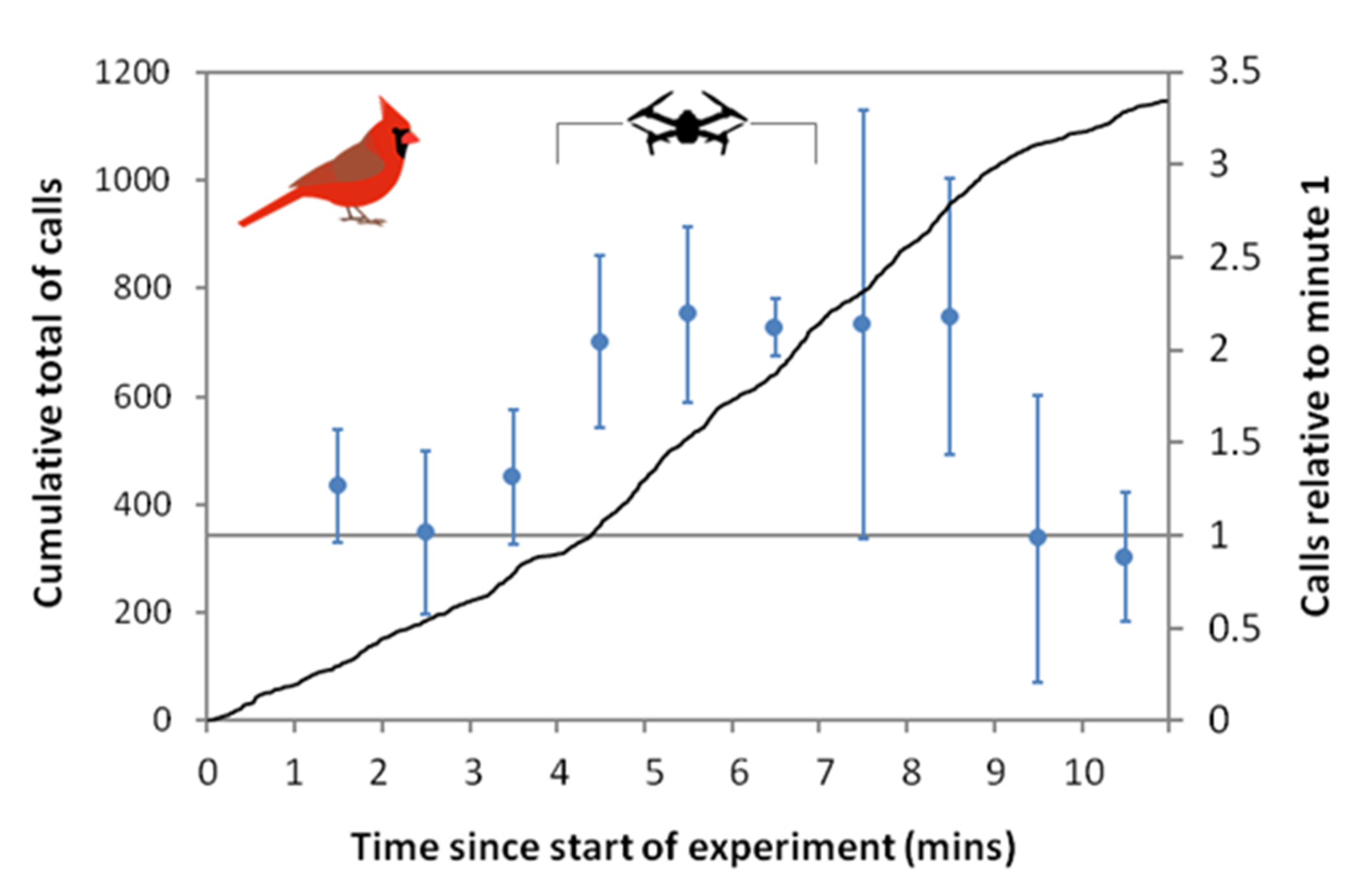
3.3. Cardinal Calls
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Ethics Statement
Conflicts of Interest
References
- Kelaher, B.P.; Colefax, A.P.; Tagliafico, A.; Bishop, M.J.; Giles, A.; Butcher, P.A. Assessing variation in assemblages of large marine fauna off ocean beaches using drones. Mar. Freshw. Res. 2020, 71, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, C.; Lejeune, P.; Lisein, J.; Sawadogo, P.; Bouché, P. Unmanned aerial survey of elephants. PLoS ONE 2013, 8, e54700. [Google Scholar] [CrossRef] [Green Version]
- Witczuk, J.; Pagacz, S.; Zmarz, A.; Cypel, M. Exploring the feasibility of unmanned aerial vehicles and thermal imaging for ungulate surveys in forests—Preliminary results. Int. J. Remote Sens. 2018, 39, 5504–5521. [Google Scholar] [CrossRef]
- Christiansen, F.; Dujon, A.M.; Sprogis, K.R.; Arnould, J.P.Y.; Bejder, L. Noninvasive unmanned aerial vehicle provides estimates of the energetic cost of reproduction in humpback whales. Ecosphere 2016, 7, e01468. [Google Scholar] [CrossRef]
- Barnas, A.F.; Darby, B.J.; VandeBerg, G.S.; Rockwell, R.F.; Ellis-Felege, S.N. A comparison of drone imagery and ground-based methods for estimating the extent of habitat destruction by lesser snow geese (Anser caerulescens caerulescens) in La Pérouse Bay. PLoS ONE 2019, 14, e0217049. [Google Scholar] [CrossRef] [Green Version]
- Ratcliffe, N.; Guihen, D.; Robst, J.; Crofts, S.; Stanworth, A.; Enderlein, P. A protocol for the aerial survey of penguin colonies using UAVs. J. Unmanned Veh. Syst. 2015, 3, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Afán, I.; Máñez, M.; Díaz-Delgado, R. Drone Monitoring of Breeding Waterbird Populations: The Case of the Glossy Ibis. Drones 2018, 2, 42. [Google Scholar] [CrossRef] [Green Version]
- Weissensteiner, M.H.; Poelstra, J.W.; Wolf, J.B.W. Low-budget ready-to-fly unmanned aerial vehicles: An effective tool for evaluating the nesting status of canopy-breeding bird species. J. Avian Biol. 2015, 46, 425–430. [Google Scholar] [CrossRef]
- Borrelle, S.B.; Fletcher, A.T. Will drones reduce investigator disturbance to surface-nesting birds? Mar. Ornithol. 2017, 45, 89–94. [Google Scholar]
- Inger, R.; Gregory, R.; Duffy, J.P.; Stott, I.; Voříšek, P.; Gaston, K.J. Common European birds are declining rapidly while less abundant species’ numbers are rising. Ecol. Lett. 2014, 18, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef]
- Musgrove, A.; Aebischer, N.; Eaton, M.; Hearn, R.; Newson, S.; Noble, D.; Parsons, M.; Risely, K.; Stroud, D. Population estimates of birds in Great Britain and the UK. Br. Birds 2013, 106, 231–232. [Google Scholar]
- Wilson, A.M.; Barr, J.; Zagorski, M. The feasibility of counting songbirds using unmanned aerial vehicles. Auk 2017, 134, 350–362. [Google Scholar] [CrossRef]
- Kloepper, L.N.; Kinniry, M. Recording animal vocalizations from a UAV: Bat echolocation during roost re-entry. Sci. Rep. 2018, 8, 7779. [Google Scholar] [CrossRef] [PubMed]
- Scholten, C.N.; Kamphuis, A.J.; Vredevoogd, K.J.; Lee-Strydhorst, K.G.; Atma, J.L.; Shea, C.B.; Lamberg, O.N.; Proppe, D.S. Real-time thermal imagery from an unmanned aerial vehicle can locate ground nests of a grassland songbird at rates similar to traditional methods. Biol. Conserv. 2019, 233, 241–246. [Google Scholar] [CrossRef]
- Mulero-Pázmány, M.; Jenni-Eiermann, S.; Strebel, N.; Sattler, T.; Negro, J.J.; Tablado, Z. Unmanned aircraft systems as a new source of disturbance for wildlife: A systematic review. PLoS ONE 2017, 12, e0178448. [Google Scholar] [CrossRef] [Green Version]
- Bibby, C.J. Ecoscope Applied Ecologists, British Trust for Ornithology, Royal Society for the Protection of Birds, and BirdLife International. In Bird Census Techniques; Academic Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Wilson, D.R.; Battiston, M.; Brzustowski, J.; Mennill, D.J. Sound Finder: A new software approach for localizing animals recorded with a microphone array. Bioacoustics 2014, 23, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Hartshorne, C. The Monotony-Threshold in Singing Birds. Auk 1956, 73, 176–192. [Google Scholar] [CrossRef]
- Wildlife Acoustics. Kaleidoscope Pro Analysis Software; Wildlife Acoustics, Inc.: Boston, MA, USA, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- McClure, C.J.W.; Ware, H.E.; Carlisle, J.; Kaltenecker, G.; Barber, J.R. An experimental investigation into the effects of traffic noise on distributions of birds: Avoiding the phantom road. Proc. R. Soc. B Boil. Sci. 2013, 280, 20132290. [Google Scholar] [CrossRef] [Green Version]
- Francis, C.D.; Ortega, C.P.; Cruz, A. Noise Pollution Changes Avian Communities and Species Interactions. Curr. Biol. 2009, 19, 1415–1419. [Google Scholar] [CrossRef] [Green Version]
- Roca, I.T.; DesRochers, L.; Giacomazzo, M.; Bertolo, A.; Bolduc, P.; Deschesnes, R.; Martin, C.A.; Rainville, V.; Rheault, G.; Proulx, R. Shifting song frequencies in response to anthropogenic noise: A meta-analysis on birds and anurans. Behav. Ecol. 2016, 27, 1269–1274. [Google Scholar] [CrossRef]
- Pieplow, N. Peterson Field Guide to Bird Sounds of Eastern North Americae; Houghton Mifflin Harcour: New York, NY, USA, 2017. [Google Scholar]
- Miljkovic, D. Methods for attenuation of unmanned aerial vehicle noise. In Proceedings of the 2018 41st International Convention on Information and Communication Technology, Electronics and Microelectronics, MIPRO 2018-Proceedings, Opatija, Croatia, 21–25 May 2018; pp. 914–919. [Google Scholar] [CrossRef]
- Seger-Fullam, K.D.; Rodewald, A.D.; Soha, J.A. Urban noise predicts song frequency in Northern Cardinals and American Robins. Bioacoustics 2011, 20, 267–276. [Google Scholar] [CrossRef]
- Dowling, J.L.; Luther, D.A.; Marra, P.P. Comparative effects of urban development and anthropogenic noise on bird songs. Behav. Ecol. 2012, 23, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Valle, R.G.; Scarton, F. Effectiveness, efficiency, and safety of censusing Eurasian oystercatchers Haematopus ostralegus by unmanned aircraft. Mar. Ornithol. 2019, 47, 81–87. [Google Scholar]
- Shannon, G.; McKenna, M.F.; Angeloni, L.M.; Crooks, K.R.; Fristrup, K.M.; Brown, E.J.; Warner, K.A.; Nelson, M.D.; White, C.; Briggs, J.; et al. A synthesis of two decades of research documenting the effects of noise on wildlife. Biol. Rev. 2016, 91, 982–1005. [Google Scholar] [CrossRef] [PubMed]
- Kight, C.R.; Saha, M.S.; Swaddle, J.P. Anthropogenic noise is associated with reductions in the productivity of breeding Eastern Bluebirds (Sialia sialis). Ecol. Appl. 2012, 22, 1989–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, D.B.; Monfils, M.J. Occupancy modeling of bird point counts: Implications of mobile animals. J. Wildl. Manag. 2015, 79, 1361–1368. [Google Scholar] [CrossRef]
- Royle, J.A.; Nichols, J.D. Estimating abundance from repeated presence–absence data or point counts. Ecology 2003, 84, 777–790. [Google Scholar] [CrossRef]
- Royle, J.A. N-mixture models for estimating population size from spatially replicated counts. Biometrics 2004, 60, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Shonfield, J.; Bayne, E.M. Autonomous recording units in avian ecological research: Current use and future applications. Avian Conserv. Ecol. 2017, 12, 14. [Google Scholar] [CrossRef]
- Campbell, M.; Francis, C.M. Using Stereo-Microphones to Evaluate Observer Variation in North American Breeding Bird Survey Point Counts. Auk 2011, 128, 303–312. [Google Scholar] [CrossRef]
- Lowther, P.E.; Celada, C.; Klein, N.K.; Rimmer, C.C.; Spector, D.A. Yellow Warbler (Setophaga petechia), version 2.0. In The Birds of North America; Poole, A.F., Gill, F.B., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 1999. [Google Scholar] [CrossRef]
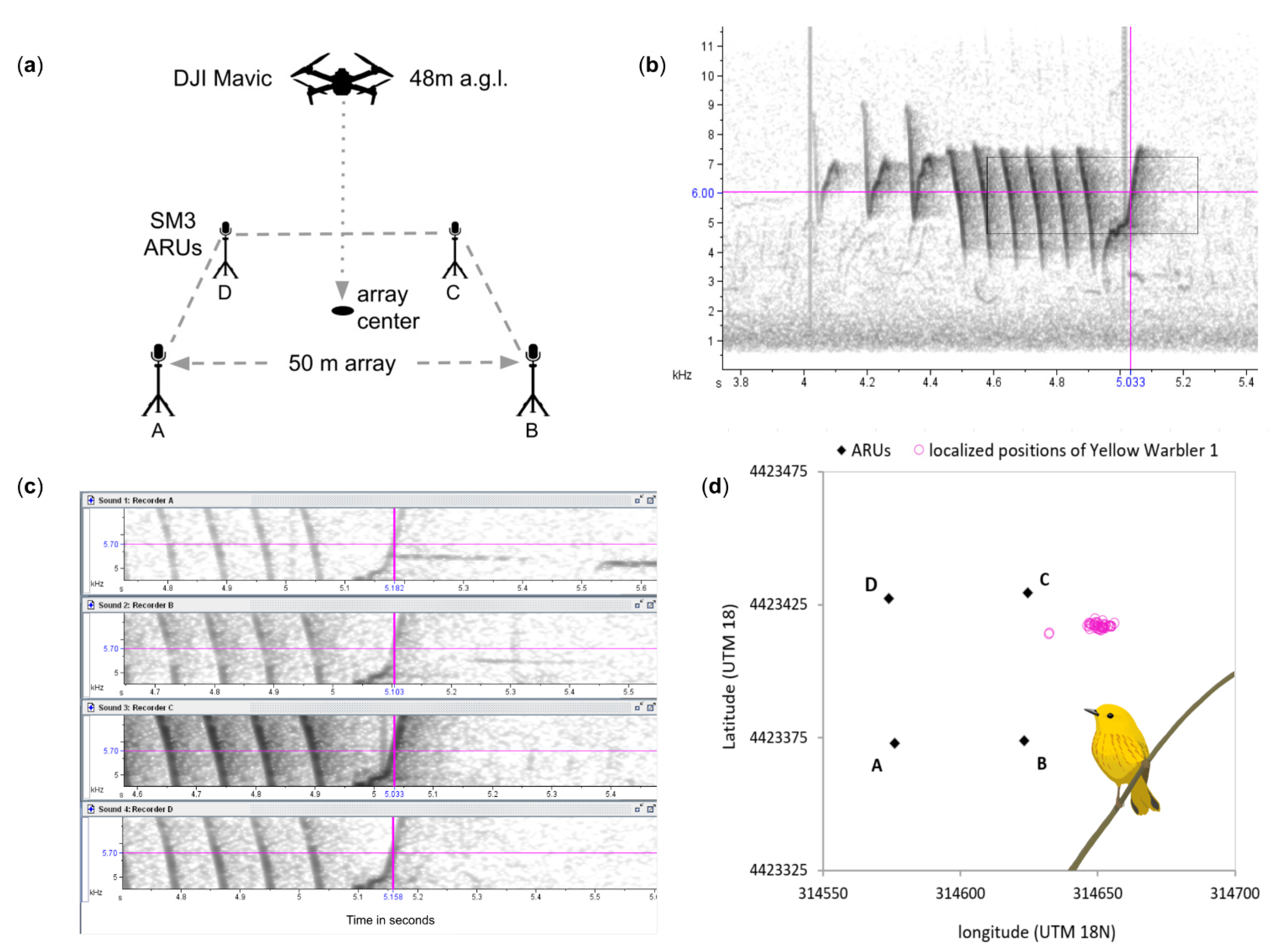




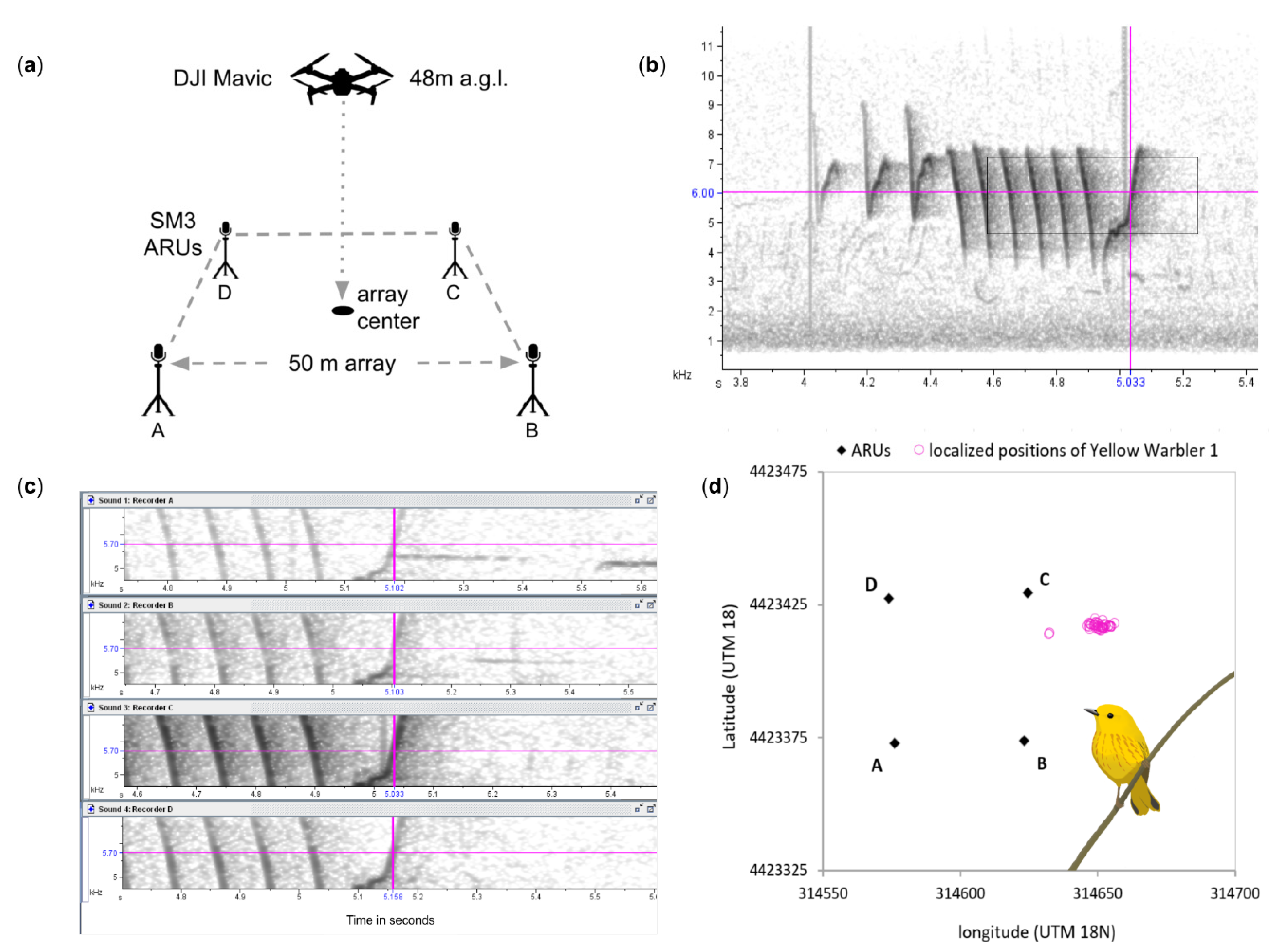{kind=link}
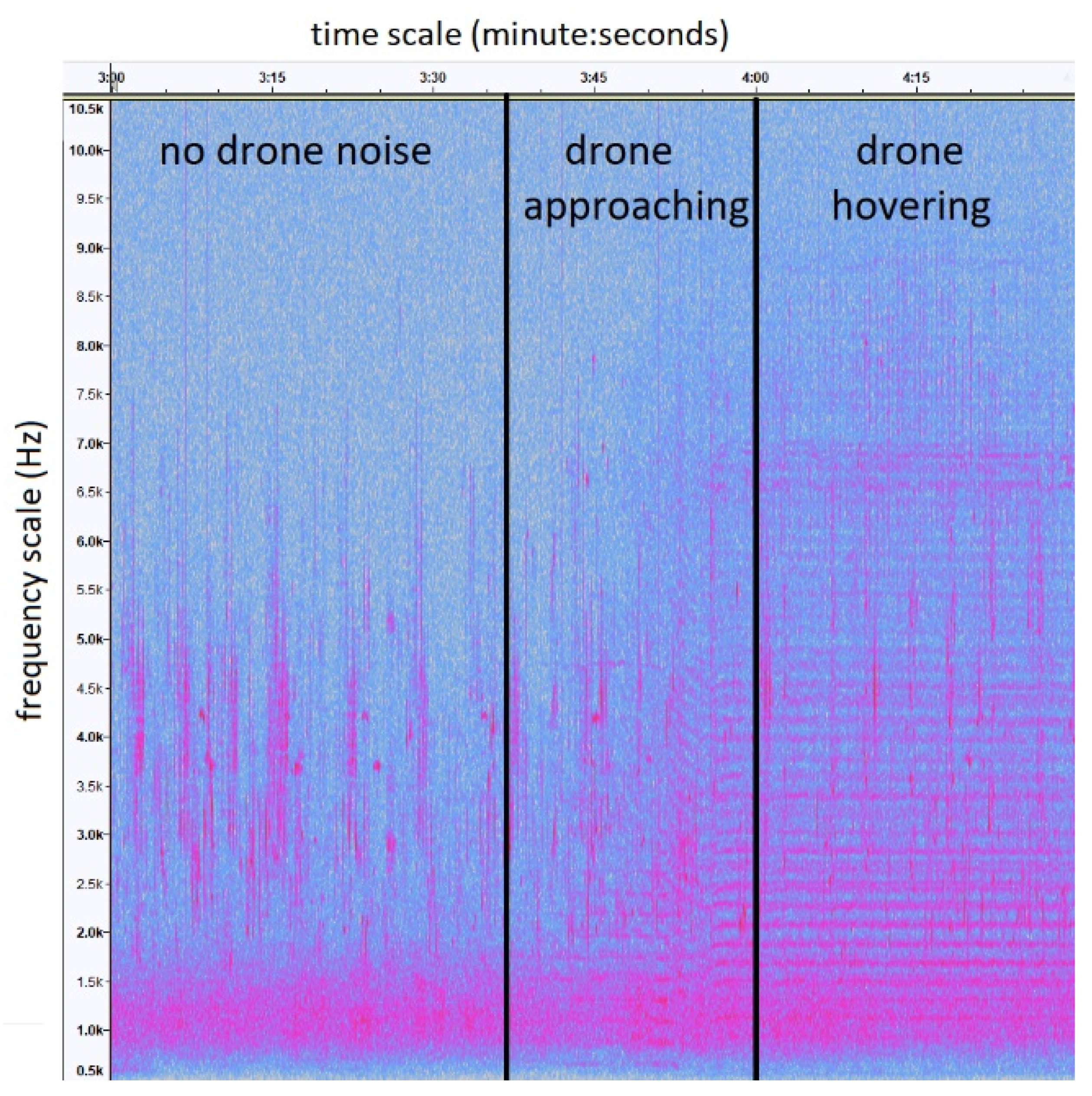
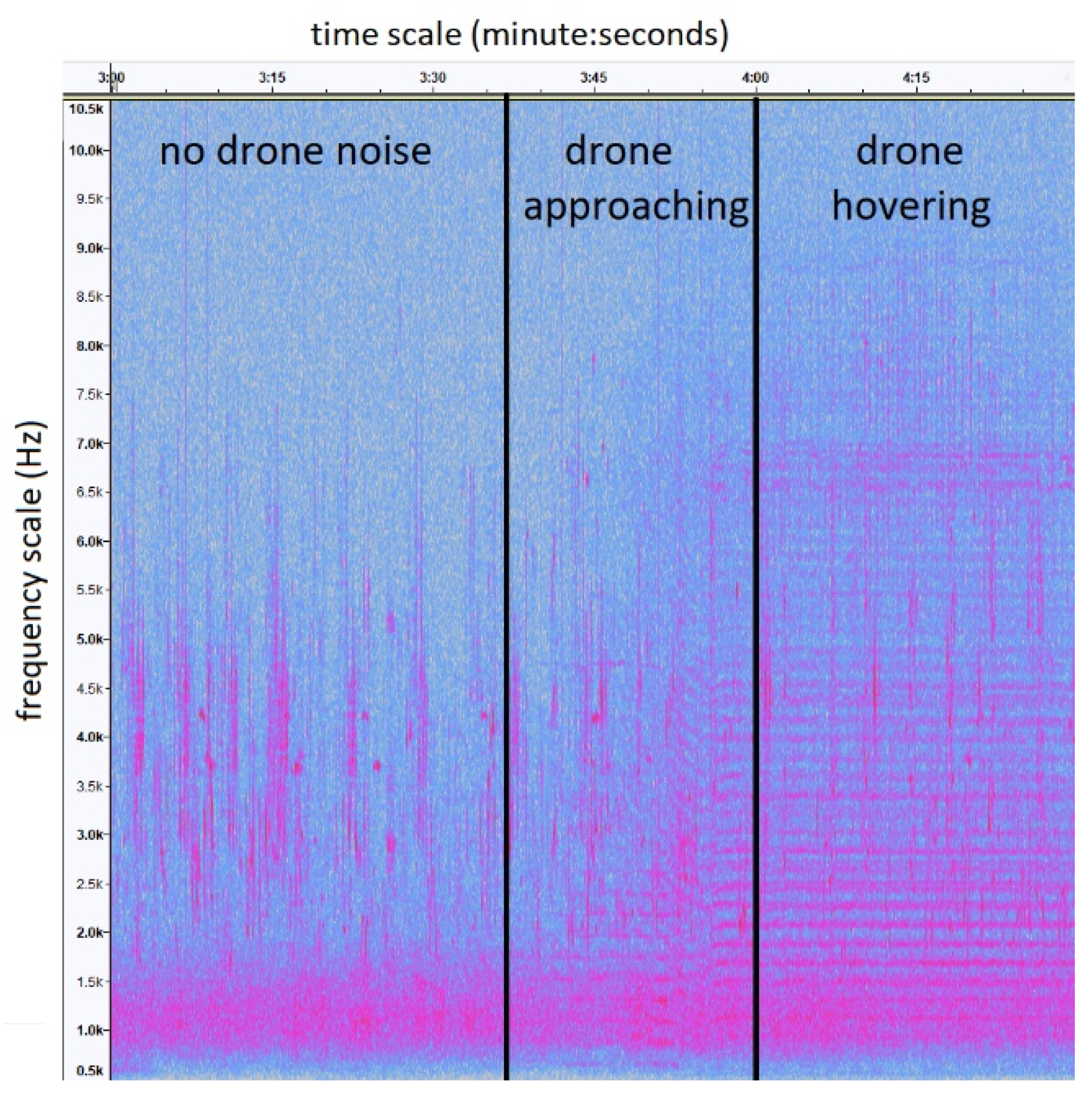{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Song Bout Detected | Localized Song Bouts | Birds within 50 m of Array Centers | ||
|---|---|---|---|---|
| Within 100 m of Array Centers | Within 50 m of Array Centers | |||
| American Robin | 1597 | 858 | 331 | 18 |
| Yellow Warbler | 1643 | 759 | 529 | 42 |
| Common Yellowthroat | 734 | 382 | 137 | 13 |
| Field Sparrow | 1388 | 532 | 388 | 31 |
| Song Sparrow | 1661 | 963 | 470 | 42 |
| Indigo Bunting | 611 | 357 | 241 | 14 |
| Northern Cardinal | 669 | 294 | 189 | 24 |
| All Seven Species | 8303 | 4145 | 2285 | 184 |
| Species | Model (See Figure 3) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 (Null) | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| AIC | ΔAIC | |||||||
| American Robin | 194.8 | 1.8 | 11.1 | 2.6 | 6.4 | 13.0 | 4.5 | 8.3 |
| Yellow Warbler | 451.5 | 1.9 | −5.3 | −6.8 | −8.0 | −3.4 | −4.9 | −6.1 |
| Common Yellowthroat | 131.8 | 0.5 | 11.8 | 3.0 | 6.7 | 12.8 | 4.0 | 7.7 |
| Field Sparrow | 316.5 | 1.9 | 6.6 | 3.2 | 6.2 | 8.5 | 5.2 | 8.1 |
| Song Sparrow | 400.6 | 0.2 | 14.0 | 3.0 | 6.9 | 14.1 | 3.2 | 7.1 |
| Indigo Bunting | 157.6 | 1.4 | 7.9 | 2.2 | 5.4 | 9.3 | 3.6 | 6.8 |
| Northern Cardinal | 224.1 | 2.0 | −2.7 | −11.7 | −8.6 | −0.7 | −9.8 | −6.7 |
| All Seven Species | 1898.6 | 0.7 | 11.4 | 1.3 | 4.1 | 12.2 | 2.0 | 4.8 |
| Yellow Warbler | ||||||
|---|---|---|---|---|---|---|
| Est | LL | UL | p | |||
| Intercept | 0.665 | 0.407 | 1.087 | 0.103 | ||
| During | { | Minute 5 | 0.662 | 0.297 | 1.472 | 0.311 |
| Minute 6 | 0.438 | 0.189 | 1.017 | 0.054 | ||
| Minute 7 | 0.189 | 0.066 | 0.495 | 0.001 | ||
| After | Minutes 9–11 | 0.483 | 0.271 | 0.860 | 0.013 | |
| Northern Cardinal | ||||||
| Est | LL | UL | p | |||
| Intercept | 0.111 | 0.050 | 0.248 | <0.001 | ||
| During | Minutes 5–6 | 1.462 | 0.544 | 3.925 | 0.450 | |
| After | Minutes 9–11 | 4.888 | 1.977 | 12.084 | <0.001 | |
| Species | Model (See Figure 3) | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 (null) | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| AIC | ΔAIC | |||||||
| American Robin | 264.1 | −4.0 | 7.2 | −0.4 | 1.8 | 2.5 | −5.5 | −4.0 |
| Yellow Warbler | 492.2 | 2.5 | 10.8 | 2.7 | 6.5 | 12.7 | 4.6 | 8.5 |
| Common Yellowthroat | 146.4 | 1.6 | 3.6 | 3.8 | 6.0 | 5.6 | 5.3 | 7.7 |
| Field Sparrow | 383.3 | 1.6 | 15.1 | 3.6 | 7.3 | 16.7 | 5.2 | 9.0 |
| Song Sparrow | 468.1 | 2.0 | 13.7 | 3.3 | 6.7 | 15.7 | 5.2 | 8.6 |
| Indigo Bunting | 202.5 | 1.7 | 10.3 | −0.1 | 3.4 | 12.2 | 1.8 | 5.4 |
| Northern Cardinal | 193.0 | 1.7 | 6.6 | 2.3 | −0.3 | 8.5 | 4.1 | 1.6 |
| All Seven Species | 2152.2 | −1.8 | 7.7 | 1.1 | 4.0 | 6.0 | −0.4 | 2.5 |
| American Robin | |||||
|---|---|---|---|---|---|
| Est | LL | UL | P | ||
| Intercept | 6.124 | 4.274 | 8.774 | <0.001 | |
| During | Minutes 5–7 | 0.847 | 0.605 | 1.188 | 0.337 |
| After | Minutes 9–11 | 1.240 | 0.922 | 1.669 | 0.155 |
| Distance | 0.984 | 0.972 | 0.995 | 0.005 | |
| Indigo Bunting | |||||
| Est | LL | Est | LL | ||
| Intercept | 4.846 | 3.786 | 6.204 | <0.001 | |
| During | Minutes 5–7 | 0.711 | 0.501 | 1.009 | 0.056 |
| After | Minutes 9–11 | 0.728 | 0.511 | 1.037 | 0.079 |
| Northern Cardinal | |||||
| Est | LL | Est | LL | ||
| Intercept | 3.792 | 2.494 | 5.764 | <0.001 | |
| During | Minute 5 | 0.759 | 0.351 | 1.640 | 0.483 |
| Minute 6 | 0.231 | 0.068 | 0.783 | 0.019 | |
| Minute 7 | 0.942 | 0.506 | 1.754 | 0.851 | |
| After | Minutes 9–11 | 0.782 | 0.491 | 1.247 | 0.302 |
| All Seven Species | |||||
| Est | LL | Est | LL | ||
| Intercept | 3.736 | 3.200 | 4.363 | <0.001 | |
| During | Minutes 5–7 | 0.907 | 0.804 | 1.024 | 0.115 |
| After | Minutes 9–11 | 0.968 | 0.862 | 1.087 | 0.586 |
| Distance | 0.996 | 0.991 | 1.000 | 0.055 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, A.M.; Boyle, K.S.; Gilmore, J.L.; Kiefer, C.J.; Walker, M.F. Species-Specific Responses of Bird Song Output in the Presence of Drones. Drones 2022, 6, 1. https://doi.org/10.3390/drones6010001
Wilson AM, Boyle KS, Gilmore JL, Kiefer CJ, Walker MF. Species-Specific Responses of Bird Song Output in the Presence of Drones. Drones. 2022; 6(1):1. https://doi.org/10.3390/drones6010001
Chicago/Turabian StyleWilson, Andrew M., Kenneth S. Boyle, Jennifer L. Gilmore, Cody J. Kiefer, and Matthew F. Walker. 2022. "Species-Specific Responses of Bird Song Output in the Presence of Drones" Drones 6, no. 1: 1. https://doi.org/10.3390/drones6010001
APA StyleWilson, A. M., Boyle, K. S., Gilmore, J. L., Kiefer, C. J., & Walker, M. F. (2022). Species-Specific Responses of Bird Song Output in the Presence of Drones. Drones, 6(1), 1. https://doi.org/10.3390/drones6010001