
Natural Food Intake and Its Contribution to Tambaqui Growth in Fertilized and Unfertilized Ponds

 ,
,  and
and 
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Design and Conditions
2.2. Plankton Availability
2.3. Stomach Content Analysis
2.4. Isotopic Analysis
2.5. Assimilation of Feed into Muscle Using Isotopic Analysis
2.6. Water Quality Analyses
2.7. Economic Analysis
2.8. Statistical Analysis
2.9. Legal and Ethical Aspects
3. Results
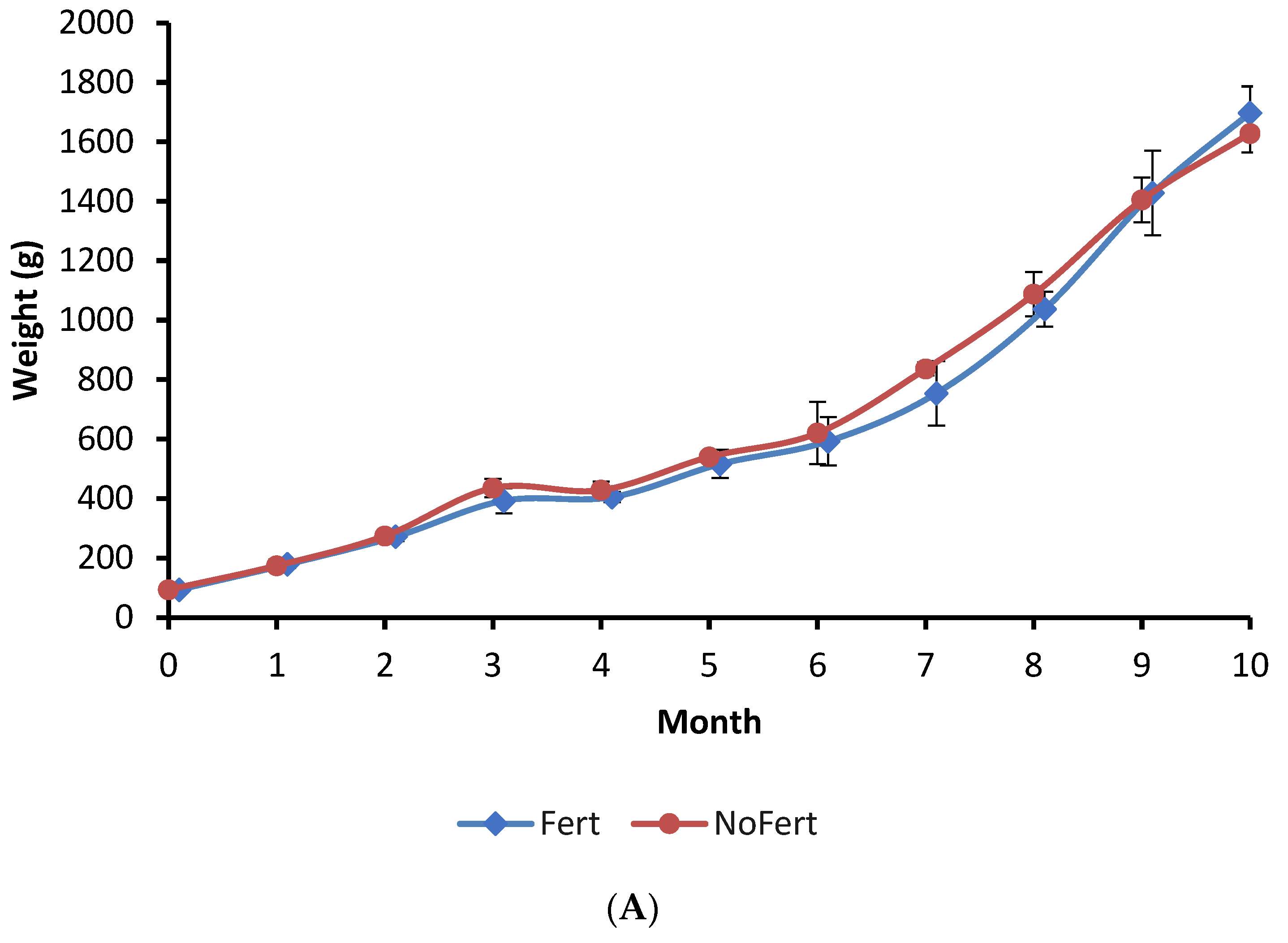
3.1. Tambaqui Performance in Ponds
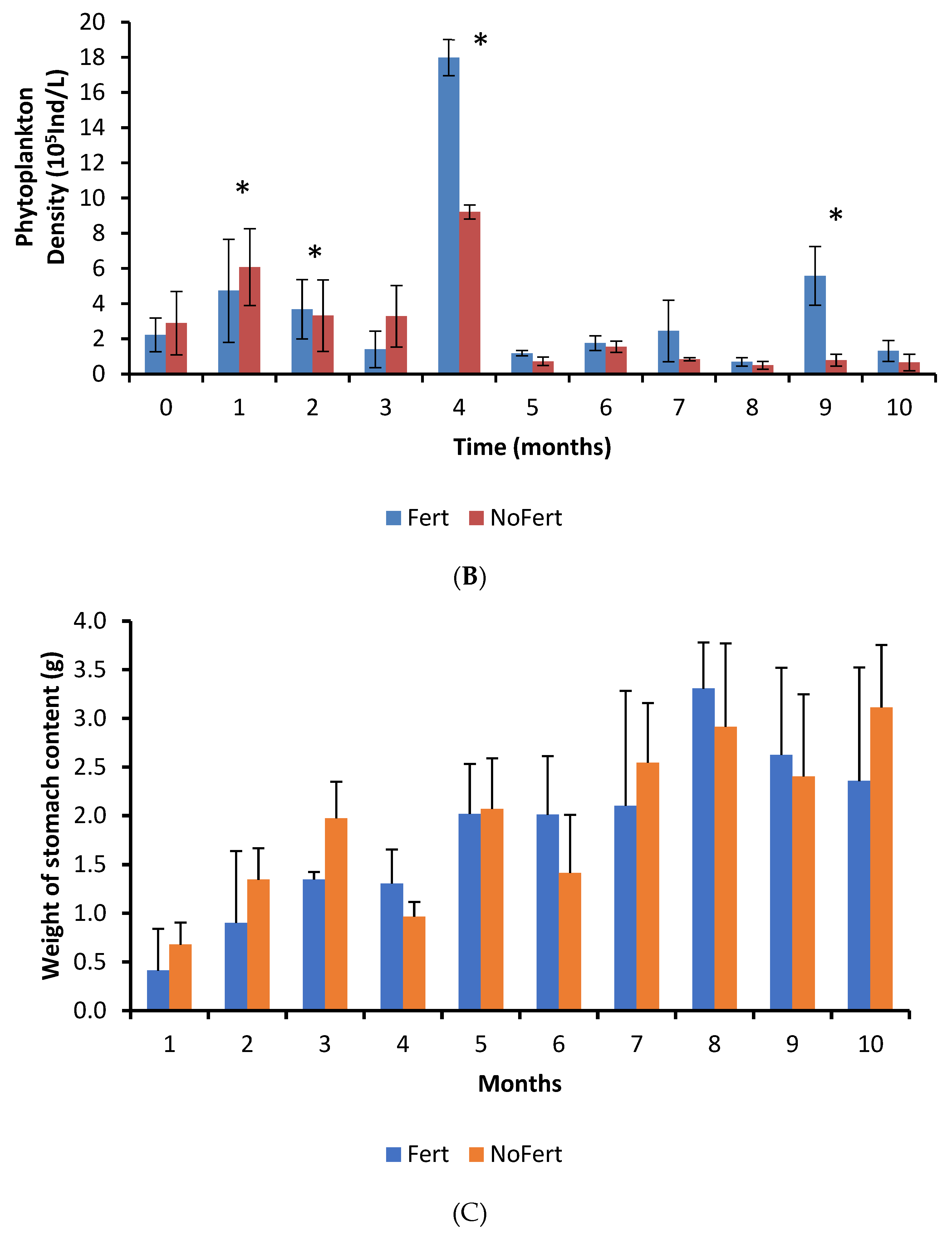
3.2. Plankton Community
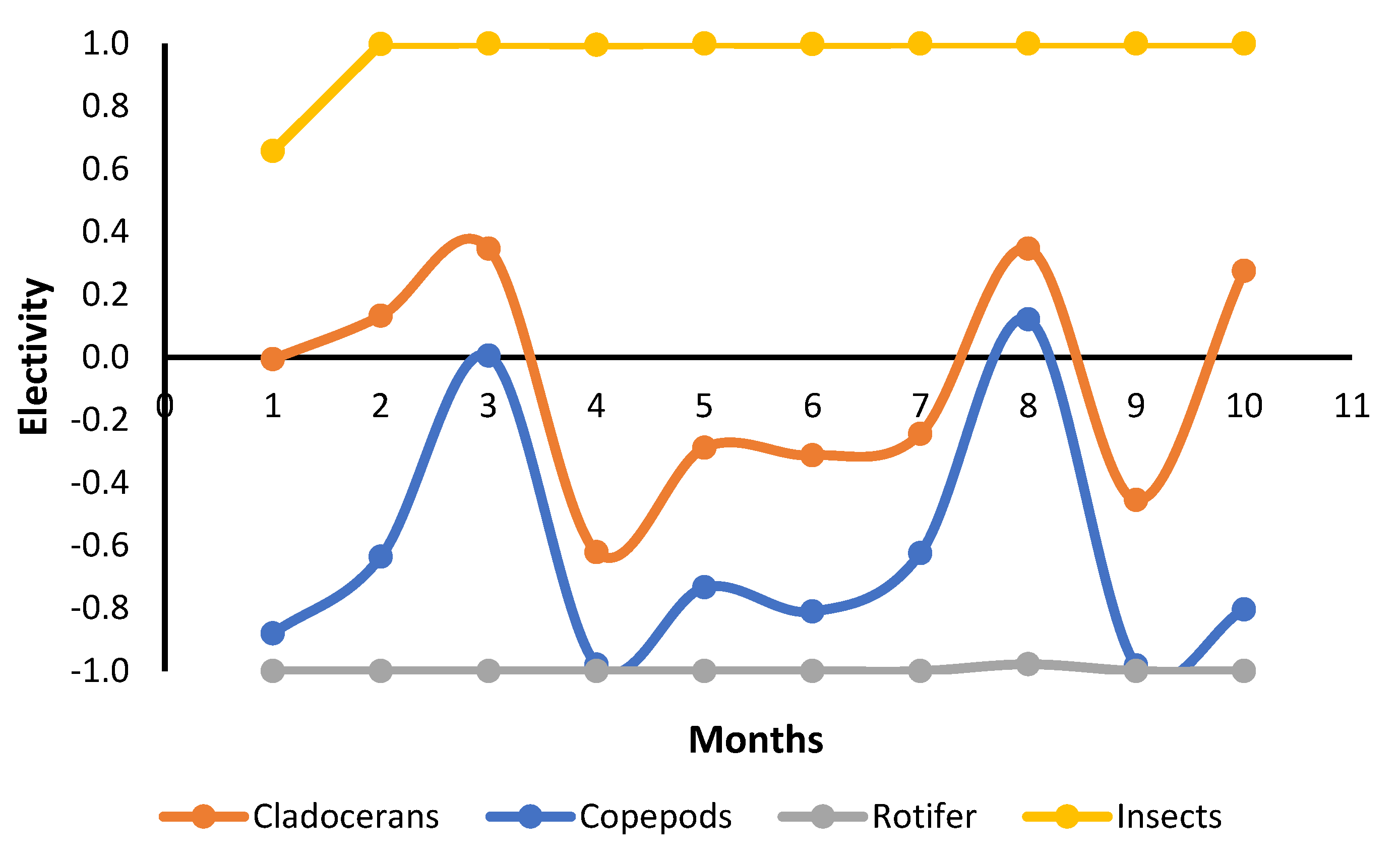
3.3. Stomach Content
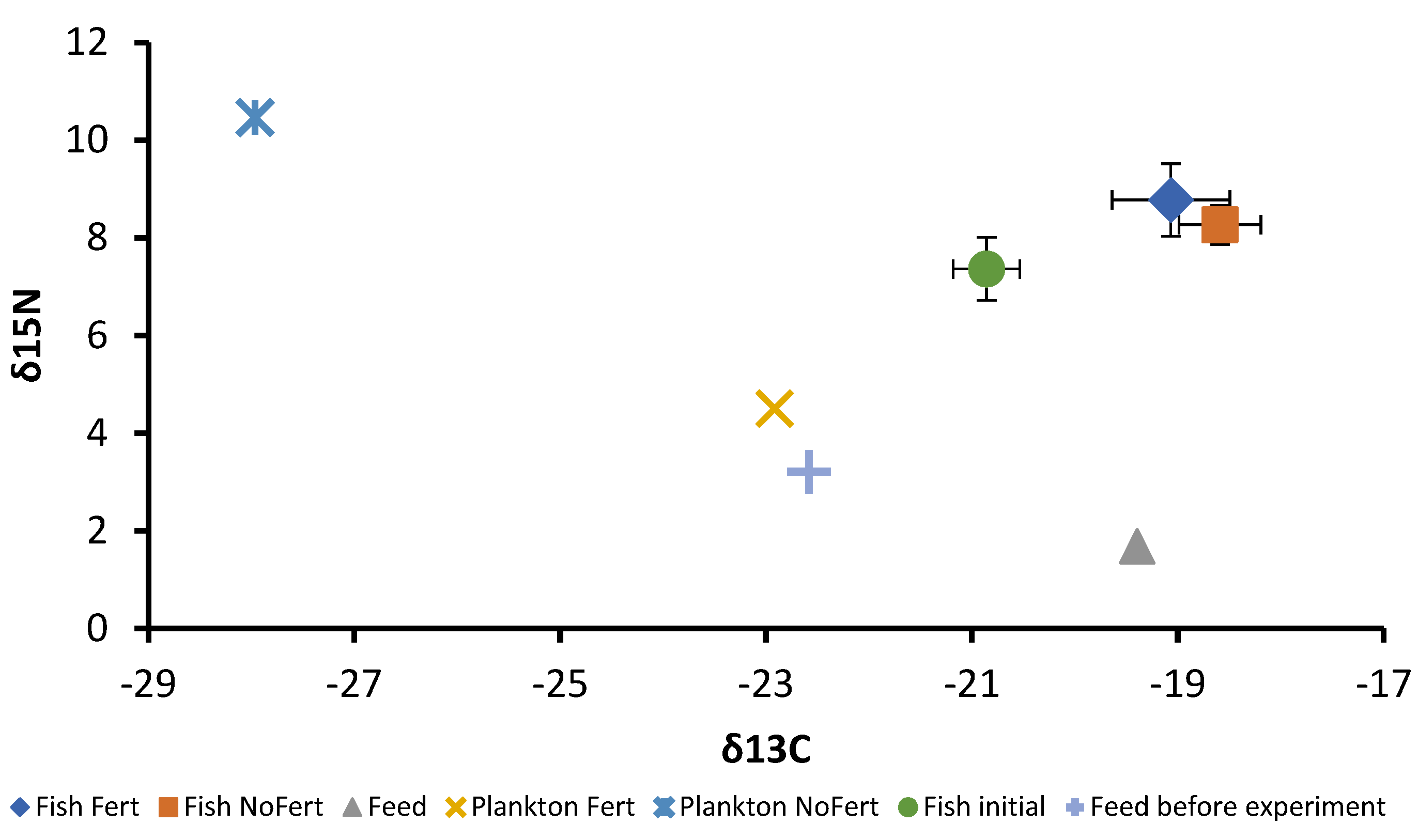
3.4. Stable Carbon and Nitrogen Isotopes as Indicators of Food Assimilation
3.5. Water Quality
3.6. Economic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alleway, H.K.; Waters, T.J.; Brummett, R.; Cai, J.; Cao, L.; Cayten, M.R.; Costa-Pierce, B.A.; Dong, Y.; Hansen, S.C.B.; Liu, S.; et al. Global principles for restorative aquaculture to foster aquaculture practices that benefit the environment. Conserv. Sci. Pract. 2023, 5, e12982. [Google Scholar] [CrossRef]
- Bhakta, J.N.; Bandyopadhyayl, P.K.; Jana, B.B. Effect of different doses of mixed fertilizer on some biogeochemical cycling bacterial population in carp culture pond. Turk. J. Fish. Aquat. Sci. 2006, 6, 165–171. Available online: http://www.trjfas.org/pdf/issue_6_2/165_171.pdf (accessed on 11 August 2021).
- Boyd, C.E.; D‘Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.J.; Teletchea, F.; Tomasso, J.R.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. J. World Aquac. Soc. 2020, 51, 578–633. [Google Scholar] [CrossRef]
- Boyd, C.E. Aquaculture pond fertilization. CAB Rev. 2018, 13, 1–12. [Google Scholar] [CrossRef]
- Garg, S.K.; Bhatnagar, A. Effect of fertilization frequency on pond productivity and fish biomass in still water ponds stocked with Cirrhinus mrigala (Ham.). Aquac. Res. 2000, 31, 409–414. [Google Scholar] [CrossRef]
- Garg, S.K.; Bhatnagar, A. Effect of different doses of organic fertilizer (cow dung) on pond productivity and fish biomass in stillwater ponds. J. Appl. Ichthyol. 1999, 15, 10–18. [Google Scholar] [CrossRef]
- Bhakta, J.N.; Sarkar, D.; Jana, S.; Jana, B.B. Optimizing fertilizer dose for rearing stage production of carps under polyculture. Aquaculture 2004, 239, 125–139. [Google Scholar] [CrossRef]
- Tabinda, A.B.; Ayub, M. Effect of high phosphate fertilization rate on pond phosphate concentrations, chlorophyll a, and fish growth in carp polyculture. Aquac. Int. 2010, 18, 285–301. [Google Scholar] [CrossRef]
- Kang’Ombe, J.; Brown, J.A.; Halfyard, L.C. Effect of using different types of organic animal manure on plankton abundance, and on growth and survival of Tilapia rendalli (Boulenger) in ponds. Aquac. Res. 2006, 37, 1360–1371. [Google Scholar] [CrossRef]
- Rahman, M.M.; Nagelkerke, L.A.J.; Verdegem, M.C.J.; Wahab, M.A.; Verreth, J.A.J. Relationships among water quality, food resources, fish diet and fish growth in polyculture ponds: A multivariate approach. Aquaculture 2008, 275, 108–115. [Google Scholar] [CrossRef]
- Milstein, A.; Azim, M.E.; Wahab, M.A.; Verdegem, M.C.J. The effects of periphyton, fish and fertilizer dose on biological processes affecting water quality in earthen fish ponds. Environ. Biol. Fishes 2003, 68, 247–260. [Google Scholar] [CrossRef]
- Woynárovich, A.; Van Anrooy, R. Field Guide to the Culture of Tambaqui Colossoma macropomum, Cuvier, 1816. In FAO Fisheries and Aquaculture Technical Paper; FAO: Rome, Italy, 2019; Volume 624, Available online: https://books.google.com.br/books?id=WbKIDwAAQBAJ&printsec=frontcover&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false (accessed on 11 November 2022).
- Azim, M.E.; Verdegem, M.C.J.; Khatoon, H.; Wahab, M.A.; Van Dam, A.A.; Beveridge, M.C.M. A comparison of fertilization, feeding and three periphyton substrates for increasing fish production in freshwater pond aquaculture in Bangladesh. Aquaculture 2002, 212, 227–243. [Google Scholar] [CrossRef]
- Azim, M.E.; Little, D.C. Intensifying aquaculture production through new approaches to manipulating natural food. CAB Rev. 2007, 1, 23. [Google Scholar] [CrossRef]
- Cremer, M.C.; Smitherman, R.O. Food habits and growth of silver and bighead carp in cages and ponds. Aquaculture 1980, 20, 57–64. [Google Scholar] [CrossRef]
- Spataru, P.; Wohlfarth, G.W.; Hulata, G. Studies on the natural food of different fish species in intensively manured polyculture ponds. Aquaculture 1983, 35, 283–298. [Google Scholar] [CrossRef]
- Chakrabarti, R.; Jana, B.B. Plankton intake as a function of body weight by common carp fry in different feeding conditions. Aquaculture 1991, 93, 21–34. [Google Scholar] [CrossRef]
- Sipauba-Tavares, L.H.; Braga, F.M.D.S. Study on feeding habits of Piaractus mesopotamicus (Pacu) larvae in fish ponds. Naga ICLARM Q. 1995, 22, 24–29. [Google Scholar]
- Mai, M.G.; Zaniboni-Filho, E. The effect of storage age in external tanks in the larvicultura performance of Salminus brasiliensis (Osteichthyes, Characidae). Acta Sci. Anim. Sci. 2005, 27, 287–296. [Google Scholar]
- Sipaúba-Tavares, L.H.; Braga, F.M.S. The feeding activity of Colossoma macropomum larvae (tambaqui) in fishponds with water hyacinth (Eichhornia crassipesEichhornia crassipes) fertilizer. Braz. J. Biol. 2007, 67, 459–466. [Google Scholar] [CrossRef]
- Ribeiro, D.F.O.; Oliveira Nuñer, A.P. Feed preferences of Salminus brasiliensis (Pisces, Characidae) larvae in fish ponds. Aquaculture 2008, 274, 65–71. [Google Scholar] [CrossRef]
- Gomes, L.C.; Silva, C.R. Impact of pond management on tambaqui, Colossoma macropomum (Cuvier), production during growth-out phase. Aquac. Res. 2009, 40, 825–832. [Google Scholar] [CrossRef]
- Castelo Branco, M.A.; Arruda, M.A.; Gamito, S. Feeding habits of Solea senegalensis in earthen ponds in Sado estuary. J. Sea Res. 2010, 64, 446–450. [Google Scholar] [CrossRef]
- Aya, F.A.; Corpuz, M.N.C.; Garcia, L.M.B. Diet composition, feed preferences and mouth morphology of early stage silver therapon (Leiopotherapon plumbeus, Kner 1864) larvae reared in outdoor tanks. J. Appl. Ichthyol. 2015, 31, 77–82. [Google Scholar] [CrossRef]
- Lima, A.F.; Tavares-Filho, A.; Moro, G.V. Natural food intake by juvenile Arapaima gigas during the grow-out phase in earthen ponds. Aquac. Res. 2018, 49, 2051–2058. [Google Scholar] [CrossRef]
- Cyrus, M.D.; Bolton, J.J.; Macey, B.M. The use of stable isotope ratios δ13C and δ15N to track the incorporation of Ulva and other important dietary ingredients into the gonads of the sea urchin Tripneustes gratilla. Aquac. Nutr. 2020, 26, 174–185. [Google Scholar] [CrossRef]
- Asano, Y.; Hayashizaki, K.I.; Eda, H.; Khonglaliang, T.; Kurokura, H. Natural foods utilized by Nile tilapia, Oreochromis niloticus, in fertilizer-based fish ponds in Lao PDR identified through stable isotope analysis. Fish. Sci. 2010, 76, 811–817. [Google Scholar] [CrossRef]
- Filbrun Jesse, E.; Reynolds, C.A.; Culver, D.A. Effects of feeding rate on habitat quality in fish rearing ponds. J. World Aquac. Soc. 2013, 44, 198–209. [Google Scholar] [CrossRef]
- Abimorad, E.G.; Ducatti, C.; Castellani, D.; Jomori, R.K.; Portella, M.C.; Sartori, M.M.P.; Carneiro, D.J. Dietary lysine deficiency increases the incorporation rate of nitrogen-free compounds in muscle of pacu (Piaractus mesopotamicus). Aquaculture 2017, 479, 522–527. [Google Scholar] [CrossRef]
- Narimbi, J.; Mazumder, D.; Sammut, J. Stable isotope analysis to quantify contributions of supplementary feed in Nile Tilapia Oreochromis niloticus (GIFT strain) aquaculture. Aquac. Res. 2018, 49, 1866–1874. [Google Scholar] [CrossRef]
- Zhou, H.; Gu, B. Using stable isotope analysis to assess the relationship among dietary protein sources, growth, nutrient turnover and incorporation in Nile tilapia (Oreochromis niloticus). Aquac. Nutr. 2020, 26, 1443–1452. [Google Scholar] [CrossRef]
- Hilsdorf, A.W.S.; Hallerman, E.; Valladão, G.M.R.; Zaminhan-Hassemer, M.; Hashimoto, D.T.; Dairiki, J.K.; Takahashi, L.S.; Corrêa Albergaria, F.C.; Gomes, M.E.S.; Venturieri, R.L.L.; et al. The farming and husbandry of Colossoma macropomum: From Amazonian waters to sustainable production. Rev. Aquac. 2022, 14, 993–1027. [Google Scholar] [CrossRef]
- Goulding, M.; Carvalho, M. Life history and management of the tambaqui (Colossoma macropomum, Characidae): An important Amazonian food fish. Rev. Bras. Zool. 1982, 1, 107–133. [Google Scholar] [CrossRef]
- Da Silva, J.A.M.; Pereira-Filho, M.; de Oliveira-Pereira, M.I. Seasonal variation of nutrients and energy in tambaqui’s (Colossoma macropomum Cuvier, 1818) natural food. Braz. J. Biol. 2000, 60, 599–605. [Google Scholar] [CrossRef]
- FAO. Fishery and Aquaculture Statistics—Yearbook 2020. In FAO Yearbook of Fishery and Aquaculture Statistics; FAO: Rome, Italy, 2023. [Google Scholar] [CrossRef]
- Valladão, G.M.R.; Gallani, S.U.; Pilarski, F. South American fish for continental aquaculture. Rev. Aquac. 2018, 10, 351–369. [Google Scholar] [CrossRef]
- Valenti, W.C.; Barros, H.P.; Moraes-Valenti, P.; Bueno, G.W.; Cavalli, R.O. Aquaculture in Brazil: Past, present and future. Aquac. Rep. 2021, 19, 100611. [Google Scholar] [CrossRef]
- Roubach, R.; Gomes, L.C.; Leão Fonseca, F.A.; Val, A.L. Eugenol as an efficacious anaesthetic for tambaqui, Colossoma macropomum (Cuvier). Aquac. Res. 2005, 36, 1056–1061. [Google Scholar] [CrossRef]
- Hyslop, E.J. Stomach contents analysis—A review of methods and their application. J. Fish Biol. 1980, 17, 411–429. [Google Scholar] [CrossRef]
- Amundsen, P.-A.; Sánchez-Hernández, J. Feeding studies take guts—Critical review and recommendations of methods for stomach contents analysis in fish. J. Fish Biol. 2019, 95, 1364–1373. [Google Scholar] [CrossRef]
- Ivlev, V.W. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961; p. 302. [Google Scholar]
- Coplen, T.B. Guidelines and recommended terms for expression of stable-isotope-ratio and gas-ratio measurement results. Rapid Commun. Mass Spectrom. 2011, 25, 2538–2560. [Google Scholar] [CrossRef]
- Brand, W.A.; Coplen, T.B. Stable isotope deltas: Tiny, yet robust signatures in nature. Isot. Environ. Health Stud. 2012, 48, 393–409. [Google Scholar] [CrossRef]
- Paul, D.; Skrzypek, G.; Fórizs, I. Normalization of measured stable isotopic compositions to isotope reference scales—A review. Rapid Commun. Mass Spectrom 2007, 21, 3006–3014. [Google Scholar] [CrossRef]
- Schimmelmann, A.; Qi, H.; Coplen, T.B.; Brand, W.A.; Fong, J.; Meier-Augenstein, W.; Kemp, H.F.; Toman, B.; Ackermann, A.; Assonov, S.; et al. Organic reference materials for hydrogen, carbon, and nitrogen stable isotope-ratio measurements: Caffeines, n-alkanes, fatty acid methyl esters, glycines, l-valines, polyethylenes, and oils. Anal. Chem. 2016, 88, 4294–4302. [Google Scholar] [CrossRef]
- Britton, J.R.; Busst, G.M.A. Stable isotope discrimination factors of omnivorous fishes: Influence of tissue type, temperature, diet composition and formulated feeds. Hydrobiologia 2018, 808, 219–234. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Waste Water, 21st ed.; American Public Health Association: Washington, DC, USA, 2017. [Google Scholar]
- Engle, C.R. Aquaculture Economics and Financing: Management and Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Veverica, K.; Bowman, J.; Gichuri, W.; Izaru, P.; Mwau, P.; Popma, T. Relative Contribution of supplemental feed and inorganic fertilizers in semi-intensive tilapia production. In Seventeenth Annual Technical Report; McElwee, K., Burke, D., Niles, M., Cummings, X., Egna, H., Eds.; Pond Dynamics/Aquaculture CRSP, Oregon State University: Corvallis, OR, USA, 2000; pp. 23–28. [Google Scholar]
- Liranço, A.D.D.S.; Romagosa, E.; Scorvo-Filho, J.D. Growth performance of Pseudoplatystoma corruscans stocked in rearing systems: Semi-intensive (ponds) and intensive (cages). Ciência Rural. 2011, 41, 524–530. Available online: http://www.scielo.br/scielo.php?pid=S0103-84782011000300027&script=sci_arttext&tlng=es (accessed on 9 March 2018). [CrossRef]
- Box, G.E.P.; Cox, D.R. An analysis of transformation. J. R. Stat. Soc. 1964, 26, 211–243. [Google Scholar] [CrossRef]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: 2021, Viena, Austria. Available online: https://www.r-project.org/ (accessed on 15 June 2023).
- Filbrun, J.E.; Culver, D.A. Stable isotopes reveal live prey support growth of juvenile channel catfish reared under intensive feeding regimens in ponds. Aquaculture 2014, 433, 125–132. [Google Scholar] [CrossRef]
- Jomori, R.K.; Ducatti, C.; Carneiro, D.J.; Portella, M.C. Stable carbon (δ13C) and nitrogen (δ15N) isotopes as natural indicators of live and dry food in Piaractus mesopotamicus (Holmberg, 1887) larval tissue. Aquac. Res. 2008, 39, 370–381. [Google Scholar] [CrossRef]
- Lima, A.F.; Rodrigues, A.P.O.; Costa, V.E. Frozen zooplankton is efficient as natural food during pirarucu Arapaima gigas weaning. Aquac. Res. 2021, 52, 4227–4236. [Google Scholar] [CrossRef]
- Oliveira, A.C.B.; Martinelli, L.A.; Moreira, M.Z.; Soares, M.G.M.; Cyrino, J.E.P. Seasonality of energy sources of Colossoma macropomum in a floodplain lake in the Amazon—Lake Camaleão, Amazonas, Brazil. Fish. Manag. Ecol. 2006, 13, 135–142. [Google Scholar] [CrossRef]
- Bateman, A.S.; Kelly, S.D. Fertilizer nitrogen isotope signatures. Isot. Environ. Health Stud. 2007, 43, 237–247. [Google Scholar] [CrossRef]
- Lomartire, S.; Marques, J.C.; Gonçalves, A.M. The key role of zooplankton in ecosystem services: A perspective of interaction between zooplankton and fish recruitment. Ecol. Indic. 2021, 129, 107867. [Google Scholar] [CrossRef]
- Brucet, S.; Boix, D.; Quintana, X.D.; Jensen, E.; Nathansen, L.W.; Trochine, C.; Meerhoff, M.; Gascón, S.; Jeppesena, E. Factors influencing zooplankton size structure at contrasting temperatures in coastal shallow lakes: Implications for effects of climate change. Limnol. Oceanogr. 2010, 55, 1697–1711. [Google Scholar] [CrossRef]
- Vakkilainen, K.; Kairesalo, T.; Hietala, J.; Balayla, D.M.; Becares, E.; Van De BUnd, W.J.; Van Donk, E.; Fernandez-Alaez, M.; Gyllstrom, M.; Hansson, L.-A.; et al. Response of zooplankton to nutrient enrichment and fish in shallow lakes: A pan-European mesocosm experiment. Freshw. Biol. 2004, 49, 1619–1632. [Google Scholar] [CrossRef]
- Sipaúba-Tavares, L.H. Análise da seletividade alimentar em larvas de tambaqui (Colossoma macropomum) e tambacu (híbrido, pacu Piaractus mesopotamicus e tambaqui Colossoma macropomum) sobre os organismos zooplanctônicos. Acta Limnol. Bras. 1993, 6, 114–132. [Google Scholar]
- Lima, A.F.; Pereira, A.S.; de Oliveira Costa-Fernandes, T.; Rodrigues, A.P.O.; Costa, V.E.; Maciel-Honda, P.O. The effect of nursery production system (in cage and pond) on performance, health status, and plankton ingestion of the low trophic level fish tambaqui, Colossoma macropomum. Aquaculture 2024, 586, 740721. [Google Scholar] [CrossRef]
- Chary, K.; van Riel, A.; Muscat, A.; Wilfart, A.; Harchaoui, S.; Verdegem, M.; Filgueira, R.; Troell, M.; Henriksson, P.J.G.; de Boer, I.J.M.; et al. Transforming sustainable aquaculture by applying circularity principles. Rev. Aquac. 2023, 16, 656–673. [Google Scholar] [CrossRef]
- Aubin, J.; Callier, M.; Rey-Valette, H.; Mathé, S.; Wilfart, A.; Legendre, M.; Slembrouck, J.; Caruso, D.; Chia, E.; Masson, G.; et al. Implementing ecological intensification in fish farming: Definition and principles from contrasting experiences. Rev. Aquac. 2019, 11, 149–167. [Google Scholar] [CrossRef]
- Watanabe, T.; Kiron, V. Prospects in larval fish dietetics. Aquaculture 1994, 124, 223–251. [Google Scholar] [CrossRef]
- Bakke, A.M.; Glover, C.; Krogdahl, Å. Feeding, digestion and absorption of nutrients. Fish Physiol. 2010, 30, 57–110. [Google Scholar] [CrossRef]
- Mitra, G.; Mukhopadhyay, P.K.; Ayyappan, S. Biochemical composition of zooplankton community grown in freshwater earthen ponds: Nutritional implication in nursery rearing of fish larvae and early juveniles. Aquaculture 2007, 272, 346–360. [Google Scholar] [CrossRef]
- da Silva Reis, Y.; Leite, J.L.R.; de Almeida, C.A.L.; Pereira, D.S.P.; Vidal, L.V.O.; de Araujo, F.G.; Fortes-Silva, R. New insights into tambaqui (Colossoma macropomum) feeding behavior and digestive physiology by the self-feeding approach: Effects on growth, dial patterns of food digestibility, amylase activity and gastrointestinal transit time. Aquaculture 2019, 498, 116–122. [Google Scholar] [CrossRef]
- Dias-Koberstein, T.C.R.; Carneiro, D.J.; Urbinati, E.C. Gastrointestinal passage time length and gastric emptying of pacu (Piaractus mesopotamicus) in different temperatures. Acta Scientiarum Anim. Sci. 2005, 27, 413–417. [Google Scholar]
- Garg, S.K.; Bhatnagar, A. Effect of varying closes of organic and inorganic fertilizers on plankton production and fish biomass in brackish water fish ponds. Aquac. Res. 1996, 27, 157–166. [Google Scholar] [CrossRef]
- Rezende, F.P.; Lima, A.F. Effect of pond fertilization on growth performance of pirarucu during grow-out phase. Lat. Am. J. Aquat. Res. 2022, 50, 22–30. [Google Scholar] [CrossRef]
- Duodu, C.; Boateng, D.; Edziyie, R. Effect of pond fertilization on productivity of tilapia pond culture in Ghana. J. Fish. Coast. Manag. 2020, 2, 56–64. [Google Scholar] [CrossRef]
- Boyd, C.E. Water Quality: An Introduction; Springer: Cham, Switzerland, 2020; 440p. [Google Scholar] [CrossRef]
- INMET. INMET Tempo. 2021. Available online: https://tempo.inmet.gov.br/ (accessed on 19 October 2021).
- Boyd, C.E.; Ammonia Nitrogen Dynamics in Aquaculture. Global Aquaculture Advocate. Available online: https://www.aquaculturealliance.org/advocate/ammonia-nitrogen-dynamics-in-aquaculture/ (accessed on 8 October 2018).
- Das, S.K.; Jana, B.B. Pond fertilization regimen: State-of-the-art. J. Appl. Aquac. 2003, 13, 35–66. [Google Scholar] [CrossRef]
- Flickinger, D.L.; Costa, G.A.; Dantas, D.P.; Moraes-Valenti, P.; Valenti, W.C. The budget of nitrogen in the grow-out of the Amazon River prawn (Macrobrachium amazonicum Heller) and tambaqui (Colossoma macropomum Cuvier) farmed in monoculture and in integrated multitrophic aquaculture systems. Aquac. Res. 2019, 50, 3444–3461. [Google Scholar] [CrossRef]
- Flickinger, D.L.; Costa, G.A.; Dantas, D.P.; Proença, D.C.; David, F.S.; Durborow, R.M.; Moraes-Valenti, P.; Valenti, W.C. The budget of carbon in the farming of the Amazon river prawn and tambaqui fish in earthen pond monoculture and integrated multitrophic systems. Aquac. Rep. 2020, 17, 100340. [Google Scholar] [CrossRef]
- Flickinger, D.L.; Dantas, D.P.; Proença, D.C.; David, F.S.; Valenti, W.C. Phosphorus in the culture of the Amazon River prawn (Macrobrachium amazonicum) and tambaqui (Colossoma macropomum) farmed in monoculture and in integrated multitrophic systems. J. World Aquac. Soc. 2020, 51, 1002–1023. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fish Weight (g) | Crude Protein (%) | Gross Energy (cal 100 g−1) | Ether Extract (%) | Crude Fiber (%) | Pellet Size (mm) | Feeding Rate (% of Body Weight/Day) |
|---|---|---|---|---|---|---|
| 60–200 | 34.4 | 336.5 | 4.1 | 7.1 | 4 | 4.5 |
| 200–500 | 36.1 | 337.7 | 4.1 | 6.8 | 6 | 3.5 |
| 500–700 | 36.1 | 337.7 | 4.1 | 6.8 | 6 | 2.5 |
| 700< | 30.3 | 337.4 | 3.4 | 6.7 | 10 | 2.5 |
| Items | Value (US$) | Unit |
|---|---|---|
| Formulated feed (4 mm) | 0.49 | US$/kg |
| Formulated feed (6 mm) | 0.49 | US$/kg |
| Formulated feed (10 mm) | 0.43 | US$/kg |
| Urea | 0.73 | US$/kg |
| Triple superphosphate | 0.30 | US$/kg |
| Rice bran | 0.18 | US$/kg |
| Labor | 1.27 | h |
| Parameters | Treatment | p Value | |
|---|---|---|---|
| Fert | NoFert | ||
| Final mean weight (g) | 1697.5 ± 88.8 a | 1628.4 ± 63.8 a | 0.2525 |
| SGR (% day−1) | 0.95 ± 0.02 a | 0.94 ± 0.01 a | 0.3903 |
| Daily feed intake (% day−1) | 1.16 ± 0.02 b | 1.25 ± 0.05 a | 0.0077 |
| Feed conversion rate | 2.04 ± 0.04 b | 2.22 ± 0.08 a | 0.0090 |
| Fish yield (t/ha/cycle) | 6.2 ± 0.3 a | 6.1 ± 0.2 a | 0.7552 |
| Survival (%) | 91.6 ± 5.6 a | 94.4 ± 1.6 a | 0.3701 |
| Food Items | FM | DIP (Individuals/L) | FO (%) | RA (%) |
|---|---|---|---|---|
| Cladocerans | Fert | 77 ± 129 | 42 ± 35 | 10 ± 13 a |
| NoFert | 39 ± 34 | 29 ± 35 | 6 ± 12 b | |
| Copepods | Fert | 458 ± 493 | 25 ± 29 | 4 ± 7 |
| NoFert | 246 ± 224 | 17 ± 27 | 4 ± 9 | |
| Rotifers | Fert | 641 ± 1199 | 2 ± 7 | 0 ± 0 |
| NoFert | 361 ± 336 | 0 ± 0 | 0 ± 0 | |
| Phytoplakton | Fert | 360 ± 376 | 6 ± 19 | 1 ± 3 |
| NoFert | 310 ± 367 | 6 ± 22 | 0 ± 2 | |
| Terrestrial Plants | Fert | NQ | 40 ± 32 | 12 ± 16 |
| NoFert | NQ | 41 ± 35 | 14 ± 21 | |
| Insects | Fert | 1 ± 1 | 62 ± 33 | 26 ± 24 |
| NoFert | 1 ± 1 | 65 ± 30 | 26 ± 21 | |
| Digested food | Fert | NQ | 58 ± 36 | 15 ± 13 |
| NoFert | NQ | 57 ± 33 | 16 ± 16 | |
| Sediment | Fert | NQ | 7 ± 15 | 1 ± 2 |
| NoFert | NQ | 11 ± 23 | 1 ± 3 | |
| Feed | Fert | 10 ± 23 | 1 ± 4 | |
| NoFert | - | 9 ± 24 | 1 ± 5 | |
| Ostracods and decapods | Fert | NQ | 10 ± 22 | 2 ± 4 |
| NoFert | NQ | 5 ± 11 | 0 ± 1 |
| Water Parameters | Fertilization Management (FM) | p Values | |||
|---|---|---|---|---|---|
| Fert | NoFert | FM | Time | F × Time | |
| Temperature (°C) | 28.58 ± 0.06 | 28.56 ± 0.05 | 0.5289 | <0.0001 | 0.0001 |
| Transparency (cm) | 66.26 ± 7.86 | 69.3 ± 12.26 | 0.3679 | <0.0001 | 0.0029 |
| Dissolved oxygen (mg L−1) | 4.09 ± 0.31 | 3.61 ± 0.36 | 0.0006 | <0.0001 | 0.0038 |
| pH | 7.26 ± 0.04 a | 7.27 ± 0.09 b | 0.0042 | <0.0001 | 0.8512 |
| TAN (mg L−1) | 0.2725 ± 0.04 | 0.2675 ± 0.02 | 0.8682 | <0.0001 | 0.2613 |
| Phosphorus (mg/L) | 0.41 ± 0.26 | 0.33 ± 0.22 | 0.0424 | <0.0001 | 0.7673 |
| Alkalinity (mg de CaCO3/L) | 20.69 ± 2.52 | 21.12 ± 1.59 | 0.7860 | - | - |
| Hardness (mg de CaCO3/L) | 15.34 ± 1.76 | 16.04 ± 2.02 | 0.6723 | - | - |
| Items | Fert | NoFert |
|---|---|---|
| Fish biomass (kg/600 m2 pond) | 370.61 | 370.61 |
| Feed conversion ratio | 2.04 | 2.22 |
| Total formulated feed (kg) | 756.04 | 822.75 |
| Formulated feed (4 mm) (US$) | 28.47 | 30.98 |
| Formulated feed (6 mm) (US$) | 127.16 | 138.38 |
| Formulated feed (8 mm) (US$) | 180.35 | 196.27 |
| Total feed costs (US$) | 335.99 | 365.63 |
| Total fertilizer costs 1 (R$) | 53.66 | 0 |
| Fertilization management labor costs 1,2 (US$) | 5.85 | 0 |
| Total cost (US$) | 395.49 | 365.63 |
| Cost increment in Fert management (%) | 8.2% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, A.F.; dos Reis, A.G.P.; Costa, V.E.; Valenti, W.C. Natural Food Intake and Its Contribution to Tambaqui Growth in Fertilized and Unfertilized Ponds. Fishes 2024, 9, 139. https://doi.org/10.3390/fishes9040139
Lima AF, dos Reis AGP, Costa VE, Valenti WC. Natural Food Intake and Its Contribution to Tambaqui Growth in Fertilized and Unfertilized Ponds. Fishes. 2024; 9(4):139. https://doi.org/10.3390/fishes9040139
Chicago/Turabian StyleLima, Adriana Ferreira, Anderson Guilherme Pereira dos Reis, Vladimir Eliodoro Costa, and Wagner Cotroni Valenti. 2024. "Natural Food Intake and Its Contribution to Tambaqui Growth in Fertilized and Unfertilized Ponds" Fishes 9, no. 4: 139. https://doi.org/10.3390/fishes9040139
APA StyleLima, A. F., dos Reis, A. G. P., Costa, V. E., & Valenti, W. C. (2024). Natural Food Intake and Its Contribution to Tambaqui Growth in Fertilized and Unfertilized Ponds. Fishes, 9(4), 139. https://doi.org/10.3390/fishes9040139