
Lipid Correction for Carbon Stable Isotope Analysis of Yellowfin Tuna


Abstract
:1. Introduction
2. Materials and Methods
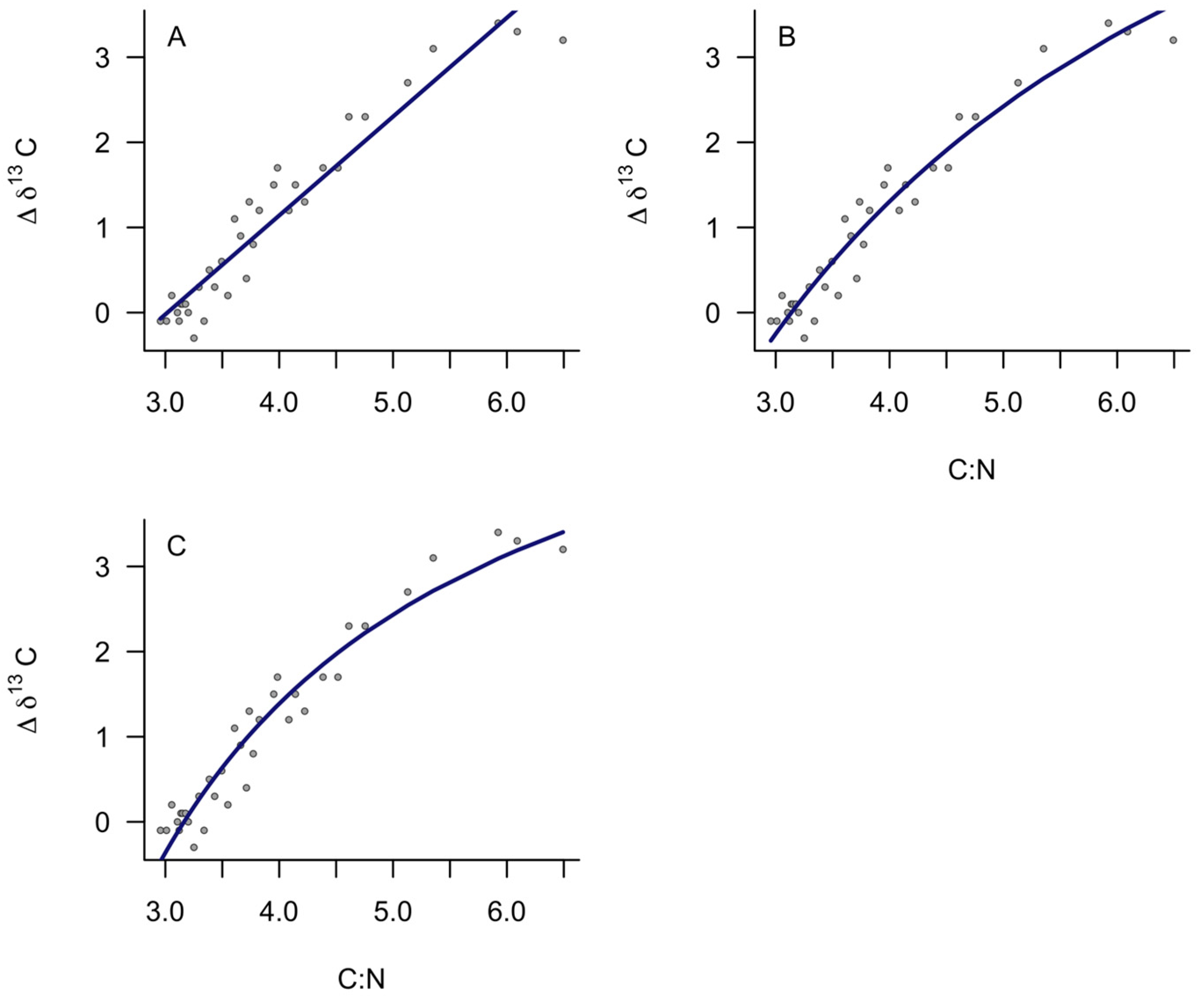
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Layman, C.A.; Arrington, D.A.; Montaña, C.G.; Post, D.M. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef] [PubMed]
- McMahon, K.W.; Hamady, L.L.; Thorrold, S.R. A review of ecogeochemistry approaches to estimating movements of marine animals. Limnol. Oceanogr. 2013, 58, 697–714. [Google Scholar] [CrossRef]
- Matsubayashi, J.; Osada, Y.; Tadokoro, K.; Abe, Y.; Yamaguchi, A.; Shirai, K.; Honda, K.; Yoshikawa, C.; Ogawa, N.O.; Ohkouchi, N.; et al. Tracking long-distance migration of marine fishes using compound-specific stable isotope analysis of amino acids. Ecol. Lett. 2020, 23, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Dance, K.M.; Rooker, J.R.; Shipley, J.B.; Dance, M.A.; Wells, R.J.D. Feeding ecology of fishes associated with artificial reefs in the northwest Gulf of Mexico. PLoS ONE 2018, 13, e0203873. [Google Scholar] [CrossRef] [PubMed]
- Lorrain, A.; Pethybridge, H.; Cassar, N.; Receveur, A.; Allain, V.; Bodin, N.; Bopp, L.; Choy, C.A.; Duffy, L.; Fry, B.; et al. Trends in tuna carbon isotopes suggest global changes in pelagic phytoplankton communities. Glob. Chang. Biol. 2020, 26, 458–470. [Google Scholar] [CrossRef] [PubMed]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Mechanism of Carbon Isotope Fractionation Associated with Lipid Synthesis. Science 1977, 197, 261–263. [Google Scholar] [CrossRef]
- Sweeting, C.J.; Polunin, N.V.C.; Jennings, S. Effects of chemical lipid extraction and arithmetic lipid correction on stable isotope ratios of fish tissues. Rapid Commun. Mass Spectrom. 2006, 20, 595–601. [Google Scholar] [CrossRef]
- Logan, J.M.; Jardine, T.D.; Miller, T.J.; Bunn, S.E.; Cunjak, R.A.; Lutcavage, M.E. Lipid corrections in carbon and nitrogen stable isotope analyses: Comparison of chemical extraction and modelling methods. J. Anim. Ecol. 2008, 77, 838–846. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montaña, C.G. Getting to the fat of the matter: Models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Hussey, N.E.; Olin, J.A.; Kinney, M.J.; McMeans, B.C.; Fisk, A.T. Lipid extraction effects on stable isotope values (δ13C and δ15N) of elasmobranch muscle tissue. J. Exp. Mar. Biol. Ecol. 2012, 434, 7–15. [Google Scholar] [CrossRef]
- Cloyed, C.S.; DaCosta, K.P.; Hodanbosi, M.R.; Carmichael, R.H. The effects of lipid extraction on δ13C and δ15N values and use of lipid-correction models across tissues, taxa and trophic groups. Methods Ecol. Evol. 2020, 11, 751–762. [Google Scholar] [CrossRef]
- Skinner, M.M.; Martin, A.A.; Moore, B.C. Is lipid correction necessary in the stable isotope analysis of fish tissues? Rapid Commun. Mass Spectrom. 2016, 30, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Layman, C.A.; Araujo, M.S.; Boucek, R.; Hammerschlag-Peyer, C.M.; Harrison, E.; Jud, Z.R.; Matich, P.; Rosenblatt, A.E.; Vaudo, J.J.; Yeager, L.A.; et al. Applying stable isotopes to examine food-web structure: An overview of analytical tools. Biol. Rev. 2012, 87, 545–562. [Google Scholar] [CrossRef]
- Lovell, M. Seasonal Variation in the Feeding Ecology of Yellowfin Tuna (Thunnus albacares) from the Northern Gulf of Mexico. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2021. [Google Scholar]
- Kim, S.L.; Koch, P.L. Methods to collect, preserve, and prepare elasmobranch tissues for stable isotope analysis. Environ. Biol. Fishes 2012, 95, 53–63. [Google Scholar] [CrossRef]
- Fry, B. Stable isotopic indicators of habitat use by Mississippi River fish. J. N. Am. Benthol Soc. 2002, 21, 676–685. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 30 June 2023).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Carlisle, A.B.; Litvin, S.Y.; Madigan, D.J.; Lyons, K.; Bigman, J.S.; Ibarra, M.; Bizzarro, J.J. Interactive effects of urea and lipid content confound stable isotope analysis in elasmobranch fishes. Can. J. Fish Aquat. Sci. 2016, 74, 419–428. [Google Scholar] [CrossRef]
- Logan, J.M.; Golet, W.; Smith, S.C.; Neilson, J.; Van Guelpen, L. Broadbill swordfish (Xiphias gladius) foraging and vertical movements in the north-west Atlantic. J. Fish Biol. 2021, 99, 557–568. [Google Scholar] [CrossRef]
- Goñi, N.; Arrizabalaga, H. Seasonal and interannual variability of fat content of juvenile albacore (Thunnus alalunga) and bluefin (Thunnus thynnus) tunas during their feeding migration to the Bay of Biscay. Prog. Oceanogr. 2010, 86, 115–123. [Google Scholar] [CrossRef]
- Logan, J.M.; Lutcavage, M.E. A comparison of carbon and nitrogen stable isotope ratios of fish tissues following lipid extractions with non-polar and traditional chloroform/methanol solvent systems. Rapid Commun. Mass Spectrom. 2008, 22, 1081–1086. [Google Scholar] [CrossRef]
- Sardenne, F.; Ménard, F.; Degroote, M.; Fouché, E.; Guillou, G.; Lebreton, B.; Hollanda, S.J.; Bodin, N. Methods of lipid-normalization for multi-tissue stable isotope analyses in tropical tuna. Rapid Commun. Mass Spectrom. 2015, 29, 1253–1267. [Google Scholar] [CrossRef]
- Kiljunen, M.; Grey, J.; Sinisalo, T.; Harrod, C.; Immonen, H.; Jones, R.I. A revised model for lipid-normalizing δ13C values from aquatic organisms, with implications for isotope mixing models. J. Appl. Ecol. 2006, 43, 1213–1222. [Google Scholar] [CrossRef]
- Hoffman, J.C.; Sutton, T.T. Lipid correction for carbon stable isotope analysis of deep-sea fishes. Deep Sea Res. Part I 2010, 57, 956–964. [Google Scholar] [CrossRef]
- Patterson, H.K.; Carmichael, R.H. The effect of lipid extraction on carbon and nitrogen stable isotope ratios in oyster tissues: Implications for glycogen-rich species. Rapid Commun. Mass Spectrom. 2016, 30, 2594–2600. [Google Scholar] [CrossRef]
- Le-Alvarado, M.; Romo-Curiel, A.E.; Sosa-Nishizaki, O.; Hernández-Sánchez, O.; Barbero, L.; Herzka, S.Z. Yellowfin tuna (Thunnus albacares) foraging habitat and trophic position in the Gulf of Mexico based on intrinsic isotope tracers. PLoS ONE 2021, 16, e0246082. [Google Scholar] [CrossRef]
- Olson, R.J.; Popp, B.N.; Graham, B.S.; López-Ibarra, G.A.; Galván-Magaña, F.; Lennert-Cody, C.E.; Bocanegra-Castillo, N.; Wallsgrove, N.J.; Gier, E.; Alatorre-Ramírez, V.; et al. Food-web inferences of stable isotope spatial patterns of copepods and yellowfin tuna in the pelagic eastern Pacific Ocean. Prog. Oceanogr. 2010, 86, 124–138. [Google Scholar] [CrossRef]
- Richards, T.M.; Gipson, E.E.; Cook, A.; Sutton, T.T.; Wells, R.J.D. Trophic ecology of meso- and bathypelagic predatory fishes in the Gulf of Mexico. ICES J. Mar. Sci. 2019, 76, 662–672. [Google Scholar] [CrossRef]


{kind=link}
| Model | Lipid Correction Equation | Parameter Estimates (SE) |
|---|---|---|
| Linear | a = 1.161 (0.059) b = −3.505 (0.236) | |
| Non-linear Equation (1) | a = 9.356 (2.310) b = −29.359 (7.108) c = 2.181 (1.875) | |
| Non-linear Equation (2) | P = 6.637 (0.249) F = 3.164 (0.031) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dance, M.A.; Lovell, M.S. Lipid Correction for Carbon Stable Isotope Analysis of Yellowfin Tuna. Fishes 2023, 8, 446. https://doi.org/10.3390/fishes8090446
Dance MA, Lovell MS. Lipid Correction for Carbon Stable Isotope Analysis of Yellowfin Tuna. Fishes. 2023; 8(9):446. https://doi.org/10.3390/fishes8090446
Chicago/Turabian StyleDance, Michael A., and Mitchell S. Lovell. 2023. "Lipid Correction for Carbon Stable Isotope Analysis of Yellowfin Tuna" Fishes 8, no. 9: 446. https://doi.org/10.3390/fishes8090446
APA StyleDance, M. A., & Lovell, M. S. (2023). Lipid Correction for Carbon Stable Isotope Analysis of Yellowfin Tuna. Fishes, 8(9), 446. https://doi.org/10.3390/fishes8090446