1. Introduction
Artificial breeding of olive flounder (
Paralichthys olivaceus) in the Republic of Korea (henceforth, Korea) was first started in 1982 [
1]. Since then, this species has become popular in the country, accounting for about 50% (39,931 metric tons) of total aquaculture production (79,651 metric tons) in 2023 [
2,
3]. As a carnivorous fish, olive flounder requires more than 50% protein in the diet [
4,
5]. Therefore, feed is an important part of the farming of this species, as it provides the protein necessary for growth, and it makes up 30–35% of the total operational costs [
3,
6,
7].
Fishmeal (FM) has been the main protein source used in fish feeds [
6,
8,
9] because it is a palatable and high-quality protein source [
10,
11]. Furthermore, FM represents a significant protein source for olive flounder [
3,
6]. However, its cost has steeply increased due to decreases in fish catch commonly used for FM [
12,
13,
14,
15]. Therefore, many studies have attempted to identify appropriate FM replacements [
6,
9,
16,
17,
18,
19].
Soybean meal (0.64 USD/kg) is less expensive than FM (2.15 USD/kg) [
20] but has problems in that it contains non-starch polysaccharides, which are disadvantageous for digestion, and high levels of antinutritional factors such as trypsin inhibitors [
19,
21,
22]. To address these problems, soy protein concentrate (SPC; 1.08 USD/kg), a product of the soybean flake leaching process [
20,
23,
24], could be utilized as a substitute. SPC contains more readily digested amino acids (AAs) and lower levels of antinutritional factors compared to soybean meal [
19]. In a previous study, it was possible to replace up to 30% FM with SPC without AA supplementation in a 60% FM-based diet without compromising the growth of olive flounder for 20 weeks [
25]. However, Deng et al. [
16] reported that SPC is not an effective substitute for FM in this species due to its unbalanced and poor nutritional profile. Generally, as plant protein sources are likely to lack limiting AAs, such as arginine, lysine, and methionine, the substitution of FM in the diet with an alternative plant protein could be improved by supplementation with these AAs. In a 10-week feeding experiment, Li et al. [
19] found that up to 40% FM could be substituted with SPC supplemented with limiting AAs (lysine and methionine) in a 68% FM-based diet without deterioration of growth of starry flounder (
Platichthys stellatus). In addition, up to 50% FM could be replaced with SPC with lysine and methionine supplementation in a 64% FM-based diet without compromising growth performance in the common sole (
Solea solea) [
26]. Nevertheless, the majority of preceding studies examining the replacement of dietary FM with SPC have been conducted over relatively brief periods (≤20 weeks) [
20,
25,
26].
The study of fish maturation is a topic that has been the subject of considerable research [
10,
25,
27]. Maturation hormones, such as follicle-stimulating hormone (FSH), luteinizing hormone (LH), and estrogen receptor (ER), are produced in the brain. FSH regulates gonadal development, and LH mainly regulates gonadal maturation [
28]. ER is taken up by oocytes and used as a nutrient for embryonic development after fertilization [
25,
29]. Vitellogenin (VIT) is synthesized after 17β-estradiol binds to the ER in the liver, is absorbed by oocytes, and is used as a nutrient for embryonic development after fertilization [
25]. However, there have been few reports regarding the maturation of fish associated with dietary FM replacement with alternative sources [
25,
27]. Furthermore, most previous studies of dietary FM replacement with alternative proteins correlated with maturation have been performed for short periods (≤16 weeks) [
15,
16,
27]. However, the effects of a low-FM diet on gonad maturation in fish should be confirmed in long-term feeding trials (≥16 weeks) [
25].
The objective of this study was to perform a long-term (12 months) trial to investigate the effects of the replacement of FM with SPC in the diet supplemented with limiting AAs on the gonadal development and growth of the olive flounder.
2. Materials and Methods
2.1. Experimental Conditions and Fish
Olive flounder (mean ± SE initial body weight 727.76 ± 7.89 g) were transferred from the Genetics and Breeding Research Center (National Fisheries Research Institute, Geoje-si, Gyeongsangnam-do, Republic of Korea) to the Feed Research Center (National Fisheries Research Institute, Pohang-si, Gyeongsangbuk-do, Republic of Korea). The fish were allowed to acclimate to the experimental conditions for 2 weeks and fed the control (Con) diet (crude protein: 53.85%, crude lipid: 9.25%). After this acclimatization period, 900 experimental fish were randomly transferred into nine 8-ton flow-through tanks (water volume: 5 tons) (100 fish/tank; n = 3 tanks per treatment). The water source was sand-filtered seawater. Water temperature ranged from 10.83 °C to 23.22 °C (mean ± SD 16.53 ± 5.61 °C), and dissolved oxygen (8.52 ± 1.00 mg/L; mean ± SD) was directly supplied to each tank through an oxygen generator. Throughout the 12-month feeding trial, dead fish were removed immediately from each tank, and their body length and weight were determined.
2.2. Experimental Diet
Three isonitrogenous (53.00%) and isoenergetic (3.93 kcal/g diet) experimental diets were formulated (
Table 1). The Con diet consisted of 60% low-temperature FM, while the LF1 and LF2 diets were formulated to match the FM content of the Con diet, with 25% and 50% FM replaced with SPC supplemented with lysine (LF1; 0.38%, LF2; 0.75%) and methionine (LF1; 0.15%, LF2; 0.29%), respectively (MP Biomedicals, Irvine, CA, USA). All feeds fulfilled the dietary nutrient requirements of olive flounder [
5,
30].
The ingredients of the experimental diets were stored in a freezer at −20 °C until use, after which they were thoroughly blended with water at a ratio of 5:1. The pressure control of the experimental feeds was dependent on the screw speed (rpm/min), barrel temperature (115–130 °C), conditioner temperature (80 °C), and steam (31.6 kg/h). The feed rate (50 kg/h) of the extruder (ATX-II twin screw extruder; Fesco, Dangu, Republic of Korea) was maintained, with the pressure set at low (885 rpm/min) and high (708 rpm/min) levels. The speeds of the discs (rpm/min) and rotors (rotor, rpm/min) of the air classifier mill (XP 50; KMTECH, Guri-si, Gyeonggi-do, Republic of Korea) were adjusted as appropriate to ensure optimal performance. All experimental diets were extruded as pellets on a biweekly basis and stored in a freezer at −20 °C until use. The fish were fed the experimental diets twice a day (08:00 and 17:00) until they appeared satiated for a period of 12 months.
2.3. Measurement of Growth and Biological Indices
All live fish from each tank were subjected to a 24 h starvation period and subsequently anesthetized using 50 ppm 2-phenoxyethanol (Sigma, St. Louis, MO, USA) prior to measurement. The number of live fish and their weight were recorded for each tank, enabling the calculation of survival and weight gain. To ascertain the biological indices (CF, VSI, HSI, and GSI), six fish were randomly selected from each tank at the outset, after four months, and at the conclusion of the 12-month experimental period. These samples were utilized to calculate the viscerosomatic index (VSI), hepatosomatic index (HSI), and gonadosomatic index (GSI). The biological indices were calculated using the following equations:
2.4. Biochemical Composition of the Experimental Diets and Proximate Composition of the Fish
The main ingredients (FM and SPC) and the experimental diets were homogenized, and the biochemical compositions were determined both before and after the production of the diets. Prior to sampling, all live fish from each tank were deprived of food for a period of 24 h. To ascertain the biochemical composition, six fish from each tank were randomly selected at the outset, after four months, and at the conclusion of the 12-month experimental period. Following homogenization, the biochemical compositions were determined. The proximate composition of the samples was analyzed in accordance with the AOAC standard method [
32]. The crude protein and crude lipid contents were determined using the Kjeldahl method (Kjeltec 2100 Distillation Unit; Foss Tecator, Hillerød, Denmark) and an ether extraction method (Soxtec TM 2043 Fat Extraction System; Foss Tecator, Hillerød, Denmark), respectively. The moisture content was determined by oven drying for 6 and 24 h at 105 °C, respectively, for the analysis of the main ingredients and experimental diets and for the fish samples. The ash content was determined by placing the samples in a muffle furnace at 550 °C for 4 h.
The AA profiles of the experimental diets and the dorsal muscle of olive flounder were hydrolyzed by the addition of 20 mL of 6 N hydrochloric acid (HCl) at 110 °C for a period of 24 h in a dry oven. The solution was filtered through a glass filter and concentrated under vacuum at 55 °C to completely evaporate the hydrochloric acid and water. The sample was dissolved with sodium citrate buffer (pH 2.20) in a 25 mL flask, filtered with a 0.45 µm membrane filter, and analyzed using an automated amino acid analyzer (Biochrom 30+; Biochrom Ltd., Cambridge, UK).
The total lipids of the experimental diets and dorsal muscle were extracted with a chloroform and methanol mixture (2:1) to methylate the fatty acids (FAs) with a 14% BF-methanol (Sigma-Aldrich, St. Louis, MO, USA) solution. The resulting FAs were then analyzed. The samples were analyzed by gas chromatography (Trace 1310; Thermo Scientific, Waltham, MA, USA) with a capillary column (SPTM-2560, 100 m × 0.25 mm; Supelco Inc., Bellefonte, PA, USA) as the detector. A mixture of 37 fatty acids (PUFA 37 Component FAME Mix; Supelco Inc.) was employed as the standard for fatty acid analysis. The methods and procedures utilized for the analysis of the samples were consistent with those previously described by [
8].
2.5. Histology of Male and Female Gonads
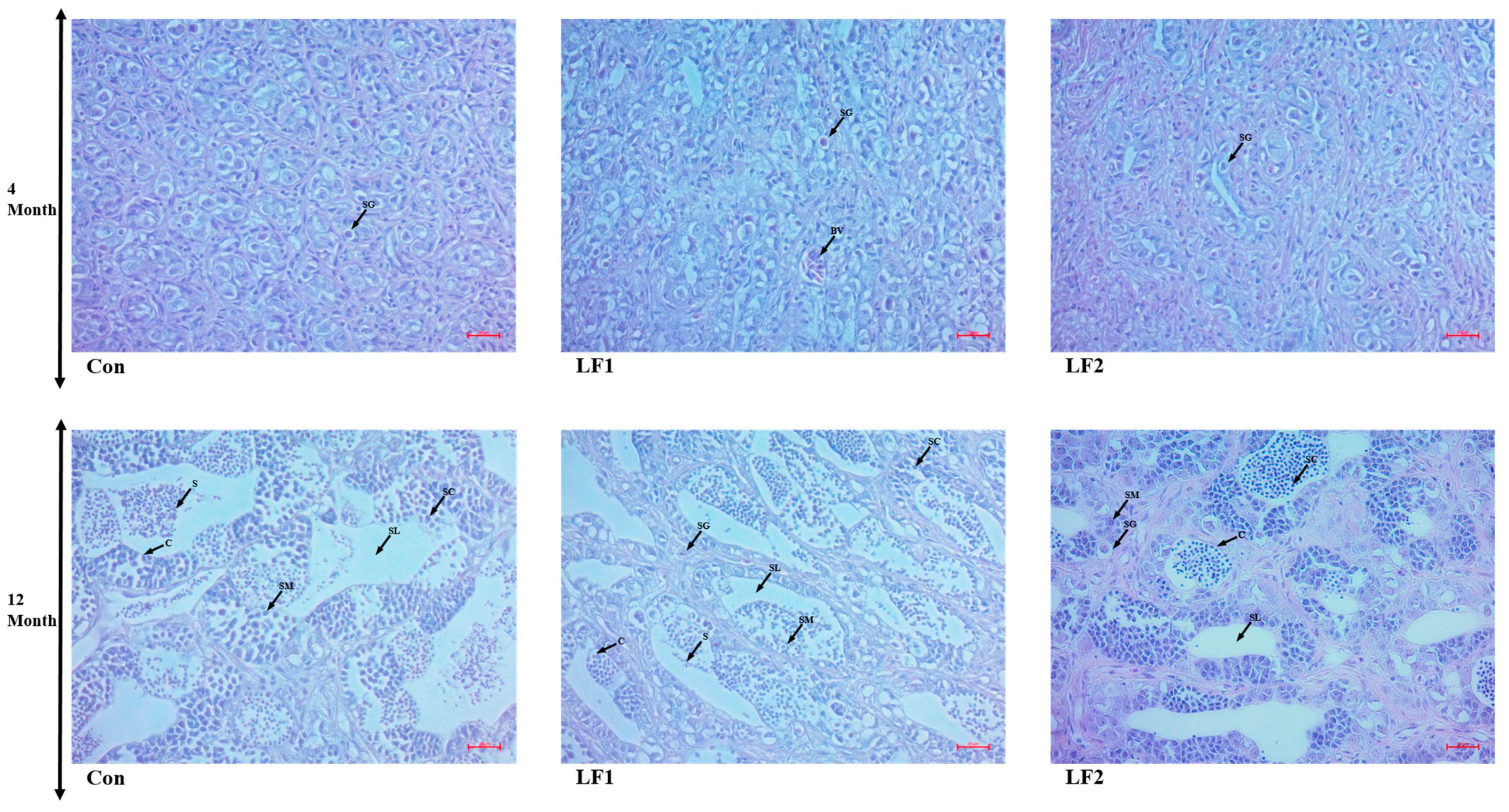
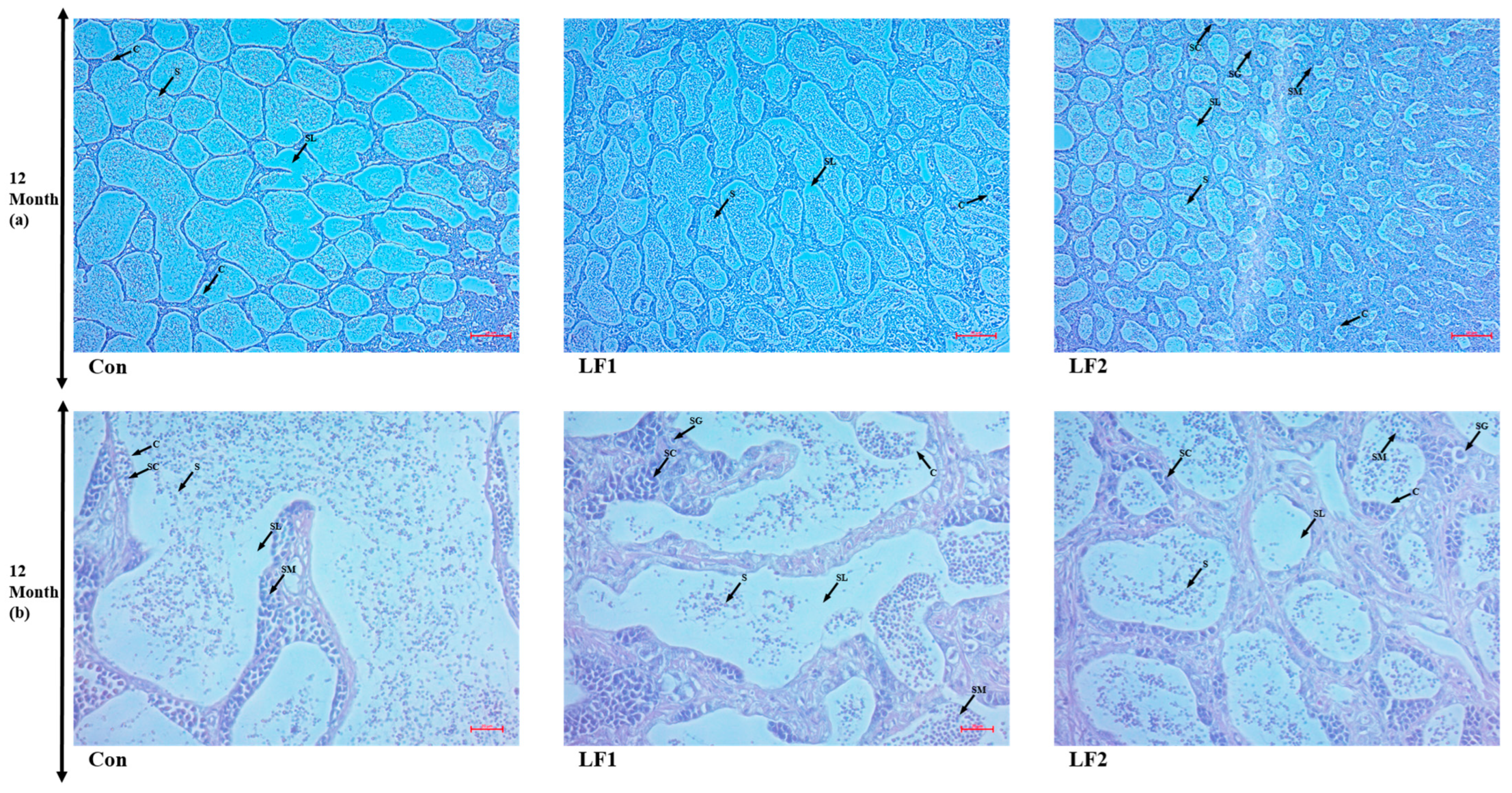
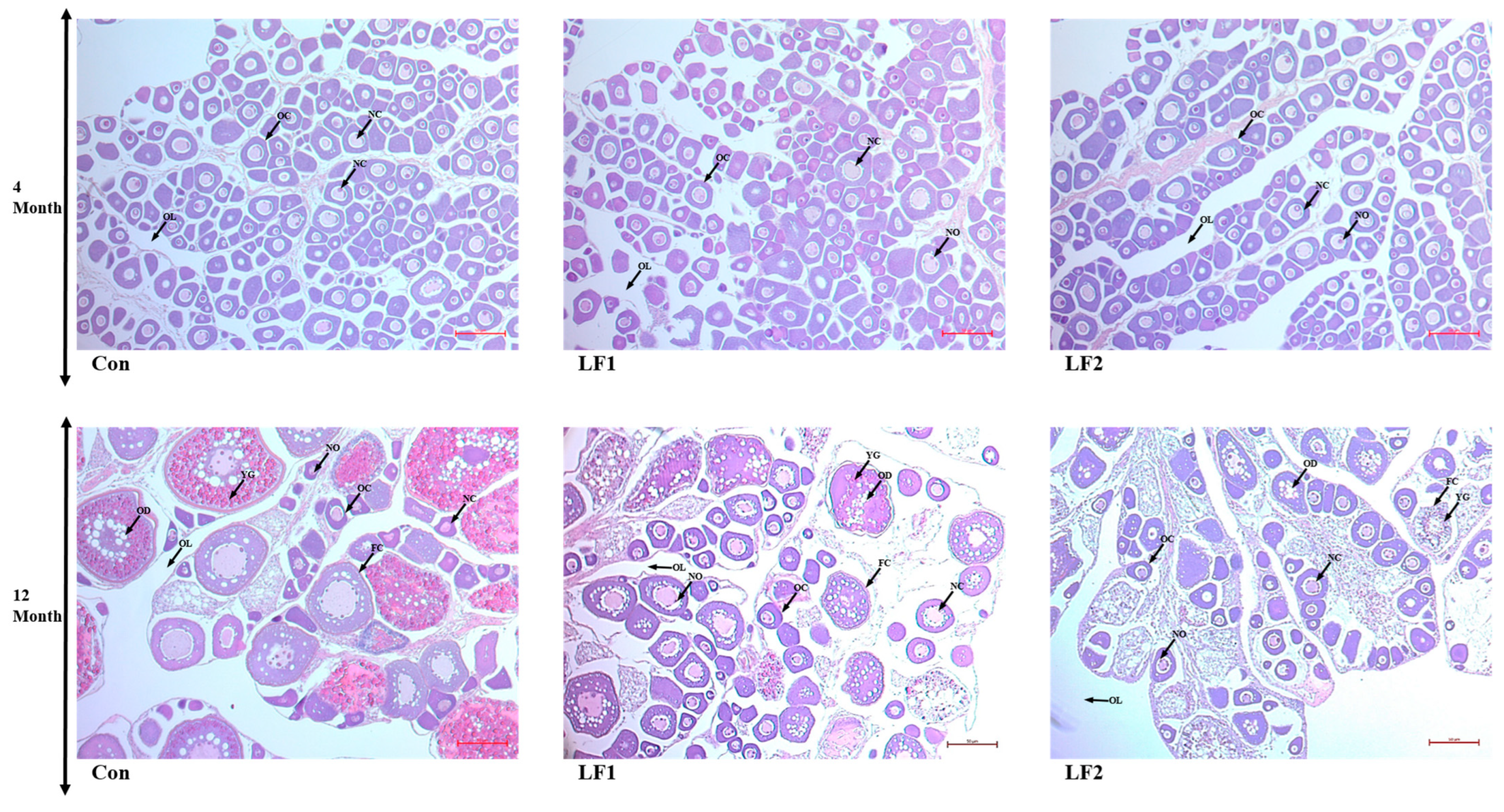
To examine gonadal development, male and female gonads were sampled at 4 and 12 months and fixed in 10% formalin (Sigma) for 24 h. The solution was exchanged twice, and the specimens were stored in 70% ethanol. The fixed tissues were embedded in paraffin to prepare tissue blocks for histological analysis. The tissues were cut into sections 4 µm thick, placed on glass slides, and deparaffinized using xylene. The gonadal tissues were stained with hematoxylin and eosin (BBC Ciochemical, Mount Vernon, WA, USA), and male and female gonads were distinguished. In males, six testes were randomly selected for each group and examined at 100× and 400× magnification (Carl Zeiss, Göttingen, Germany). In females, six ovaries were randomly selected for each group and examined at 200× magnification (Carl Zeiss). The sections were observed and photographed using Zen program v. 3.7 (Carl Zeiss, Göttingen, Germany).
2.6. Quantitative Real-Time PCR
The brains, gonads, livers, and middle intestines of six randomly selected fish from each tank were sampled, immersed in five volumes of Trizol reagent (Ambion, Carlsbad, CA, USA), and stored at −80 °C until use. The stored tissue was thawed before RNA extraction and reverse-transcribed to cDNA using RT & GO master mix (MP Biomedicals). The concentration of cDNA used in the analyses was 100 ng/µL.
The 18S rRNA gene (GenBank, EF126037.1) of olive flounder showed more stable results than other housekeeping genes, so we selected it as a control housekeeping gene. The primers used for mRNA gene expression analysis are shown in
Table 2. The levels of expression of growth (GH and IGF) and maturation-related (FSH, LH, ERα, and ERβ) genes in the brain, VIT genes in the liver and gonads, and digestive enzyme-related (amylase, chymotrypsin2, trypsin2, trypsin3, and lipase) genes in the middle intestine were examined. cDNA (100 ng/µL) was subjected to real-time PCR with an initial denaturation step of 2 min at 98 °C followed by 40 amplification cycles of denaturation at 98 °C for 10 s, annealing at 60 °C for 10 s, and extension at 68 °C for 30 s.
2.7. Statistical Analysis
Statistical analyses were performed using one-way ANOVA and Duncan’s multiple range test [
33] in SPSS version 24.0 (SPSS Inc., Chicago, IL, USA). All values are expressed as the mean ± SE of triplicate samples. In all analyses,
p < 0.05 was taken to indicate statistical significance. SE was used for differences between statistical analyses in results, and SD for general differences such as water temperature and dissolved oxygen.
4. Discussion
SPC is gaining traction as an alternative source of protein to FM for addition to commercial fish diets, overcoming the problems associated with the increasing cost of FM and poor digestibility of soybean [
19,
20,
37]. Weight gain and SGR of olive flounder fed the LF1 and LF2 diets were comparable to those fed the Con diet, indicating that 50% FM could be substituted with SPC supplemented with limiting AAs (lysine and methionine) in the 60% FM-based diet without compromising growth performance. This is in line with previous studies on gilthead seabream (
Sparus aurata L.) [
9] and starry flounder (
Platichthys stellatus) [
19].
Arginine (2.04–2.10% of the diet) [
34] and lysine (1.50–2.10% of the diet) [
35] requirements in the diet of olive flounder were met in all experimental diets in the present study. However, methionine levels (1.17–1.37% of the diet) in all experimental diets, including the Con diet, were slightly lower than requirements (1.44–1.49% of the diet in the presence of 0.06% cysteine) [
36]. Previous studies have demonstrated that cysteine supplementation can spare about 40–50% of the dietary methionine requirement for
Catla catla [
38] and stinging catfish (
Heteropneustes fossilis) [
39]. Therefore, the relatively high (0.28–0.61%) cysteine content in the diets used in this study was able to lower the dietary methionine requirement of olive flounder. Increased substitution of dietary FM with SPC increased ∑SFA but decreased ∑MUFA and ∑n-3 HUFA. ∑n-3 HUFA is crucial for the optimal growth and health of fish [
40]. The Con and LF1 diets satisfied the dietary ∑n-3 HUFA requirement of olive flounder (8.16–10.20% of total FA) [
4], but the LF2 diet had a slightly low ∑n-3 HUFA content. Nevertheless, this does not have a negative effect on growth performance as it does for
P. olivaceus, in line with previous studies [
39,
40].
Replacement of 50% FM in the diet with SPC supplemented with limiting AAs did not affect the DFI, FE, and PER of olive flounder. This is partially in line with [
41], who reported that the growth performance and FE of red sea bream (
Pagrus major) fed a similar percentage of an SPC-based diet with AA supplementation (for 60 days) were higher than in fish fed the same diet without supplementation; however, the latter fish had similar parameters after 153 days of feeding. In another study, the DFI, FE, and PER of black sea bream (
Acanthopagrus schlegelii) fed a diet with a replacement of 40% FM with SPC were comparable to fish fed a 60% FM-based diet [
42]. In another study, there were no differences in growth or FE between gilthead seabream fed a 60% FM-based diet or the same diet in which up to 40% FM was replaced by SPC supplemented with methionine for 10 weeks [
9]. Similarly, in [
19], there were no changes in the DFI of starry flounder fed a diet with substitution of up to 60% FM by SPC with AA supplementation compared to fish fed a control diet of 68% FM. In this study, CF, VSI, or HSI did not decrease in treatment groups with supplementation, in line with previous studies on olive flounder [
39] and yellow croaker (
Larimichthys crocea) [
20]. However, we found that GSI results at 12 months were higher in both sexes fed the control diet. This may have been due to ∑n-3 HUFA levels and ER gene expression, which are very important for growth and maturation [
43,
44,
45,
46]. Changes in dietary FA profile affect steroid production in the African catfish (
Clarias gariepinus) [
27], and adding appropriate amounts of n-3 or n-6 ∑HUFA is important for improving egg quality [
45]. VIT production is induced via the ER and accumulates in egg yolk [
25,
29]. However, ref. [
25] reported no differences in the GSI of female olive flounder fed a 63% FM-based diet or diets with replacement of 20% and 30% FM with SPC. This may have been due to the difference in size of the fish between that study (initial weight 150 g) and ours (initial weight 727 g), as female olive flounder can reach maturity in the second year of life [
47].
In this study, AA and FA profiles of dorsal muscles were found to be affected by dietary treatment, which is in line with several previous studies [
16,
19,
24,
41,
44,
48,
49,
50]. In this study, the proximate lipid composition of olive flounder was not influenced by dietary treatment, but high levels of crude lipid were found in the dorsal muscles of LF2 fish, probably due to the high level of protein in that diet. Similarly, in a previous study, the crude protein and total lipid compositions of rainbow trout (
Oncorhynchus mykiss) differed in fish fed different FM or plant protein diets [
48]. Similar findings have been reported in common sole [
26].
Regarding gonads, as noted in the results, there were no differences at 4 months, but the substitute diet resulted in a greater proportion of spermatogonia and abnormal transformation from spermatogonia to spermatozoa at 12 months; in addition, oocytes failed to absorb yolk globules in such fish. Maturation is influenced by photoperiod and water temperature [
47,
51], and gonadal development in olive flounder occurs in the spring [
52]. October was the fourth month of this experiment, so there were no differences in the histological results at that time point, only after a year [
25,
53]. In addition, the GSI and ∑n-3 HUFA levels of dorsal muscle were lower in LF2 fish than in controls, while, as already noted, the crude lipid level was lower in controls. Ref. [
44] reported that mature fish utilize fat absorbed from digestion, leading to a decrease in their body fat. ∑n-3 HUFA is essential for both maturation [
44] and oocyte development [
54]. Previous studies have shown that dietary and muscle ∑n-3 HUFA levels play important roles in the maturation of gilthead sea bream [
43] and Atlantic salmon (
Salmo salar) [
45].
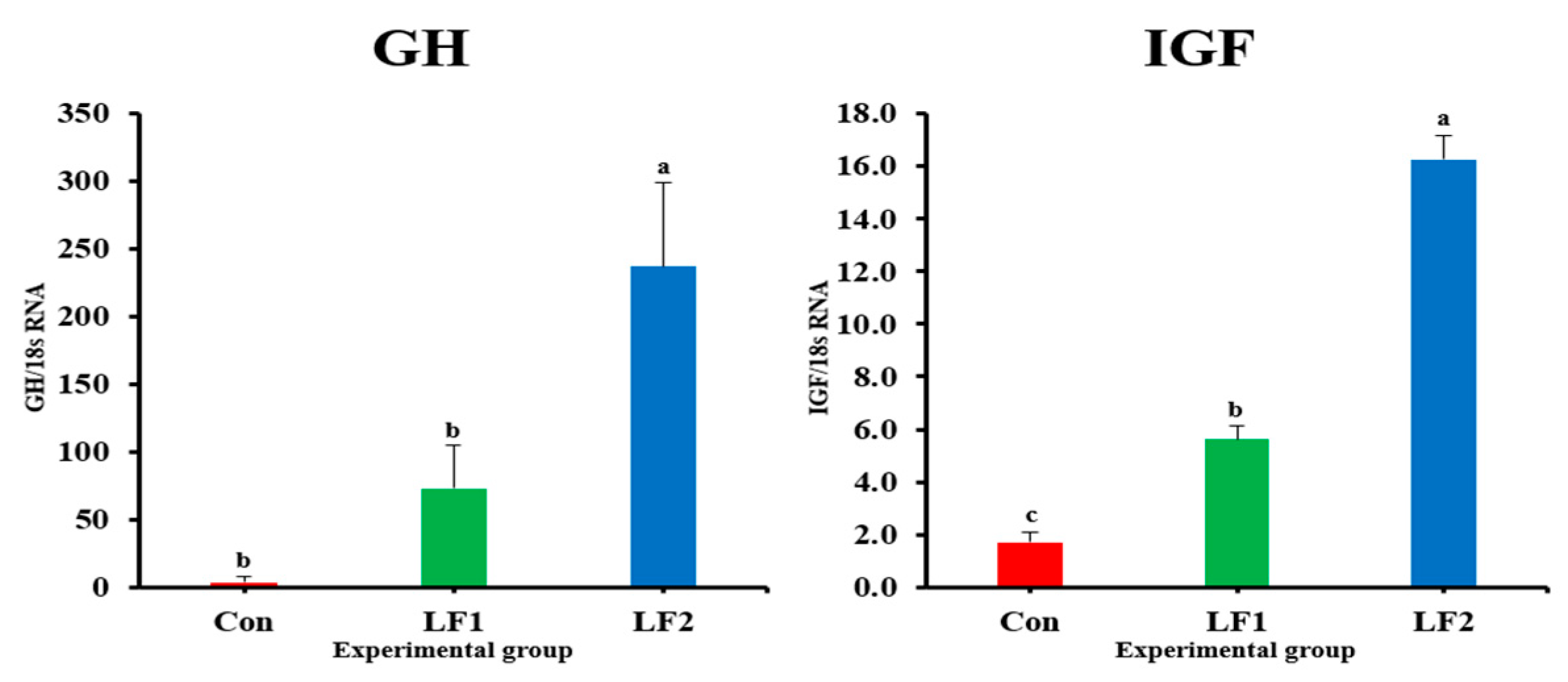
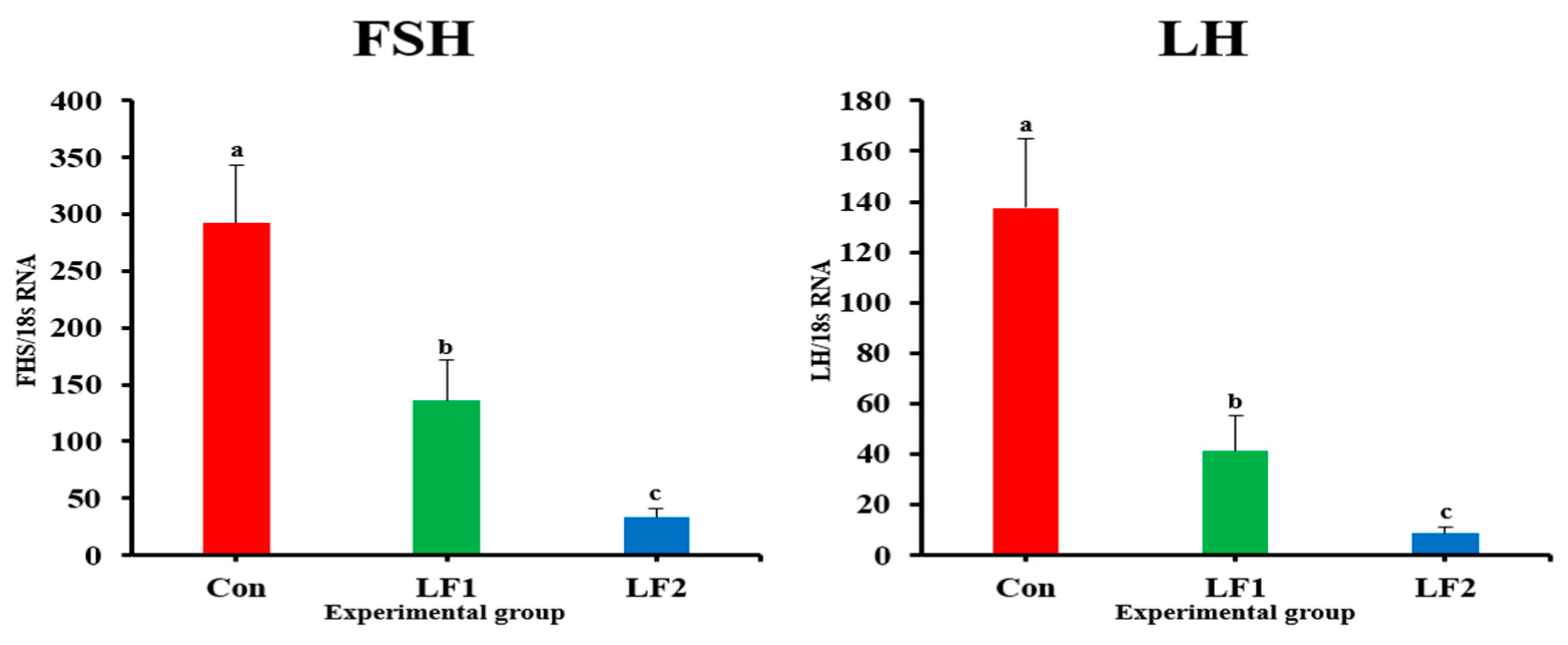
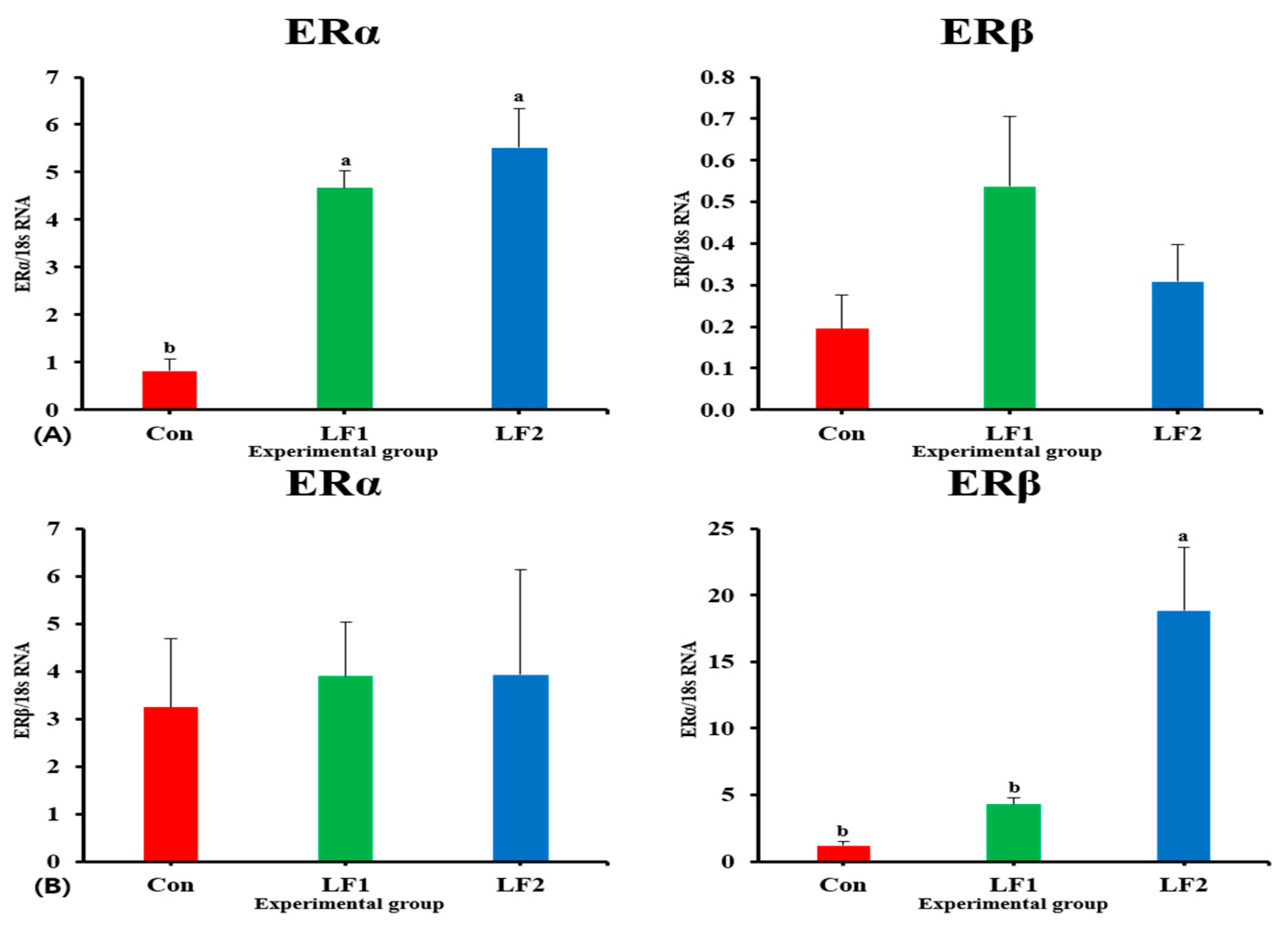
The expression levels of genes that encode growth hormones (GH and IGF) in fish fed the LF2 diet were higher than in controls. However, maturity-related genes showed the opposite trend. Previous studies have shown that growth slows down during maturation [
44,
55] and maturation-related gene expression suppresses growth-related gene expression [
25]. Maturation occurs along the brain–pituitary–gonad axis, and maturation-related genes are expressed in the brain [
28,
54]. The expressions of FSH, LH, ERα, and ERβ affect ovarian development and act as part of a positive steroid feedback loop [
25,
28,
56]. FSH induces gonadal growth, and LH mainly regulates gonadal maturation [
28,
57,
58]. The low levels of maturation-related indicators (GSI and results of histological analysis) appeared to be related to decreased FSH and LH gene expression in fish, which is a consequence of increased dietary FM replacement by SPC. ER is involved in the production of VIT and plays an important role in its accumulation in the egg yolk [
25,
29]. GH and E2 gene expression occur at the same time [
55], and the ER is a receptor of E2 [
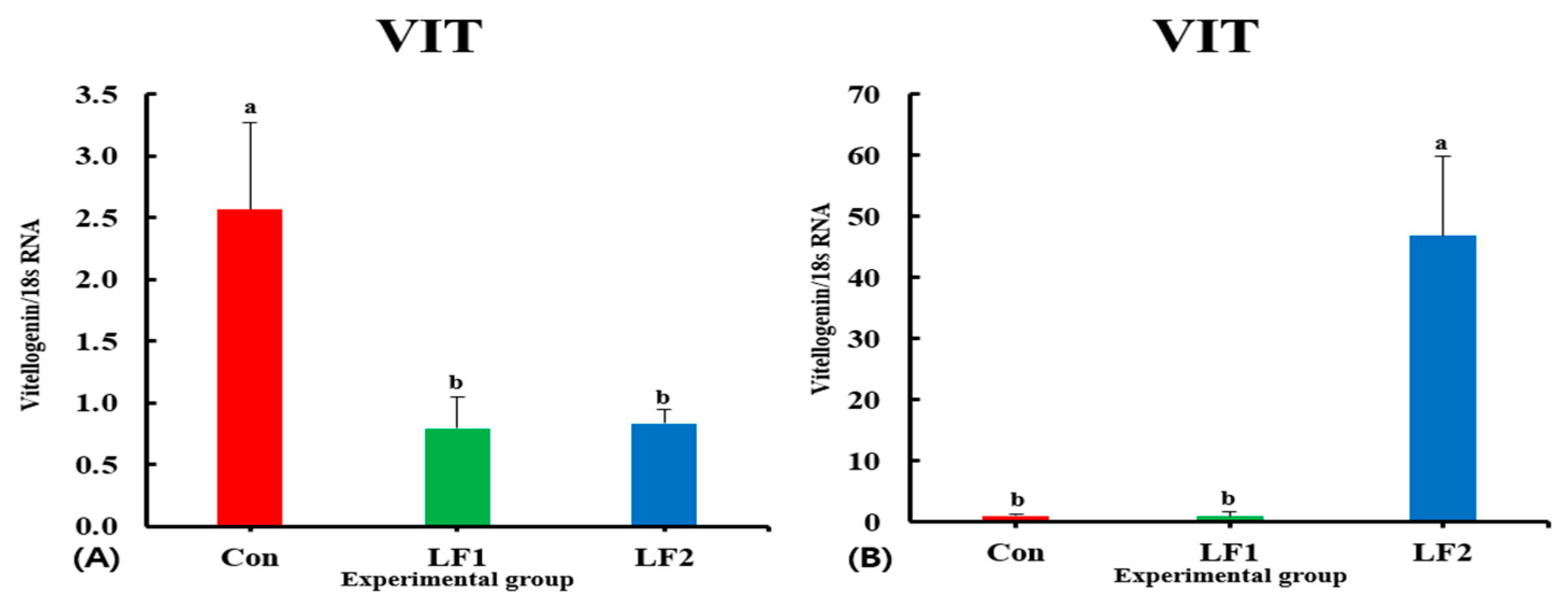
25]. This explains the highest level of ER gene expression in the LF2 diet group. VIT is produced in the liver and absorbed by oocytes, and it is used as a nutrient for embryonic development after fertilization [
25]. Gonadal VIT gene expression level was highest in the Con group, in line with two previous studies [
25,
44]. However, the expression of the VIT gene in the liver was highest in LF2 fish. Ref. [
59] reported that VIT expression in the liver was highest at the late-VIT (LV) stage and was highest in the ovary at the post-VIT (PV) stage in turbot (
Scophthalmus maximus). The fish in the Con group would have been considered to be in the PV stage, LF1 fish would be in the LV stage, and LF2 fish would be in the early VIT (EV) stage. VIT gene expression in the ovary, but not in the liver, was the same as in [
59]. This may have been due to the influence of ER gene expression, which was highest in the LF2 group. ER gene expression is significantly higher at the beginning of the reproduction cycle in rainbow trout [
60]. Based on these results, we assume that increased dietary FM replacement with SPC made it more difficult for fish to digest lipids and accumulate liver-produced VIT in the oocytes or delayed the timing of VIT production relative to the Con diet group.
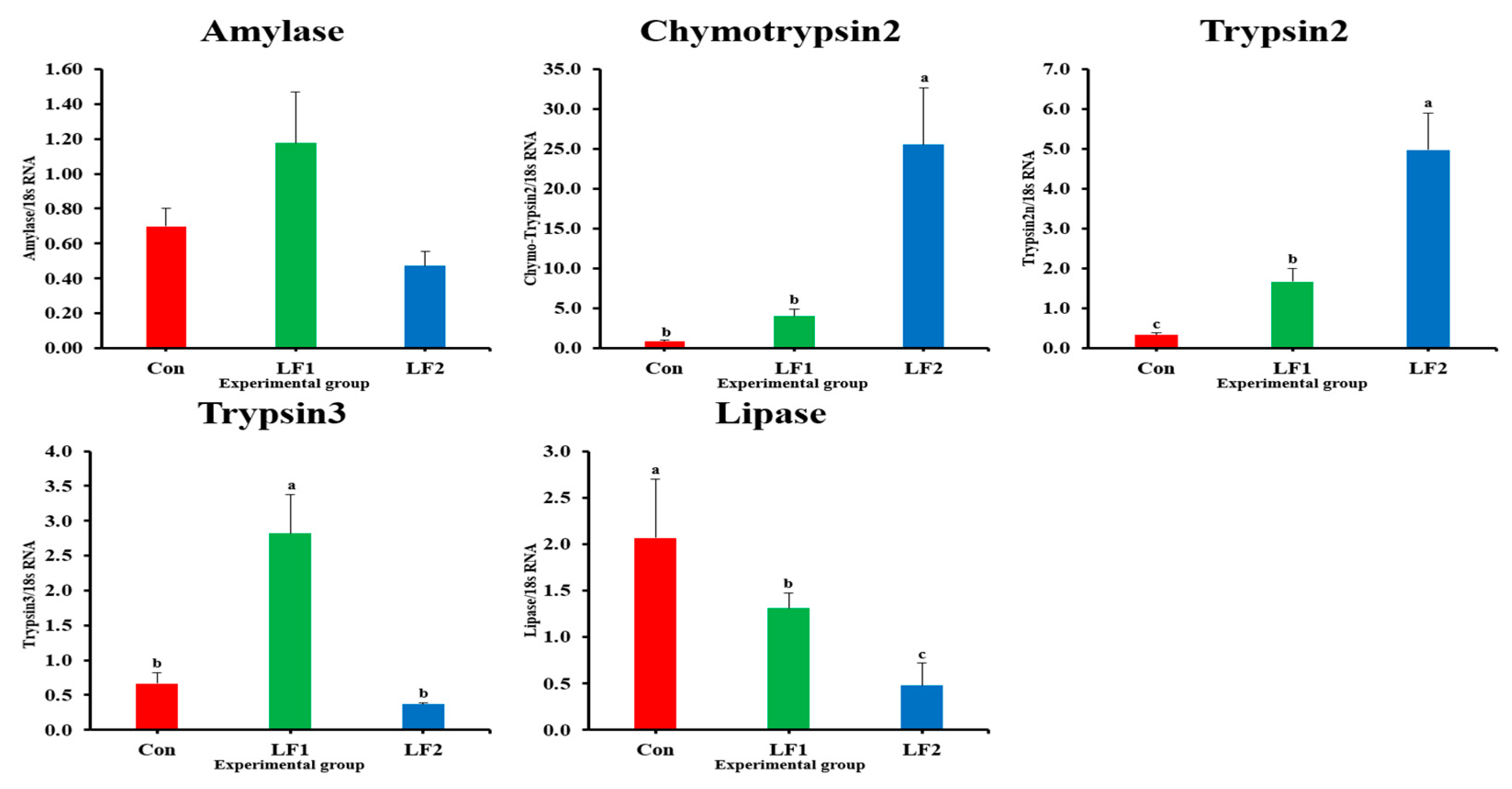
The expression levels of the chymotrypsin2 and trypsin2 genes in the middle intestine increase with an increase in replacement with SPC in this study. Similarly, ref. [
19] reported such an increase in the trypsin enzyme. Two previous studies of proteolytic enzymes showed that 40% dietary FM replacement with SPC increased the activities of pepsin and trypsin in rainbow trout and starry flounder [
19,
61]. However, the apparent digestibility coefficient (ADC) of protein decreases with increased dietary FM replacement with SPC [
16]. However, despite the difference in the amounts of digestive enzymes in the intestine and ADC results, there was no difference in the crude protein content in the dorsal muscle [
16,
19]. It has been postulated that an imbalance in the amino acid content of the diet may result in a reduction in the digestive rate [
16]. However, this has not been demonstrated to affect growth in this study, and numerous previous studies have indicated that such an imbalance does not influence digestive processes [
16,
21,
47,
61]. Nevertheless, previous studies suggest that the trace level of trypsin inhibitor present in SPC may stimulate the production of protein digestive enzymes [
19,
61]. Therefore, the high levels of chymotrypsin2 and trypsin2 gene expression in LF2 fish would increase the digestibility of SPC in the diet. However, the expression of the lipase gene was lowest in LF2 fish, similar to reports on juvenile hybrid grouper (
Epinephelus fuscoguttatus♀ ×
E.
lanceolatus♂) [
62]. Fat absorbed through digestion triggers maturation in fish [
25,
43]. Therefore, the lowest expression of the lipase gene in the LF2 group may have been related to maturation. It is imperative to assess the complexity of the individual and the subsequent generation that provides the feed.
Prior research has indicated that the supplementation of FM with heated soybeans in African catfish diets has resulted in reduced fertilization, hatching, and survival rates [
61]. The quality of dietary protein has been demonstrated to influence ovarian size and the viability of eggs produced [
27,
63,
64]. This assessment should be conducted in order to determine the feasibility of replacing FM with SPC through future verification.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}