
Estimates of the Age, Growth, and Mortality of Triplophysa scleroptera (Herzenstein, 1888) in the Upper Reaches of the Yellow River, China

Abstract
:1. Introduction
2. Materials and Methods
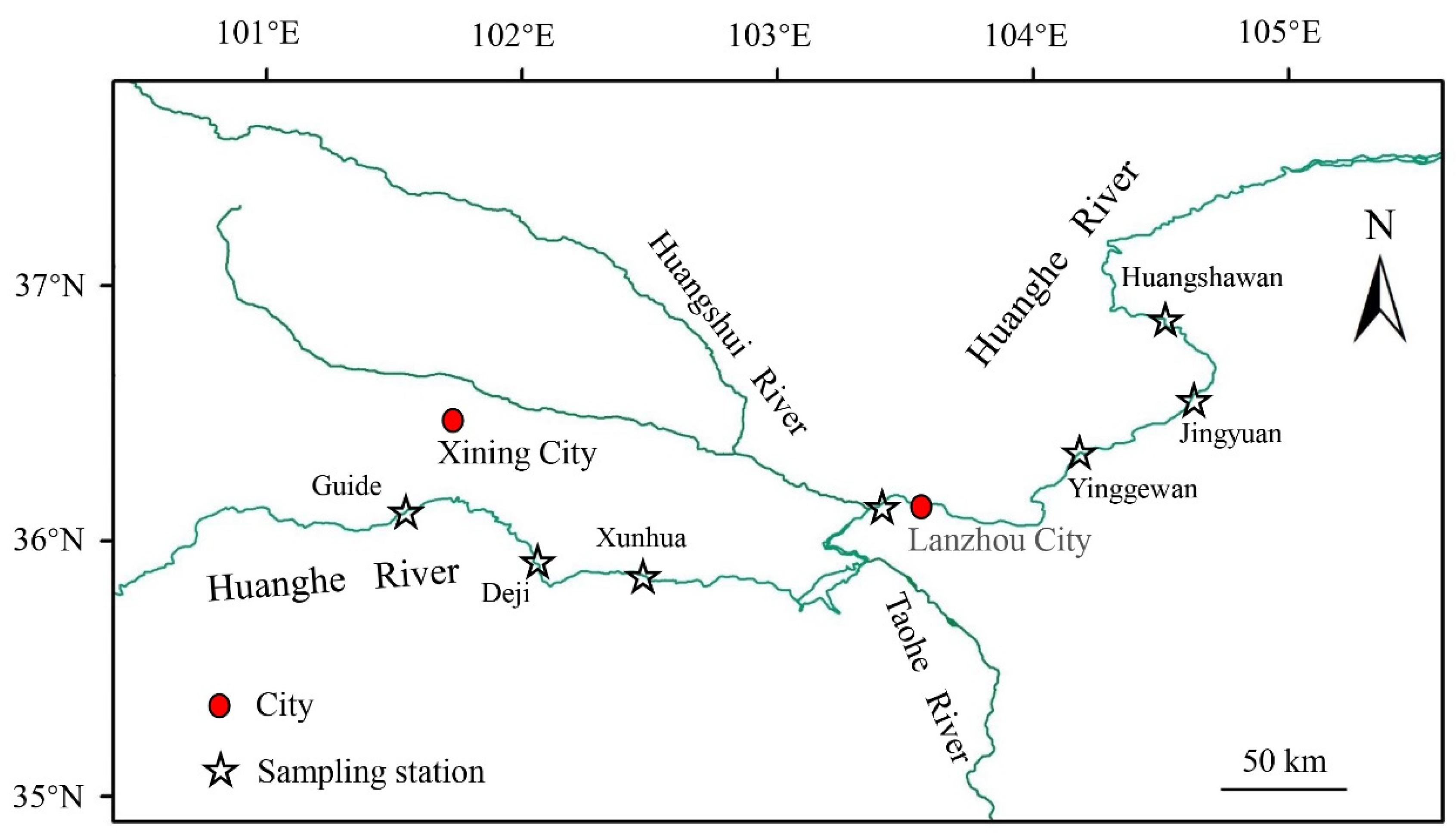
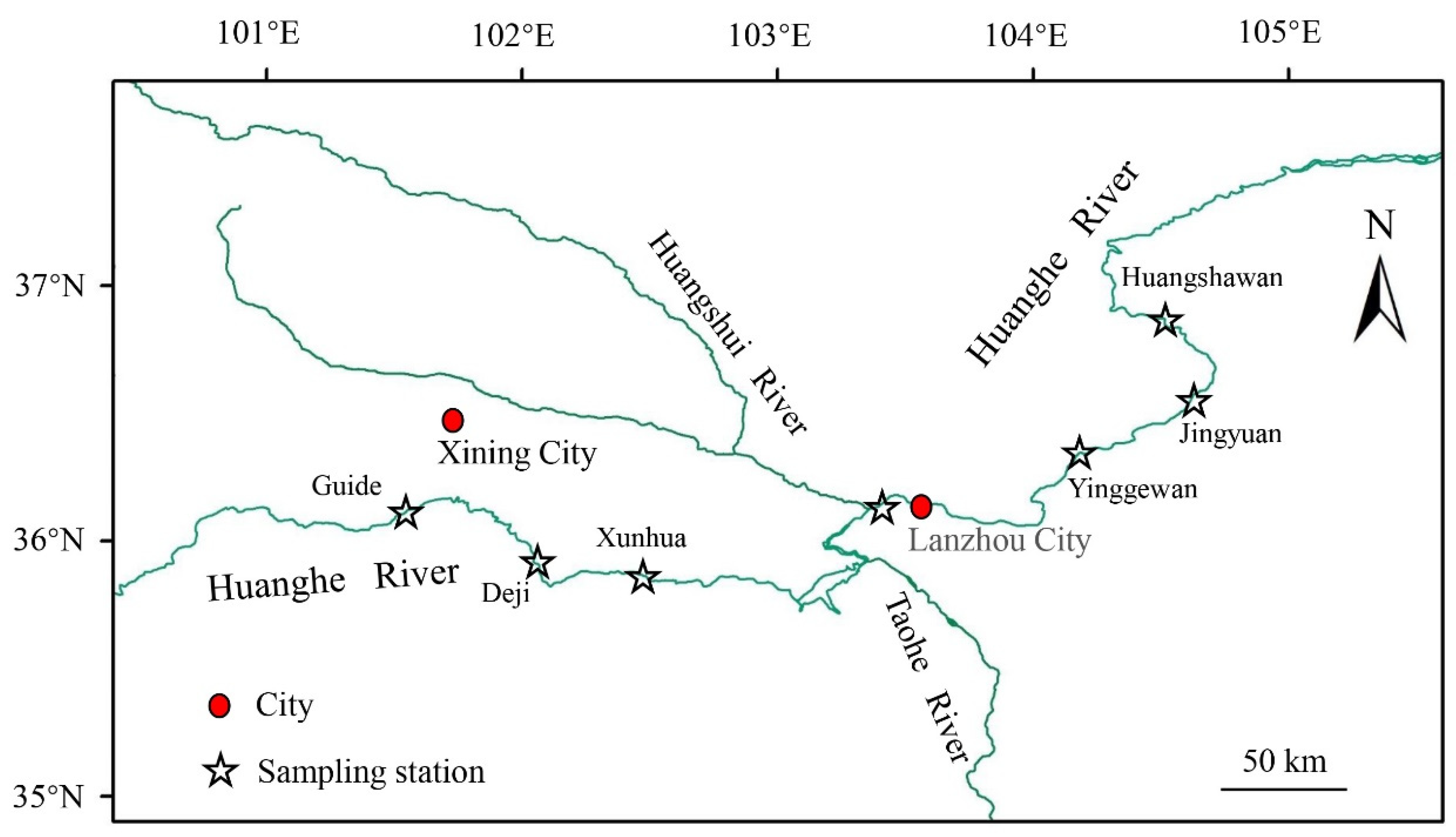
2.1. Study Area and Sample Collection
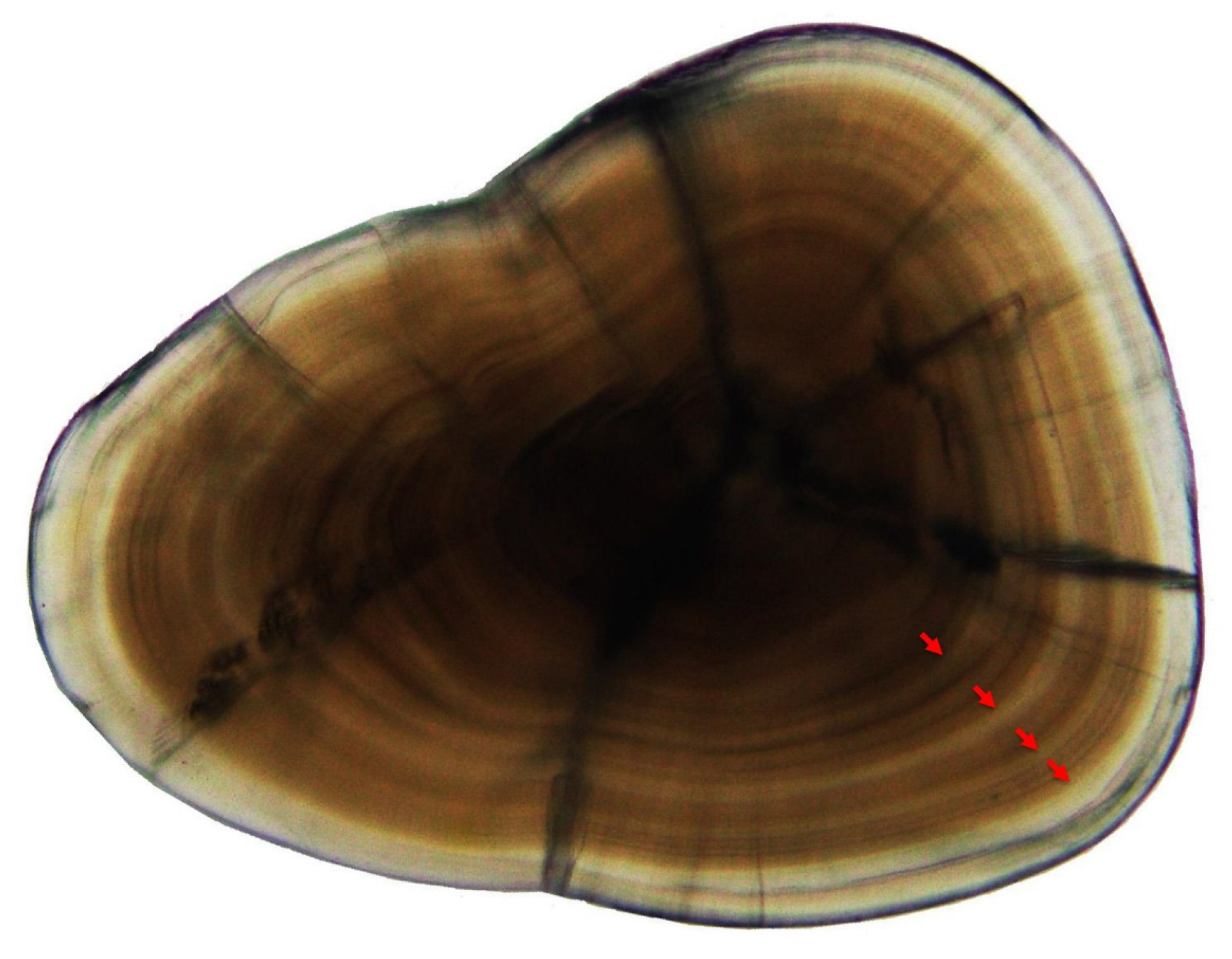
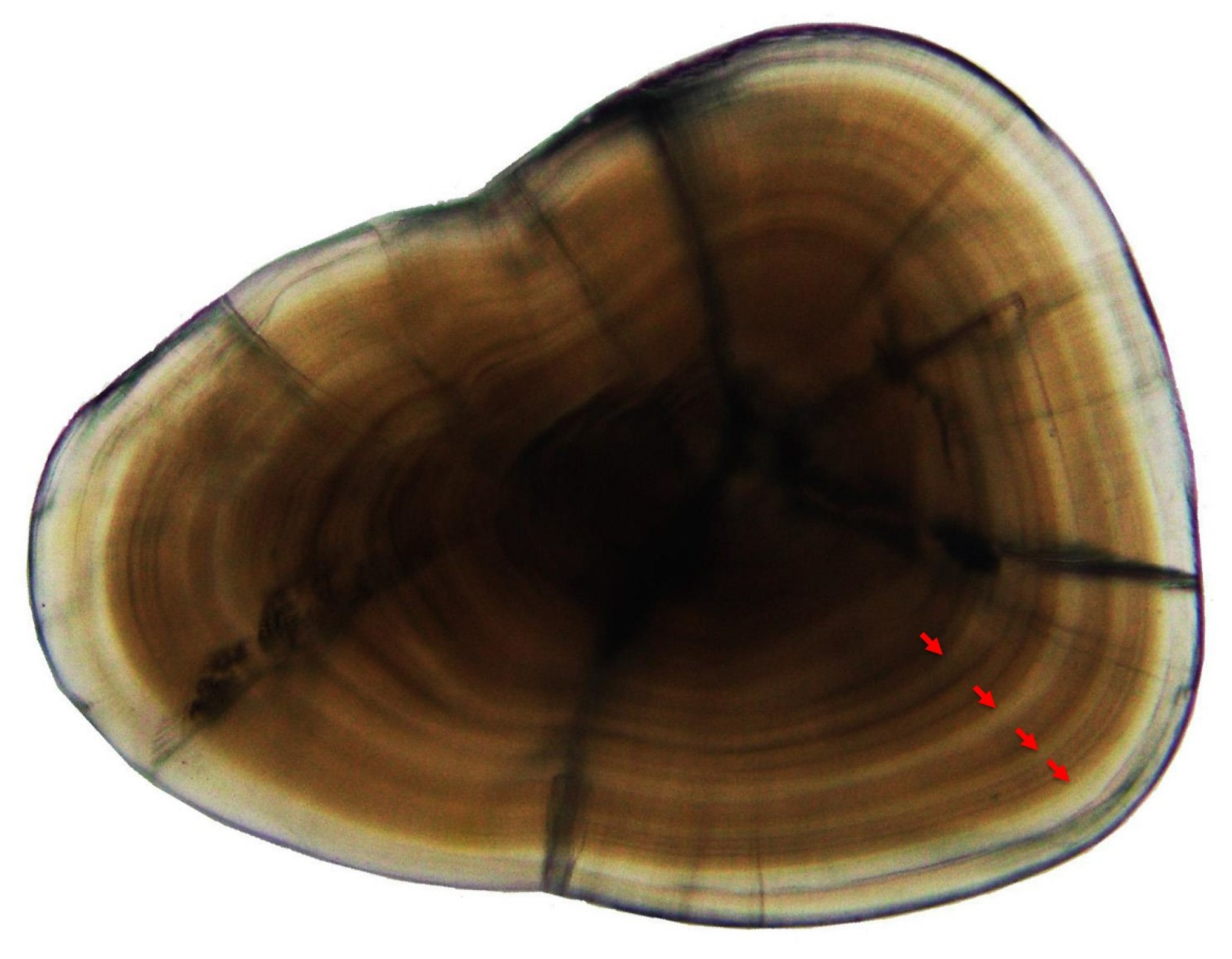
2.2. Age Estimation
2.3. Length–Weight Relationship
2.4. Estimation of Growth Equation Parameters
2.5. Total Mortality, Fishing Mortality and Exploitation Rate
3. Result
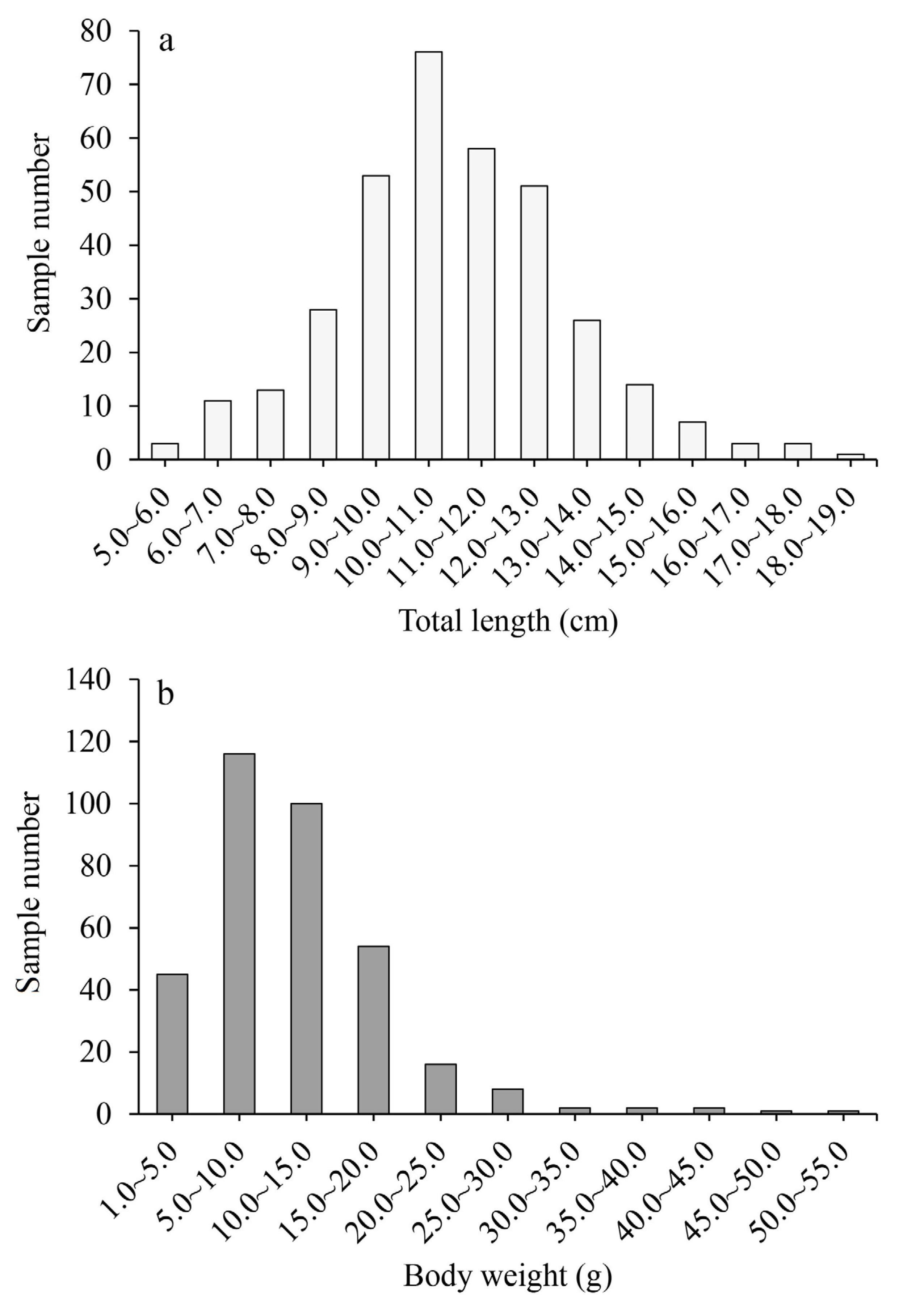
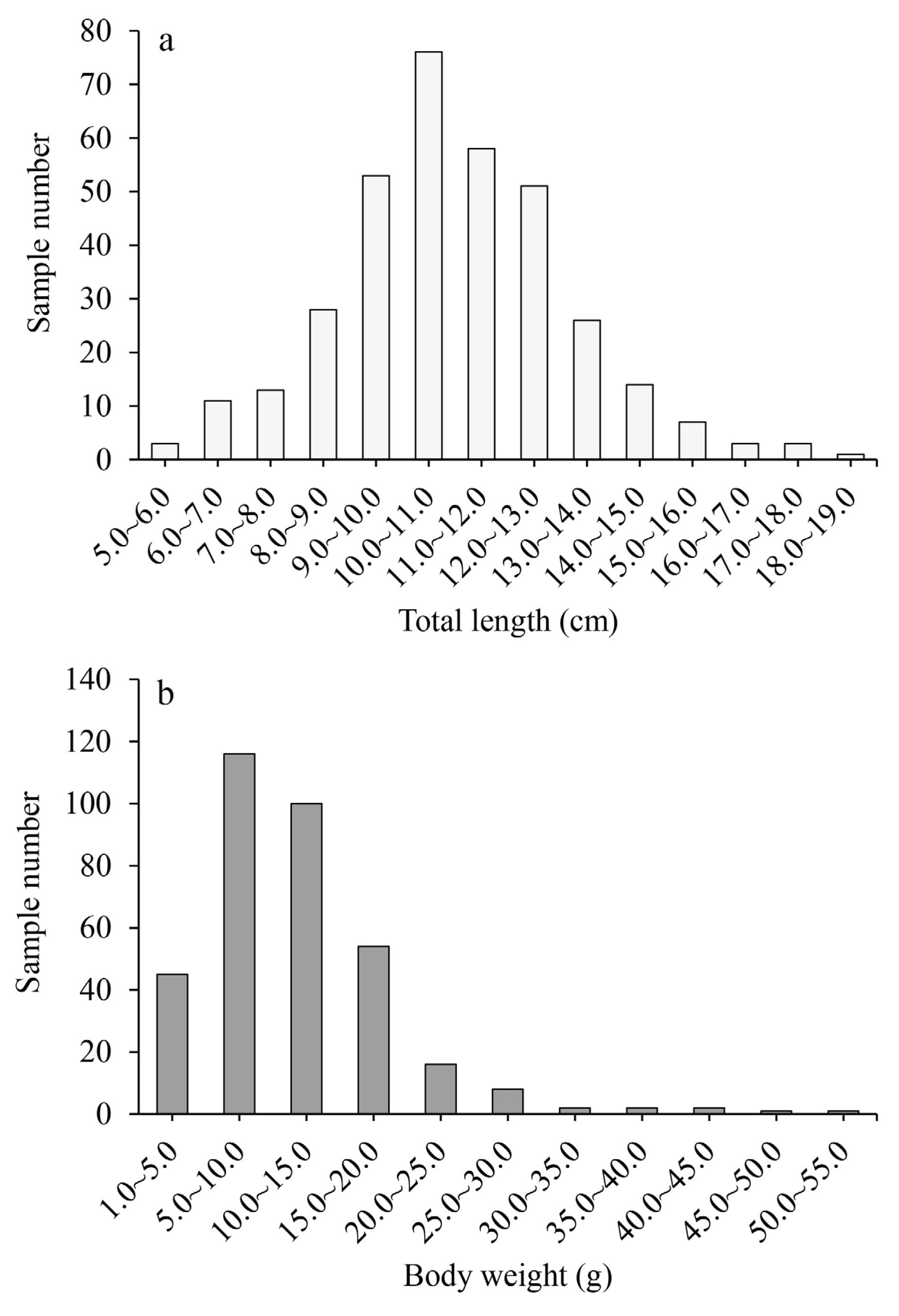
3.1. Population Structure
3.2. Age Distribution
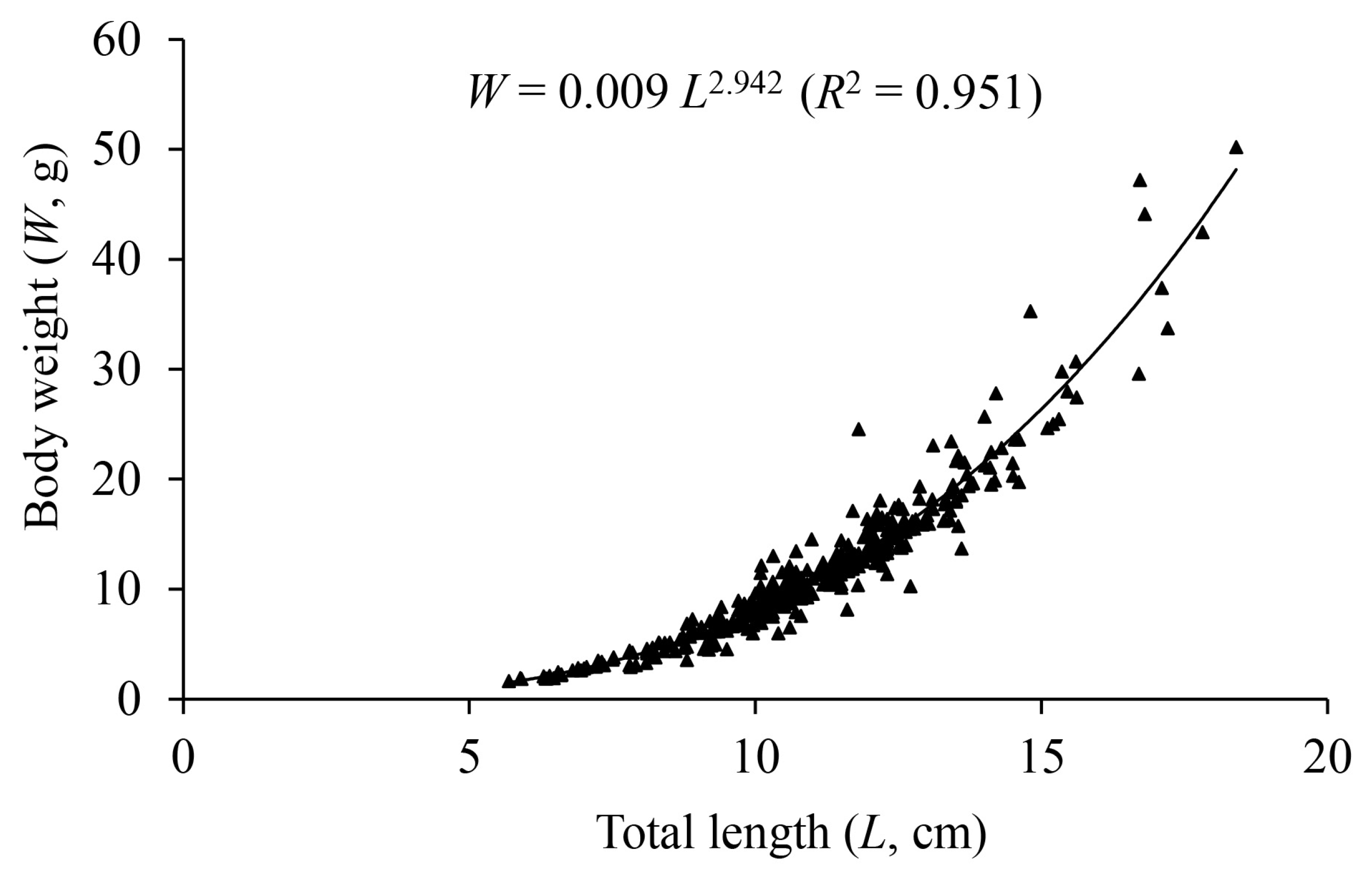
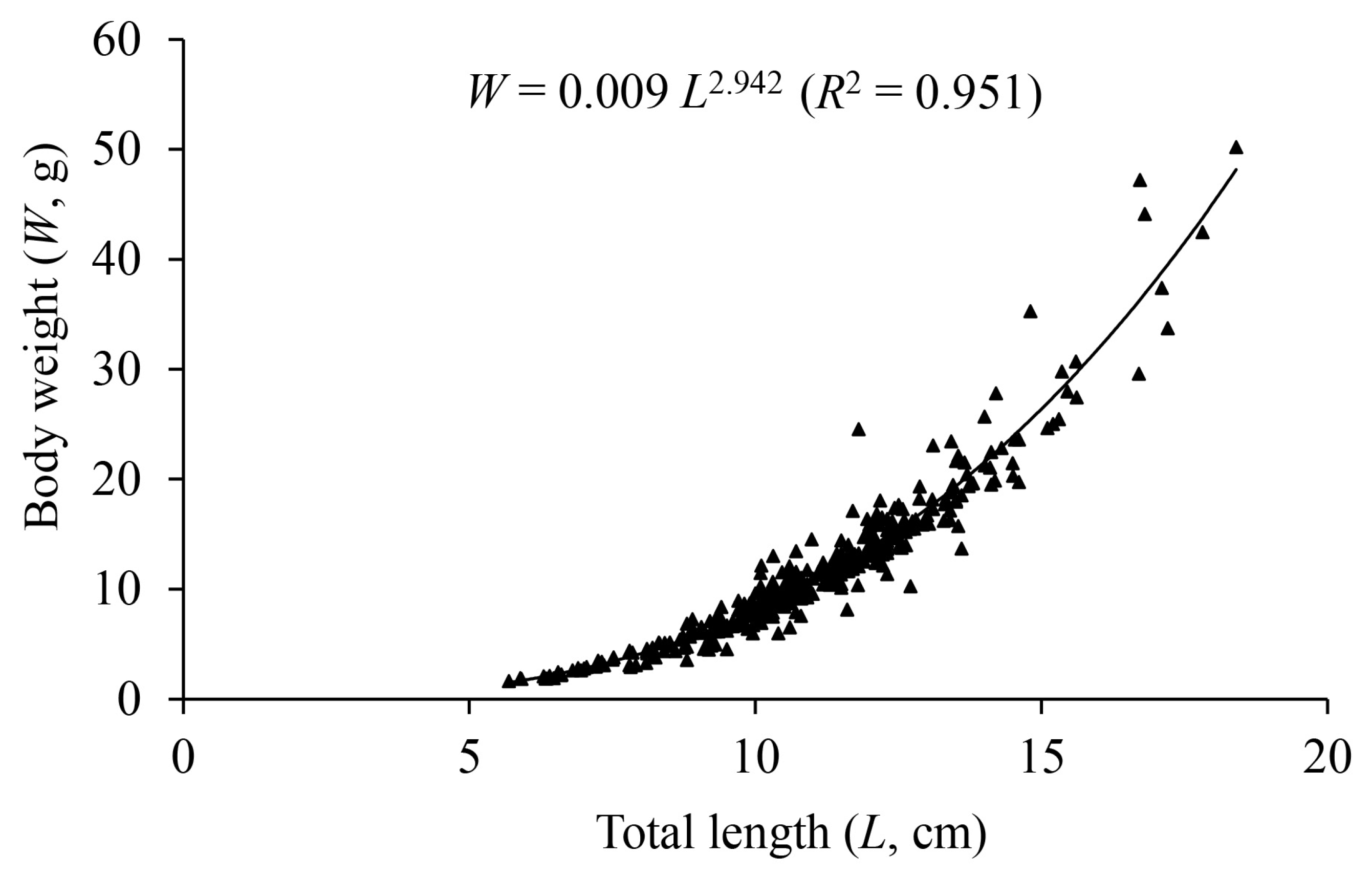
3.3. Length–Weight Relationship
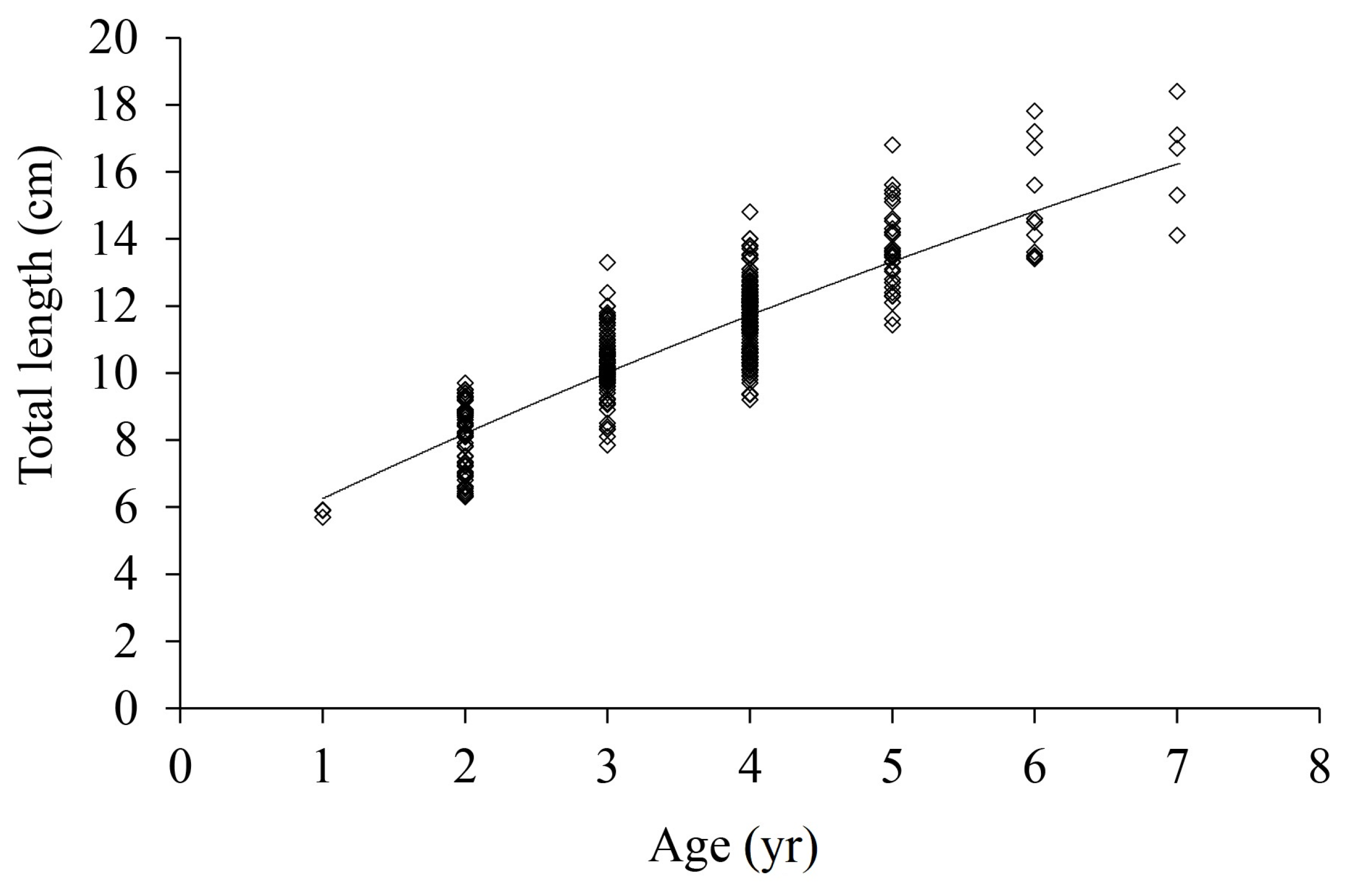
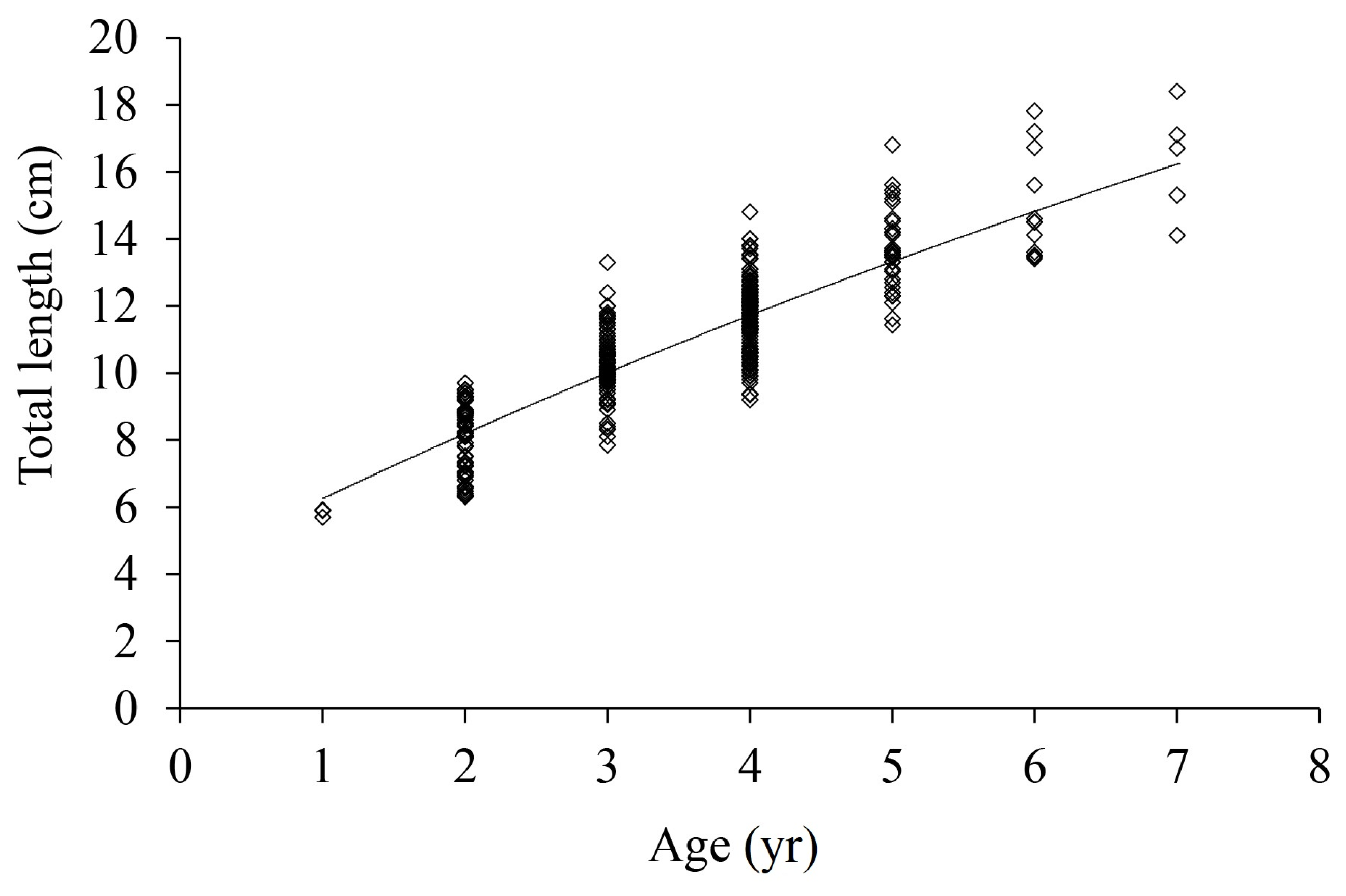
3.4. Growth Equation
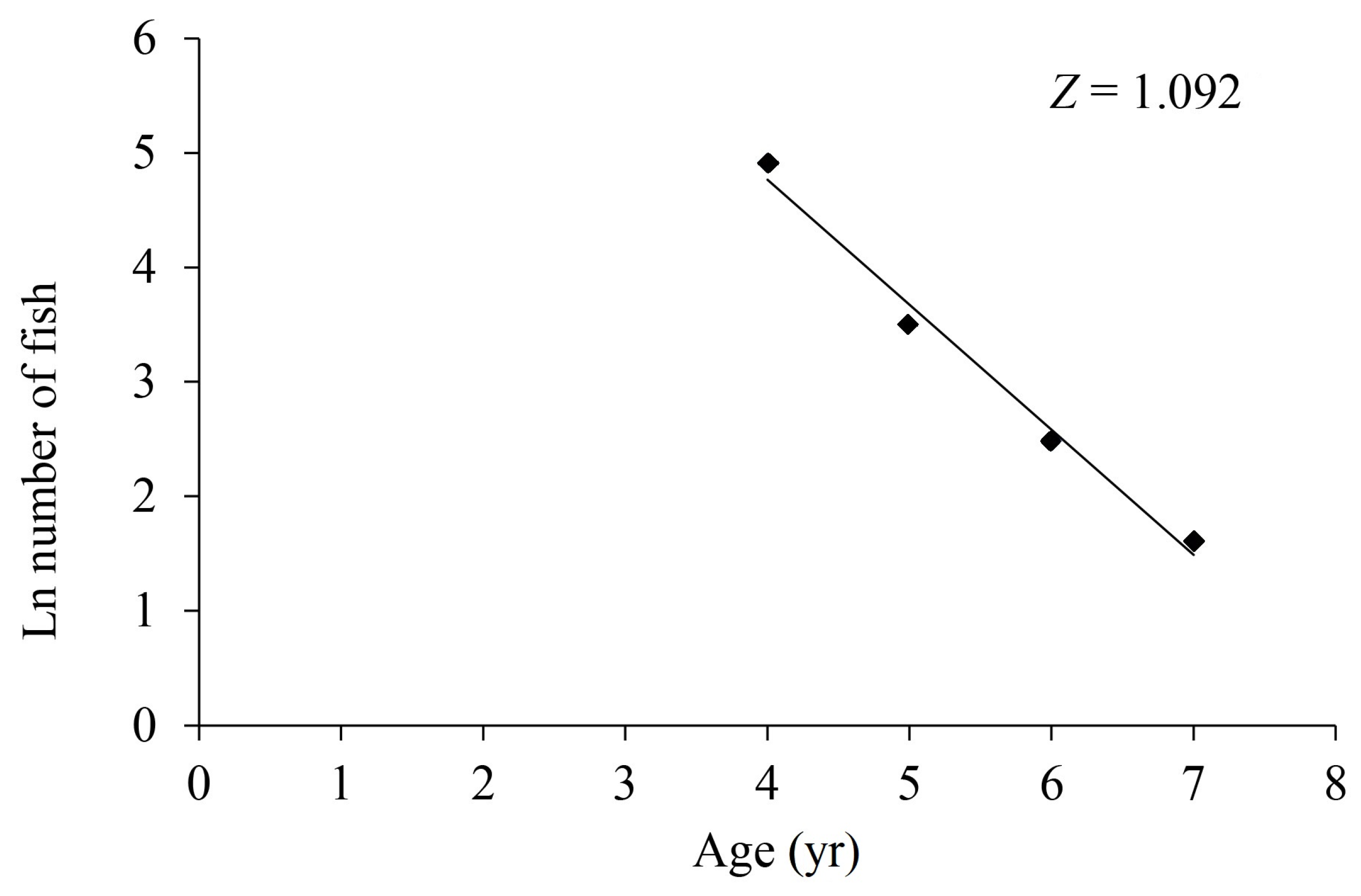
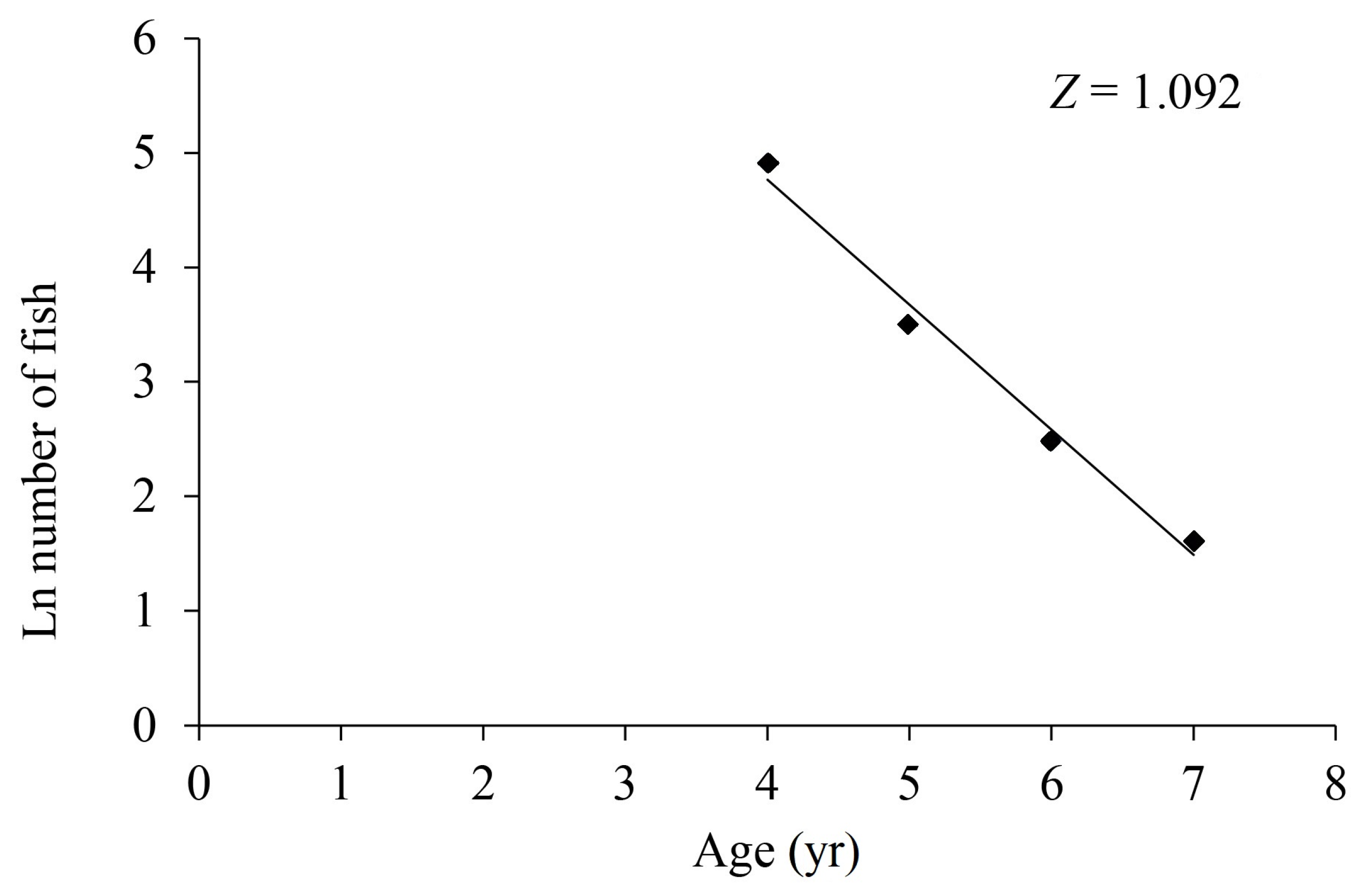
3.5. Mortality and Exploitation Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, T.; Zhang, Y.P.; Yang, Z.Y.; Liu, Z.; Du, Y.Y. DNA barcoding reveals cryptic diversity in the underestimated genus Triplophysa (Cypriniformes: Cobitidae, Nemacheilinae) from the northeastern Qinghai-Tibet Plateau. Bmc. Evol. Biol. 2020, 20, 151. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Liu, N.F.; Yang, J.X. A brief review of Triplophysa (Cypriniformes: Balitoridae) species from the Tarim Basin in Xinjiang, China, with description of a new species. Zootaxa 2007, 1605, 47–58. [Google Scholar] [CrossRef]
- Hu, J.X.; Liu, M.D.; He, D.K. Phylogeography of Triplophysa stenura (Nemacheilidae): Responded to the Mid-Pleistocene Climate Transition in the Qinghai-Tibetan Plateau. Zool. Stud. 2020, 30, e67. [Google Scholar]
- Wang, Y.; Shen, Y.J.; Feng, C.G.; Zhao, K.; Song, Z.B.; Zhang, Y.P.; Yang, L.D.; He, S.P. Mitogenomic perspectives on the origin of Tibetan loaches and their adaptation to high altitude. Sci. Rep. 2016, 6, 29690. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.G.; Jiang, J.P.; Wang, Y.Z.; Zhang, E.; Zhang, Y.Z.; Li, L.L.; Xie, F.; Cai, B.; Cao, L.; Zheng, G.M.; et al. Red List of China’s Vertebrates. Biodivers. Sci. 2016, 24, 500–551. (In Chinese) [Google Scholar]
- Ding, R.H. The Fishes of Sichuan, China; Sichuan Publishing House of Science and Technology: Chengdu, China, 1994; pp. 73–75. (In Chinese) [Google Scholar]
- Qi, W.H.; Guo, Y.S.; Wang, H.; He, M.; Chen, Y.L.; Yu, T.L. Study on biology of Triplophysa scleroptera in Xiaman Nature Reserve, Sichuan. Sichuan. J. Zool. 2008, 27, 1157–1162. (In Chinese) [Google Scholar]
- Zhu, S.H.; Yan, S.; Chen, Z.L. Study on the relationship between oxygen consumption rate and asphyxiation piont and body mass of Triplophysa scleroptera. J. Qinghai. Univ. 2020, 38, 34–38. (In Chinese) [Google Scholar]
- Jackson, G.D. Advances in defining the life histories of myopsid squid. Mar. Freshwater. Res. 2004, 55, 357–365. [Google Scholar] [CrossRef]
- Wang, J.L.; Liu, W.; Li, P.L.; Tang, F.J.; Lu, W.Q. Estimation of Coregonus ussuriensis age, growth, and maturation in China’s Amur River. PeerJ. 2022, 10, e12817. [Google Scholar] [CrossRef]
- Labbaci, A.; Chaoui, L.; Kara, M.H. Age, growth and reproduction of the Mediterranean killifish Aphanius fasciatus Nardo, 1827 in Mellah Lagoon (Eastern Algeria). Environ. Biol. Fish. 2019, 102, 663–674. [Google Scholar] [CrossRef]
- Andres, M.J.; Slack, W.T.; Peterson, M.S.; Kimmel, K.D.; Lewis, B.R.; Grammer, P.O. Growth estimation of western population segment gulf sturgeon using length-at-age and mark–recapture data. T Am. Fish. Soc. 2019, 148, 176–190. [Google Scholar] [CrossRef]
- Fan, J.J.; Zhao, G.J.; Mu, X.M.; Lu, A.; Tian, P.; Gao, P.; Sun, W.Y. Effects of cascading reservoirs on streamflow and sediment load with machine learning reconstructed time series in the upper Yellow River basin. Catena 2023, 225, 107008. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, Y.P.; Guan, L.H.; Du, Y.Y.; Lou, Z.Y.; Jiao, W.L. Current freshwater fish resources and the application of DNA barcoding in species identification in Gansu Province. Biodivers. Sci. 2015, 23, 306–313. (In Chinese) [Google Scholar] [CrossRef]
- Li, L.T.; Yang, X.F.; Yang, R.B.; Fan, Q.X.; Wei, K.J.; Jiang, H. Age structure and growth characteristics of Triplophysa orientalis in the middle of the Yarlung Tsangpo River, Tibet. J. Huazhong Agric. Univ. 2016, 35, 117–123. (In Chinese) [Google Scholar]
- Froese, R.; Tsikliras, A.C.; Stergiou, K.I. Editorial note on weight–length relations of fishes. Acta. Ichthyol. Piscat. 2011, 41, 261–263. [Google Scholar] [CrossRef]
- Tian, N.N.; Yang, R.B.; Tan, B.Z.; Zeng, X.L.; He, L.Q.; Xu, Z.L.; Zhu, Z.; Liu, H.P.; Yang, X.F. Age, growth, and reproductive characteristics of Triplophysa stewarti in Lake Chugutso, Tibet. J. Fish. Sci. China 2022, 29, 1013–1021, (In Chinese with English abstract). [Google Scholar]
- Ricker, W.E. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can. 1975, 191, 1–382. [Google Scholar]
- von Bertalanffy, L. A quantitative theory of organic growth (inquiries on growth laws. II). Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Chen, Y.; Jackson, D.A.; Harvey, H.H. A comparison of von Bertalanffy and polynomial functions in the modeling fish growth data. Can. J. Fish. Aquat. Sci. 1992, 49, 1228–1235. [Google Scholar] [CrossRef]
- Pauly, D.; Moreau, J.; Abad, N. Comparison of age-structured and length-converted catch curves of brown trout Salmo trutta in two French rivers. Fish. Res. 1995, 22, 197–204. [Google Scholar] [CrossRef]
- Beverton, R.J.H.; Holt, S.J. On the Dynamics of Exploited Fish Populations; Chapman and Hall: London, UK, 1957; p. 533. [Google Scholar]
- Pauly, D. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. J. Conserv. Inter. Explor. Marit. 1980, 39, 175–192. [Google Scholar] [CrossRef]
- Zhan, B.Y.; Lou, D.C.; Zhong, J.S. An assessment of the filefish population and rational exploitation of the resource. J. Fish. China 1986, 10, 409–418. (In Chinese) [Google Scholar]
- Ralston, S. Mortality rates of snappers and groupers. In Tropical Snappers and Grouper: Biology and Fisheries Management; Polovina, J.J., Ralston, S., Eds.; Westview Press: Boulder, CO, USA, 1987; pp. 375–404. [Google Scholar]
- Gray, C.A.; Barnes, L.M.; Robbins, W.D.; Van Der Meulen, D.E.; Ochwada–Doyle, F.A.; Kendall, B.W. Length–and age–based demographics of exploited populations of stout whiting, Sillago robusta Stead, 1908. J. Appl. Ichthyol. 2017, 33, 1073–1082. [Google Scholar] [CrossRef]
- Zhang, X.F.; He, C.L.; Song, Z.B. Age and Growth of Triplophysa markehenensis from the Markehe River in Upper Reaches of the Dadu River. Chin. J. Zool. 2010, 45, 11–20. (In Chinese) [Google Scholar]
- Deng, H.T.; Yue, X.J.; Chen, D.Q.; Tian, H.W.; Liu, S.P. Growth characteristics and feed habit of Triplophysa atenura in Nujiang River. Freshw. Fish. 2010, 40, 26–33. (In Chinese) [Google Scholar]
- Wang, C.; Liang, Y.Q. Age and growth of Triplophysa anterodorsalis Zhu & Cao, 1989 in the Heishui River, China. J. Appl. Ichthyol. 2017, 33, 1215–1217. [Google Scholar]
- Yao, N.; Wang, M.; Gong, Y.R.; Ju, M.H.; Wang, S.; Chen, S.A.; Song, Y. Biological characteristics of Triplophysa bombifrons in Tarim River Basin. Jiangsu Agr. Sci. 2018, 46, 146–149. (In Chinese) [Google Scholar]
- Yao, N.; Ma, L.; Jing, S.S.; Chen, S.A. Growth characteristics of Triplophysa siluroides (Herzenstein) in Beichuanhe Basin of Qinghai. J. Inn. Mong. Agr. Univ. 2019, 40, 5–10. (In Chinese) [Google Scholar]
- Jin, S.S.; Wang, X.Y.; Lin, X.; Chen, S.A.; Liu, M.C.; Xie, C.X. Age and Growth of Triplophysa tenuis in Kaidu River, Xinjiang. Xinjiang Agr. Sci. 2020, 57, 181–189. (In Chinese) [Google Scholar]
- Zhao, J.F.; Qiu, L.H.; Zhou, Q.; Zhou, X.Y.; Shen, J.Z. Age structure and growth characteristics of Triplophysa yarkandensis in the Qarqan River, Xinjiang. J. Huazhong Agr. Univ. 2023, 42. Available online: https://kns.cnki.net/kcms2/detail/42.1181.S.20230801.1115.002.html (accessed on 1 August 2023). (In Chinese).
- Campana, S.E. Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J. Fish. Biol. 2001, 59, 197–242. [Google Scholar] [CrossRef]
- Phelps, Q.E.; Edwards, K.R.; Willis, D.W. Precision of five structures for estimating age of common carp. N. Am. J. Fish. Manag. 2007, 27, 103–105. [Google Scholar] [CrossRef]
- Campana, S.E.; Thorrold, S.R. Otoliths, increments, andelements: Keys to a comprehensive understanding of fish populations. Can. J. Fish. Aquat. Sci. 2001, 58, 30–38. [Google Scholar] [CrossRef]
- Duan, Y.J.; Xie, C.X.; Zhou, X.J.; Ma, S.S.; Huo, B. Age and growth characteristics of Schizopygopsis younghusbandi Regan, 1905 in the Yarlung Tsangpo River in Tibet, China. J. Appl. Ichthyol. 2014, 30, 948–954. [Google Scholar] [CrossRef]
- Chen, Y.F.; He, D.K.; Duan, Z.H. Annuli characters of selincuo schizothoracine fish (Gymnocypris selincuoensis) in Selincuo Lake, Qinghai–Tibetan Plateau. Acta. Zool. Sin. 2002, 48, 384–392, (In Chinese with English abstract). [Google Scholar]
- Zeng, L.; Tang, W.Q. Discussion on age determination methods for two esquamate Triplophysa fishes. Chin. J. Zool. 2010, 45, 94–103, (In Chinese with English abstract). [Google Scholar]
- Tsagarakis, K.; Basusta, A.; Basusta, N.; Biandolino, F.; Bostanci, D.; Buz, K.; Djodjo, Z.; Dulcic, J.; Gokoglu, M.; Gucu, A.; et al. New Fisheries-related data from the Mediterranean Sea (October 2015). Mediterr. Mar. Sci. 2015, 16, 703–713. [Google Scholar] [CrossRef]
- Yedier, S.; Kontaş, S.; Bostancı, D. Marmara Denizi’nde Yaşayan Pagellus acarne (Risso, 1827)’nin Kondisyon Faktörü, Boy-Boy ve Boy-Ağırlık İlişkileri. J. Anatol. Environ. Anim. Sci. 2019, 4, 82–88. [Google Scholar]
- Szymon, S.; Berg, F. Varying relationships between fish length and scale size under changing environmental conditions—Multidecadal perspective in Atlantic herring. Ecol. Indic. 2022, 134, 108494. [Google Scholar]
- Froese, R. Cube law, condition factor and weight–length relationships: History, meta–analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, C.; Tian, Y.J.; Watanabe, Y. Age, growth, and mortality rate of the yellow goosefish Lophius litulon (Jordan, 1902) in the Yellow Sea. J. Oceanol. Limnol. 2021, 39, 732–740. [Google Scholar] [CrossRef]
- Onikura, N.; Nakajima, J. Age, growth and habitat use of the topmouth gudgeon, Pseudorasbora parva in irrigation ditches on northwestern Kyushu Island, Japan. J. Appl. Ichthyol. 2013, 29, 186–192. [Google Scholar] [CrossRef]
- Xie, J.Y. Length-weight and length-length relationships of four endemic fish species from the upper Yellow River in the Tibetan Plateau, China. J. Appl. Ichthyol. 2015, 31, 958–960. [Google Scholar] [CrossRef]
- Chen, S.; Xie, C.; Li, D.; Yao, N.; Ding, H.; Zhang, Z. Length–weight relationships of five Triplophysa species from the northwest of China. J. Appl. Ichthyol. 2017, 33, 1234–1236. [Google Scholar] [CrossRef]
- Naveed, A.; Muhammad, F.K. Length–weight relationships of four Triplophysa species from northern, Pakistan. J. Appl. Ichthyol. 2018, 34, 1223–1224. [Google Scholar]
- Munro, J.; Pauly, D. A simple method for comparing the growth of fishes and invertebrates. Fishbyte 1983, 1, 5–6. [Google Scholar]
- Pauly, D.; Moreau, J.; Prein, M. A comparison of overall growth performance of tilapia in open waters and aquaculture. In Proceedings of the ICLARM Conference Proceedings of the Second International Symposium on Tilapia in Aquaculture, Bangkok, Thailand, 16–20 March 1987; pp. 469–479. [Google Scholar]
- Stearns, S.C. The Evolution of life history traits: A critique of the theory and a review of the data. Annu. Rev. Ecol. Evol. S. 1977, 8, 145–171. [Google Scholar] [CrossRef]
- Mims, M.C.; Olden, J.D. Life history theory predicts fish assemblage response to hydrologic regimes. Ecology 2012, 93, 35–45. [Google Scholar] [CrossRef]
- Li, X.Q.; Chen, Y.F. Age structure, growth and mortality estimates of an endemic Ptychobarbus dipogon (Regan, 1905) (Cyprinidae: Schizothoracinae) in the Lhasa River, Tibet. Environ. Biol. Fish. 2009, 86, 97–105. [Google Scholar] [CrossRef]
- Ma, B.S.; Nie, Y.Y.; Wei, K.J.; Xu, B.; Zhu, X.Y.; Xu, J. Estimates on age, growth, and mortality of Gymnocypris firmispinatus (Cyprinidae: Schizothoracinae) in the Anning River, China. J. Oceanol. Limnol. 2019, 37, 736–744. [Google Scholar] [CrossRef]
- Lu, W.Q.; Li, P.L.; Ma, B.; Huo, T.B.; Yin, Z.Q.; Tang, F.J.; Wang, J.L. Assessment of fishery management parameters for major prey fish species in the lower reaches of the Songhua River. Front. Mar. Sci. 2023, 10, 1166634. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | N | Proportion (%) | L (cm) | W (g) | ||
|---|---|---|---|---|---|---|
| Range | Mean ± S.D. | Range | Mean ± S.D. | |||
| 1 | 3 | 0.86 | 5.69–5.91 | 5.83 ± 0.12 | 1.65–1.92 | 1.81 ± 0.14 |
| 2 | 61 | 17.58 | 6.30–9.70 | 8.19 ± 1.03 | 1.88–8.37 | 4.58 ± 1.76 |
| 3 | 97 | 27.96 | 7.85–13.29 | 10.25 ± 0.94 | 4.24–17.13 | 8.23 ± 2.74 |
| 4 | 136 | 39.19 | 9.20–14.80 | 11.62 ± 1.11 | 5.04–35.29 | 13.20 ± 4.19 |
| 5 | 33 | 9.51 | 11.43–16.8 | 13.59 ± 1.27 | 11.37–44.11 | 20.14 ± 6.58 |
| 6 | 12 | 3.46 | 13.40–17.81 | 14.92 ± 1.55 | 13.68–47.2 | 25.72 ± 10.46 |
| 7 | 5 | 1.44 | 14.10–18.40 | 16.32 ± 1.66 | 21.07–50.22 | 32.75 ± 11.47 |
| Species | Investigation Area | Age Determination Materials | Sex | Age (yr) | N | Growth Parameters | Sources | |||
|---|---|---|---|---|---|---|---|---|---|---|
| L∞ (cm) | K (yr−1) | t0 (yr) | φ | |||||||
| T. orientalis | Yarlung Tsangpo River | Otolith | ♀ | 3–9 | 101 | 15.165 | 0.133 | −0.018 | 1.486 | Li et al. [15] |
| ♂ | 3–9 | 82 | 12.519 | 0.177 | −0.069 | 1.443 | ||||
| T. stewarti | Chugutso Lake | Otolith | ♀ and ♂ | 1–6 | 695 | 13.891 | 0.168 | −2.895 | 1.510 | Tian et al. [17] |
| T. markehenensis | Dadu River | Otolith | ♀ and ♂ | 2–8 | 103 | 17.312 | 0.159 | −0.533 | 1.404 | Zhang et al. [27] |
| T. stenura | Nujiang River | Otolith and Vertebra | ♀ and ♂ | 3–14 | 172 | 24.694 | 0.059 | −0.169 | 1.556 | Deng et al. [28] |
| T. anterodorsalis | Heishui River | Vertebra | ♀ and ♂ | 1–5 | 247 | 9.880 | 0.260 | −0.209 | 1.404 | Wang et al. [29] |
| T. bombifrons | Tarim River | Otolith and Vertebra | ♀ and ♂ | 1–6 | 52 | 63.310 | 0.050 | −1.437 | 2.302 | Yao et al. [30] |
| T. siluroides | Beichuan River | Otolith and Scale | ♀ and ♂ | 2–6 | 53 | 10.320 | 0.334 | −0.835 | 1.551 | Yao et al. [31] |
| T. tenuis | Kaidu River | Otolith and Vertebra | ♀ | 2–5 | 83 | 23.750 | 0.640 | −0.700 | 2.558 | Jin et al. [32] |
| ♂ | 1–6 | 56 | ||||||||
| T. yarkandensis | Qarqan River | Otolith and Opercular bone | ♀ | 1–10 | 157 | 40.726 | 0.112 | −0.930 | 2.269 | Zhao et al. [33] |
| ♂ | 1–11 | 148 | 32.791 | 0.157 | −0.715 | 2.227 | ||||
| T. scleroptera | Yellow River | Otolith | ♀ and ♂ | 1–7 | 347 | 37.536 | 0.064 | −1.849 | 1.955 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Liu, J.; Wang, T.; Wang, J. Estimates of the Age, Growth, and Mortality of Triplophysa scleroptera (Herzenstein, 1888) in the Upper Reaches of the Yellow River, China. Fishes 2023, 8, 457. https://doi.org/10.3390/fishes8090457
Li P, Liu J, Wang T, Wang J. Estimates of the Age, Growth, and Mortality of Triplophysa scleroptera (Herzenstein, 1888) in the Upper Reaches of the Yellow River, China. Fishes. 2023; 8(9):457. https://doi.org/10.3390/fishes8090457
Chicago/Turabian StyleLi, Peilun, Jiacheng Liu, Tai Wang, and Jilong Wang. 2023. "Estimates of the Age, Growth, and Mortality of Triplophysa scleroptera (Herzenstein, 1888) in the Upper Reaches of the Yellow River, China" Fishes 8, no. 9: 457. https://doi.org/10.3390/fishes8090457
APA StyleLi, P., Liu, J., Wang, T., & Wang, J. (2023). Estimates of the Age, Growth, and Mortality of Triplophysa scleroptera (Herzenstein, 1888) in the Upper Reaches of the Yellow River, China. Fishes, 8(9), 457. https://doi.org/10.3390/fishes8090457