
Vitamin A in Fish Well-Being: Integrating Immune Strength, Antioxidant Capacity and Growth


Abstract
1. Introduction
- Role of vitamin A in fish immunology.
- Vitamin A status and oxidative stress.
- Impact of vitamin A on fish growth.
- Integrative discussion.
- Future directions.
2. Role of Vitamin A in Fish Immunology
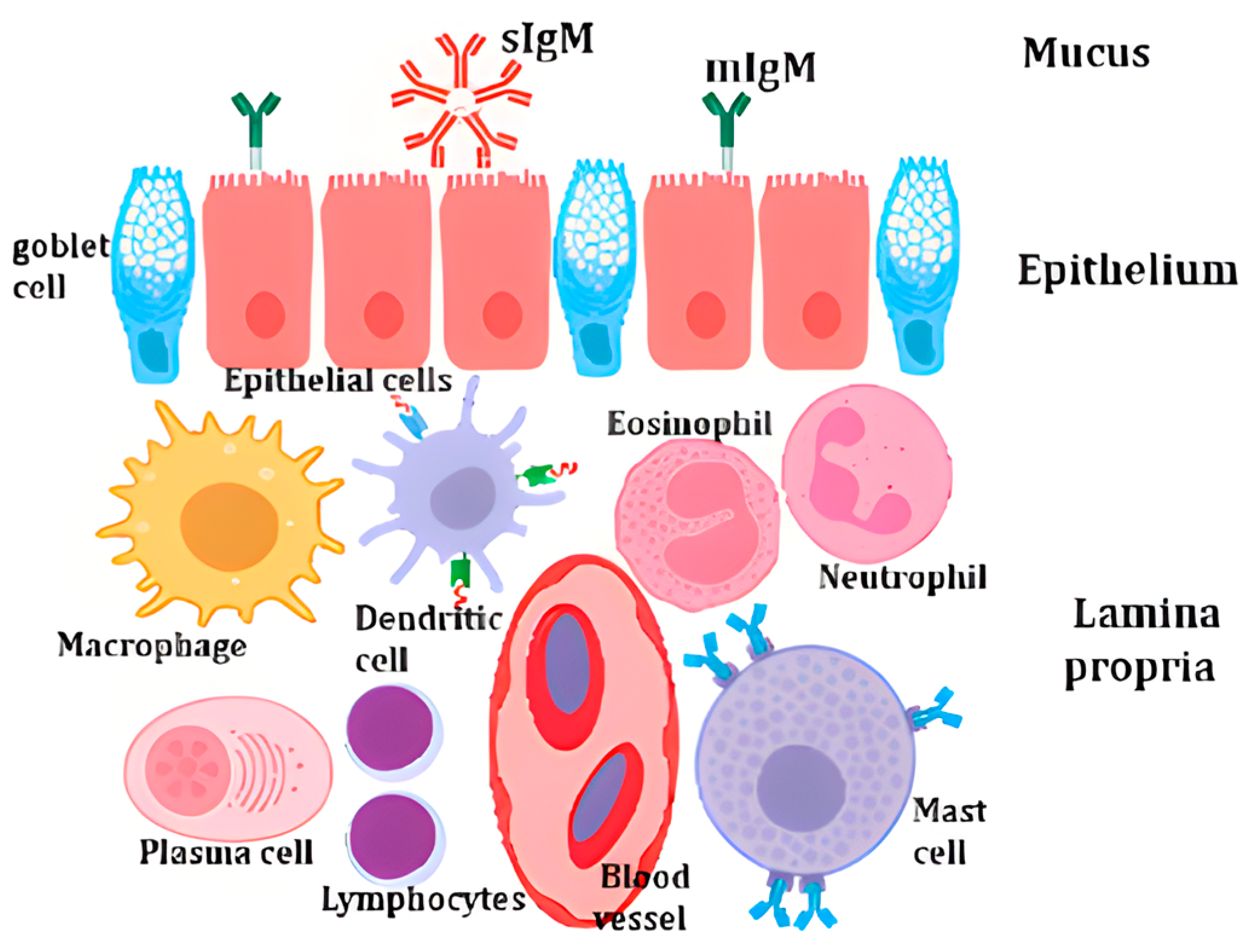
2.1. Overview of Fish Immune System
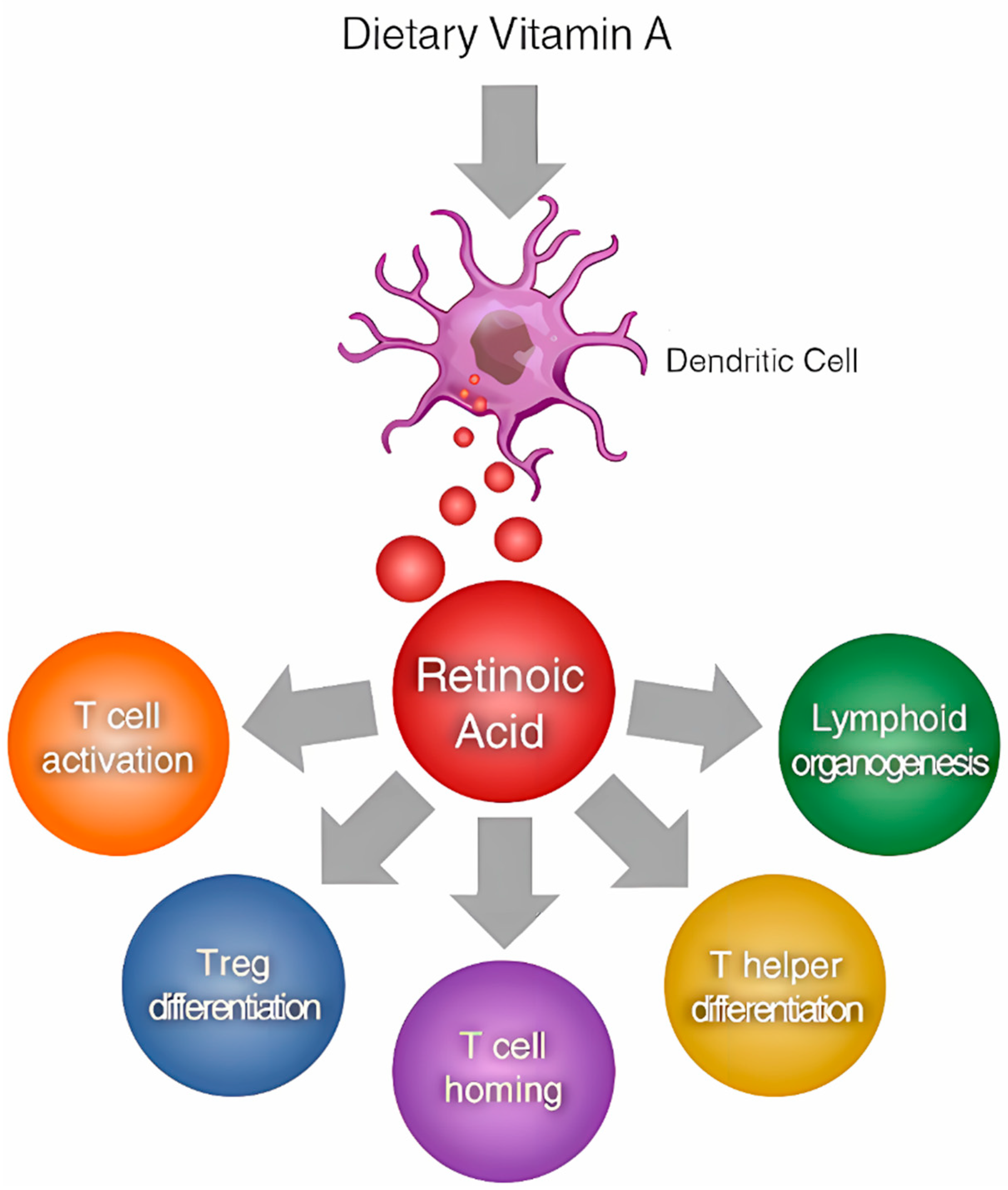
2.2. Vitamin A and Immune Function
2.3. Vitamin A Modulation of Immune Responses
2.4. Summary of Key Studies and Findings
3. Vitamin A Status and Oxidative Stress
3.1. Oxidative Stress in Fish
3.2. Importance of Vitamin A in Combating Oxidative Stress
- Scavenging Free Radicals: Retinol exhibits direct antioxidant properties owing to its hydrophobic polyene chains, allowing it to quench singlet oxygen and neutralize radicals [10,12]. Despite being prone to auto-oxidation in high-oxygen environments, retinol remains effective under physiological oxygen levels [5,92,93]. Palace et al. [94] detailed how retinol acts as a chain-breaking antioxidant by intercepting peroxyl radicals, thereby halting lipid peroxidation and the formation of hydroperoxides. It efficiently scavenges peroxyl radicals in various lipid models, including liposomes mimicking cell membranes.
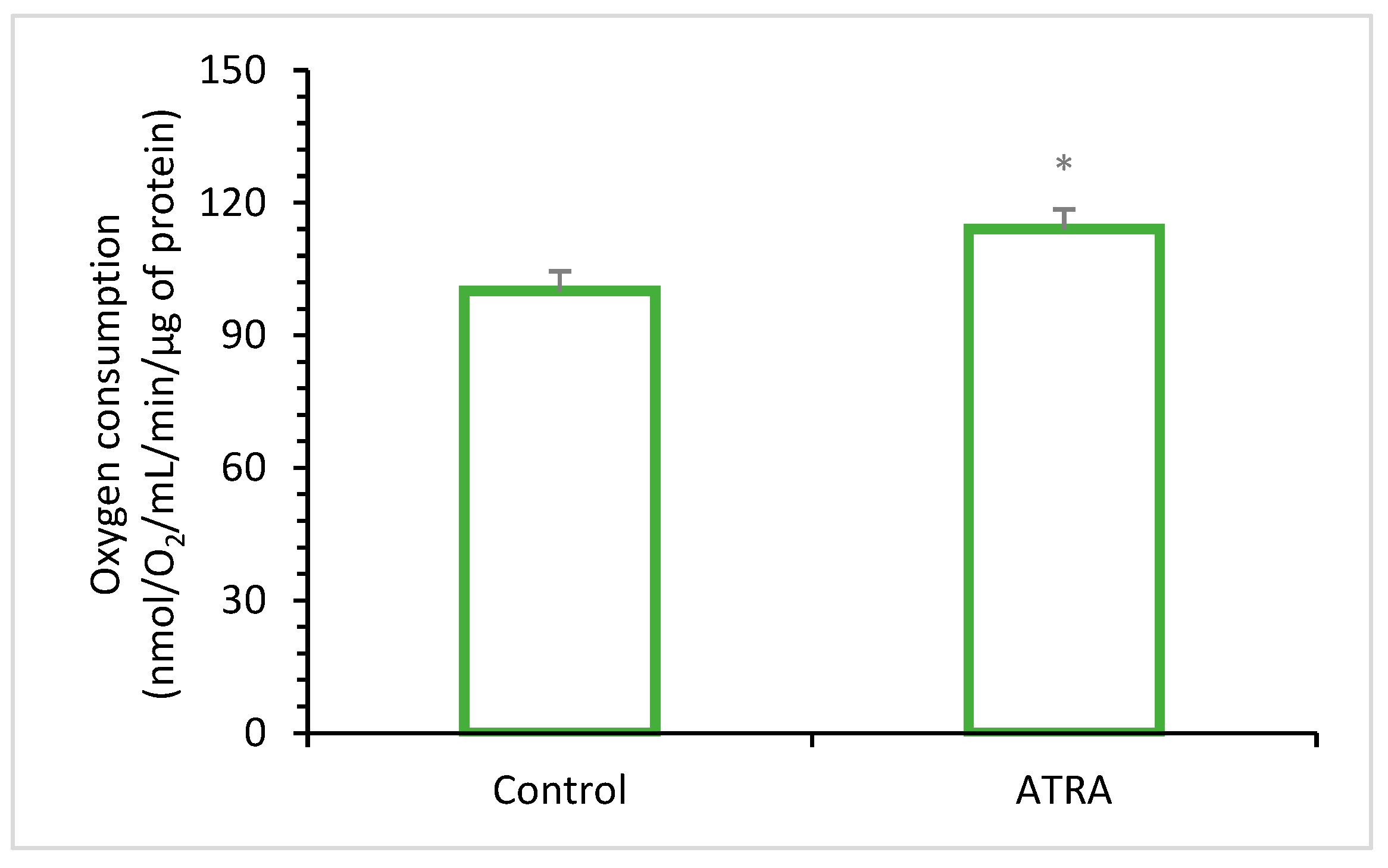
- Enhancing Antioxidant Enzyme Activity: ATRA, a metabolite of vitamin A, serves as a potent transcriptional regulator that influences the expression of genes involved in antioxidant processes [95,96] (Figure 3). Specifically, ATRA has been shown to upregulate the expression and activity of key antioxidant enzymes such as SOD, CAT, and GPx [5,90,97,98].
- 3.
- Inducing Autophagy: ATRA has been found to induce autophagy, a critical cellular mechanism that helps cells cope with nutrient scarcity or oxidative stress, as observed in fish and other organisms [100,101]. Research by Rajawat et al. [100] suggests that ATRA promotes the formation of autophagosomes via a pathway that does not involve conventional nuclear retinoid receptors. Specifically, ATRA triggers the relocation of the cation-independent mannose-6-phosphate receptor from the trans-Golgi region to maturing autophagosomes, leading to their acidification. This autophagic process plays a crucial role in managing redox balance and maintaining cellular stability [102].
3.3. Case Studies: Effects of Vitamin A Supplementation on Antioxidant Capacity
4. Impact of Vitamin A on Fish Growth
4.1. Growth Metrics in Fish
4.2. Vitamin A and Growth Regulation
4.3. Effects of Vitamin A Supplementation on Growth Rates and Body Composition
5. Integrative Discussion
5.1. Interconnectedness of Immunology, Antioxidant Capacity, and Growth
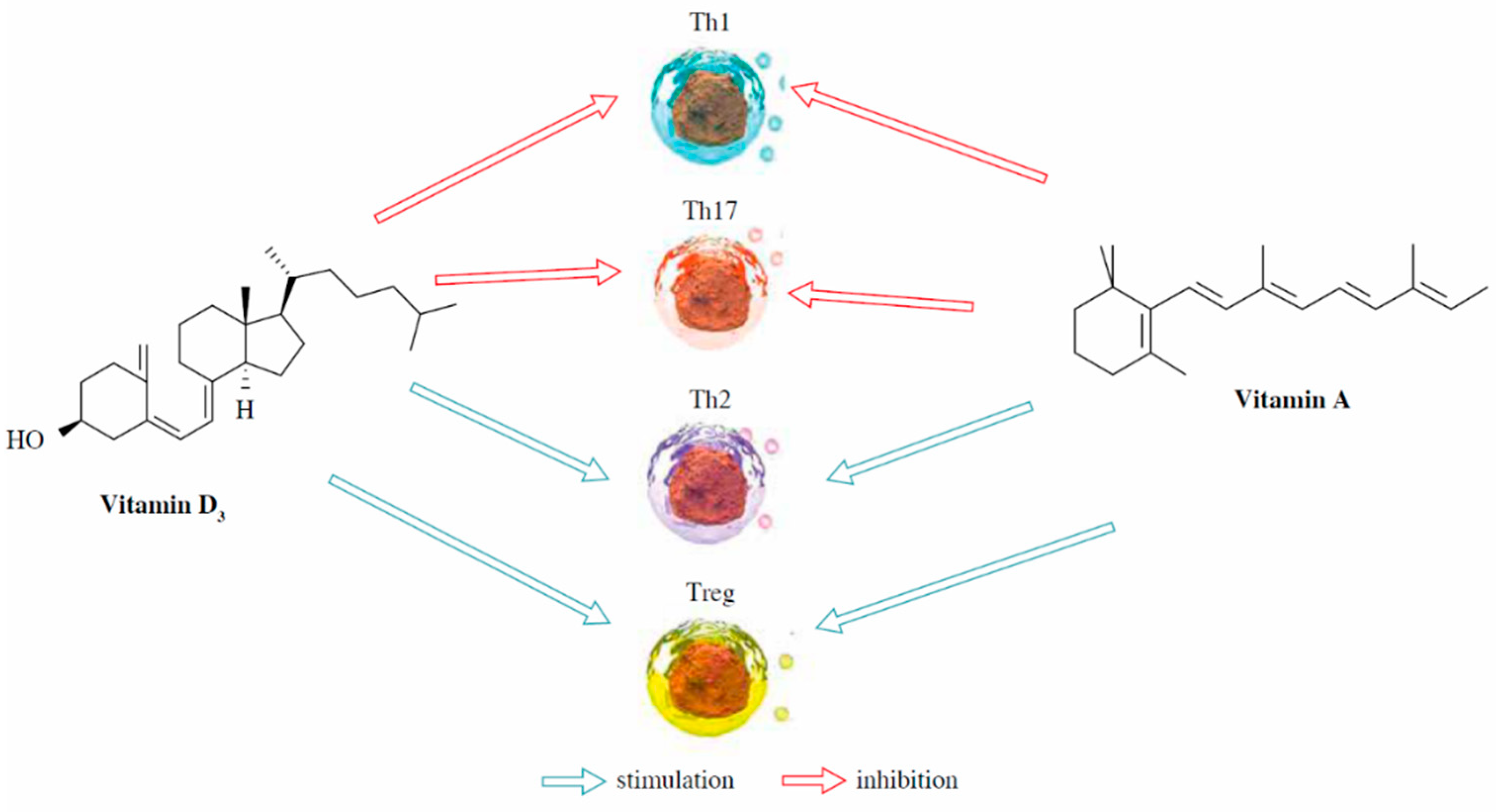
5.2. Interaction of Vitamin A with Other Vitamins
5.3. Recommendations for Integrated Management Practices in Aquaculture
- Nutritionally Balanced Diets: Formulating diets that contain sufficient vitamin A and account for its interactions with vitamins E, D, and C is essential. Using stable and bioaccessible forms of vitamin A in high-quality feeds enhances bioavailability, supporting immune function, antioxidant capacity, and growth [151,152].
- Water Quality Management: Maintaining optimal water conditions is fundamental for nutrient absorption and metabolic processes [153,154]. The regular monitoring and adjustment of water parameters prevent conditions that could impair vitamin A metabolism, ensuring efficient utilization and maximizing fish health [155,156].
- Monitoring and Adjustment: The continuous monitoring of fish health indicators and environmental parameters enables proactive adjustments in management [163,164]. Biomarker assessments of vitamin A levels and immune function offer insights into the effectiveness of dietary strategies, facilitating timely interventions to optimize fish health.
- Integrated Health Management: Integrating nutrition, water quality management, and disease prevention measures ensures a comprehensive approach to maximizing the benefits of vitamin A. Strategies such as vaccination programs, biosecurity measures, and probiotic use enhance immune competence and overall resilience in fish populations [165,166].
6. Future Directions
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 4-HNE | 4-hydroxynonenal |
| APCs | Antigen-presenting cells |
| ATRA | All-trans-retinoic acid |
| CAT | Catalase |
| FCR | Feed conversion ratio |
| FR | Feeding rate |
| GPx | Glutathione peroxidase |
| IFN-γ | Interferon-gamma |
| IL | Interleukin |
| ILCs | Innate lymphoid cells |
| IU | International Unit |
| Keap1 | Kelch-like ECH-associated protein 1 |
| MDA | Malondialdehyde |
| MPO | Myeloperoxidase |
| Nrf 2 | Nuclear factor-erythroid 2 p45-related factor 2 |
| RAREs | Retinoic acid response elements |
| RARs | Retinoic acid receptors |
| ROS | Reactive oxygen species |
| RXRs | Retinoid X receptors |
| SGR | Specific growth rate |
| SOD | Superoxide dismutase |
| SR | Survival rate |
| Tregs | T cells |
References
- Clugston, R.D.; Blaner, W.S. Vitamin A (retinoid) metabolism and actions: What we know and what we need to know about amphibians. Zoo Biol. 2014, 33, 527–535. [Google Scholar] [CrossRef]
- Hernandez, L.H.; Hardy, R.W. Vitamin A functions and requirements in fish. Aquac Res. 2020, 51, 3061–3071. [Google Scholar] [CrossRef]
- Martin, S.A.M.; Krol, E. Nutrigenomics and immune function in fish: New insights from omics technologies. Dev. Comp. Immunol. 2017, 75, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.; Ahmed, I.; Wani, G.B. Effect of Supplementation of Vitamin A on Growth, Haemato-Biochemical Composition, and Antioxidant Ability in Cyprinus carpio var. communis. Aquac. Nutr. 2022, 2022, 8446092. [Google Scholar] [CrossRef] [PubMed]
- Shastak, Y.; Gordillo, A.; Pelletier, W. The relationship between vitamin A status and oxidative stress in animal production. J. Appl. Anim. Res. 2023, 51, 546–553. [Google Scholar] [CrossRef]
- Samrani, L.M.M.; Pennings, J.L.A.; Hallmark, N.; Bars, R.; Tinwell, H.; Pallardy, M.; Piersma, A.H. Dynamic regulation of gene expression and morphogenesis in the zebrafish embryo test after exposure to all-trans retinoic acid. Reprod. Toxicol. 2023, 115, 8–16. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. The role of vitamin A in non-ruminant immunology. Front. Anim. Sci. 2023, 4, 1197802. [Google Scholar] [CrossRef]
- Wold, H.L.; Wake, K.; Higashi, N.; Wang, D.; Kojima, N.; Imai, K.; Blomhoff, R.; Senoo, H. Vitamin A distribution and content in tissues of the lamprey, Lampetra japonica. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 276, 134–142. [Google Scholar] [CrossRef]
- Shidoji, Y.; Muto, Y. Vitamin A transport in plasma of the non-mammalian vertebrates: Isolation and partial characterization of piscine retinol-binding protein. J. Lipid Res. 1977, 18, 679–691. [Google Scholar] [CrossRef]
- Landete, J.M. Dietary intake of natural antioxidants: Vitamins and polyphenols. Crit. Rev. Food Sci. Nutr. 2013, 53, 706–721. [Google Scholar] [CrossRef]
- Roos, N.; Chamnan, C.; Loeung, D.; Jakobsen, J.; Thilsted, S.H. Freshwater fish as a dietary source of vitamin A in Cambodia. Food Chem. 2007, 103, 1104–1111. [Google Scholar] [CrossRef]
- Alsop, D.; Van Der Kraak, G.J.; Brown, S.B.; Eales, J.G. The biology and toxicology of retinoids in fish. In Biocheminstry and Molecular Biology of Fishes; Mommsen, T.P., Moon, T.W., Eds.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2005; Volume 6 Environmental toxicology; pp. 413–428. [Google Scholar] [CrossRef]
- Wagner, H.J. Retinal structure of fishes. In The Visual System of Fish; Douglas, R., Djamgoz, M., Eds.; Springer: Dordrecht, The Netherlands, 1990. [Google Scholar] [CrossRef]
- Jami, R.; Mérour, E.; Lamoureux, A.; Bernard, J.; Millet, J.K.; Biacchesi, S. Deciphering the Fine-Tuning of the Retinoic Acid-Inducible Gene-I Pathway in Teleost Fish and Beyond. Front. Immunol. 2021, 12, 679242. [Google Scholar] [CrossRef]
- Jiang, W.D.; Zhang, L.; Feng, L.; Wu, P.; Liu, Y.; Kuang, S.Y.; Li, S.W.; Tang, L.; Mi, H.F.; Zhang, L.; et al. New Insight on the Immune Modulation and Physical Barrier Protection Caused by Vitamin A in Fish Gills Infected with Flavobacterium columnare. Front Immunol. 2022, 13, 833455. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zhang, L.; Jiang, W.; Liu, Y.; Jiang, J.; Kuang, S.; Li, S.; Tang, L.; Tang, W.; Zhou, X.; et al. Dietary Vitamin A Improved the Flesh Quality of Grass Carp (Ctenopharyngodon idella) in Relation to the Enhanced Antioxidant Capacity through Nrf2/Keap 1a Signaling Pathway. Antioxidants 2022, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.W.Y.; Lai, R.W.S.; Zhou, G.J.; Leung, K.M.Y. Concentration-response of six marine species to all-trans-retinoic acid and its ecological risk to the marine environment. Ecotoxicol. Environ. Saf. 2022, 235, 113455. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, M.R.; Wingert, R.A. Zebrafish as a Model to Study Retinoic Acid Signaling in Development and Disease. Biomedicines 2023, 11, 1180. [Google Scholar] [CrossRef]
- Guilland, J.-C. Les interactions entre les vitamines A, D, E et K: Synergie et/ou competition. OCL 2011, 18, 59–67. [Google Scholar] [CrossRef]
- Kane, M.A. Analysis, occurrence, and function of 9-cis-retinoic acid. Biochim. Biophys. Acta 2012, 1821, 10–20. [Google Scholar] [CrossRef]
- Buck, J.; Grun, F.; Derguini, F.; Chen, Y.; Kimura, S.; Noy, N.; Hammerling, U. Anhydroretinol: A naturally occur-ring inhibitor of lymphocyte physiology. J. Exp. Med. 1993, 178, 675–680. [Google Scholar] [CrossRef]
- Blomhoff, R.; Blomhoff, H.K. Overview of retinoid metabolism and function. Overview of retinoid metabolism and function. J. Neurobiol. 2006, 66, 606–630. [Google Scholar] [CrossRef]
- Fernández, I.; Gisbert, E. The effect of vitamin a on flatfish development and skeletogenesis: A review. Aquaculture 2011, 315, 34–48. [Google Scholar] [CrossRef]
- Hernandez de-Dios, M.A.; Tovar-Ramírez, D.; Maldonado García, D.; Galaviz-Espinoza, M.A.; Spanopoulos Zarco, M.; Maldonado-García, M.C. Functional Additives as a Boost to Reproductive Performance in Marine Fish: A Review. Fishes 2022, 7, 262. [Google Scholar] [CrossRef]
- Zhang, L.; Feng, L.; Jiang, W.-D.; Liu, Y.; Wu, P.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Zhou, X.-Q. Vitamin A deficiency suppresses fish immune function with differences in different intestinal segments: The role of transcriptional factor NF-κB and p38 mitogen-activated protein kinase signalling pathways. Br. J. Nutr. 2017, 117, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Hilton, J.W. Hypervitaminosis A in rainbow trout (Salmo gairdneri): Toxicity signs and maximum tolerable level. J. Nutr. 1983, 113, 1737–1745. [Google Scholar] [CrossRef] [PubMed]
- Furuita, H.; Tanaka, H.; Yamamoto, T.; Shiraishi, M.; Takeuchi, T. Effects of high dose of vitamin A on reproduction and egg quality of Japanese flounder Paralichthys olivaceus. Fish. Sci. 2001, 67, 606–613. [Google Scholar] [CrossRef]
- NASEM (National Academies of Sciences, Engineering, and Medicine). Nutrient Requirements of Fish and Shrimp; National Academic Science: Washington, DC, USA, 2011. [Google Scholar]
- Taveekijakarn, P.; Miyazaki, T.; Matsumoto, M.; Arai, S. Vitamin A Deficiency in Cherry Salmon. J. Aquat. Anim. Health 1994, 6, 251–259. [Google Scholar] [CrossRef]
- Saleh, G.; Eleraku, W.; Gropp, J.M. A short note on the effects of vitamin A hypervitaminosis on health and growth of Tilapia nilotica (Oreochromis niloticus). J. Appl. Ichthyol. 1995, 11, 382–385. [Google Scholar] [CrossRef]
- Dedi, J.; Takeuchi, T.; Seikai, T.; Watanabe, T.; Hosoya, K. Hyper vitaminosis A during vertebral morphogenesis in larval Japanese flounder. Fish. Sci. 1997, 63, 466–473. [Google Scholar] [CrossRef]
- Ørnsrud, R.; Graff, I.E.; Høie, S.; Totland, G.K.; Hemre, G.-I. Hypervitaminosis A in first-feeding fry of the Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2002, 8, 7–13. [Google Scholar] [CrossRef]
- Campeche, D.F.B.; Catharino, R.R.; Godoy, H.T.; Cyrino, J.E.P. Vitamin A in diets for Nile tilapia. Sci. Agric. 2009, 66, 751–756. [Google Scholar] [CrossRef]
- Dominguez, D.; Montero, D.; Zamorano, M.J.; Castro, P.L.; da Silva, J.; Fontanillas, R.; Izquierdo, M. High Levels of Vitamin A in Plant-Based Diets for Gilthead Seabream (Sparus aurata) Juveniles, Effects on Growth, Skeletal Anomalies, Bone Molecular Markers, and Histological Morphology. Aquac. Nutr. 2023, 2023, 5788432. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, P.; Jameel, F.; Rani, D.; Serajuddin, M. Deficiency of protein, fat and vitamins in freshwater catfish, Clarias batrachus: Morphological symptoms and impact on growth performance. Borneo J. Mar. Sci. Aquac. 2019, 3, 9–12. [Google Scholar]
- Yang, Q.-H.; Zhou, X.-Q.; Jiang, J.; Liu, Y. Effect of dietary vitamin A deficiency on growth performance, feed utilization and immune responses of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Res. 2008, 39, 902–906. [Google Scholar] [CrossRef]
- Ma, J.; Liu, Y.; Li, Y.; Gu, J.; Liu, J.; Tang, J.; Wang, J.; Ryffel, B.; Shen, Y.; Brand, D.; et al. Differential role of all-trans retinoic acid in promoting the development of CD4+ and CD8+ regulatory T cells. J. Leukoc. Biol. 2014, 95, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Mariu, A.; Chatha, A.M.M.; Naz, S.; Khan, M.F.; Safdar, W.; Ashraf, I. Effect of Temperature, pH, Salinity and Dissolved Oxygen on Fishes. J. Zool. Syst. 2023, 1, 1–12. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Delving into Vitamin A Supplementation in Poultry Nutrition: Current Knowledge, Functional Effects, and Practical Implications. Worlds Poult. Sci. J. 2023, 79, 109–131. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organisation of the United Nations). Record Fisheries and Aquaculture Production Makes Critical Contribution to Global Food Security. 2022. Available online: https://www.fao.org/newsroom/detail/record-fisheries-aquaculture-production-contributes-food-security-290622/en (accessed on 6 July 2024).
- Palace, V.P.; Werner, J. Vitamins A and E in the maternal diet influence egg quality and early life stage development in fish: A review. Sci. Mar. 2006, 70, 41–57. [Google Scholar] [CrossRef]
- Firdaus-Nawi, M.; Zamri-Saad, M. Major Components of Fish Immunity: A Review. Pertanika J. Trop. Agric. Sci. 2016, 39, 393–420. [Google Scholar]
- Mokhtar, D.M.; Zaccone, G.; Alesci, A.; Kuciel, M.; Hussein, M.T.; Sayed, R.K.A. Main Components of Fish Immunity: An Overview of the Fish Immune System. Fishes 2023, 8, 93. [Google Scholar] [CrossRef]
- Smith, N.C.; Rise, M.L.; Christian, S.L. A Comparison of the Innate and Adaptive Immune Systems in Cartilaginous Fish, Ray-Finned Fish, and Lobe-Finned Fish. Front. Immunol. 2019, 10, 2292. [Google Scholar] [CrossRef]
- Semple, S.L.; Dixon, B. Salmonid Antibacterial Immunity: An Aquaculture Perspective. Biology 2020, 9, 331. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, F.A.; Oettgen, H.C. Adaptive immunity. J. Allergy Clin. Immunol. 2010, 125 (Suppl. S2), S33–S40. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Xu, H.; Yu, Y.; Xu, Z. Regulatory roles of cytokines in T and B lymphocytes-mediated immunity in teleost fish. Dev. Comp. Immunol. 2023, 144, 104621. [Google Scholar] [CrossRef]
- Magnadottir, B. Immunological Control of Fish Diseases. Mar. Biotechnol. 2010, 12, 361–379. [Google Scholar] [CrossRef]
- Barbosa, L.M.G.; Moraes, G.; Anibal, F.F.; Machado, C.M.M. Effect of environmental thermal fluctuations on innate immune responses in pacu Piaractus mesopotamicus juveniles. Aquac. Rep. 2020, 17, 100303. [Google Scholar] [CrossRef]
- Magadan, S.; Sunyer, O.J.; Boudinot, P. Unique Features of Fish Immune Repertoires: Particularities of Adaptive Immunity within the Largest Group of Vertebrates. Results Probl. Cell Differ. 2015, 57, 235–264. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.; Banu, H.; Prakash, A.; Tripathi, G. Immune System of Fish: An Evolutionary Perspective. In Antimicrobial Immune Response; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Green, H.N.; Mellanby, E. Vitamin A as an anti-infective agent. Brit. Med. J. 1928, 2, 691. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Vitamin A supply in swine production: A review of current science and practical considerations. Appl. Anim. Sci. 2023, 39, 289–305. [Google Scholar] [CrossRef]
- Vijayaram, S.; Ringø, E.; Zuorro, A.; van Doan, H.; Sun, Y. Beneficial roles of nutrients as immunostimulants in aquaculture: A review. Aquac. Fish. 2023, 9, 707–720. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Pet Wellness and Vitamin A: A Narrative Overview. Animals 2024, 14, 1000. [Google Scholar] [CrossRef]
- Thompson, I.; Fletcher, T.C.; Houlihan, D.F.; Secombes, C.J. The effect of dietary vitamin A on the immunocompetence of Atlantic salmon (Salmo salar L.). Fish Physiol. Biochem. 1994, 12, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Gomez, D.; Sunyer, J.O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish. Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of Vitamin A in the Immune System. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [PubMed]
- Speirs, Z.C.; Loynes, C.A.; Mathiessen, H.; Elks, P.M.; Renshaw, S.A.; Jørgensen, L.V.G. What can we learn about fish neutrophil and macrophage response to immune challenge from studies in zebrafish. Fish Shellfish. Immunol. 2024, 148, 109490. [Google Scholar] [CrossRef]
- Jiang, W.-D.; Zhang, L.; Feng, L.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Inconsistently impairment of immune function and structural integrity of head kidney and spleen by vitamin A deficiency in grass carp (Ctenopharyngodon idella). Fish Shellfish. Immunol. 2020, 99, 243–256. [Google Scholar] [CrossRef]
- Guimaraes, I.G.; Lim, C.; Yildirim-Aksoy, M.; Li, M.H.; Klesius, P.H. Effects of dietary levels of vitamin A on growth, hematology, immune response and resistance of Nile tilapia (Oreochromis niloticus) to Streptococcus iniae. Anim. Feed. Sci. Technol. 2014, 188, 126–136. [Google Scholar] [CrossRef]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin Effects on the Immune System: Vitamins A and D Take Centre Stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef]
- Nagpal, I.; Wei, L.N. All-trans Retinoic Acid as a Versatile Cytosolic Signal Modulator Mediated by CRABP1. Int. J. Mol. Sci. 2019, 20, 3610. [Google Scholar] [CrossRef]
- White, J.A.; Boffa, M.B.; Jones, B.; Petkovich, M. A zebrafish retinoic acid receptor expressed in the regenerating caudal fin. Development 1994, 120, 1861–1872. [Google Scholar] [CrossRef]
- Balmer, J.E.; Blomhoff, R. A robust characterization of retinoic acid response elements based on a comparison of sites in three species. J. Steroid Biochem. Mol. Biol. 2005, 96, 347–354. [Google Scholar] [CrossRef]
- Fernández, I.; Tiago, D.M.; Laizé, V.; Leonor Cancela, M.; Gisbert, E. Retinoic acid differentially affects in vitro proliferation, differentiation and mineralization of two fish bone-derived cell lines: Different gene expression of nuclear receptors and ECM proteins. J. Steroid Biochem. Mol. Biol. 2014, 140, 34–43. [Google Scholar] [CrossRef]
- Lu, L.; Ma, J.; Li, Z.; Lan, Q.; Chen, M.; Liu, Y.; Xia, Z.; Wang, J.; Han, Y.; Shi, W.; et al. All-Trans Retinoic Acid Promotes TGF-β-Induced Tregs via Histone Modification but Not DNA Demethylation on Foxp3 Gene Locus. PLoS ONE 2011, 6, e24590. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-M.; Wang, K.-P.; Ma, J.; Zheng, S.G. The role of all-trans retinoic acid in the biology of Foxp3+ regulatory T cells. Cell. Mol. Immunol. 2015, 12, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Indrevær, R.L.; Moskaug, J.; Paur, I.; Bøhn, S.K.; Jørgensen, S.F.; Blomhoff, R.; Aukrust, P.; Fevang, B.; Blomhoff, H.K. IRF4 Is a Critical Gene in Retinoic Acid-Mediated Plasma Cell Formation and Is Deregulated in Common Variable Immunodeficiency-Derived B Cells. J. Immunol. 2015, 195, 2601–2611. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Fu, S.; Yin, X.; Guo, Z.; Wang, A.; Ye, J. Long-Lived Plasma Cells Secrete High-Affinity Antibodies Responding to a T-Dependent Immunization in a Teleost Fish. Front. Immunol. 2019, 10, 2324. [Google Scholar] [CrossRef]
- Bono, M.R.; Tejon, G.; Flores-Santibañez, F.; Fernandez, D.; Rosemblatt, M.; Sauma, D. Retinoic Acid as a Modulator of T Cell Immunity. Nutrients 2016, 8, 349. [Google Scholar] [CrossRef]
- Duriancik, D.M.; Lackey, D.E.; Hoag, K.A. Vitamin A as a regulator of antigen presenting cells. J. Nutr. 2010, 140, 1395–1399. [Google Scholar] [CrossRef]
- Iliev, D.B.; Thim, H.; Lagos, L.; Olsen, R.; Jørgensen, J.B. Homing of Antigen-Presenting Cells in Head Kidney and Spleen—Salmon Head Kidney Hosts Diverse APC Types. Front. Immunol. 2013, 4, 137. [Google Scholar] [CrossRef]
- Lewis, K.L.; Del Cid, N.; Traver, D. Perspectives on antigen presenting cells in zebrafish. Dev. Comp. Immunol. 2014, 46, 63–73. [Google Scholar] [CrossRef]
- Conserva, M.R.; Anelli, L.; Zagaria, A.; Specchia, G.; Albano, F. The Pleiotropic Role of Retinoic Acid/Retinoic Acid Receptors Signaling: From Vitamin A Metabolism to Gene Rearrangements in Acute Promyelocytic Leukemia. Int. J. Mol. Sci. 2019, 20, 2921. [Google Scholar] [CrossRef]
- Sakai, M.; Hikima, J.; Kono, T. Fish cytokines: Current research and applications. Fish. Sci. 2021, 87, 1–9. [Google Scholar] [CrossRef]
- Hernandez, L.H.; Teshima, S.-I.; Koshio, S.; Ishikawa, M.; Tanaka, Y.; Alam, S. Effects of vitamin A on growth, serum anti-bacterial activity and transaminase activities in the juvenile Japanese flounder, Paralichthys olivaceus. Aquaculture 2007, 262, 444–450. [Google Scholar] [CrossRef]
- Cuesta, A.; Ortuno, J.; Rodrigues, A.; Esteban, M.A.; Meseguer, J. Changes in some innate defense parameters of seabream (Sparus aurata L.) induced by retinol acetate. Fish Shellfish. Immunol. 2002, 13, 279–291. [Google Scholar] [CrossRef]
- Biller, J.D.; Takahashi, L.S. Oxidative stress and fish immune system: Phagocytosis and leukocyte respiratory burst activity. An. Da Acad. Bras. De Ciênc. 2018, 90, 3403–3414. [Google Scholar] [CrossRef] [PubMed]
- Shastak, Y.; Pelletier, W. Captivating Colors, Crucial Roles: Astaxanthin’s Antioxidant Impact on Fish Oxidative Stress and Reproductive Performance. Animals 2023, 13, 3357. [Google Scholar] [CrossRef] [PubMed]
- Filho, D.W. Reactive oxygen species, antioxidants and fish mitochondria. Front. Biosci. 2007, 12, 1229–1237. [Google Scholar] [CrossRef]
- Checa, J.; Aran, J.M. Reactive Oxygen Species: Drivers of Physiological and Pathological Processes. J. Inflamm. Res. 2020, 13, 1057–1073. [Google Scholar] [CrossRef]
- Li, X.; Naseem, S.; Hussain, R.; Ghaffar, A.; Li, K.; Khan, A. Evaluation of DNA Damage, Biomarkers of Oxidative Stress, and Status of Antioxidant Enzymes in Freshwater Fish (Labeo rohita) Exposed to Pyriproxyfen. Oxidative Med. Cell. Longev. 2022, 2022, 5859266. [Google Scholar] [CrossRef]
- Alghazeer, R.; Howell, N.K. Formation of 4-hydroxynonenal (4-HNE) in frozen mackerel (Scomber scombrus) in the presence and absence of green tea. J. Sci. Food Agric. 2008, 88, 1128–1134. [Google Scholar] [CrossRef]
- Bastos, F.F.; Tobar, S.A.; Dantas, R.F.; Silva, E.S.; Nogueira, N.P.; Paes, M.C.; Righi, B.D.; Bastos, J.C.; Bastos, V.L. Melatonin affects conjugation of 4-hydroxynonenal with glutathione in liver of pacu, a hypoxia-tolerant fish. Fish Physiol. Biochem. 2013, 39, 1205–1214. [Google Scholar] [CrossRef]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxidative Med Cell Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef]
- Garcia, D.; Lima, D.; da Silva, D.G.H.; de Almeida, E.A. Decreased malondialdehyde levels in fish (Astyanax altiparanae) exposed to diesel: Evidence of metabolism by aldehyde dehydrogenase in the liver and excretion in water. Ecotoxicol. Environ. Saf. 2020, 190, 110107. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant Defenses in Fish: Biotic and Abiotic Factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Zommara, M.; Eweedah, N.M.; Helal, A.I.; Aboel-Darag, M.A. The potential role of nano-selenium and vitamin C on the performances of Nile tilapia (Oreochromis niloticus). Environ. Sci. Pollut. Res. 2020, 27, 9843–9852. [Google Scholar] [CrossRef] [PubMed]
- Zengin, H. The effects of feeding and starvation on antioxidant defence, fatty acid composition and lipid peroxidation in reared Oncorhynchus mykiss fry. Sci. Rep. 2021, 11, 16716. [Google Scholar] [CrossRef] [PubMed]
- Battisti, E.K.; Marasca, S.; Durigon, E.G.; Villes, V.S.; Schneider, T.L.S.; Uczay, J.; Peixoto, N.C.; Lazzari, R. Growth and oxidative parameters of Rhamdia quelen fed dietary levels of vitamin A. Aquaculture 2017, 474, 11–17. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- Dao, D.Q.; Ngo, T.C.; Thong, N.M.; Nam, P.C. Is vitamin A an antioxidant or a pro-oxidant? J. Phys. Chem. B 2017, 121, 9348–9357. [Google Scholar] [CrossRef]
- Palace, V.P.; Khaper, N.; Qin, Q.; Singal, P.K. Antioxidant potentials of vitamin A and carotenoids and their relevance to heart disease. Free Radic. Biol. Med. 1999, 26, 746–761. [Google Scholar] [CrossRef]
- Blaner, W.S.; Shmarakov, I.O.; Traber, M.G. Vitamin A and vitamin E: Will the real antioxidant please stand up? Annu. Rev. Nutr. 2021, 41, 105–131. [Google Scholar] [CrossRef]
- Wang, J.; Zou, L.; Jiang, P.; Yao, M.; Xu, Q.; Hong, Q.; Zhu, J.; Chi, X. Vitamin A ameliorates valproic acid-induced autism-like symptoms in developing zebrafish larvae by attenuating oxidative stress and apoptosis. Neurotoxicology 2024, 101, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Park, U.-H.; Han, H.S.; Um, E.; An, X.-H.; Kim, E.-J.; Um, S.J. Redox regulation of transcriptional activity of retinoic acid receptor by thioredoxin glutathione reductase (TGR). Biochem. Biophys. Res. Commun. 2009, 390, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Flohé, L. Regulatory phenomena in the glutathione peroxidase superfamily. Antioxid. Redox Signal. 2020, 33, 498–516. [Google Scholar] [CrossRef] [PubMed]
- Tourniaire, F.; Musinovic, H.; Gouranton, E.; Astier, J.; Marcotorchino, J.; Arreguin, A.; Bernot, D.; Palou, A.; Bonet, M.L.; Ribot, J.; et al. All-trans retinoic acid induces oxidative phosphorylation and mitochondria biogenesis in adipocytes. J. Lipid Res. 2015, 56, 1100–1109. [Google Scholar] [CrossRef]
- Rajawat, Y.; Hilioti, Z.; Bossis, I. Autophagy: A target for retinoic acids. Autophagy 2010, 6, 1224–1226. [Google Scholar] [CrossRef]
- Valenzuela, C.A.; Azúa, M.; Álvarez, C.A.; Schmitt, P.; Ojeda, N.; Mercado, L. Evidence of the Autophagic Process during the Fish Immune Response of Skeletal Muscle Cells against Piscirickettsia salmonis. Animals 2023, 13, 880. [Google Scholar] [CrossRef]
- Zhou, Z.; He, Y.; Wang, S.; Wang, Y.; Shan, P.; Li, P. Autophagy regulation in teleost fish: A double-edged sword. Aquaculture 2022, 558, 738369. [Google Scholar] [CrossRef]
- Fontagné, S.; Lataillade, E.; Brèque, J.; Kaushik, S. Lipid peroxidative stress and antioxidant defence status during ontogeny of rainbow trout (Oncorhynchus mykiss). Br. J. Nutr. 2008, 100, 102–111. [Google Scholar] [CrossRef]
- Jiang, W.D.; Zhou, X.Q.; Zhang, L.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.Y.; Tang, L.; Tang, W.N.; Zhang, Y.A.; et al. Vitamin A deficiency impairs intestinal physical barrier function of fish. Fish Shellfish. Immunol. 2019, 87, 546–558. [Google Scholar] [CrossRef]
- Wu, F.; Zhu, W.; Liu, M.; Chen, C.; Chen, J.; Tan, Q. Effects of Dietary Vitamin A on Growth Performance, Blood Biochemical Indices and Body Composition of Juvenile Grass Carp (Ctenopharyngodon Idellus). Turk. J. Fish. Aquat. Sci. 2016, 16, 339–345. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, M.; Liang, X.-F.; Peng, D.; Xie, R.; Wu, D. Dietary supplementation of VA enhances growth, feed utilization, glucose and lipid metabolism, appetite, and antioxidant capacity of Chinese perch (Siniperca chuatsi). Fish Physiol. Biochem. 2024, 50, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Deering, M.J.; Paradis, H.; Ahmad, R.; Al-Mehiawi, A.S.; Gendron, R.L. The role of dietary vitamin A in mechanisms of cataract development in the teleost lumpfish (Cyclopterus lumpus L). J. Fish Dis. 2024, 47, e13899. [Google Scholar] [CrossRef]
- Liang, D.; Yang, Q.; Tan, B.; Dong, X.; Chi, S.; Liu, H.; Zhang, S. Dietary vitamin A deficiency reduces growth performance, immune function of intestine, and alters tight junction proteins of intestine for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Fish Shellfish. Immunol. 2020, 107 Pt A, 346–356. [Google Scholar] [CrossRef]
- Devlin, R.H.; Leggatt, R.A.; Benfey, T.J. Chapter 7—Genetic modification of growth in fish species used in aquaculture: Phenotypic and physiological responses. In Fish Physiology; Benfey, T.J., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 38, pp. 237–272. [Google Scholar] [CrossRef]
- Breck, J.E. Body composition in fishes: Body size matters. Aquaculture 2014, 433, 40. [Google Scholar] [CrossRef]
- Young, T.; Laroche, O.; Walker, S.P.; Miller, M.R.; Casanovas, P.; Steiner, K.; Esmaeili, N.; Zhao, R.; Bowman, J.P.; Wilson, R.; et al. Prediction of Feed Efficiency and Performance-Based Traits in Fish via Integration of Multiple Omics and Clinical Covariates. Biology 2023, 12, 1135. [Google Scholar] [CrossRef] [PubMed]
- FAO (Food and Agriculture Organisation of the United Nations). The Measurement of Fish and Shellfish. 2022. Available online: https://www.fao.org/4/F0752E/F0752E03.htm (accessed on 7 July 2024).
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Crane, D.P.; Ogle, D.H.; Shoup, D.E. Use and misuse of a common growth metric: Guidance for appropriately calculating and reporting specific growth rate. Rev. Aquacult. 2020, 12, 1542–1547. [Google Scholar] [CrossRef]
- Ahmed, I.; Jan, K.; Fatma, S.; Dawood, M.A.O. Muscle proximate composition of various food fish species and their nutritional significance: A review. J. Anim. Physiol. Anim. Nutr. 2022, 106, 690–719. [Google Scholar] [CrossRef]
- Peng, C.; Wang, Q.; Chen, J.; Yang, H.; Zhang, W.; Wang, D.; Li, S.; Tao, M.; Shi, H.; Lin, H.; et al. Retinoic acid and androgen influence germ cells development and meiotic initiation in juvenile orange-spotted grouper, Epinephelus coioides. Gen. Comp. Endocrinol. 2020, 289, 113379. [Google Scholar] [CrossRef]
- Fraher, D.; Mann, R.J.; Dubuisson, M.J.; Ellis, M.K.; Yu, T.; Walder, K.; Ward, A.C.; Winkler, C.; Gibert, Y. The endocannabinoid system and retinoic acid signaling combine to influence bone growth. Mol. Cell. Endocrinol. 2021, 529, 111267. [Google Scholar] [CrossRef]
- Brown, G. Retinoic acid receptor regulation of decision-making for cell differentiation. Front. Cell Dev. Biol. 2023, 11, 1182204. [Google Scholar] [CrossRef]
- Qiu, J.; Nordling, S.; Vasavada, H.H.; Butcher, E.C.; Hirschi, K.K. Retinoic Acid Promotes Endothelial Cell Cycle Early G1 State to Enable Human Hemogenic Endothelial Cell Specification. Cell Rep. 2020, 33, 108465. [Google Scholar] [CrossRef] [PubMed]
- Haga, Y.; Du, S.-J.; Satoh, S.; Kotani, T.; Fushimi, H.; Takeuchi, T. Analysis of the mechanism of skeletal deformity in fish larvaeusing a vitamin A-induced bone deformity model. Aquaculture 2011, 315, 26–33. [Google Scholar] [CrossRef]
- Jackman, W.R.; Gibert, Y. Retinoic Acid Signaling and the Zebrafish Dentition during Development and Evolution. Subcell. Biochem. 2020, 95, 175–196. [Google Scholar] [CrossRef] [PubMed]
- Mackowetzky, K.; Dicipulo, R.; Fox, S.C.; Philibert, D.A.; Todesco, H.; Doshi, J.D.; Kawakami, K.; Tierney, K.; Waskiewicz, A.J. Retinoic acid signaling regulates late stages of semicircular canal morphogenesis and otolith maintenance in the zebrafish inner ear. Dev. Dyn. 2022, 251, 1798–1815. [Google Scholar] [CrossRef]
- Paulissen, E.; Palmisano, N.J.; Waxman, J.S.; Martin, B.L. Somite morphogenesis is required for axial blood vessel formation during zebrafish embryogenesis. eLife 2022, 11, e74821. [Google Scholar] [CrossRef]
- Durbin, L.; Sordino, P.; Barrios, A.; Gering, M.; Thisse, C.; Thisse, B.; Brennan, C.; Green, A.; Wilson, S.; Holder, N. Anteroposterior patterning is required within segments for somite boundary formation in developing zebrafish. Development 2000, 27, 1703–1713. [Google Scholar] [CrossRef]
- Stickney, H.L.; Barresi, M.J.F.; Devoto, S.H. Somite development in zebrafish. Dev. Dyn. 2000, 219, 287–303. [Google Scholar] [CrossRef]
- Ward, A.B.; Mehta, R.S. Axial Elongation in Fishes: Using Morphological Approaches to Elucidate Developmental Mechanisms in Studying Body Shape. Integr. Comp. Biol. 2010, 50, 1106–1119. [Google Scholar] [CrossRef]
- Lleras Forero, L.; Narayanan, R.; Huitema, L.F.; Van Bergen, M.; Apschner, A.; Peterson-Maduro, J.; Logister, I.; Valentin, G.; Morelli, L.G.; Oates, A.C.; et al. Segmentation of the zebrafish axial skeleton relies on notochord sheath cells and not on the segmentation clock. eLife 2018, 7, e33843. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development (OECD). Detailed Review Paper on the Retinoid System. OECD Series on Testing and Assessment, Nr.343, Paris. 2021. Available online: https://one.oecd.org/document/ENV/CBC/MONO(2021)20/en/pdf (accessed on 11 February 2024).
- Isabella, A.J.; Barsh, G.R.; Stonick, J.A.; Dubrulle, J.; Moens, C.B. Retinoic Acid Organizes the Zebrafish Vagus Motor Topographic Map via Spatiotemporal Coordination of Hgf/Met Signaling. Dev. Cell 2020, 53, 344–357.e5. [Google Scholar] [CrossRef]
- Udo, I.U. Effects of dietary vitamin a level on growth, feed utilization and survival of juvenile North African catfish (Clarias gariepinus). Livest. Res. Rural. Dev. 2017, 29, 1–8. Available online: https://www.lrrd.org/lrrd29/2/udo29022.htm#:~:text=gariepinus%20should%20contain%20vitamin%20A,growth%20and%20efficient%20feed%20utilization (accessed on 7 July 2024).
- Hu, C.-J.; Chen, S.-M.; Pan, C.-H.; Huang, C.-H. Effects of dietary vitamin A or β-carotene concentrations on growth of juvenile hybrid tilapia, Oreochromis niloticus × O. aureus. Aquaqulture 2006, 253, 602–607. [Google Scholar] [CrossRef]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef] [PubMed]
- Shekhova, E. Mitochondrial Reactive Oxygen Species as Major Effectors of Antimicrobial Immunity. PLoS Pathog. 2020, 16, e1008470. [Google Scholar] [CrossRef]
- Victor, V.M.; Rocha, M.; De la Fuente, M. Immune Cells: Free Radicals and Antioxidants in Sepsis. Int. Immunopharmacol. 2004, 4, 327–347. [Google Scholar] [CrossRef]
- Tesoriere, L.; Bongiorno, A.; Pintaudi, A.M.; D’Anna, R.; D’Arpa, D.; Livrea, M.A. Synergistic interactions between vitamin A and vitamin E against lipid peroxidation in phosphatidylcholine liposomes. Arch. Biochem. Biophys. 1996, 326, 57–63. [Google Scholar] [CrossRef]
- Tesoriere, L.; Ciaccio, M.; Bongiorno, A.; Riccio, A.; Pintaudi, A.; Livrea, M. antioxidant activity of all-trans-retinol in homogeneous solution and in phosphatidylcholine liposomes. Arch. Biochem. Biophys. 1993, 307, 217–223. [Google Scholar] [CrossRef]
- Moore, T. The effect of vitamin E deficiency on vitamin A reserves of the rat. Biochem. J. 1940, 34, 1321. [Google Scholar] [CrossRef]
- Wang, Z.-L.; Pang, S.-J.; Zhang, K.-W.; Li, P.-G.; Yang, C. Dietary vitamin A modifies the gut microbiota and intestinal tissue transcriptome, impacting intestinal permeability and the release of inflammatory factors, thereby influencing Aβ pathology. Front. Nutr. 2024, 11, 1367086. [Google Scholar] [CrossRef]
- Fernández, I.; Ortiz-Delgado, J.B.; Sarasquete, C.; Gisbert, E. Vitamin A effects on vertebral bone tissue homeostasis in gilthead sea bream (Sparus aurata) juveniles. J. Appl. Ichthyol. 2012, 28, 419–426. [Google Scholar] [CrossRef]
- Lucock, M.; Jones, P.; Martin, C.; Beckett, E.; Yates, Z.; Furst, J.; Veysey, M. Vitamin D: Beyond Metabolism. J. Evid. Based Complement. Altern. Med. 2015, 20, 310–322. [Google Scholar] [CrossRef]
- Cheng, K.; Huang, Y.; Wang, C.; Ali, W.; Karrow, N.A. Physiological function of vitamin D3 in fish. Rev. Aquac. 2023, 15, 1732–1748. [Google Scholar] [CrossRef]
- MacDonald, P.N.; Dowd, D.R.; Nakajima, S.; Galligan, M.A.; Reeder, M.C.; Haussler, C.A.; Ozato, K.; Haussler, M.R. Retinoid X receptors stimulate and 9-cis retinoic acid inhibits 1,25-dihydroxyvitamin D3-activated expression of the rat osteocalcin gene. Mol. Cell. Biol. 1993, 13, 5907–5917. [Google Scholar]
- Pierens, S.L.; Fraser, D.R. The origin and metabolism of vitamin D in rainbow trout. J. Steroid Biochem. Mol. Biol. 2015, 145, 58–64. [Google Scholar] [CrossRef]
- Džopalić, T.; Božić-Nedeljković, B.; Jurišić, V. The role of vitamin A and vitamin D in modulation of the immune response with a focus on innate lymphoid cells. Cent. Eur. J. Immunol. 2021, 46, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Ruiter, B.; Patil, S.U.; Shreffler, W.G. Vitamins A and D have antagonistic effects on expression of effector cytokines and gut-homing integrin in human innate lymphoid cells. Clin. Exp. Allergy 2015, 45, 1214–1225. [Google Scholar] [CrossRef]
- De Tullio, M.C. The Mystery of Vitamin C. Nat. Educ. 2010, 3, 48. [Google Scholar]
- Ching, B.; Chew, S.F.; Ip, Y.K. Ascorbate synthesis in fishes: A review. IUBMB Life 2015, 67, 69–76. [Google Scholar] [CrossRef]
- Lee, K.W.; Yoo, H.K.; Kim, S.-S.; Han, G.S.; Jung, M.M.; Kim, H.S. Effect of Dietary Vitamin C Supplementation on Growth Performance and Biochemical Parameters in Grower Walleye Pollock, Gadus chalcogrammus. Animals 2024, 14, 1026. [Google Scholar] [CrossRef]
- Kanter, M.; Coskun, O.; Armutcu, F.; Uz, Y.H.; Kizilay, G. Protective effects of vitamin C, alone or in combination with vitamin A, on endotoxin-induced oxidative renal tissue damage in rats. Tohoku J. Exp. Med. 2005, 206, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.M.; Yu, H.R.; Li, L.Y.; Li, M.; Qiu, X.Y.; Fan, X.Q.; Fan, Y.L.; Shan, L.L. Effects of Dietary Vitamin C on the Growth Performance, Biochemical Parameters, and Antioxidant Activity of Coho Salmon Oncorhynchus kisutch (Walbaum, 1792) Postsmolts. Aquac. Nutr. 2022, 2022, 6866578. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; To, V.P.T.H.; Santigosa, E.; Dumas, A.; Hernandez, J.M. Vitamin nutrition in salmonid aquaculture: From avoiding deficiencies to enhancing functionalities. Aquaculture 2022, 561, 738654. [Google Scholar] [CrossRef]
- Manam, V.K. Fish feed nutrition and its management in aquaculture. Int. J. Fish Aquat. Stud. 2023, 11, 58–61. [Google Scholar] [CrossRef]
- Terech-Majewska, E.; Pajdak, J.; Siwicki, A.K. Water as a source of macronutrients and micronutrients for fish, with special emphasis on the nutritional requirements of two fish species: The common carp (Cyprinus carpio) and the rainbow trout (Oncorhynchus mykiss). J. Elem. 2016, 21, 947–961. [Google Scholar]
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef]
- Akhter, F.; Siddiquei, H.R.; Alahi, M.E.E.; Mukhopadhyay, S.C. Recent Advancement of the Sensors for Monitoring the Water Quality Parameters in Smart Fisheries Farming. Computers 2021, 10, 26. [Google Scholar] [CrossRef]
- Lindholm-Lehto, P. Water quality monitoring in recirculating aquaculture systems. Aquac. Fish Fish. 2023, 3, 113–131. [Google Scholar] [CrossRef]
- Sneddon, L.U.; Wolfenden, D.C.C.; Thomson, J.S. Stress management and welfare. Fish. Physiol. 2016, 35, 463–539. [Google Scholar]
- Long, L.; Zhang, H.; Ni, Q.; Liu, H.; Wu, F.; Wang, X. Effects of stocking density on growth, stress, and immune responses of juvenile Chinese sturgeon (Acipenser sinensis) in a recirculating aquaculture system. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 219, 25–34. [Google Scholar] [CrossRef]
- Dara, M.; Carbonara, P.; La Corte, C.; Parrinello, D.; Cammarata, M.; Parisi, M.G. Fish Welfare in Aquaculture: Physiological and Immunological Activities for Diets, Social and Spatial Stress on Mediterranean Aqua Cultured Species. Fishes 2023, 8, 414. [Google Scholar] [CrossRef]
- Gerber, B.; Stamer, A.; Stadtlander, T. Environmental Enrichment and Its Effects on Welfare in Fish. FiBL Schweiz. 2015. Available online: https://orgprints.org/id/eprint/29142/1/Gerber-etal-2015-Environmental-Enrichment-and-its-effects-on-welfare-in-fish-FiBL-Review.pdf (accessed on 9 July 2024).
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Caballero-Froilán, J.C.; Jiménez-García, M.; Capó, X.; Tejada, S.; Saraiva, J.L.; Sureda, A.; Moranta, D. Enriched environments enhance cognition, exploratory behaviour and brain physiological functions of Sparus aurata. Sci. Rep. 2020, 10, 11252. [Google Scholar] [CrossRef] [PubMed]
- Browning, H. Improving welfare assessment in aquaculture. Front. Vet. Sci. 2023, 10, 1060720. [Google Scholar] [CrossRef]
- Hemal, M.M.; Rahman, A.; Nurjahan; Islam, F.; Ahmed, S.; Kaiser, M.S.; Ahmed, M.R. An Integrated Smart Pond Water Quality Monitoring and Fish Farming Recommendation Aquabot System. Sensors 2024, 24, 3682. [Google Scholar] [CrossRef] [PubMed]
- Gudding, R.; Lillehaug, A.; Tavornpanich, S. Immunoprophylaxis in Biosecurity Programs. J. Appl. Aquac. 2015, 27, 220–227. [Google Scholar] [CrossRef]
- Wang, B.; Thompson, K.D.; Wangkahart, E.; Yamkasem, J.; Bondad-Reantaso, M.G.; Tattiyapong, P.; Jian, J.; Surachetpong, W. Strategies to enhance tilapia immunity to improve their health in aquaculture. Rev. Aquac. 2023, 15 (Suppl. S1), 41–56. [Google Scholar] [CrossRef]
- Varghese, T.; Gopan, A.; Rejish Kumar, V. Reinventing the Micronutrients beyond Nutrition: Functions in Immune Modulation and Stress Mitigation of Fish. In Biotechnological Advances in Aquaculture Health Management; Gupta, S.K., Giri, S.S., Eds.; Springer: Singapore, 2021. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organisation of the United Nations). Causes and Effects of Pollution on Fish. 2024. Available online: https://www.fao.org/fishery/docs/CDrom/aquaculture/a0844t/docrep/009/T1623E/T1623E03.htm (accessed on 11 July 2024).
- Brooks, P.R.; Crowe, T.P. Combined Effects of Multiple Stressors: New Insights Into the Influence of Timing and Sequence. Front. Ecol. Evol. 2019, 7, 387. [Google Scholar] [CrossRef]
- Keleştemur, G.T.; Özdemir, Y. Effect of Dietary Vitamin A and E on Tissues Vitamin Concentrations and Lipid Peroxidation of Juvenile Rainbow Trout at Different Flow Rates. Turk. J. Sci. Technol. 2016, 11, 21–29. [Google Scholar]
- Zhou, C.; Yang, S.; Ka, W.; Gao, P.; Li, Y.; Long, R.; Wang, J. Association of Gut Microbiota With Metabolism in Rainbow Trout Under Acute Heat Stress. Front. Microbiol. 2022, 13, 846336. [Google Scholar] [CrossRef]
- Kong, M.; Zhao, W.; Wang, C.; Qi, J.; Liu, J.; Zhang, Q. A Well-Established Gut Microbiota Enhances the Efficiency of Nutrient Metabolism and Improves the Growth Performance of Trachinotus ovatus. Int. J. Mol. Sci. 2024, 25, 5525. [Google Scholar] [CrossRef] [PubMed]
- Maurya, V.K.; Shakya, A.; Bashir, K.; Kushwaha, S.C.; McClements, D.J. Vitamin A fortification: Recent advances in encapsulation technologies. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2772–2819. [Google Scholar] [CrossRef] [PubMed]
- Shastak, Y.; Pelletier, W.; Kuntz, A. Insights into Analytical Precision: Understanding the Factors Influencing Accurate Vitamin A Determination in Various Samples. Analytica 2024, 5, 54–73. [Google Scholar] [CrossRef]
- Shastak, Y.; Pelletier, W. Nutritional Balance Matters: Assessing the Ramifications of Vitamin A Deficiency on Poultry Health and Productivity. Poultry 2023, 2, 493–515. [Google Scholar] [CrossRef]
- Arshad, R.; Gulshad, L.; Haq, I.U.; Farooq, M.A.; Al-Farga, A.; Siddique, R.; Manzoor, M.F.; Karrar, E. Nanotechnology: A novel tool to enhance the bioavailability of micronutrients. Food Sci. Nutr. 2021, 9, 3354–3361. [Google Scholar] [CrossRef]
- Vettom, E.J.; Danve, A.; Khalasi, Y. Precision Aquaculture: Integrating Technology for Enhanced Production and Sustainability. Agric. Mag. 2024, 3, 203–211. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Form | Function | Sources |
|---|---|---|
| Retinyl esters | Storage | [8] |
| All-trans-retinol | Transport; antioxidant; precursor of active forms | [9,10] |
| 13-cis retinol | Transport; precursor of active forms | [11] |
| 3-dehydroretinol | Intermediate and precursor of active forms | [12] |
| Retinaldehyde (11-cis-retinal) | Vision | [13] |
| All-trans-retinoic acid | Cellular proliferation and specialization, immunomodulation, oxidative stress defense, reproductive and embryonic development, skeletal integrity, morphogenesis, neurogenesis, hematopoiesis | [14,15,16,17,18] |
| 13-cis-retinoic acid | Morphogen, modulation of the effects of calcitriol, central nervous system patterning, signaling molecule | [18,19] |
| 9-cis-retinoic acid | Embryonic development, hormone regulation | [20] |
| 14-hydroxy-4,14-retro-retinol | Cofactor for growth of B lymphocytes and for activation of T lymphocytes | [21,22] |
| All-trans-4-hydroxy retinoic acid and 4-oxo-retinoic acid | Gap junctional cell communication, favored form to facilitate elimination of all-trans-retinoic acid | [12] |
| Signs of Deficiency | Reference | Signs of Excess | Reference |
|---|---|---|---|
| Compromised immune function | [4,25] | Slower growth | [26] |
| Abnormalities in reproduction | [27] | Visual impairment | [28] |
| Degeneration of skeletal and cardiac muscle fibers | [29] | Abnormal vertebral growth and muscle pain | [28,30,31,32] |
| Hemorrhagic granulomatous spleen; necrotic granulomatous liver; decreased hepatosomatic index | [12,33] | Lower liver iron levels and pale-yellow livers | [26] |
| Hindered bone development, poorly formed bony tissue in the skull and vertebral protuberance | [29,34] | Increased vitamin A levels in the broodstock and eggs | [27] |
| Loss of appetite | [35] | Increased mortality | [26] |
| Stunted growth | [33] | Dry epithelia | [28] |
| Increased mortality | [12,29] | Fin abnormalities | [26] |
| Internal as well as subcutaneous hemorrhage at the fin bases and within the dermal layers | [30] | Skin disorders | [28] |
| Protruding, impaired visual acuity, exophthalmia | [33,35] | Reduced fat stores | [32] |
| Keratinization of epithelial tissues | [36] | ||
| Deficiency-related anemia | [33] | ||
| Abdominal depigmentation and edema, sometimes with ascites | [30,33] |
| Fish Species | Dietary Vitamin A Supplementation Levels | Challenge * | Vitamin A Effects on Immune Criteria | Reference |
|---|---|---|---|---|
| Grass carp (Ctenopharyngodon idella) | 0, 600, 1200, 1800, 2800, and 3800 IU/kg | Yes | Increased contents of complements, activities of acid phosphatase and lysozyme, mRNAs of β-defensin-1, liver-expressed antimicrobial peptide 2A and 2B, hepcidin, and anti-inflammatory cytokines like transforming growth factor β1 (TGF-β1), TGF-β2, interleukin-10 (IL-10), and IL-11. | [15] |
| Nile tilapia (Oreochromis niloticus) | 0, 2500, 5000, 10,000, and 20,000 IU/kg | Yes | Increased lysozyme activity and superoxide anion production | [61] |
| Japanese flounder (Paralichthys olivaceus) | 0, 10,000, and 25,000 IU/kg | No | Improved serum antibacterial activity | [77] |
| Gilthead seabream (Sparus aurata L.) | 0, 150 and 300 mg of retinyl acetate/kg | No | Enhanced respiratory burst activity in head-kidney leucocytes | [78] |
| Grass carp (Ctenopharyngodon idella) | 0.206, 0.413, 0.619, 0.963, and 1.307 mg of retinyl acetate/kg | Yes | Improved intestinal innate humoral immune response, and lower intestinal inflammation | [25] |
| Common carp (Cyprinus carpio var.) | 0, 0.03, 0.07, 0.11, 0.15, and 0.19 g as retinyl acetate/kg | No | Lower white blood cell count | [4] |
| Atlantic salmon (Salmo salar) | 0.37, 1.95, and 15 mg of retinyl acetate/kg | No | Improved kidney leucocyte migration, serum bactericidal activity, enhanced serum antiprotease activity | [56] |
| Stressor | Permissible Limit | Effects on Fish |
|---|---|---|
| Oxygen depletion | Dissolved oxygen levels should not fall below 5 mg/L for most freshwater fish. | Low dissolved oxygen can lead to fish suffocation and reduced growth. |
| Temperature fluctuations | Diurnal fluctuations in water temperature should not exceed a certain threshold, which is species-specific. | Rapid temperature changes can stress fish and impact metabolism. |
| Pollutants (heavy metals) | Varies by metal and species. In general, allowable concentrations are low (micrograms per liter or lower). | Heavy metals like lead, mercury, and cadmium can accumulate in fish tissues and harm health. |
| Pesticides and herbicides | Varies by chemical and species. Generally, very low concentrations are allowed (parts per billion). | These chemicals can disrupt fish physiology and impair reproduction. |
| Ammonia | Total ammonia nitrogen levels should be below 0.02 mg/L for freshwater fish. | High ammonia can damage fish gills and cause respiratory distress. |
| pH | Optimal pH ranges from 6.5 to 9.0, depending on the fish species. | Extreme pH levels can stress fish, affecting ion balance and survival. |
| Salinity | Varies widely by fish species. Some tolerate freshwater, while others require high salinity. | Salinity outside a fish’s tolerance range can cause osmotic stress. |
| UV radiation | Exposure should be limited, especially in shallow, clear waters. | Prolonged UV exposure can damage fish skin and eyes. |
| Microorganisms (pathogens) | Presence of pathogens like bacteria, viruses, and parasites should be minimized. | Infections can weaken fish and lead to disease outbreaks. |
| Toxic algal blooms | Concentrations of harmful algae should be monitored and controlled. | Toxins produced by algae can harm fish and other aquatic organisms. |
| Fish Species | Dietary Vitamin A Supplementation Levels | Challenge * | Vitamin A Effects on Oxidative Stress Status | Reference |
|---|---|---|---|---|
| Grass carp (Ctenopharyngodon idella) | 0, 600, 1200, 1800, 2800, and 3800 IU/kg | Yes | Lowered concentrations of reactive oxygen species, malondialdehyde, and protein carbonyl, increased both the activities and mRNAs of copper/zinc superoxide dismutase, glutathione transferases, glutathione peroxidase, and glutathione reductase. | [15] |
| Grass carp (Ctenopharyngodon Idellus) | 68, 328, 599, 1327, 2584 and 4769 IU/kg | No | Increased serum superoxide dismutase activity | [105] |
| Grass carp (Ctenopharyngodon idella) | 19, 607, 1209, 1798, 2805, and 3796 IU/kg | No | Decreased levels of muscle reactive oxygen species, malondialdehyde, and protein carbonyl, enhanced activities of antioxidative enzyme (catalase, copper/zinc superoxide dismutase, MnSOD, glutathione peroxidase, and glutathione reductase), as well as increased content of glutathione | [16] |
| Japanese flounder (Paralichthys olivaceus) | 0, 10,000, and 25,000 IU/kg | No | Higher glutamic-pyruvic transaminase activity | [77] |
| Common carp (Cyprinus carpio var.) | 0, 0.03, 0.07, 0.11, 0.15, and 0.19 g as retinyl acetate/kg | No | Reduced serum malondialdehyde levels | [4] |
| Chinese perch (Siniperca chuatsi) | 0, 20, 40, 60, and 80 mg of retinyl acetate/kg | No | Upregulation of antioxidant genes (Nrf2 and GPx) | [106] |
| Lumpfish (Cyclopterus lumpus L.) | 5000, 15,000 and 120,000 IU/kg | No | Involvement in reducing oxidative stress in the eye lens at the moderate supplementation levels | [107] |
| Hybrid grouper (Epinephelus fuscoguttatus×Epinephelus lanceolatus) | 317, 1136, 2038, 4142, 7715, 15,204 IU/kg | No | Higher blood serum SOD activity, total antioxidant capacity and reduced malondialdehyde level | [108] |
| Fish Species | Dietary Vitamin A Supplementation Levels | Challenge * | Vitamin A Effects on Growth Parameters | Reference |
|---|---|---|---|---|
| Nile tilapia (Oreochromis niloticus) | 0, 2500, 5000, 10,000, and 20,000 IU/kg | Yes | Higher weight gain and feed intake; improved feed efficiency ratio. | [61] |
| African catfish (Clarias gariepinus) | 0, 833, and 1666 IU/kg | No | Improved final weight | [130] |
| Japanese flounder (Paralichthys olivaceus) | 0, 10,000, and 25,000 IU/kg | No | Improved weight gain and specific growth rate | [77] |
| Grass carp (Ctenopharyngodon Idellus) | 68, 328, 599, 1327, 2584 and 4769 IU/kg | No | Increased specific growth rate | [104] |
| Nile tilapia (Oreochromis niloticus) | 0, 600, 1200, 1800, 2400, 3000, 3600, 4200, 4800 and 5400 IU/kg | No | Improved final weight and weight gain | [33] |
| Grass carp (Ctenopharyngodon idella) | 0.206, 0.413, 0.619, 0.963, and 1.307 mg of retinyl acetate/kg | Yes | Improved growth performance | [25] |
| Hybrid tilapia (Oreochromis niloticus × O. aureus) | 0, 1000, 2000, 6000, 18,000, and 50,000 IU/kg | No | Improved weight gain | [131] |
| Common carp (Cyprinus carpio var.) | 0, 0.03, 0.07, 0.11, 0.15, and 0.19 g as retinyl acetate/kg | No | Improvements in growth parameters such as live weight gain, feed conversion ratio, protein efficiency ratio, specific growth rate, and body protein deposition | [4] |
| Chinese perch (Siniperca chuatsi) | 0, 20, 40, 60, and 80 mg of retinyl acetate/kg | No | Improved growth and feed utilization | [106] |
| Hybrid grouper (Epinephelus fuscoguttatus × Epinephelus lanceolatus) | 317, 1136, 2038, 4142, 7715, 15,204 IU/kg | No | Improved weight gain, specific growth rate, feed conversion ratio and protein efficiency ratio | [108] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shastak, Y.; Pelletier, W. Vitamin A in Fish Well-Being: Integrating Immune Strength, Antioxidant Capacity and Growth. Fishes 2024, 9, 330. https://doi.org/10.3390/fishes9080330
Shastak Y, Pelletier W. Vitamin A in Fish Well-Being: Integrating Immune Strength, Antioxidant Capacity and Growth. Fishes. 2024; 9(8):330. https://doi.org/10.3390/fishes9080330
Chicago/Turabian StyleShastak, Yauheni, and Wolf Pelletier. 2024. "Vitamin A in Fish Well-Being: Integrating Immune Strength, Antioxidant Capacity and Growth" Fishes 9, no. 8: 330. https://doi.org/10.3390/fishes9080330
APA StyleShastak, Y., & Pelletier, W. (2024). Vitamin A in Fish Well-Being: Integrating Immune Strength, Antioxidant Capacity and Growth. Fishes, 9(8), 330. https://doi.org/10.3390/fishes9080330