
Incorporating Geographical Scale and Multiple Environmental Factors to Delineate the Breeding Distribution of Sea Turtles




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
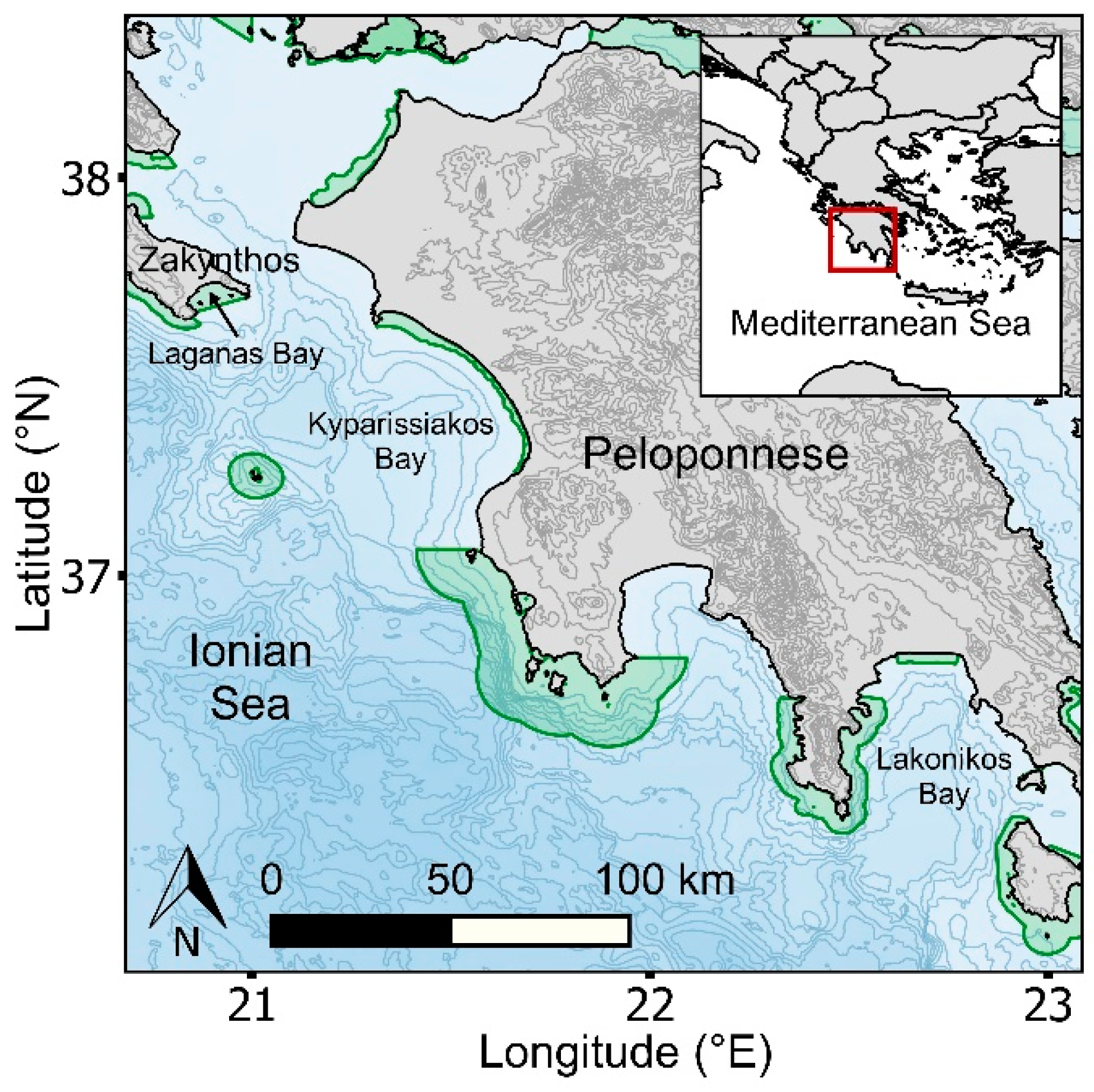
2.1. Study Region and Species
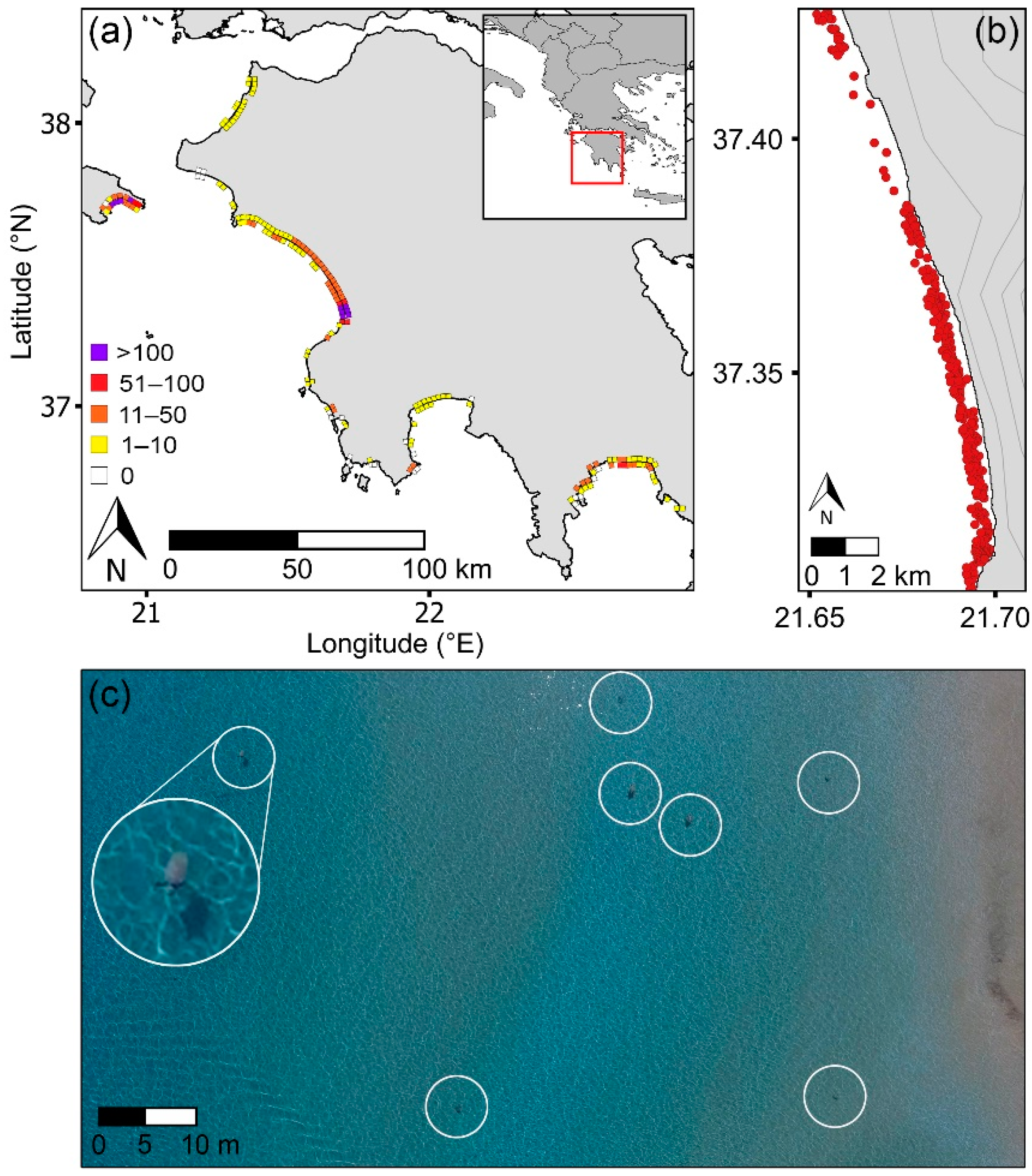
2.2. UAS Surveys and Data Processing
2.3. Temperature Records
2.4. Wind and Temperature Effects
2.5. Statistical Analysis
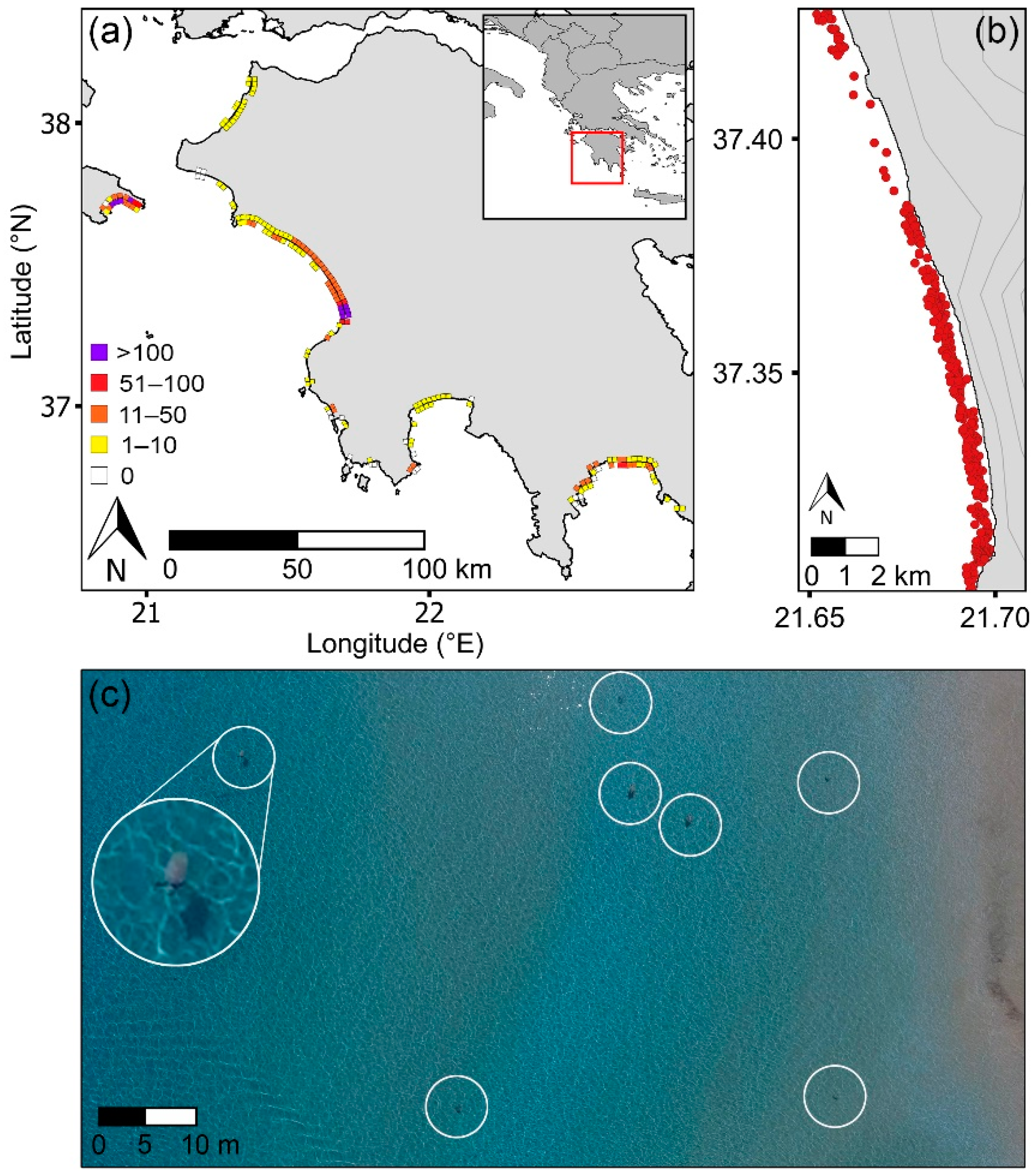
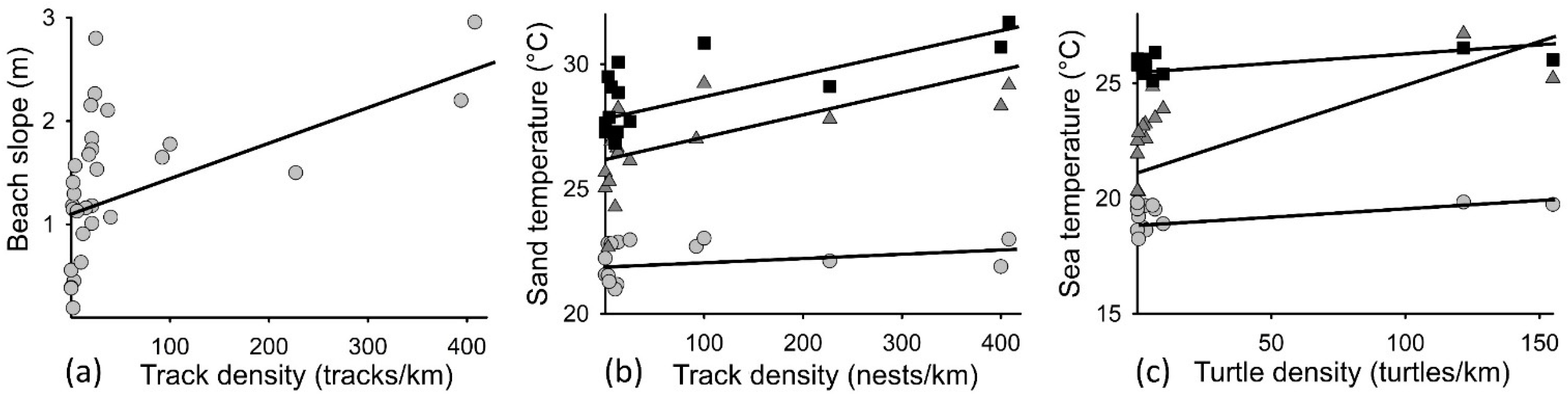
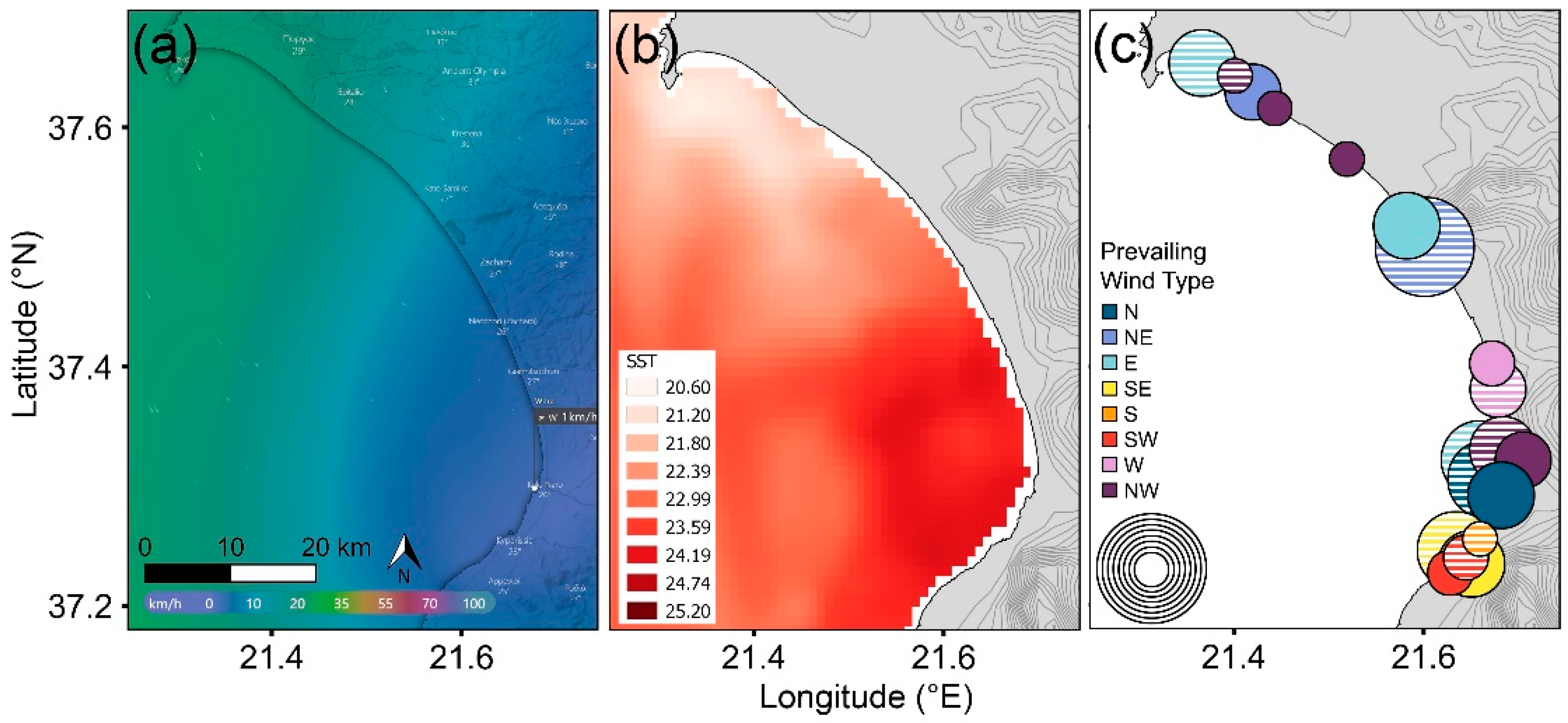
3. Results
3.1. Regional Distribution of Sea Turtles and Nests
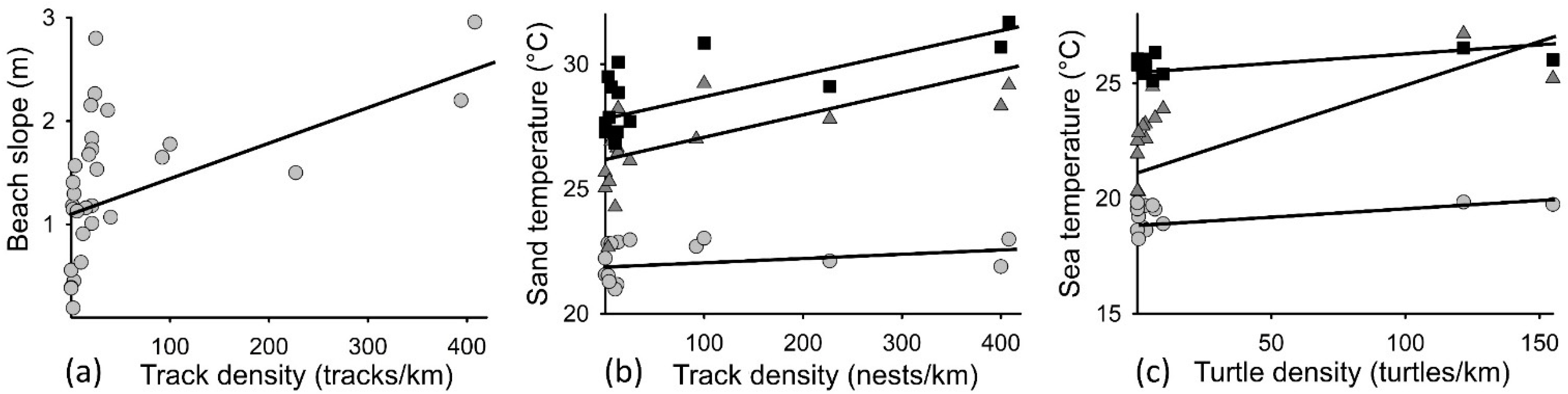
3.2. Inwater Turtle and Beach Track Densities Related to Temperature and Beach Slope
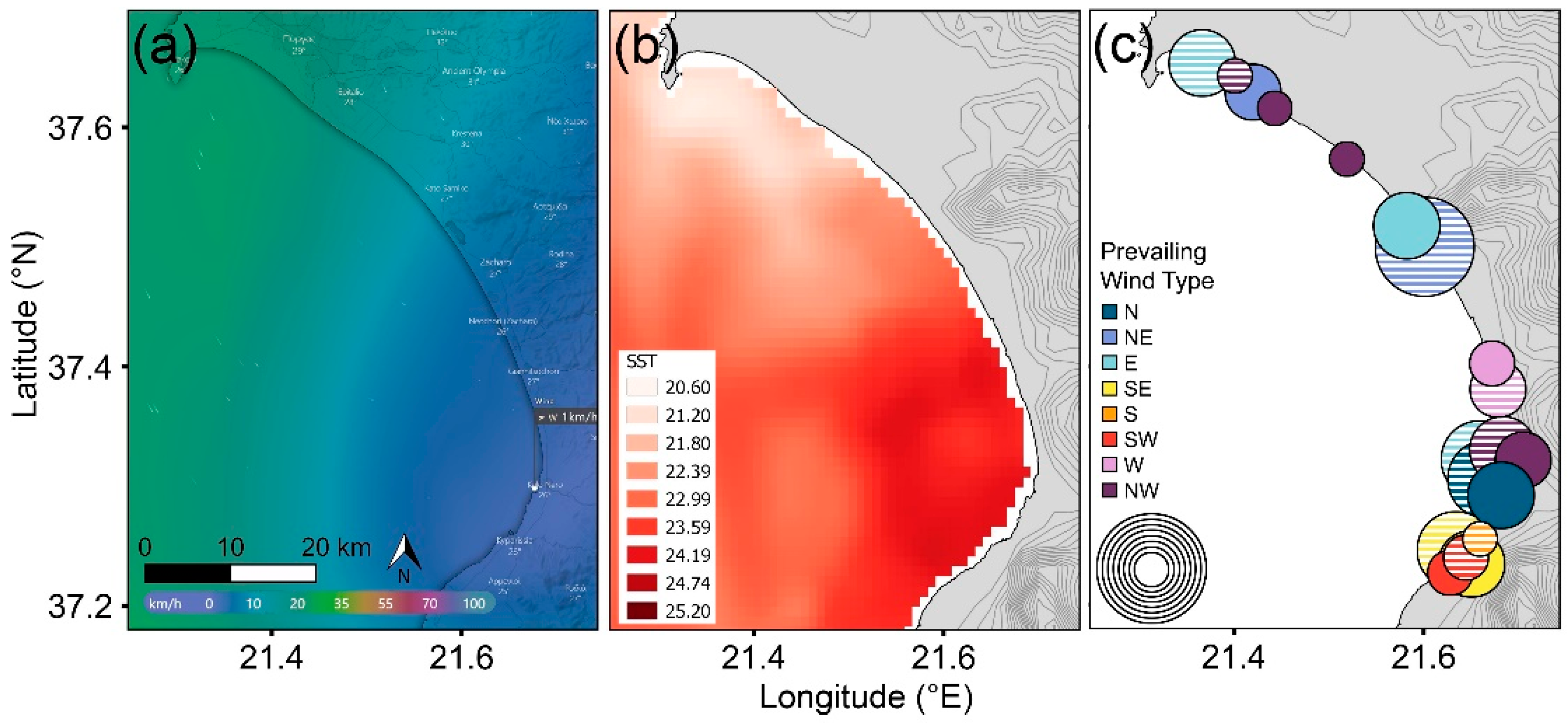
3.3. Rookery Locations Versus Wind and Sea Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harley, C.D.; Randall Hughes, A.; Hultgren, K.M.; Miner, B.G.; Sorte, C.J.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The impacts of climate change in coastal marine systems. Ecol. Lett. 2006, 9, 228–241. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Chivers, W.J.; Walne, A.W.; Hays, G.C. Mismatch between marine plankton range movements and the velocity of climate change. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Hays, G.C.; Bailey, H.; Bograd, S.J.; Bowen, W.D.; Campagna, C.; Carmichael, R.H.; Casale, P.; Chiaradia, A.; Costa, D.P.; Cuevas, E.; et al. Translating marine animal tracking data into conservation policy and management. Trends Ecol. Evol. 2019, 34, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.T.; Schoeman, D.S.; Buckley, L.B.; Moore, P.; Poloczanska, E.S.; Brander, K.M.; Brown, C.; Bruno, J.F.; Duarte, C.M.; Halpern, B.S.; et al. The pace of shifting climate in marine and terrestrial ecosystems. Science 2011, 334, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Carr, M.H.; Neigel, J.E.; Estes, J.A.; Andelman, S.; Warner, R.R.; Largier, J.L. Comparing marine and terrestrial ecosystems: Implications for the design of coastal marine reserves. Ecol. Appl. 2003, 13, S90–S107. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.C.; Hill, J.K.; Ohlemuller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Brown, C.J.; Sydeman, W.J.; Kiessling, W.; Schoeman, D.S.; Moore, P.J.; Brander, K.; Bruno, J.F.; Buckley, L.B.; Burrows, M.T.; et al. Global imprint of climate change on marine life. Nat. Clim. Chang. 2013, 3, 919–925. [Google Scholar] [CrossRef]
- Bates, A.E.; Helmuth, B.; Burrows, M.T.; Duncan, M.I.; Garrabou, J.; Guy-Haim, T.; Lima, F.; Queiros, A.M.; Seabra, R.; Marsh, R.; et al. Biologists ignore ocean weather at their peril. Nature 2018, 560, 299–301. [Google Scholar] [CrossRef] [Green Version]
- Siepielski, A.M.; Morrissey, M.B.; Buoro, M.; Carlson, S.M.; Caruso, C.M.; Clegg, S.M.; Coulson, T.; DiBattista, J.; Gotanda, K.M.; Francis, C.D.; et al. Precipitation drives global variation in natural selection. Science 2017, 355, 959–962. [Google Scholar] [CrossRef] [Green Version]
- VanDerWal, J.; Murphy, H.T.; Kutt, A.S.; Perkins, G.C.; Bateman, B.L.; Perry, J.J.; Reside, A.E. Focus on poleward shifts in species’ distribution underestimates the fingerprint of climate change. Nat. Clim. Chang. 2012, 3, 239–243. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Bruno, J.F. The impact of climate change on the world’s marine ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, J.M.; Hebblewhite, M.; Loison, A.; Fuller, M.; Powell, R.; Basille, M.; Van Moorter, B. Habitat-performance relationships: Finding the right metric at a given spatial scale. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2255–2265. [Google Scholar] [CrossRef]
- Pullin, A.S.; Knight, T.M.; Stone, D.A.; Charman, K. Do conservation managers use scientific evidence to support their decision-making? Biol. Conserv. 2004, 119, 245–252. [Google Scholar] [CrossRef]
- Grüss, A.; Kaplan, D.M.; Guénette, S.; Roberts, C.M.; Botsford, L.W. Consequences of adult and juvenile movement for marine protected areas. Biol. Conserv. 2011, 144, 692–702. [Google Scholar] [CrossRef]
- Hussey, N.E.; Kessel, S.T.; Aarestrup, K.; Cooke, S.J.; Cowley, P.D.; Fisk, A.T.; Harcourt, R.G.; Holland, K.N.; Iverson, S.J.; Kocik, J.F.; et al. Aquatic animal telemetry: A panoramic window into the underwater world. Science 2015, 348, 1255642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, D.W. Unoccupied Aircraft Systems in marine science and conservation. Annu. Rev. Mar. Sci. 2019, 11, 439–463. [Google Scholar] [CrossRef] [Green Version]
- Raoult, V.; Colefax, A.P.; Allan, B.M.; Cagnazzi, D.; Castelblanco-Martínez, N.; Ierodiaconou, D.; Johnston, D.W.; Landeo-Yauri, S.; Lyons, M.; Pirotta, V.; et al. Operational protocols for the use of drones in marine animal research. Drones 2020, 4, 64. [Google Scholar] [CrossRef]
- Schofield, G.; Esteban, N.; Katselidis, K.A.; Hays, G.C. Drones for research on sea turtles and other marine vertebrates—A review. Biol. Conserv. 2019, 238, 108214. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Colefax, A.P.; Tagliafico, A.; Bishop, M.J.; Giles, A.; Butcher, P.A. Assessing variation in assemblages of large marine fauna off ocean beaches using drones. Mar. Freshw. Res. 2019, 71, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Christie, K.S.; Gilbert, S.L.; Brown, C.L.; Hatfield, M.; Hanson, L. Unmanned aircraft systems in wildlife research: Current and future applications of a transformative technology. Front. Ecol. Environ. 2016, 14, 241–251. [Google Scholar] [CrossRef]
- Linchant, J.; Lisein, J.; Semeki, J.; Lejeune, P.; Vermeulen, C. Are unmanned aircraft systems (UASs) the future of wildlife monitoring? A review of accomplishments and challenges. Mammal. Rev. 2015, 45, 239–252. [Google Scholar] [CrossRef]
- Hays, G.C.; Hawkes, L.A. Satellite tracking sea turtles: Opportunities and challenges to address key questions. Front. Mar. Sci. 2018, 5, 432. [Google Scholar] [CrossRef]
- Mazaris, A.D.; Schofield, G.; Gkazinou, C.; Almpanidou, V.; Hays, G.C. Global sea turtle conservation successes. Sci. Adv. 2017, 3, e1600730. [Google Scholar] [CrossRef] [Green Version]
- Casale, P.; Tucker, A.D. Caretta Caretta (Amended Version of 2015 Assessment). 2017. Available online: https://www.biosfera1.com/wp-content/uploads/2021/01/Categoria-de-Amea%C3%A7a-Caretta-caretta-2017-10.2305_IUCN.UK_.2017-2.RLTS_.T3897A119333622.en_.pdf (accessed on 18 November 2021).
- Weishampel, J.F.; Bagley, D.A.; Ehrhart, L.M. Earlier nesting by loggerhead sea turtles following sea surface warming. Glob. Chang. Biol. 2004, 10, 1424–1427. [Google Scholar] [CrossRef]
- Rees, A.F.; Alfaro-Shigueto, J.; Barata, P.C.R.; Bjorndal, K.A.; Bolten, A.B.; Bourjea, J.; Broderick, A.C.; Campbell, L.M.; Cardona, L.; Carreras, C.; et al. Are we working towards global research priorities for management and conservation of sea turtles? Endanger. Species Res. 2016, 31, 337–382. [Google Scholar] [CrossRef] [Green Version]
- Culver, M.; Gibeaut, J.C.; Shaver, D.J.; Tissot, P.; Starek, M. Using LiDAR data to assess the relationship between beach geomorphology and Kemp’s Ridley (Lepidochelys kempii) nest site selection along Padre Island, TX, United States. Front. Mar. Sci. 2020, 7, 214. [Google Scholar] [CrossRef]
- Schofield, G.; Lilley, M.K.S.; Bishop, C.M.; Brown, P.; Katselidis, K.A.; Dimopoulos, P.; Pantis, J.D.; Hays, G.C. Conservation hotspots: Implications of intense spatial area use by breeding male and female loggerheads at the Mediterranean’s largest rookery. Endanger. Species Res. 2009, 10, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Schofield, G.; Bishop, C.M.; Katselidis, K.A.; Dimopoulos, P.; Pantis, J.D.; Hays, G.C. Microhabitat selection by sea turtles in a dynamic thermal marine environment. J. Anim. Ecol. 2009, 78, 14–21. [Google Scholar] [CrossRef]
- Weishampel, J.F.; Bagley, D.A.; Ehrhart, L.M. Intra-annual Loggerhead and Green Turtle Spatial Nesting Patterns. Southeast. Nat. 2006, 5, 453–462. [Google Scholar] [CrossRef]
- Almpanidou, V.; Schofield, G.; Kallimanis, A.S.; Türkozan, O.; Hays, G.C.; Mazaris, A.D. Using climatic suitability thresholds to identify past, present and future population viability. Ecol. Indic. 2016, 71, 551–556. [Google Scholar] [CrossRef]
- Dunkin, L.; Reif, M.; Altman, S.; Swannack, T. A spatially explicit, multi-criteria decision support model for loggerhead sea turtle nesting habitat suitability: A remote sensing-based approach. Remote Sens. 2016, 8, 573. [Google Scholar] [CrossRef] [Green Version]
- Bannister, N.; Holland, J.; Farrelly, T. Nest site fidelity of Flatback Turtles (Natator depressus) on Bare Sand Island, Northern Territory, Australia. North. Territ. Nat. 2016, 27, 47–53. [Google Scholar]
- Koch, A.U.; Guinea, M.L.; Whiting, S.D. Effects of sand erosion and current harvest practices on incubation of the flatback sea turtle (Natator depressus). Aust. J. Zool. 2007, 55, 97–105. [Google Scholar] [CrossRef]
- Koch, A.U.; Guinea, M.L. Lower nesting success of flatback turtles caused by disorientation. Mar. Turt. Newsl. 2006, 114, 16. [Google Scholar]
- Garcon, J.S.; Grech, A.; Moloney, J.; Hamann, M. Relative Exposure Index: An important factor in sea turtle nesting distribution. Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 140–149. [Google Scholar] [CrossRef]
- Fonseca, J.G.; Cordeiro, C.A.M.M.; Ferreira, C.E.L. Spatial distribution of sea turtles on South Atlantic subtropical reefs. Mar. Ecol. Prog. Ser. 2021, 678, 125–138. [Google Scholar] [CrossRef]
- Schofield, G.; Bishop, C.M.; MacLean, G.; Brown, P.; Baker, M.; Katselidis, K.A.; Dimopoulos, P.; Pantis, J.D.; Hays, G.C. Novel GPS tracking of sea turtles as a tool for conservation management. J. Exp. Mar. Biol. Ecol. 2007, 347, 58–68. [Google Scholar] [CrossRef]
- Hawkes, L.A.; Broderick, A.C.; Godfrey, M.H.; Godley, B.J. Climate change and marine turtles. Endanger. Species Res. 2009, 7, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Witt, M.J.; Hawkes, L.A.; Godfrey, M.H.; Godley, B.J.; Broderick, A.C. Predicting the impacts of climate change on a globally distributed species: The case of the loggerhead turtle. J. Exp. Biol. 2010, 213, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, M.M.P.B.; Maynard, J.A.; Guinea, M.; Bell, I.P.; Werdell, P.J.; Hamann, M. Proxy indicators of sand temperature help project impacts of global warming on sea turtles in northern Australia. Endanger. Species Res. 2009, 9, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Casale, P.; Broderick, A.C.; Camiñas, J.A.; Cardona, L.; Carreras, C.; Demetropoulos, A.; Fuller, W.J.; Godley, B.J.; Hochscheid, S.; Kaska, Y.; et al. Mediterranean sea turtles: Current knowledge and priorities for conservation and research. Endanger. Species Res. 2018, 36, 229–267. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Casale, P. Loggerhead Turtle (Caretta caretta)—Mediterranean Subpopulation. 2015. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwjts9bLgLX0AhXmrlYBHXitC40QFnoECAMQAQ&url=https%3A%2F%2Fwww.iucnredlist.org%2Fspecies%2Fpdf%2F83646294%2Fattachment&usg=AOvVaw2OD_PogM4JBFHcTK7HZDu6 (accessed on 18 November 2021).
- Casale, P.; Margaritoulis, D. Sea Turtles in the Mediterranean: Distribution, Threats and Conservation Priorities; International Union for Conservation of Nature (IUCN): Gland, Switzerland, 2010. [Google Scholar]
- Dodd, C.K. Synopsis of the Biological Data on the Loggerhead Sea Turtle Caretta Caretta (Linnaeus 1758); United States Fish and Wildlife Service: Washington, DC, USA, 1988.
- Margaritoulis, D. Nesting activity and reproductive output of loggerhead sea turtles, Caretta caretta, over 19 seasons (1984–2002) at Laganas Bay, Zakynthos, Greece: The largest rookery in the Mediterranean. Chelonian Conserv. Biol. 2005, 4, 916–929. [Google Scholar]
- Schofield, G.; Dimadi, A.; Fossette, S.; Katselidis, K.A.; Koutsoubas, D.; Lilley, M.K.S.; Luckman, A.; Pantis, J.D.; Karagouni, A.D.; Hays, G.C.; et al. Satellite tracking large numbers of individuals to infer population level dispersal and core areas for the protection of an endangered species. Divers. Distrib. 2013, 19, 834–844. [Google Scholar] [CrossRef]
- Hays, G.C.; Broderick, A.C.; Glen, F.; Godley, B.J.; Houghton, J.D.R.; Metcalfe, J.D. Water temperature and internesting intervals for loggerhead (Caretta caretta) and green (Chelonia mydas) sea turtles. J. Therm. Biol. 2002, 27, 429–432. [Google Scholar] [CrossRef]
- Fossette, S.; Schofield, G.; Lilley, M.K.S.; Gleiss, A.C.; Hays, G.C. Acceleration data reveal the energy management strategy of a marine ectotherm during reproduction. Funct. Ecol. 2012, 26, 324–333. [Google Scholar] [CrossRef]
- Katselidis, K.A.; Schofield, G.; Stamou, G.; Dimopoulos, P.; Pantis, J.D.; Katzner, T. Females first? Past, present and future variability in offspring sex ratio at a temperate sea turtle breeding area. Anim. Conserv. 2012, 15, 508–518. [Google Scholar] [CrossRef]
- Schofield, G.; Scott, R.; Dimadi, A.; Fossette, S.; Katselidis, K.A.; Koutsoubas, D.; Lilley, M.K.S.; Pantis, J.D.; Karagouni, A.D.; Hays, G.C. Evidence-based marine protected area planning for a highly mobile endangered marine vertebrate. Biol. Conserv. 2013, 161, 101–109. [Google Scholar] [CrossRef]
- Schofield, G.; Katselidis, K.A.; Lilley, M.K.S.; Reina, R.D.; Hays, G.C. Detecting elusive aspects of wildlife ecology using drones: New insights on the mating dynamics and operational sex ratios of sea turtles. Funct. Ecol. 2017, 31, 2310–2319. [Google Scholar] [CrossRef]
- Dujon, A.M.; Ierodiaconou, D.; Geeson, J.J.; Arnould, J.P.Y.; Allan, B.M.; Katselidis, K.A.; Schofield, G.; Scales, K.; Bouchet, P. Machine learning to detect marine animals in UAV imagery: Effect of morphology, spacing, behaviour and habitat. Remote. Sens. Ecol. Conserv. 2021, 7, 341–354. [Google Scholar] [CrossRef]
- Katselidis, K.A.; Schofield, G.; Stamou, G.; Dimopoulos, P.; Pantis, J.D. Employing sea-level rise scenarios to strategically select sea turtle nesting habitat important for long-term management at a temperate breeding area. J. Exp. Mar. Biol. Ecol. 2014, 450, 47–54. [Google Scholar] [CrossRef]
- Agisoft Metashape, 1.6.5; Agisoft LLC: Saint Petersburg, Russia, 2021.
- Carvalho, R.C.; Kennedy, D.M.; Niyazi, Y.; Leach, C.; Konlechner, T.M.; Ierodiaconou, D. Structure-from-motion photogrammetry analysis of historical aerial photography: Determining beach volumetric change over decadal scales. Earth Surf. Process. Landf. 2020, 45, 2540–2555. [Google Scholar] [CrossRef]
- QGIS Geographic Information System, 3.16.8; QGIS Association. 2021. Available online: https://www.qgis.org/en/site/getinvolved/governance/charter/index.html (accessed on 18 November 2021).
- Katselidis, K.A.; Schofield, G.; Stamou, G.; Dimopoulos, P.; Pantis, J.D. Evidence-based management to regulate the impact of tourism at a key marine turtle rookery on Zakynthos Island, Greece. Oryx 2013, 47, 584–594. [Google Scholar] [CrossRef] [Green Version]
- Buongiorno Nardelli, B.; Tronconi, C.; Pisano, A.; Santoleri, R. High and ultra-high resolution processing of satellite sea surface temperature data over southern European seas in the framework of MyOcean project. Remote Sens. Environ. 2013, 129, 1–16. [Google Scholar] [CrossRef]
- Amante, C.; Eakins, B.W. ETOPO1 arc-minute global relief model: Procedures, data sources and analysis. NOAA technical memorandum NESDIS NGDC-24. Natl. Geophys. Data Cent. NOAA 2009, 10, V5C8276M. [Google Scholar]
- GADM Database of Global Administrative Areas, Version 2.0. Available online: www.gadm.org (accessed on 15 May 2020).
- Androulidakis, Y.S.; Kombiadou, K.D.; Makris, C.V.; Baltikas, V.N.; Krestenitis, Y.N. Storm surges in the Mediterranean Sea: Variability and trends under future climatic conditions. Dyn. Atmos. Ocean 2015, 71, 56–82. [Google Scholar] [CrossRef]
- Makris, C.; Galiatsatou, P.; Tolika, K.; Anagnostopoulou, C.; Kombiadou, K.; Prinos, P.; Velikou, K.; Kapelonis, Z.; Tragou, E.; Androulidakis, Y.; et al. Climate change effects on the marine characteristics of the Aegean and Ionian Seas. Ocean Dyn. 2016, 66, 1603–1635. [Google Scholar] [CrossRef]
- Vagenas, C.; Anagnostopoulou, C.; Tolika, K. Climatic study of the marine surface wind field over the Greek seas with the use of a high resolution RCM focusing on extreme winds. Climate 2017, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Windyty, S.E. Available online: www.windy.com (accessed on 30 September 2020).
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Grenyer, R.; Orme, C.D.; Jackson, S.F.; Thomas, G.H.; Davies, R.G.; Davies, T.J.; Jones, K.E.; Olson, V.A.; Ridgely, R.S.; Rasmussen, P.C.; et al. Global distribution and conservation of rare and threatened vertebrates. Nature 2006, 444, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Sirami, C.; Seymour, C.; Midgley, G.; Barnard, P. The impact of shrub encroachment on savanna bird diversity from local to regional scale. Divers. Distrib. 2009, 15, 948–957. [Google Scholar] [CrossRef]
- Forey, E.; Chapelet, B.; Vitasse, Y.; Tilquin, M.; Touzard, B.; Michalet, R. The relative importance of disturbance and environmental stress at local and regional scales in French coastal sand dunes. J. Veg. Sci. 2008, 19, 493–502. [Google Scholar] [CrossRef]
- Lowen, J.B.; McKindsey, C.W.; Therriault, T.W.; DiBacco, C. Effects of spatial resolution on predicting the distribution of aquatic invasive species in nearshore marine environments. Mar. Ecol. Prog. Ser. 2016, 556, 17–30. [Google Scholar] [CrossRef]
- Harley, C.D.G.; Helmuth, B.S.T. Local- and regional-scale effects of wave exposure, thermal stress, and absolute versus effective shore level on patterns of intertidal zonation. Limnol. Oceanogr. 2003, 48, 1498–1508. [Google Scholar] [CrossRef]
- Fauchald, P.; Erikstad, K.E.; Skarsfjord, H. Scale-dependent predator–prey Interactions: The hierarchical spatial distribution of seabirds and prey. Ecology 2000, 81, 773–783. [Google Scholar] [CrossRef]
- Wood, D.W.; Bjorndal, K.A.; Ross, S.T. Relation of temperature, moisture, salinity, and slope to nest site selection in loggerhead sea turtles. Copeia 2000, 2000, 119. [Google Scholar] [CrossRef] [Green Version]
- Hamel, M.A.; McMahon, C.R.; Bradshaw, C.J.A. Flexible inter-nesting behaviour of generalist olive ridley turtles in Australia. J. Exp. Mar. Biol. Ecol. 2008, 359, 47–54. [Google Scholar] [CrossRef]
- Sato, K.; Matsuzawa, Y.; Tanaka, H.; Bando, T.; Minamikawa, S.; Sakamoto, W.; Naito, Y. Internesting intervals for loggerhead turtles, Caretta caretta, and green turtles, Chelonia mydas, are affected by temperature. Can. J. Zool. 1998, 76, 1651–1662. [Google Scholar] [CrossRef]
- Laloe, J.O.; Esteban, N.; Berkel, J.; Hays, G.C. Sand temperatures for nesting sea turtles in the Caribbean: Implications for hatchling sex ratios in the face of climate change. J. Exp. Mar. Biol. Ecol. 2016, 474, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Godley, B.J.; Broderick, A.C.; Glen, F.; Hays, G.C. Temperature-dependent sex determination of Ascension Island green turtles. Mar. Ecol. Prog. Ser. 2002, 226, 115–124. [Google Scholar] [CrossRef]
- Hughes, D.A.; Richard, J.D. The nesting of the Pacific Ridley Turtle Lepidochelys olivacea on Playa Naneite, Costa Rica. Mar. Biol. 1974, 24, 97–107. [Google Scholar] [CrossRef]
- Godley, B.J.; Broderick, A.C.; Frauenstein, R.; Glen, F.; Hays, G.C. Reproductive seasonality and sexual dimorphism in green turtles. Mar. Ecol. Prog. Ser. 2002, 226, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Arendt, M.D.; Segars, A.L.; Byrd, J.I.; Boynton, J.; Whitaker, J.D.; Parker, L.; Owens, D.W.; Blanvillain, G.; Quattro, J.M.; Roberts, M.A. Distributional patterns of adult male loggerhead sea turtles (Caretta caretta) in the vicinity of Cape Canaveral, Florida, USA during and after a major annual breeding aggregation. Mar. Biol. 2011, 159, 101–112. [Google Scholar] [CrossRef]
- Hays, G.C.; Luschi, P.; Papi, F.; del Seppia, C.; Marsh, R. Changes in behaviour during the inter-nesting period and post-nesting migration for Ascension Island green turtles. Mar. Ecol. Prog. Ser. 1999, 189, 263–273. [Google Scholar] [CrossRef]
- Kaplan, D.M.; Largier, J.L.; Navarrete, S.; Guiñez, R.; Castilla, J.C. Large diurnal temperature fluctuations in the nearshore water column. Estuar. Coast. Shelf Sci. 2003, 57, 385–398. [Google Scholar] [CrossRef]
- Barnes, R.S.K.; Mann, K.H. Fundamentals of Aquatic Ecology; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 1444314106. [Google Scholar]
- Narváez, D.A.; Navarrete, S.A.; Largier, J.; Vargas, C.A. Onshore advection of warm water, larval invertebrate settlement, and relaxation of upwelling off central Chile. Mar. Ecol. Prog. Ser. 2006, 309, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Tapia, F.J.; Pineda, J.; Ocampo-Torres, F.J.; Fuchs, H.L.; Parnell, P.E.; Montero, P.; Ramos, S. High-frequency observations of wind-forced onshore transport at a coastal site in Baja California. Cont. Shelf Res. 2004, 24, 1573–1585. [Google Scholar] [CrossRef]
- Weimerskirch, H.; Louzao, M.; de Grissac, S.; Delord, K. Changes in wind pattern alter albatross distribution and life-history traits. Science 2012, 335, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Barlow, D.R.; Klinck, H.; Ponirakis, D.; Garvey, C.; Torres, L.G. Temporal and spatial lags between wind, coastal upwelling, and blue whale occurrence. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Dehnhard, N.; Ludynia, K.; Poisbleau, M.; Demongin, L.; Quillfeldt, P. Good days, bad days: Wind as a driver of foraging success in a flightless seabird, the southern rockhopper penguin. PLoS ONE 2013, 8, e79487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiting, S.D.; Long, J.L.; Hadden, K.M.; Lauder, A.D.K.; Koch, A.U. Insights into size, seasonality and biology of a nesting population of the Olive Ridley turtle in northern Australia. Wildl. Res. 2007, 34, 200–210. [Google Scholar] [CrossRef]
- Shaver, D.J.; Tissot, P.E.; Streich, M.M.; Walker, J.S.; Rubio, C.; Amos, A.F.; George, J.A.; Pasawicz, M.R. Hypothermic stunning of green sea turtles in a western Gulf of Mexico foraging habitat. PLoS ONE 2017, 12, e0173920. [Google Scholar] [CrossRef] [Green Version]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Rodwell, M.J.; Rowell, D.P.; Folland, C.K. Oceanic forcing of the wintertime North Atlantic Oscillation and European climate. Nature 1999, 398, 320–323. [Google Scholar] [CrossRef]
- Brown, A.C.; McLachlan, A. The Ecology of Sandy Shores; Elsevier: Amsterdam, The Netherlands, 2006; ISBN 978-0-12-372569-1. [Google Scholar]
- McLachlan, A. Sandy beach ecology—A review. In Sandy Beaches as Ecosystems. Developments in Hydrobiology; Springer: Dordrecht, The Netherlands, 1983; Volume 19, pp. 321–380. ISBN 978-90-481-8521-4. [Google Scholar]
- Babcock, R.C.; Bustamante, R.H.; Fulton, E.A.; Fulton, D.J.; Haywood, M.D.E.; Hobday, A.J.; Kenyon, R.; Matear, R.J.; Plagányi, E.E.; Richardson, A.J.; et al. Severe continental-scale impacts of climate change are happening now: Extreme climate events impact marine habitat forming communities along 45% of Australia’s coast. Front. Mar. Sci. 2019, 6, 411. [Google Scholar] [CrossRef]
- Gill, P.C.; Morrice, M.G.; Page, B.; Pirzl, R.; Levings, A.H.; Coyne, M. Blue whale habitat selection and within-season distribution in a regional upwelling system off southern Australia. Mar. Ecol. Prog. Ser. 2011, 421, 243–263. [Google Scholar] [CrossRef] [Green Version]
- Rubidge, E.M.; Monahan, W.B.; Parra, J.L.; Cameron, S.E.; Brashares, J.S. The role of climate, habitat, and species co-occurrence as drivers of change in small mammal distributions over the past century. Glob. Chang. Biol. 2011, 17, 696–708. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Yates, K.L.; Bouchet, P.J.; Caley, M.J.; Mengersen, K.; Randin, C.F.; Parnell, S.; Fielding, A.H.; Bamford, A.J.; Ban, S.; Barbosa, A.M.; et al. Outstanding challenges in the transferability of ecological models. Trends Ecol. Evol. 2018, 33, 790–802. [Google Scholar] [CrossRef] [Green Version]
- Switzer, P.V. Site fidelity in predictable and unpredictable habitats. Evol. Ecol. 1993, 7, 533–555. [Google Scholar] [CrossRef]
- Weishampel, J.F.; Bagley, D.A.; Ehrhart, L.M.; Rodenbeck, B.L. Spatiotemporal patterns of annual sea turtle nesting behaviors along an East Central Florida beach. Biol. Conserv. 2003, 110, 295–303. [Google Scholar] [CrossRef]
- Hays, G.C.; Ashworth, J.S.; Barnsley, M.J.; Broderick, A.C.; Emery, D.R.; Godley, B.J.; Henwood, A.; Jones, E.L. The importance of sand albedo for the thermal conditions on sea turtle nesting beaches. Oikos 2001, 93, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Patrício, A.R.; Marques, A.; Barbosa, C.; Broderick, A.C.; Godley, B.J.; Hawkes, L.A.; Rebelo, R.; Regalla, A.; Catry, P. Balanced primary sex ratios and resilience to climate change in a major sea turtle population. Mar. Ecol. Prog. Ser. 2017, 577, 189–203. [Google Scholar] [CrossRef]
- Long, T.M.; Angelo, J.; Weishampel, J.F. LiDAR-derived measures of hurricane- and restoration-generated beach morphodynamics in relation to sea turtle nesting behaviour. Int. J. Remote Sens. 2011, 32, 231–241. [Google Scholar] [CrossRef]
- Dawson, T.P.; Jackson, S.T.; House, J.I.; Prentice, I.C.; Mace, G.M. Beyond predictions: Biodiversity conservation in a changing climate. Science 2011, 332, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Hawkes, L.A.; Broderick, A.C.; Godfrey, M.H.; Godley, B.J. Investigating the potential impacts of climate change on a marine turtle population. Glob. Chang. Biol. 2007, 13, 923–932. [Google Scholar] [CrossRef]
- Edgar, G.J.; Stuart-Smith, R.D.; Willis, T.J.; Kininmonth, S.; Baker, S.C.; Banks, S.; Barrett, N.S.; Becerro, M.A.; Bernard, A.T.; Berkhout, J.; et al. Global conservation outcomes depend on marine protected areas with five key features. Nature 2014, 506, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, L.; Day, J.O.N.; Lewis, A.; Slegers, S.; Kerrigan, B.; Breen, D.A.N.; Cameron, D.; Jago, B.; Hall, J.; Lowe, D.; et al. Establishing representative no-take areas in the Great Barrier Reef: Large-scale implementation of theory on Marine Protected Areas. Conserv. Biol. 2005, 19, 1733–1744. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, R.L. Systematic Conservation Planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Pike, D.A. Climate influences the global distribution of sea turtle nesting. Glob. Ecol. Biogeogr. 2013, 22, 555–566. [Google Scholar] [CrossRef]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T.; et al. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579–583. [Google Scholar] [CrossRef]
- Opdam, P.; Wascher, D. Climate change meets habitat fragmentation: Linking landscape and biogeographical scale levels in research and conservation. Biol. Conserv. 2004, 117, 285–297. [Google Scholar] [CrossRef]
- Oliver, T.A.; Palumbi, S.R. Distributions of stress-resistant coral symbionts match environmental patterns at local but not regional scales. Mar. Ecol. Prog. Ser. 2009, 378, 93–103. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dickson, L.C.; Katselidis, K.A.; Eizaguirre, C.; Schofield, G. Incorporating Geographical Scale and Multiple Environmental Factors to Delineate the Breeding Distribution of Sea Turtles. Drones 2021, 5, 142. https://doi.org/10.3390/drones5040142
Dickson LC, Katselidis KA, Eizaguirre C, Schofield G. Incorporating Geographical Scale and Multiple Environmental Factors to Delineate the Breeding Distribution of Sea Turtles. Drones. 2021; 5(4):142. https://doi.org/10.3390/drones5040142
Chicago/Turabian StyleDickson, Liam C., Kostas A. Katselidis, Christophe Eizaguirre, and Gail Schofield. 2021. "Incorporating Geographical Scale and Multiple Environmental Factors to Delineate the Breeding Distribution of Sea Turtles" Drones 5, no. 4: 142. https://doi.org/10.3390/drones5040142
APA StyleDickson, L. C., Katselidis, K. A., Eizaguirre, C., & Schofield, G. (2021). Incorporating Geographical Scale and Multiple Environmental Factors to Delineate the Breeding Distribution of Sea Turtles. Drones, 5(4), 142. https://doi.org/10.3390/drones5040142