
A Mathematical Investigation of Sex Differences in Alzheimer’s Disease


Abstract
1. Introduction
2. Materials and Methods
2.1. Mathematical Model
2.2. Numerical Scheme
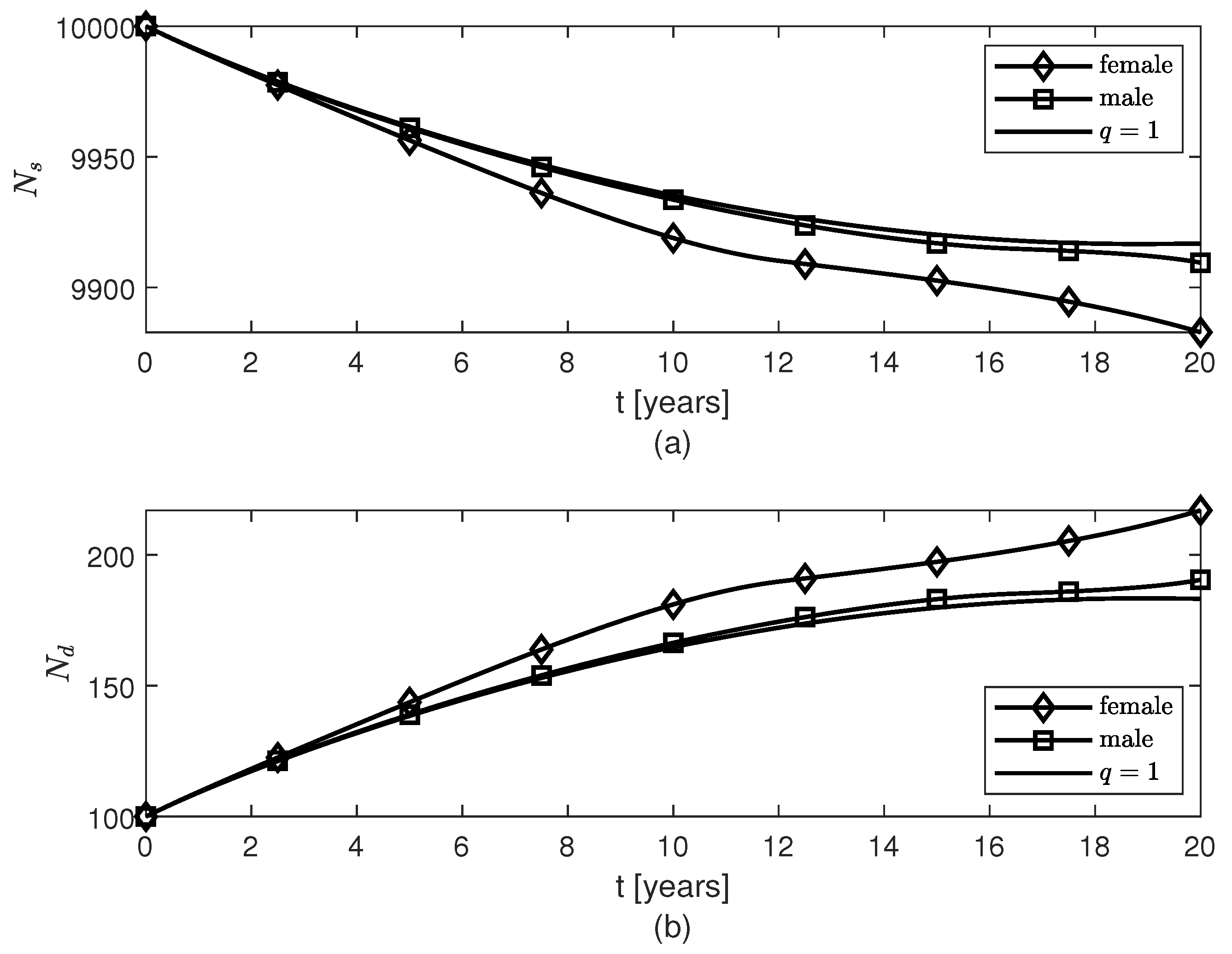
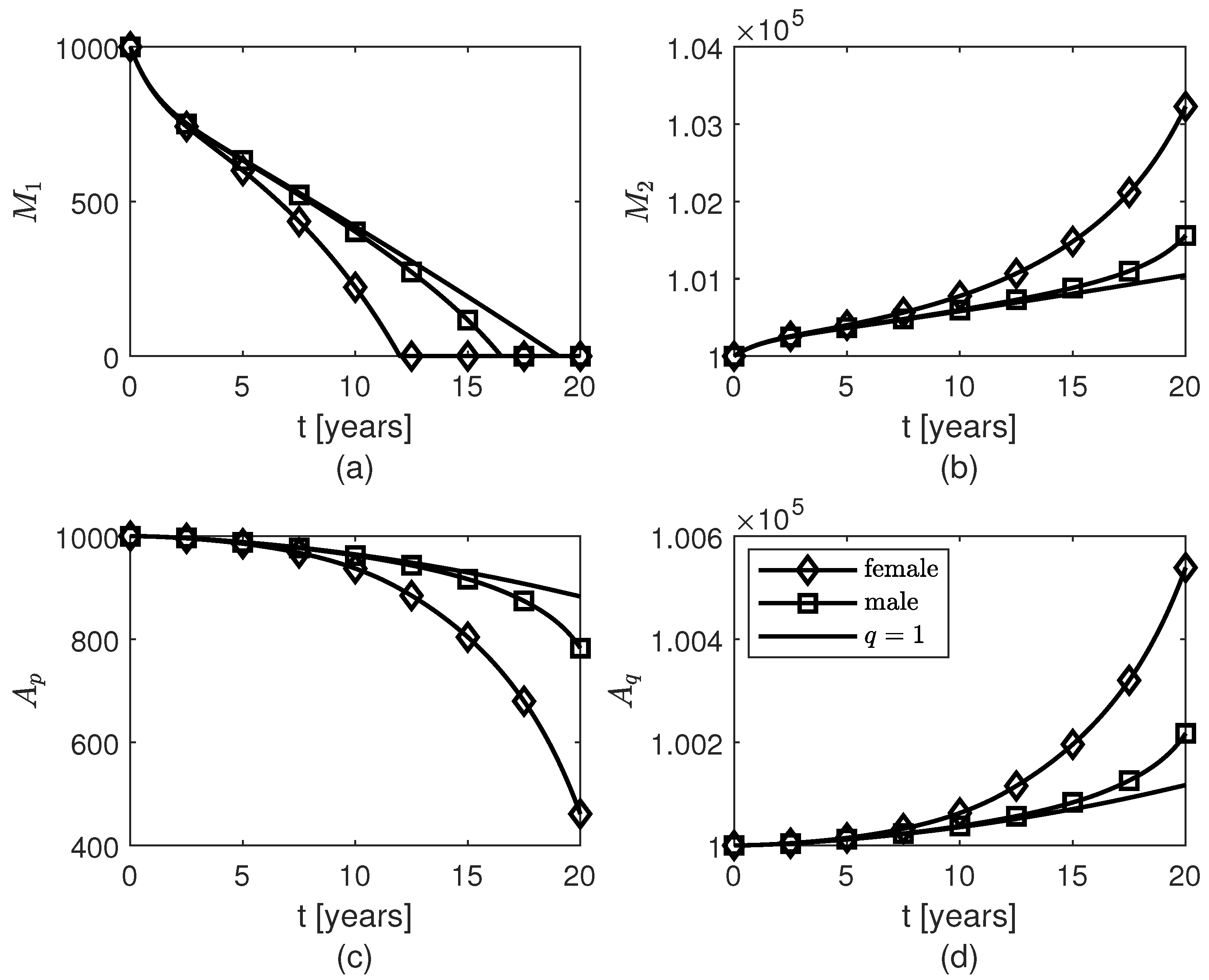
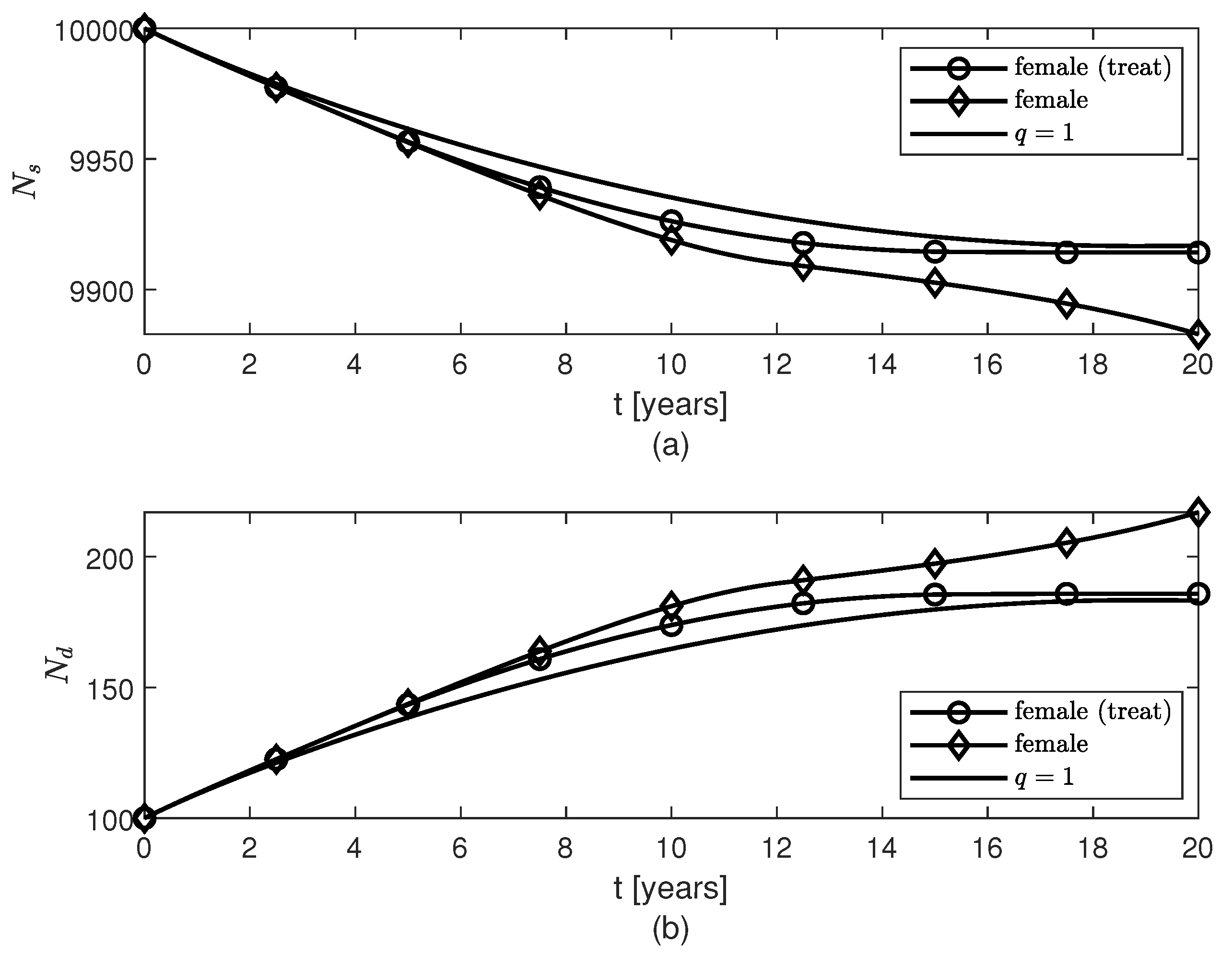
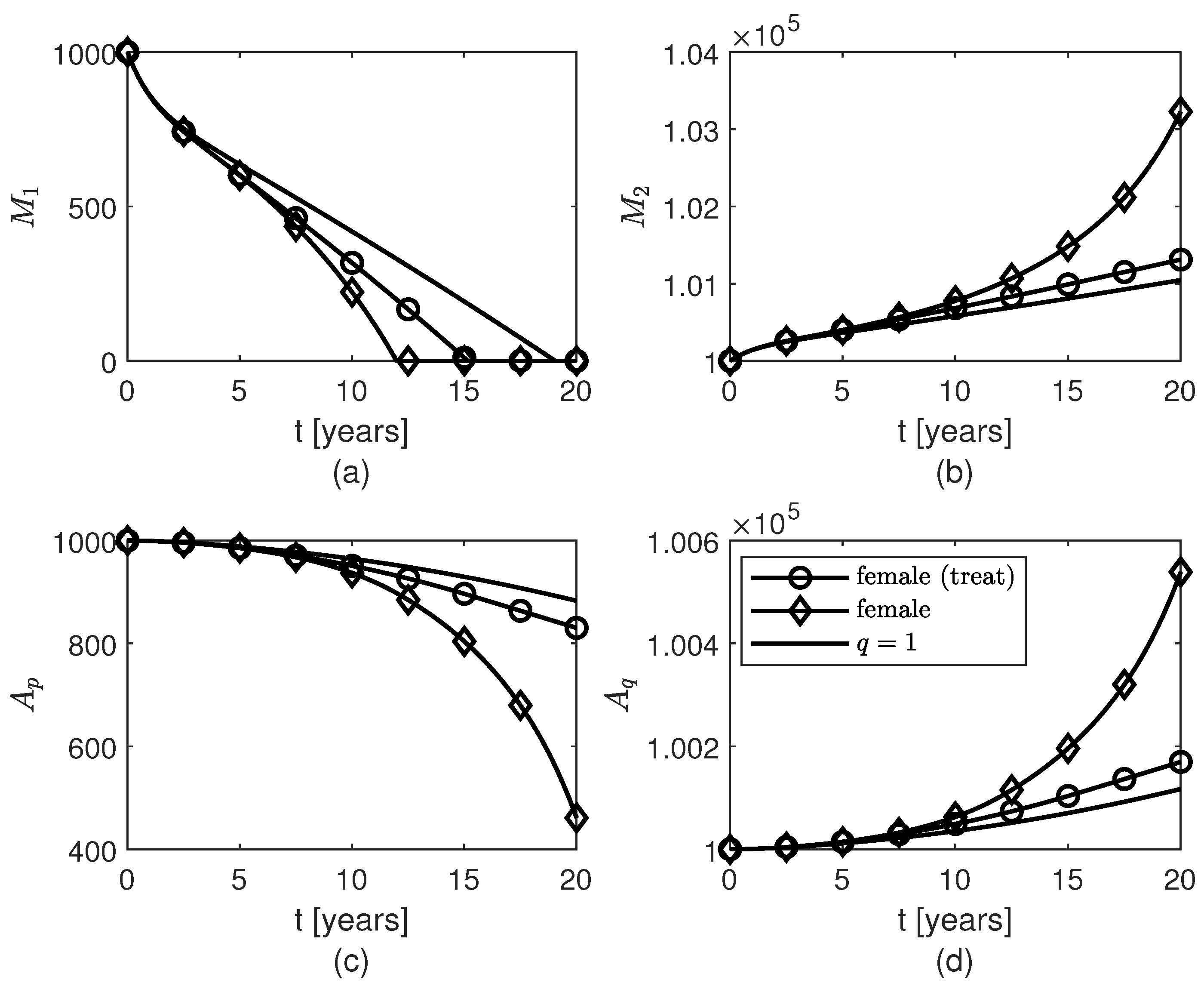
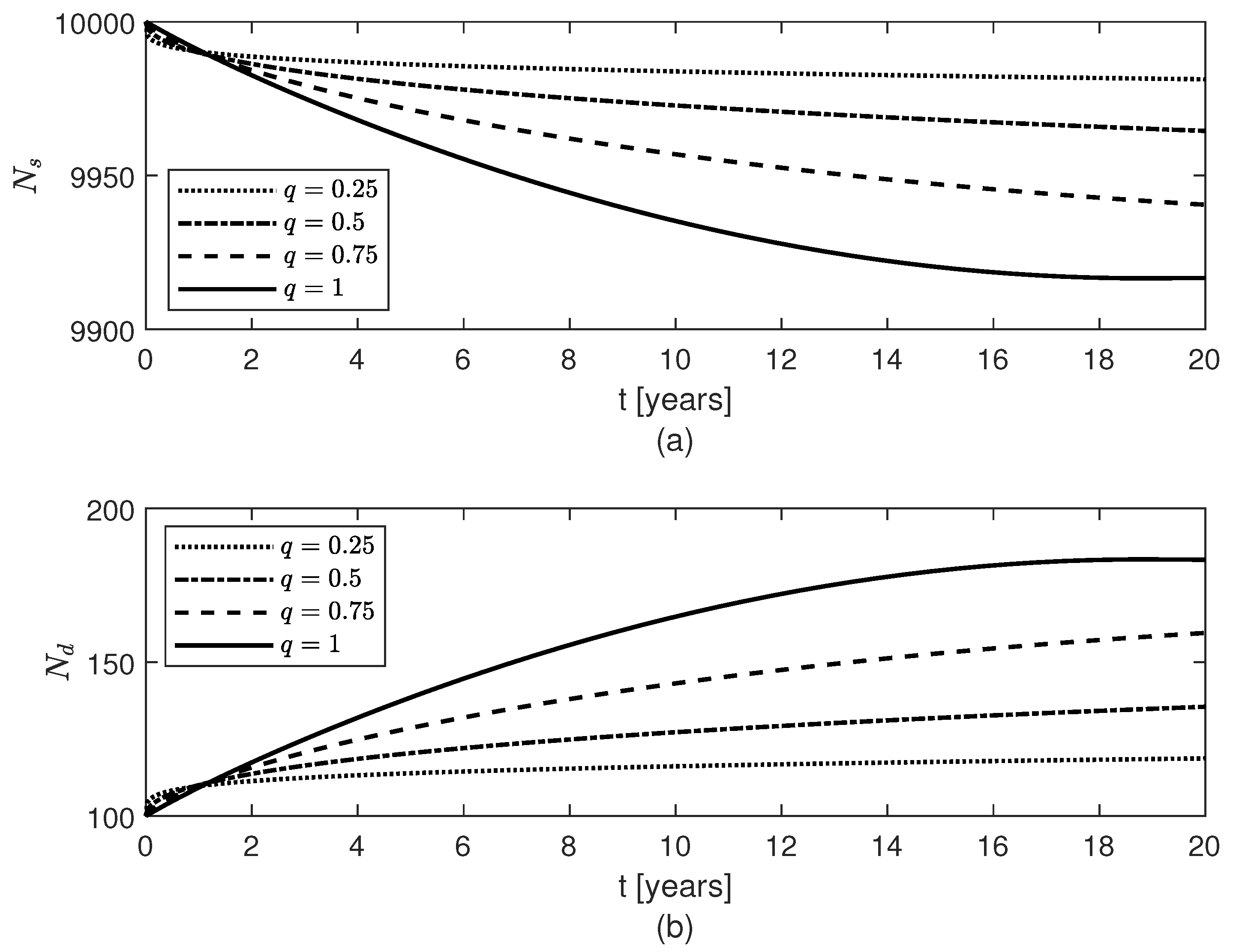
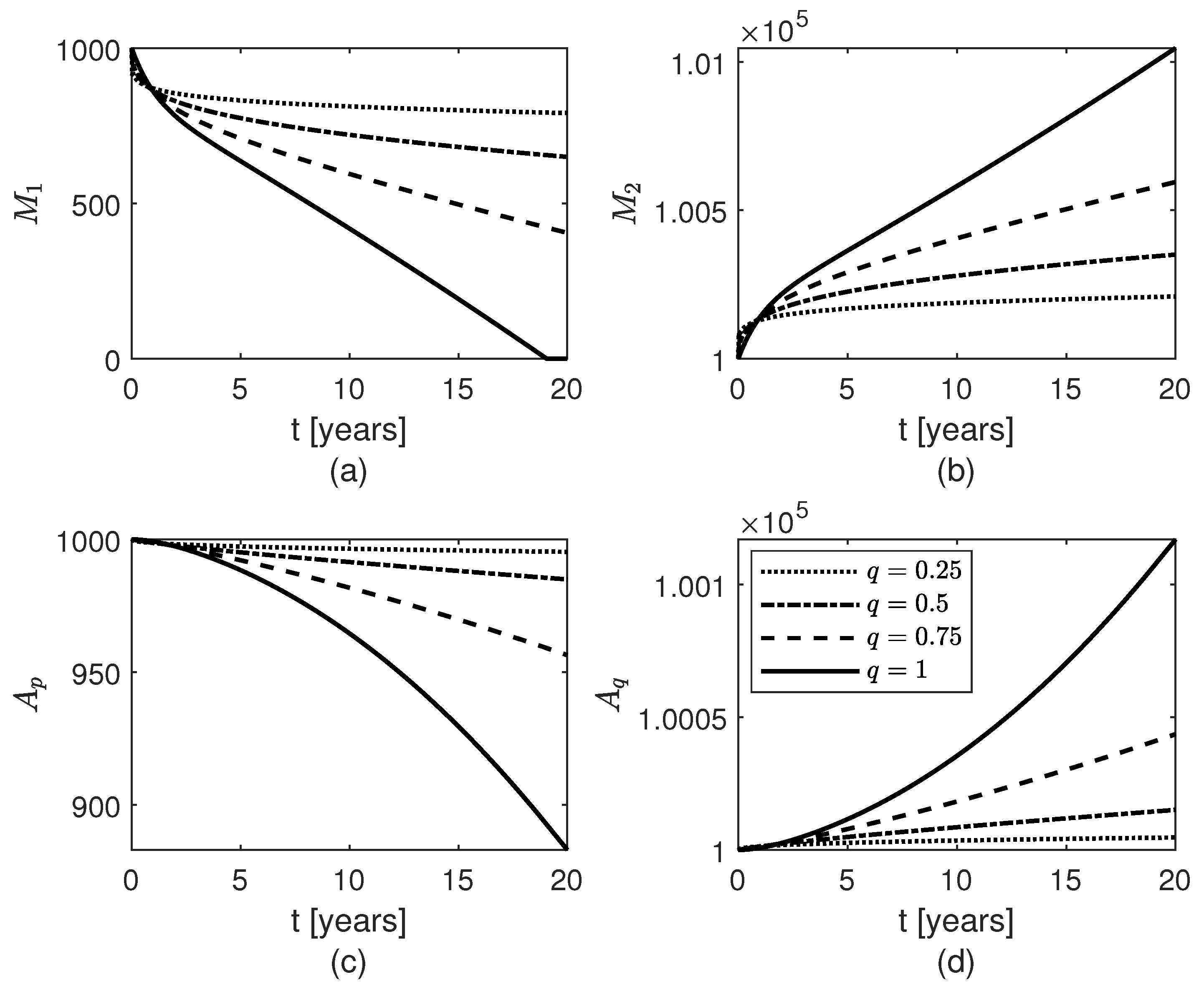
3. Results
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| Proliferative reactive astrocytes (in activated state) | |
| Quiescent astrocytes (in resting state) | |
| Amyloid- | |
| Activated microglia in pro-inflammatory state | |
| Activated microglia in anti-inflammatory state | |
| Surviving neurons | |
| Dead neurons |
References
- Pospich, S.; Raunser, S. The molecular basis of Alzheimer’s plaques. Science 2017, 358, 45–46. [Google Scholar] [CrossRef] [PubMed]
- Nebel, R.A.; Aggarwal, N.T.; Barnes, L.L.; Gallagher, A.; Goldstein, J.M.; Kantarci, K.; Mallampalli, M.P.; Mormino, E.C.; Scott, L.; Yu, W.H.; et al. Understanding the impact of sex and gender in Alzheimer’s disease: A call to action. Alzheimers Dement. 2018, 14, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Disease Facts and Figures. Available online: https://www.alz.org/alzheimers-dementia/facts-figures (accessed on 28 June 2022).
- Dementia. Available online: https://www.who.int/news-room/fact-sheets/detail/dementia (accessed on 28 June 2022).
- Wang, Y.; Mishra, A.; Brinton, R.D. Transitions in metabolic and immune systems from pre-menopause to post-menopause: Implications for age-associated neurodegenerative diseases. F1000Research 2020, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Zarate, S.; Stevnsner, T.; Gredilla, R. Role of estrogen and other sex hormones in brain aging. Neuroprotection and DNA repair. Front Aging Neurosci. 2017, 9, 430. [Google Scholar] [CrossRef]
- Mishra, A.; Brinton, R.D. Inflammation: Bridging age, menopause and APOEϵ4 genotype to Alzheimer’s disease. Front. Aging Neurosci. 2018, 10, 312. [Google Scholar] [CrossRef]
- Liu, C.C.; Liu, C.C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms and therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef]
- Dubal, D.B. Sex difference in Alzheimer’s disease: An updated, balanced and emerging perspective on differing vulnerabilities. Handb. Clin. Neurol. 2020, 175, 261–273. [Google Scholar] [CrossRef]
- Pontifex, M.G.; Martinsen, A.; Saleh, R.N.M.; Harden, G.; Tejera, N.; Muller, M.; Fox, C.; Vauzour, D.; Minihane, A.-M. APOE4 genotype exacerbates the impact of menopause on cognition and synaptic plasticity in APOE-TR mice. FASEB J. 2021, 35, e21583. [Google Scholar] [CrossRef]
- Corder, E.H.; Ghebremedhin, E.; Taylor, M.G.; Thal, D.R.; Ohm, T.G.; Braak, H. The biphasic relationship between regional brain senile plaque and neurofibrillary tangle distributions: Modification by age, sex, and APOE polymorphism. Ann. N. Y. Acad. Sci. 2004, 1019, 24–28. [Google Scholar] [CrossRef]
- Delage, C.I.; Simoncicova, E.; Tremblay, M.E. Microglial heterogeneity in aging and Alzheimer’s disease: Is sex relevant? J. Pharmacol. Sci. 2021, 146, 169–181. [Google Scholar] [CrossRef]
- Navakkode, S.; Gaunt, J.R.; Pavon, M.V.; Bansal, V.A.; Abraham, R.P.; Chong, Y.S.; Ch’ng, T.H.; Sajikumar, S. Sex-specific accelerated decay in time/activity-dependent plasticity and associative memory in an animal model of Alzheimer’s disease. Aging Cell. 2021, 20, e13502. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Park, J.; Choi, Y.K. The role of astrocytes in the central nervous system focused on BK channel and heme oxygenase metabolites: A review. Antioxidants 2019, 8, 121. [Google Scholar] [CrossRef]
- Wake, H.; Moorhouse, A.J.; Jinno, S.; Kohsaka, S.; Nabekura, J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J. Neurosci. 2009, 29, 3974–3980. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.-J.; Chung, W.-S. Phagocytic roles of glial cells in healthy and diseased brains. Biomol. Ther. 2018, 26, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Dubal, D.B.; Wise, P.M. Estrogen and neuroprotection: From clinical observations to molecular mechanisms. Dialogues Clin. Neurosci. 2002, 4, 149–161. [Google Scholar] [CrossRef]
- Brann, D.W.; Dhandapani, K.; Wakade, C.; Mahesh, V.B.; Khan, M.M. Neurotrophic and neuroprotective actions of estrogen: Basic mechanisms and clinical implications. Steroids 2007, 72, 381–405. [Google Scholar] [CrossRef]
- What Are the Symptoms of High Estrogen? Available online: https://www.medicalnewstoday.com/articles/323280#treatment (accessed on 30 June 2022).
- Lynch, A.M.; Murphy, K.J.; Deighan, B.F.; O’Reilly, J.A.; Gun’ko, Y.K.; Cowley, T.R.; Gonzalez-Reyes, R.E.; Lynch, M.A. The impact of glial activation in the aging brain. Aging Dis. 2010, 1, 262–278. [Google Scholar]
- Chun, H.; Marriott, I.; Lee, C.J.; Cho, H. Elucidating the interactive roles of glia in Alzheimer’s disease using established and newly developed experimental models. Front. Neurol. 2018, 9, 797. [Google Scholar] [CrossRef]
- Jiwaji, Z.; Tiwari, S.S.; Aviles-Reyes, R.X.; Hooley, M.; Hampton, D.; Torvell, M.; Johnson, D.A.; McQueen, J.; Baxter, P.; Sabari-Sankar, K.; et al. Reactive astrocytes acquire neuroprotective as well as deleterious signatures in response to Tau and Aβ pathology. Nat Commun. 2022, 13, 135. [Google Scholar] [CrossRef]
- Varnum, M.M.; Ikezu, T. The classification of microglial activation phenotypes on neurodegeneration and regeneration in Alzheimer’s disease brain. Arch. Immunol. Ther. Exp. 2012, 60, 251–266. [Google Scholar] [CrossRef]
- Klohs, J. An integrated view on vascular dysfunction in Alzheimer’s disease. Neurodegener. Dis. 2019, 19, 109–127. [Google Scholar] [CrossRef] [PubMed]
- Korte, N.; Nortley, R.; Attwell, D. Cerebral blood flow decrease as an early pathological mechanism in Alzheimer’s disease. Acta Neuropathol. 2020, 140, 793–810. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.L.; Pritchard, H.A.T.; Walsh, K.R.; Strangward, P.; White, C.; Hill-Eubanks, D.; Alakrawi, M.; Hennig, G.W.; Allan, S.M.; Nelson, M.T.; et al. Functionally linked potassium channel activity in cerebral endothelial and smooth muscle cells is compromised in Alzheimer’s disease. Pharmacology 2022, 119, e2204581119. [Google Scholar] [CrossRef]
- Wang, D.; Chen, F.; Han, Z.; Yin, Z.; Ge, X.; Lei, P. Relationship between amyloid-β deposition and blood-brain barrier dysfunction in Alzheimer’s disease. Front Cell Neurosci. 2021, 15, 695479. [Google Scholar] [CrossRef]
- Prins, N.; Scheltens, P. White matter hyperintensities, cognitive impairment and dementia: An update. Nat. Rev. Neurol. 2015, 11, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Lohner, V.; Pehlivan, G.; Sanroma, G.; Miloschewski, A.; Schirmer, M.D.; Stocker, T.; Reuter, M.; Breteler, M.M.B. The relation between sex, menopause, and white matter hyperintensities: The Rhineland study. Neurology 2022. [Google Scholar] [CrossRef]
- Alzheimer, A. Über einen eigenartigen schweren Erkrankungsprozeß der Hirnrincle. Neurol. Central. 1906, 25, 1134. [Google Scholar]
- Pallitto, M.M.; Murphy, R.M. A mathematical model of the kinetics of beta-amyloid fibril growth from the denatured state. Biophys J. 2001, 81, 1805–1822. [Google Scholar] [CrossRef]
- Helal, M.; Hingant, E.; Pujo-Menjouet, L.; Webb, G.F. Alzheimer’s disease: Analysis of a mathematical model incorporating the role of prions. J. Math. Biol. 2014, 69, 1207–1235. [Google Scholar] [CrossRef]
- Craft, D.L.; Wein, L.M.; Selkoe, D.J. A mathematical model of the impact of novel treatments on the A beta burden in the Alzheimer’s brain, CSF and plasma. Bull. Math. Biol. 2002, 64, 1011–1031. [Google Scholar] [CrossRef] [PubMed]
- Puri, I.K.; Li, L. Mathematical modeling for the pathogenesis of Alzheimer’s disease. PLoS ONE 2010, 5, e15176. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Friedman, A. Mathematical model on Alzheimer’s disease. BMC Syst. Biol. 2016, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Bertsch, M.; Franchi, B.; Marcello, N.; Tesi, M.C.; Tosin, A. Alzheimer’s disease: A mathematical model for onset and progression. Math. Med. Biol. 2017, 34, 193–214. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Melnik, R. Nonlocal models in the analysis of brain neurodegenerative protein dynamics with application to Alzheimer’s disease. Sci. Rep. 2022, 12, 7328. [Google Scholar] [CrossRef]
- Vosoughi, A.; Sadigh-Eteghad, S.; Ghorbani, M.; Shahmorad, S.; Farhoudi, M.; Rafi, M.A.; Omidi, Y. Mathematical models to shed light on amyloid-beta and tau protein dependent pathologies in Alzheimer’s disease. Neuroscience 2020, 424, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Holtzman, D.M. Biomarker modeling of Alzheimer’s disease. Neuron 2013, 80, 1347–1358. [Google Scholar] [CrossRef]
- Young, A.L.; Oxtoby, N.P.; Daga, P.; Cash, D.M.; Fox, N.C.; Ourselin, S.; Schott, J.M.; Alexander, D.C. A data-driven model of biomarker changes in sporadic Alzheimer’s disease. Brain J. Neurol. 2014, 137, 2564–2577. [Google Scholar] [CrossRef]
- Macdonald, A.; Pritchard, D. A mathematical model of Alzheimer’s disease and the Apoe gene. ASTIN Bull. 2000, 30, 69–110. [Google Scholar] [CrossRef]
- Hane, F.; Augusta, C.; Bai, O. A hierarchical Bayesian model to predict APOE4 genotype and the age of Alzheimer’s disease onset. PLoS ONE 2018, 13, e0200263. [Google Scholar] [CrossRef]
- Perez, C.; Ziburkus, J.; Ullah, G. Analyzing and modeling the dysfunction of inhibitory neurons in Alzheimer’s disease. PLoS ONE 2016, 11, e0168800. [Google Scholar] [CrossRef]
- Proctor, C.J.; Boche, D.; Gray, D.A.; Nicoll, J.A.R. Investigating interventions in Alzheimer’s disease with computer simulation models. PLoS ONE 2013, 8, e73631. [Google Scholar] [CrossRef]
- Hadjichrysanthou, C.; Ower, A.K.; de Wolf, F.; Anderson, R.M. Alzheimer’s disease neuroimaging initiative. The development of a stochastic mathematical model of Alzheimer’s disease to help improve the design of clinical trials of potential treatments. PLoS ONE 2018, 13, e0190615. [Google Scholar] [CrossRef]
- Samko, S.G.; Kilbas, A.A.; Marichev, O.I. Fractional Integrals and Derivatives; Gordon and Breach: Yverdon, Switzerland, 1993. [Google Scholar]
- Podlubny, I. Fractional Differential Equations; Mathematics in Science and Engineering 198; Academic Press: San Diego, CA, USA, 1999. [Google Scholar]
- Hilfer, R. Applications of Fractional Calculus in Physics; World Scientific: River Edge, NJ, USA, 2000. [Google Scholar]
- Oldham, K.B.; Spanier, J. The Fractional Calculus: Theory and Applications of Differentiation and Integration to Arbitrary Order; Dover: Mineola, NY, USA, 2006. [Google Scholar]
- Tarasov, V.E. Fractional Dynamics: Applications of Fractional Calculus to Dynamics of Particles, Fields and Media; Springer: Heidelberg, Germany, 2010. [Google Scholar]
- Mainardi, F. Fractional Calculus and Waves in Linear Viscoelasticity: An Introduction to Mathematical Models; Imperial College Press: London, UK, 2010. [Google Scholar]
- Baleanu, D.; Diethelm, K.; Scalas, E.; Trujillo, J.J. Fractional Calculus: Models and Numerical Methods; Series on Complexity, Nonlinearity and Chaos 3; World Scientific: Hackensack, NJ, USA, 2012. [Google Scholar]
- West, B.J. Fractional Calculus View of Complexity Tomorrow’S Science; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- West, B.J. Nature’s Patterns and the Fractional Calculus; Series Fractional Calculus in Applied Sciences and Engineering 2; De Gruyter: Berlin, Germany, 2017. [Google Scholar]
- Evangelista, L.R.; Lenzi, E.K. Fractional Diffusion Equations and Anomalous Diffusion; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Yang, X.-Y.; Yang, G.J. General Fractional Derivatives with Applications in Viscoelasticity; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Patnaik, S.; Hollkamp, J.P.; Semperlotti, F. Applications of variable-order fractional operators: A review. Proc. R. Soc. A 2020, 476, 20190498. [Google Scholar] [CrossRef] [PubMed]
- Wangersky, P.J. Lotka-Volterra population models. Ann. Rev. Ecol. Syst. 1978, 9, 189–218. [Google Scholar] [CrossRef]
- Cushing, J.M. Volterra integrodifferential equations in population dynamics. In Mathematics of Biology; Iannelli, M., Ed.; CIME Summer Schools 80; Springer: Berlin/Heidelberg, Germany, 2010; pp. 81–148. [Google Scholar]
- Ruan, S. Delay differential equations in single species dynamics. In Delay Differential Equations and Applications; Arino, O., Hbid, M.L., Ait Dads, E., Eds.; NATO science series. Series II, Mathematics, physics, and chemistry 205; Springer: Dordrecht, The Netherlands, 2006; pp. 477–517. [Google Scholar]
- Roberts, R.O.; Aakre, J.A.; Kremers, W.K.; Vassilaki, M.; Knopman, D.S.; Mielke, M.M.; Alhurani, R.; Geda, Y.E.; Machulda, M.M.; Coloma, P.; et al. Prevalence and outcomes of amyloid positivity among persons without dementia in a longitudinal, population-based detting. JAMA Neurol. 2018, 75, 970–979. [Google Scholar] [CrossRef]
- Sturchioa, A.; Dwivedic, A.K.; Youngd, C.B.; Malme, T.; Marsilia, L.; Sharmaa, J.S.; Mahajana, A.; Hilla, E.J.; Andaloussif, S.E.L.; Postond, K.L.; et al. High cerebrospinal amyloid-β 42 is associated with normal cognition in individuals with brain amyloidosis. E. Clin. Med. 2021, 38, 100988. [Google Scholar] [CrossRef]
- Song, Y.; Li, S.; Li, X.; Chen, X.; Wei, Z.; Liu, Q.; Cheng, Y. The effect of estrogen replacement therapy on Alzheimer’s disease and Parkinson’s disease in post-menopausal women: A meta-analysis. Front. Neurosci. 2020, 14, 157. [Google Scholar] [CrossRef]
- Vinogradova, Y.; Dening, T.; Hippisley-Cox, J.; Taylor, L.; Moore, M.; Coupland, C. Use of menopausal hormone therapy and risk of dementia: Nested case-control studies using QResearch and CPRD databases. BMJ 2021, 374, n2182. [Google Scholar] [CrossRef]
- Lorenzo, C.F.; Hartley, T.T. Variable order and distributed order fractional operators. Nonlinear Dyn. 2002, 29, 57–98. [Google Scholar] [CrossRef]
- Sun, H.; Chen, W.; Wei, H.; Chen, Y. A comparative study of constant-order and variable- order fractional models in characterizing memory property of systems. Eur. Phys. J. Spec. Top. 2011, 193, 185–192. [Google Scholar] [CrossRef]
- Ramirez, L.; Coimbra, C. On the selection and meaning of variable order operators for dynamic modeling. Int. J. Diff. Equ. 2010, 2010, 846107. [Google Scholar] [CrossRef]
- De Barros, L.C.; Lopes, M.M.; Pedro, F.S.; Esmi, E.; dos Santos, J.P.C.; Sanchez, D.E. The memory effect on fractional calculus: An application in the spread of COVID-19. Comput. Appl. Math. 2021, 40, 72. [Google Scholar] [CrossRef]
- When Does Menopause Start? Understanding the Symptoms by Age. Available online: https://www.healthpartners.com/blog/menopause-symptoms-by-age/ (accessed on 26 June 2022).
- Moghaddam, B.P.; Machado, J.A.T. Extended algorithms for approximating variable order fractional derivatives with applications. J. Sci. Comput. 2017, 71, 1351–1374. [Google Scholar] [CrossRef]
- Lawson, C.L.; Hanson, R.J. Solving Least Squares Problems; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1995; p. 161. [Google Scholar]
- Garrappa, R. Numerical solution of fractional differential equations: A survey and a software tutorial. Mathematics 2018, 6, 16. [Google Scholar] [CrossRef]
- Almeida, R.; Malinowska, A.B.; Odzijewicz, T. On systems of fractional differential equations with the ψ-Caputo derivative and their applications. Math. Meth. Appl. Sci. 2021, 44, 8026–8041. [Google Scholar] [CrossRef]
- Almeida, R.; Torres, D.F.M. An expansion formula with higher-order derivatives for fractional operators of variable order. Sci. World J. 2013, 2013, 915437. [Google Scholar] [CrossRef]
- Hale, J. Functional Differential Equations; Applied Mathematical Sciences 3; Springer: New York, NY, USA, 1971. [Google Scholar]
- Berezansky, L.; Braverman, E. On the existence of positive solutions for systems of differential equations with a distributed delay. Comput. Math. Appl. 2012, 63, 1256–1265. [Google Scholar] [CrossRef][Green Version]
- Berezansky, L.; Braverman, E. On nonoscillation and stability for systems of differential equations with a distributed delay. Automatica 2012, 48, 612–618. [Google Scholar] [CrossRef]
- Teschl, G. Ordinary Differential Equations and Dynamical Systems; Graduate Studies in Mathematics 140; American Mathematical Society: Providence, RI, USA, 2012. [Google Scholar]
- Pooseh, S.; Almeida, R.; Torres, D.F.M. A numerical scheme to solve fractional optimal control problems. Conf. Pap. Sci. 2013, 2013, 165298. [Google Scholar] [CrossRef]
- Rivers-Auty, J.; Mather, A.E.; Peters, R.; Lawrence, C.B.; Brough, D. Anti-inflammatories in Alzheimer’s disease-potential therapy or spurious correlate? Brain Commun. 2020, 2, fcaa109. [Google Scholar] [CrossRef]
- Arevalo-Rodriguez, I.; Smailagic, N.; Roqué-Figuls, M.; Ciapponi, A.; Sanchez-Perez, E.; Giannakou, A.; Pedraza, O.L.; Bonfill Cosp, X.; Cullum, S. Mini-Mental State Examination (MMSE) for the early detection of dementia in people with mild cognitive impairment (MCI). Cochrane Database Syst Rev. 2021, 7, CD010783. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Knopman, D.S.; Jagust, W.J.; Shaw, L.M.; Aisen, P.S.; Weiner, M.W.; Petersen, R.C.; Trojanowski, J.Q. Hypothetical model of dynamic biomarkers of the Alzheimer’s pathological cascade. Lancet Neurol. 2010, 9, 119–128. [Google Scholar] [CrossRef]
- Makin, S. The amyloid hypothesis on trial. Nature 2018, 559, S4–S7. [Google Scholar] [CrossRef] [PubMed]
- Furman, J.L.; Sama, D.M.; Gant, J.C.; Beckett, T.L.; Murphy, M.P.; Bachstetter, A.D.; Van Eldik, L.J.; Norris, C.M. Targeting astrocytes ameliorates neurologic changes in amouse model of Alzheimer’s disease. J Neurosci. 2012, 32, 16129–16140. [Google Scholar] [CrossRef]
- Chou, R.C.; Kane, M.; Ghimire, S.; Gautam, S.; Gui, J. Treatment for rheumatoid arthritis and risk of Alzheimer’s disease: A nested case-control analysis. CNS Drugs 2016, 30, 1111–1120. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Atkin, S.L.; Pirro, M.; Sahebkar, A. A review of the anti-inflammatory properties of antidiabetic agents providing protective effects against vascular complications in diabetes. J. Cell Physiol. 2019, 234, 8286–8294. [Google Scholar] [CrossRef]
- Munoz-Jimenez, M.; Zaarkti, A.; Garcia-Arnes, J.A.; Garcia-Casares, N. Antidiabetic drugs in Alzheimer’s disease and mild cognitive impairment: A systematic review. Dement. Geriatr. Cogn. Disord. 2020, 49, 423–434. [Google Scholar] [CrossRef]
- Chowen, J.A.; Garcia-Segura, L.M. Role of glial cells in the generation of sex differences in neurodegenerative diseases and brain aging. Mech. Ageing Dev. 2021, 196, 111473. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drapaca, C.S. A Mathematical Investigation of Sex Differences in Alzheimer’s Disease. Fractal Fract. 2022, 6, 457. https://doi.org/10.3390/fractalfract6080457
Drapaca CS. A Mathematical Investigation of Sex Differences in Alzheimer’s Disease. Fractal and Fractional. 2022; 6(8):457. https://doi.org/10.3390/fractalfract6080457
Chicago/Turabian StyleDrapaca, Corina S. 2022. "A Mathematical Investigation of Sex Differences in Alzheimer’s Disease" Fractal and Fractional 6, no. 8: 457. https://doi.org/10.3390/fractalfract6080457
APA StyleDrapaca, C. S. (2022). A Mathematical Investigation of Sex Differences in Alzheimer’s Disease. Fractal and Fractional, 6(8), 457. https://doi.org/10.3390/fractalfract6080457