
Effect of Inorganic Mercury on Semen Quality, Embryo and Larval Development of Bocachico Prochilodus magdalenae



Abstract
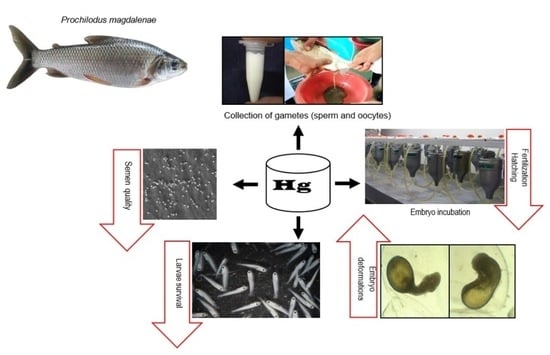
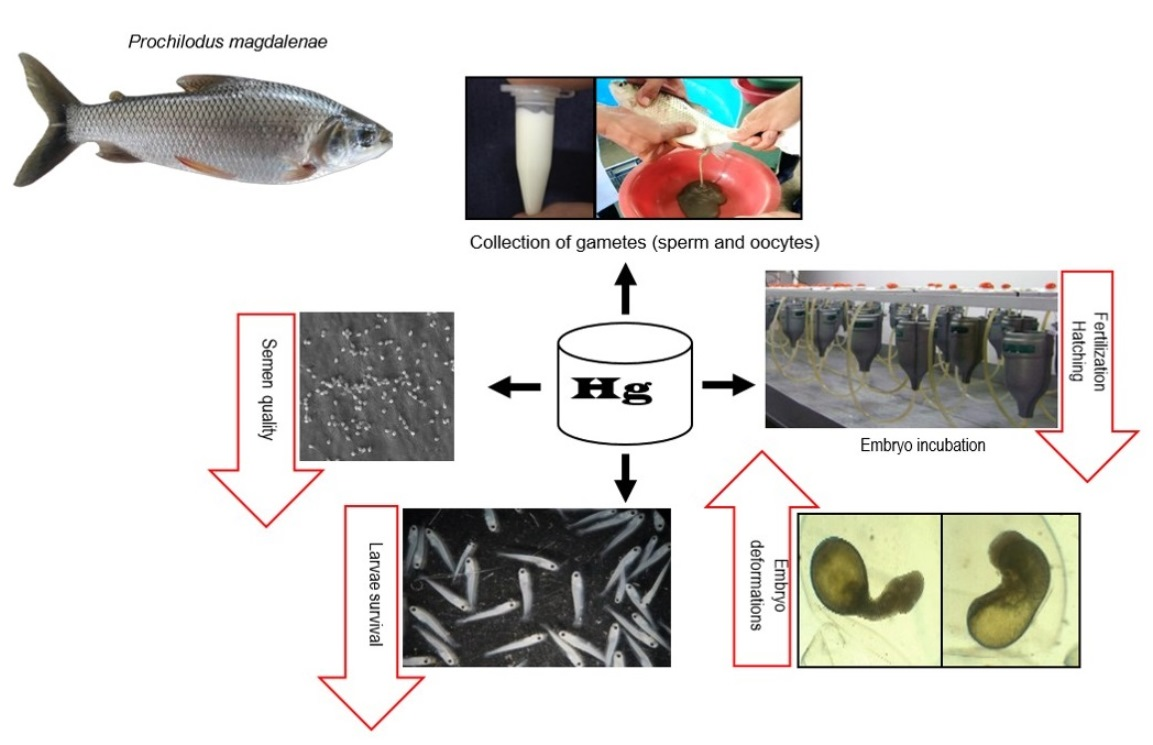
1. Introduction
2. Materials and Methods
2.1. Semen and Egg Collection
2.2. Experiment 1
2.3. Experiment 2
2.4. Embryonic Deformations
2.5. Fertilization and Hatching Rates
2.6. Larval Survival
2.7. Water Quality
2.8. Statistical Analysis
3. Results
3.1. Semen Quality
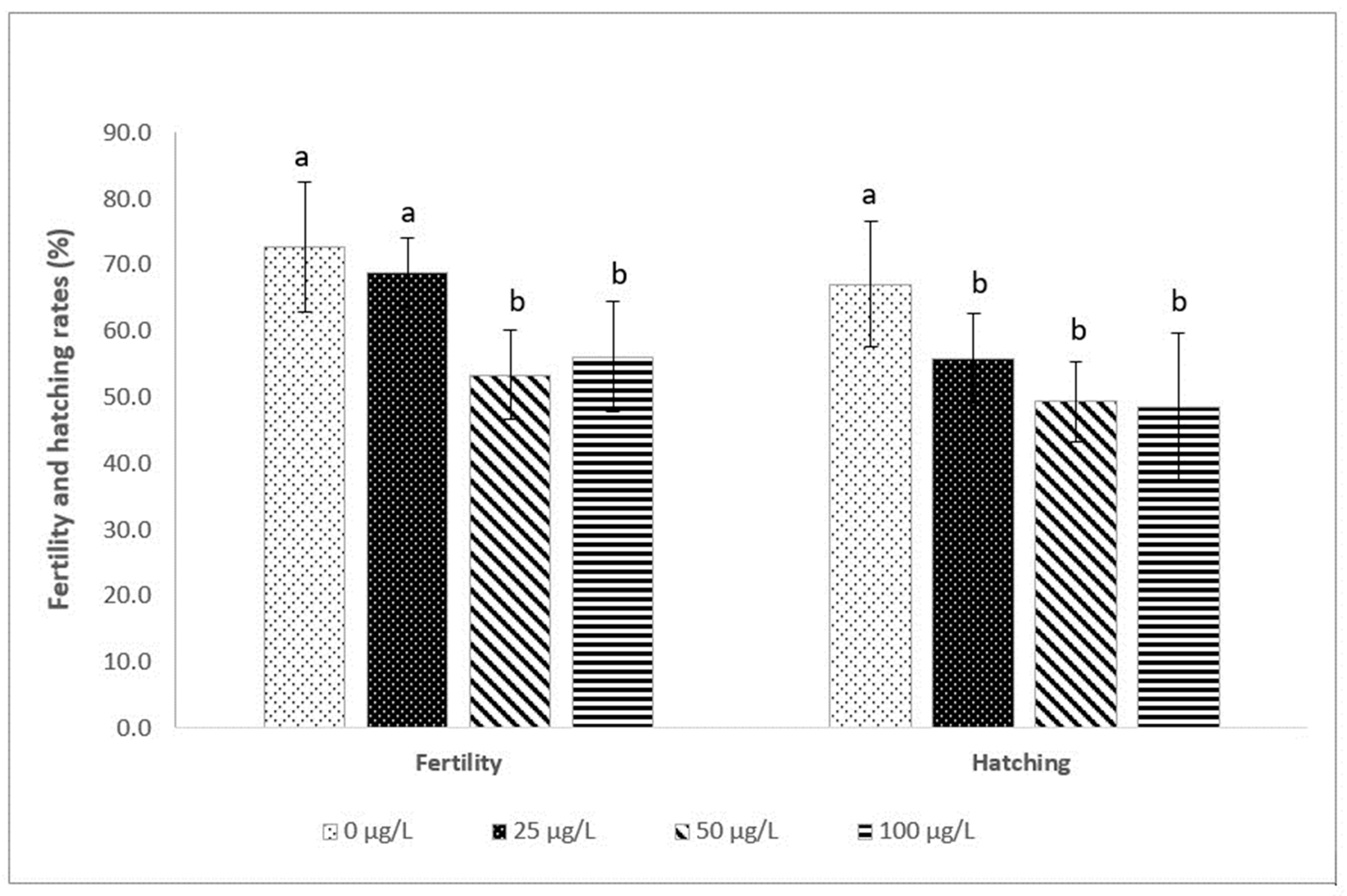
3.2. Fertilization and Hatching Rates
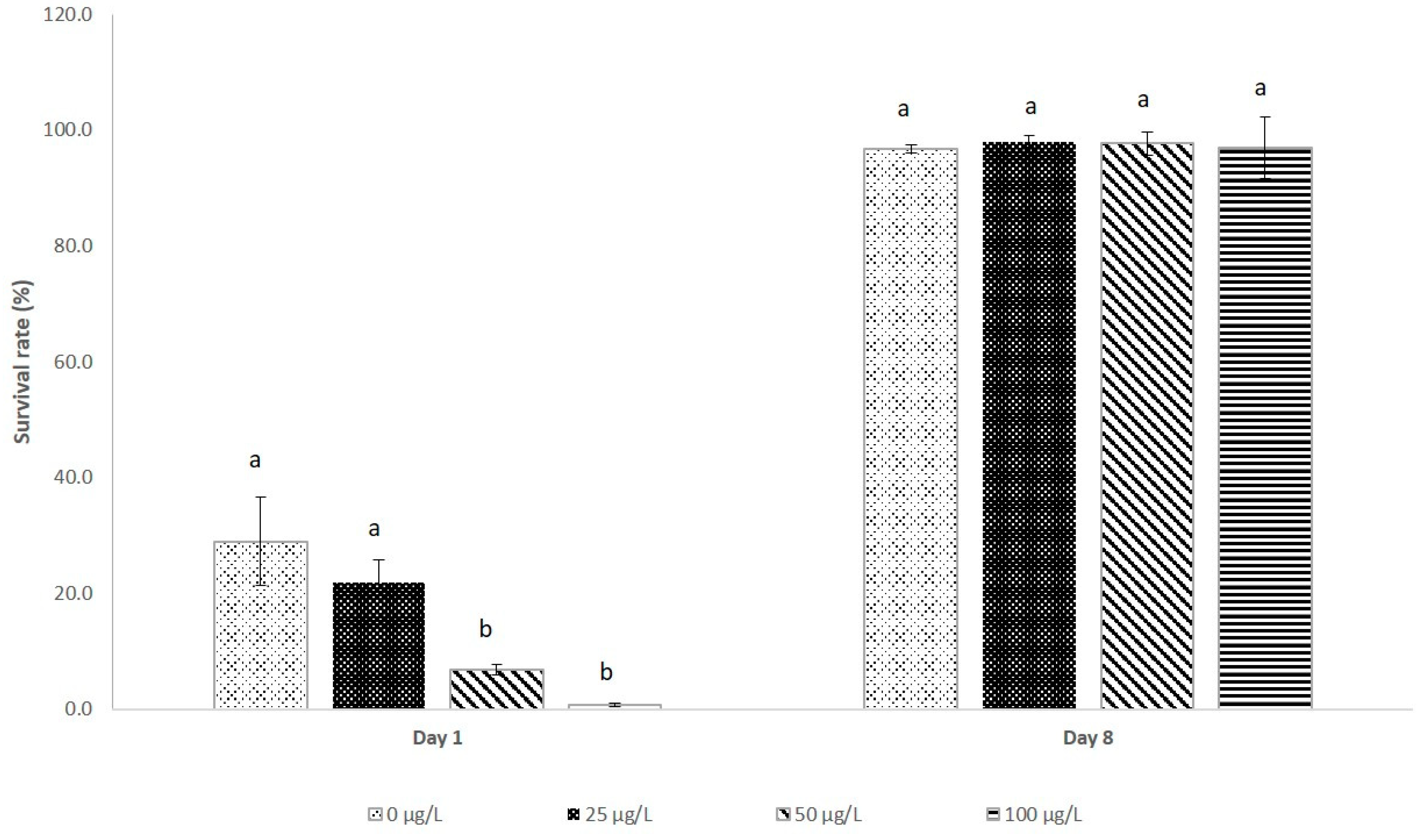
3.3. Larval Survival
3.4. Egg Diameters, Mercury Absorption by Eggs and Larvae, and Size of Newly Hatched Larvae
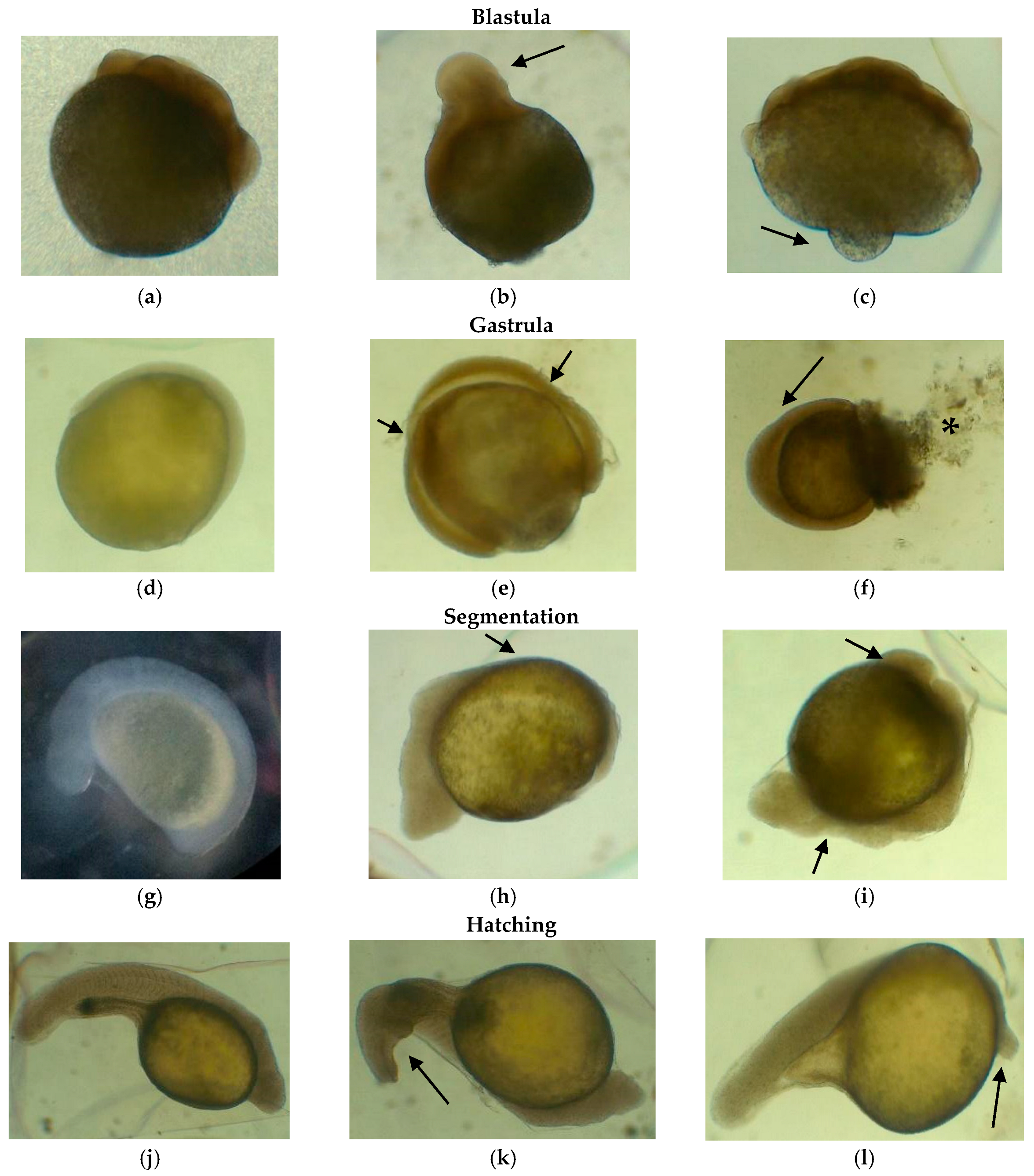
3.5. Deformation of Embryos and Larvae Exposed to Inorganic Mercury
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaioli, M.; Amoedo, D.; González, D. Impact of mercury on human health and the environment. Arch. Argent. Pediatr. 2012, 110, 259–264. [Google Scholar] [CrossRef]
- Casas, I.C.; Gómez, E.; Rodríguez, L.M.; Girón, S.L.; Mateus, J.C. Hacia un plan nacional para el control de los efectos en salud del mercurio en Colombia. Biomedica 2015, 35, 30–37. [Google Scholar] [CrossRef][Green Version]
- Bera, T.; Kumar, S.V.; Devi, M.S.; Kumar, V.; Behera, B.K.; Das, B.K. Effect of heavy metals in fish reproduction: A review. J. Environ. Biol. 2022, 43, 631–642. [Google Scholar] [CrossRef]
- Chen, Q.; An, J.; Xie, D.; Gong, S.; Lian, X.; Liu, Z.; Shen, Y.; Li, Y. Suppression and recovery of reproductive behavior induced by early life exposure to mercury in zebrafish. Comp. Biochem. Physiol. Pt. C Toxicol. Pharmacol. 2021, 239, 108876. [Google Scholar] [CrossRef] [PubMed]
- Brraich, O.S.; Jangu, S. Some aspects of reproductive biology on effect of heavy metal pollution on the histopathological structure of gonads in Labeo rohita (Hamilton-Buchanan) from Harike wetland, India. Int. J. Fish. Aquac. 2015, 7, 9–14. [Google Scholar] [CrossRef]
- Crump, K.L.; Trudeau, V.L. Mercury-induced reproductive impairment in fish. Environ. Toxicol. Chem. 2009, 28, 895–907. [Google Scholar] [CrossRef]
- Telmer, K.H.; Veiga, M.M. World emissions of mercury from artisanal and small scale gold mining. In Mercury Fate and Transport in the Global Atmosphere; Springer US: Boston, MA, USA, 2009; pp. 131–172. [Google Scholar]
- ANM (Agencia Nacional de Minería). Boletín Estadístico Minero: 2020–2021; ANM, Ministerio de Minas y Energía: Bogotá, Colombia, 2021. [Google Scholar]
- Cordy, P.; Veiga, M.; Salih, I.; Al-Saadi, S.; Console, S.; García, O.; Mesa, L.A.; Velásquez-López, P.C.; Roeser, M. Mercury contamination from artisanal gold mining in Antioquia, Colombia: The world’s highest per capita mercury pollution. Sci. Total Environ. 2011, 410–411, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.J.S. All that glitters is not gold or platinum: Institutions and the use of mercury in mining in Chocó, Colombia. Extr. Ind. Soc. 2018, 5, 308–318. [Google Scholar] [CrossRef]
- Londoño Franco, L.F.; Londoño Muñoz, P.T.; Muñoz García, F.G. Los riesgos de los metales pesados en la salud humana y animal. Biotecnol. Sect. Agropecu. Agroind. 2016, 14, 145–153. [Google Scholar] [CrossRef]
- Díaz Arriaga, F.A. Mercurio en la minería del oro: Impacto en las fuentes hídricas destinadas para consumo humano. Rev. Salud Pública 2015, 16, 947–957. [Google Scholar] [CrossRef]
- Enamorado-Montes, G.; Reino-Causil, B.; Urango-Cardenas, I.; Marrugo-Madrid, S.; Marrugo-Negrete, J. Mercury Accumulation in Commercial Varieties of Oryza sativa L. Cultivated in Soils of La Mojana Region, Colombia. Toxics 2021, 9, 304. [Google Scholar] [CrossRef]
- Gutiérrez, F.; de la Parra, A. Contaminación del agua en la cuenca del rio Magdalena (Colombia) y su relación con los peces. In Peces de la Cuenca del río Magdalena, Colombia: Diversidad, Conservación y Uso Sostenible; Jiménez-Segura, L., Lasso, C.A., Eds.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2020; pp. 239–269. [Google Scholar] [CrossRef]
- Atencio-García, V.J.; Madariaga-Mendoza, D.L.; Ortiz-Bedoya, A.; Guerrero-Durango, H.; Marrugo-Negrete, J.L. Fish health in the Cauca River (Colombia). In Proceedings of the IX International Symposium on Aquatic Animal Health: Enhancing Aquatic Animal Health towards One Health, Santiago, Chile, 5–8 September 2022. [Google Scholar]
- Moreno-Arias, C.; López-Casas, S.; Rogeliz-Prada, C.; Jiménez-Segura, L. Protection of spawning habitat for potamodromous fish, an urgent need for the hydropower planning in the Andes. Neotrop. Ichthyol. 2021, 19, e210027. [Google Scholar] [CrossRef]
- Marrugo-Negrete, J.; Navarro-Frómeta, A.; Ruiz-Guzmán, J. Total mercury concentrations in fish from Urrá reservoir (Sinú river, Colombia): Six years of monitoring. Rev. MVZ Córdoba 2015, 20, 4754–4765. [Google Scholar] [CrossRef][Green Version]
- Marrugo-Negrete, J.; Lans, E.; Benítez, L. Hallazgo de mercurio en peces de la Ciénaga de Ayapel, Córdoba, Colombia. Rev. MVZ Córdoba 2007, 12, 878–886. [Google Scholar] [CrossRef]
- USEPA (US Environmental Protection Agency). Updates 1995: Water Quality Criteria Documents for the Protection of Aquatic Life in Ambient Water; EPA-820-B-96-001; Office of Water: Washington, DC, USA, 1996. [Google Scholar]
- Porthro, M.G. Office of Water Policy and Technical Guidance on Interpretation and Implementation of Aquatic Life Metal Criteria. 1993. Available online: https://www.epa.gov/wqc/office-water-policy-and-technical-guidance-interpretation-and-implementation-aquatic-life-metals (accessed on 21 November 2022).
- Boyd, C.E. Water Quality, 2nd ed.; Springer: London, UK, 2015; Volume 2, pp. 277–311. [Google Scholar] [CrossRef]
- Kennedy, C.J. The toxicology of metals in fishes. In Encyclopedia of Fish Physiology: From Genome to Environment; Academic Press: San Diego, CA, USA, 2011. [Google Scholar] [CrossRef]
- Kotnik, J.; Horvat, M.; Begu, E.; Shlyapnikov, Y.; Sprovieri, F.; Pirrone, N. Dissolved gaseous mercury (DGM) in the Mediterranean Sea: Spatial and temporal trends. Mar. Chem. 2017, 193, 8–19. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, Z.; Žagar, D. Mercury transport and fate models in aquatic systems: A review and synthesis. Sci. Total Environ. 2018, 639, 538–549. [Google Scholar] [CrossRef]
- USEPA (US Environmental Protection Agency). Mercury Study Report to Congress; EPA-452/R-97-003; Office of Water: Washington, DC, USA, 1997; Volume I. [Google Scholar]
- Jezierska, B.; Ługowska, K.; Witeska, M. The effects of heavy metals on embryonic development of fish (a review). Fish Physiol. Biochem. 2009, 35, 625–649. [Google Scholar] [CrossRef]
- Drąg-Kozak, E.; Łuszczek-Trojnar, E.; Socha, M.; Bojarski, B. Effects of melatonin on cadmium accumulation and haematological parameters in cadmium intoxicated Prussian carp (Carassius gibelio B.). Ann. Anim. Sci. 2021, 21, 899–923. [Google Scholar] [CrossRef]
- Naz, S.; Hussain, R.; Ullah, Q.; Chatha, A.M.; Shaheen, A.; Khan, R.U. Toxic effect of some heavy metals on hematology and histopathology of major carp (Catla catla). Environ. Sci. Pollut. Res. 2021, 28, 6533–6539. [Google Scholar] [CrossRef]
- Suchana, S.A.; Ahmed, M.S.; Islam, S.M.; Rahman, M.L.; Rohani, M.F.; Ferdusi, T.; Ahmmad, A.K.; Fatema, M.K.; Badruzzaman, M.; Shahjahan, M. Chromium exposure causes structural aberrations of erythrocytes, gills, liver, kidney, and genetic damage in striped catfish Pangasianodon hypophthalmus. Biol. Trace Elem. Res. 2021, 199, 3869–3885. [Google Scholar] [CrossRef]
- Shahjahan, M.D.; Taslima, K.; Rahman, M.S.; Al-Emran, M.D.; Alam, S.I.; Faggio, C. Effects of heavy metals on fish physiology—A review. Chemosphere 2022, 300, 134519. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.M.; Rohani, M.F.; Zabed, S.A.; Islam, M.T.; Jannat, R.; Akter, Y.; Shahjahan, M. Acute effects of chromium on hemato-biochemical parameters and morphology of erythrocytes in striped catfish Pangasianodon hypophthalmus. Toxicol. Rep. 2020, 7, 664–670. [Google Scholar] [CrossRef]
- Hatef, A.; Alavix, S.M.H.; Golshan, M.; Linhart, O. Toxicity of environmental contaminants to fish spermatozoa function in vitro—A review. Aquat. Toxicol. 2013, 140-141, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, G.J.; Dietrich, M.; Kowalski, R.K.; Dobosz, S.; Karol, H.; Demianowicz, W.; Glogowski, J. Exposure of rainbow trout milt to mercury and cadmium alters sperm motility parameters and reproductive success. Aquat. Toxicol. 2010, 10, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Coward, K.; Bromage, N.K.; Hibbit, O.; Parrington, J. Gamete physiology, fertilization and activation in teleost fish. Rev. Fish Biol. Fish. 2002, 12, 33–58. [Google Scholar] [CrossRef]
- Rawson, D.W.; Zhang, T.; Kalicharan, D.; Jongebloed, W.L. Field emission scanning electron microscopy and transmission electron microscopy studies of the chorion, plasma membrane and syncytial layers of the gastrula-stage embryo of the zebrafish Brachydanio rerio: A consideration of the structural and functional relationships with respect to cryoprotectant penetration. Aquac. Res. 2000, 31, 325–336. [Google Scholar] [CrossRef]
- Jaramillo, R.; Goicoechea, O.; Garrido, O.; Molinari, E. Salmo salar: Morfología ultraestructural de la pared del corión en ovas normales y con problemas de eclosión. Arch. Med. Vet. 2009, 41, 67–71. [Google Scholar] [CrossRef]
- Villalobos, S.A.; Hamm, J.T.; Teh, S.J.; Hinton, D.E. Thiobencarb-induced embryotoxicity in medaka (Oryzias latipes): Stage-specific toxicity and the protective role of chorion. Aquat. Toxicol. 2000, 48, 309–326. [Google Scholar] [CrossRef]
- Atencio-García, V.J.; Arabia, R.F.; Aristizábal, R.J. Desarrollo embrionario y larvario de dorada Brycon sinuensis. In Memorias del IV Congreso Iberoamericano Virtual de Acuicultura CIVA; Universidad de Zaragoza: Zaragoza, Spain, 2007. [Google Scholar]
- Boglione, C.; Gisbert, E.; Gavaia, P.; Witten, P.; Moren, M.; Fontagn, S.; Koumoundouros, G. Skeletal anomalies in reared European fish larvae and juveniles. Part 2: Main typologies, occurrences and causative factors. Rev. Aquac. 2013, 5 (Suppl. S1), 121–167. [Google Scholar] [CrossRef]
- Sfakianakis, D.G.; Renieri, E.; Kentouri, M.; Tsatsakis, A.M. Effect of heavy metals on fish larvae deformities: A review. Environ. Res. 2015, 137, 246–255. [Google Scholar] [CrossRef]
- Favaloro, E.; Mazzola, A. Meristic variation and skeletal anomalies of wild and reared sharpsnout seabream juveniles (Diplodus puntazzo, Cetti 1777) off coastal Sicily, Mediterranean Sea. Aquac. Rev. 2003, 34, 575–579. [Google Scholar] [CrossRef]
- Yepes-Blandón, J.A.; Bian, C.; Benítez-Galeano, M.J.; Aristizabal-Regino, J.L.; Estrada-Posada, A.L.; Mir, D.; Vásquez-Machado, G.; Atencio-García, V.J.; Shi, Q.; Rodríguez-Osorio, N. Draft genome assembly for the Colombian freshwater bocachico fish, Prochilodus magdalenae. Front. Genet. 2023, 13, 989788. [Google Scholar] [CrossRef] [PubMed]
- Lasso, C.; Agudelo, E.; Jiménez-Segura, L.F.; Ramírez-Gil, H.; Morales-Betancourt, M.; Ajiaco-Martínez, R.; Gutiérrez, F.; Usma, J.; Muñoz, S.; Sanabria, A. Catalogo de Los Recursos Pesqueros Continentales; Serie Editorial Recursos Hidrobiológicos y Pesqueros Continentales de Colombia; Instituto de Investigacion de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2011. [Google Scholar]
- Duarte, L.; Cuervo, C.; Vargas, O.; Gil-Manrique, B.; Cuello, F.; De León, G.; Isaza, E.; Tejada, K.; Manjarrez-Martínez, L.; Reyes-Ardila, H. Estadísticas de Desembarco y Esfuerzo de las Pesquerías Artesanales de Colombia, Santa Marta, Colombia. 2020. Available online: http://sepec.aunap.gov.co/Archivos/Boletines-2020/SEPEC_Boletin_Pesca_Artesanal_2020.pdf (accessed on 10 October 2022).
- Mojica, J.; Castellanos, C.; Usma, J.; Álvarez, R.; Lasso, C. Libro Rojo de Peces Dulceacuícolas de Colombia; Serie Libros Rojos de Especies Amenazadas de Colombia; Universidad Nacional de Colombia, WWF: Manizales, Colombia, 2012. [Google Scholar]
- Atencio-García, V. Impactos de la Hidroeléctrica Urra en los peces migratorios del río Sinú. Rev. Temas Agrar. 2000, 5, 29–40. [Google Scholar]
- Ochoa-Orrego, L.; Jiménez-Segura, L.F.; Palacio, J. Ictioplancton en la ciénaga de Ayapel, Río San Jorge (Colombia): Cambios espacio-temporales. Bol. Cient. Mus. Hist. Nat. 2015, 19, 103–114. [Google Scholar] [CrossRef]
- Jiménez-Segura, L.F. Aspectos Diferenciales de las Comunidades de Peces en Grandes Ríos Tropicales y Sus Lagunas Marginales; Seminario de ecología de comunidades; Universidad Federal de Minas Gerais: Belo Horizonte, Brazil, 1998. [Google Scholar]
- Atencio-García, V.J.; Espinosa, J.A.; Martínez, J.G.; Pardo-Carrasco, S.C. Insemination of bocachico fish (Prochilodus magdalenae) with fresh or cryopreserved semen: Effect of spermatozoa/oocyte ratio. Rev. Colomb. Cienc. Pecu. 2015, 28, 347–355. [Google Scholar] [CrossRef]
- Atencio, V.; Kerguelén, E.; Naar, E.; Petro, R. Desempeño reproductivo del bocachico Prochilodus magdalenae inducido dos veces en un mismo año. Rev. MVZ Córdoba 2013, 18, 3304–3310. [Google Scholar] [CrossRef]
- Hayati, A.; Wulansari, E.; Armando, D.S.; Sofiyanti, A.; Amin, M.H.; Pramudy, M. Effects of in vitro exposure of mercury on sperm quality and fertility of tropical fish Cyprinus carpio L. Egypt. J. Aquat. Res. 2019, 45, 189–195. [Google Scholar] [CrossRef]
- Pataki, B.; Roberta, B.I.; Gazsi, G.; Urbányi, B.; Kollár, T.; Horváth, A. Effect of age on the mercury sensitivity of zebrafish (Danio rerio) sperm. Fish Physiol. Biochem. 2021, 47, 687–695. [Google Scholar] [CrossRef]
- Hatef, A.; Alavi, S.M.; Butts, A.E.; Policar, T.; Linhart, O. Mechanism of action of mercury on sperm morphology, adenosine triphosphate content, and motility in Perca fluviatilis (Percidae; Teleostei). Environ. Toxicol. Chem. 2011, 30, 905–914. [Google Scholar] [CrossRef]
- Xin, M.; Niksirat, H.; Shaliutina-Kolešová, A.; Siddique, M.; Sterba, J.; Boryshpolets, S.; Linhart, O. Molecular and subcellular cryoinjury of fish spermatozoa and approaches to improve cryopreservation. Rev. Aquac. 2020, 12, 909–924. [Google Scholar] [CrossRef]
- Cabrita, E.; Martínez-Páramo, S.; Gavaia, P.J.; Riesco, M.F.; Valcarce, D.G.; Sarasquete, C.; Herráez, M.P.; Robles, V. Factors enhancing fish sperm quality and emerging tools for sperm analysis. Aquaculture 2014, 432, 389–401. [Google Scholar] [CrossRef]
- Jaishankar, M.; Tseten, T.; Anbalagan, N.; Mathew, B.B.; Beeregowda, K.N. Toxicity mechanism and health effects of some heavy metals. Interdiscip. Toxicol. 2014, 7, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Weis, J.S. Reproductive, developmental, and neurobehavioral effects of methylmercury in fishes. J. Environ. Sci. Health Part C 2009, 27, 212–225. [Google Scholar] [CrossRef]
- Linhart, O.; Alavi, S.M.H.; Rodina, M.; Gela, D.; Cosson, J. Comparison of sperm velocity, motility and fertilizing ability between firstly and secondly activated spermatozoa of common carp (Cyprinus carpio). J. Appl. Ichthyol. 2008, 24, 386–392. [Google Scholar] [CrossRef]
- Martínez, C.S.; Escobar, A.G.; Torres, J.G.; Brum, D.S.; Santos, W.F.; Alonso, M.J.; Salaices, M.; Vassallo, D.V.; Peçanha, F.M.; Leivas, F.G.; et al. Chronic exposure to low doses of mercury impairs sperm quality and induces oxidative stress in rats. J. Toxicol. Environ. Health A 2014, 77, 143–154. [Google Scholar] [CrossRef]
- Pieterse, G.M. Histopathological Changes in the Testis of Oreochromis Mossambicus (Cichlidae) as a Biomarker of Heavy Metal Pollution. Ph.D. Thesis, Rand Afrikaans University, Johannesburg, South Africa, 2004. [Google Scholar]
- Monsalvo-Spencer, P.; Salinas-Zavala, C.A.; Reynoso-Granados, T. Morfología de la membrana coriónica de los huevos de Octopus bimaculoides y Octopus hubbsorum (Cephalopoda: Octopodidae). Hidrobiologica 2013, 23, 124–129. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter (Unit) | 0 µg/L | 25 µg/L | 50 µg/L | 100 µg/L |
|---|---|---|---|---|
| Duration of motility (s) | 35.4 ± 3.0 a | 32.4 ± 2.0 b | 31.0 ± 2.5 b | 30.2 ± 2.3 b |
| Total motility (%) | 94.8 ± 3.3 a | 82.3 ± 9.2 b | 85.5 ± 9.2 b | 82.2 ± 10.3 b |
| Rapid (%) | 39.2 ± 11.5 a | 20.3 ± 10.4 b | 24.3 ± 8.8 b | 24.5 ± 8.1 b |
| Medium (%) | 31.9 ± 6.5 a | 23.9 ± 4.9 b | 23.8 ± 4.1 b | 23.6 ± 4.1 b |
| Slow (%) | 23.8 ± 7.4 b | 38.1 ± 11.6 a | 37.5 ± 4.7 a | 35.5 ± 6.7 a |
| Immotile (%) | 5.2 ± 2.3 b | 17.7 ± 9.2 a | 14.5 ± 9.2 a | 17.8 ± 10.3 a |
| Total progressivity (%) | 57.3 ± 11.3 a | 32.9 ± 13.8 b | 35.1 ± 10.7 b | 36.1 ± 11.0 b |
| VCL (µm/s) | 92.7 ± 15.0 a | 68.4 ± 16.8 b | 74.0 ± 12.9 b | 75.9 ± 10.6 b |
| VSL (µm/s) | 46.5 ± 8.6 a | 37.8 ± 12.5 a | 39.4 ± 8.3 a | 44.8 ± 13.2 a |
| STR (%) | 62.2 ± 4.7 a | 64.2 ± 6.1 a | 62.5 ± 5.1 a | 63.3 ± 4.0 a |
| LIN (%) | 52.6 ± 4.7 a | 50.5 ± 9.0 a | 49.1 ± 6.2 a | 51.1 ± 6.0 a |
| WOB (%) | 82.0 ± 4.8 a | 73.6 ± 8.3 b | 73.8 ± 5.0 b | 75.3 ± 5.4 b |
| ALH (µm) | 1.6 ± 0.1 a | 1.3 ± 0.1 b | 1.4 ± 0.2 a | 1.4 ± 0.1 b |
| BCF (Hz) | 10.4 ± 0.6 a | 8.5 ± 1.4 b | 8.7 ± 1.2b | 8.9 ± 1.0 b |
| Parameters | 0 µg/L | 25 µg/L | 50 µg/L | 100 µg/L |
|---|---|---|---|---|
| Egg diameters (mm) | 4.3 ± 0.3 a | 4.5 ± 0.3 b | 4.9 ± 0.4 c | 4.9 ± 0.5 c |
| iHg absorption by eggs (µg/kg of eggs) | 1.1 ± 0.2 c | 64.5 ± 3.5 b | 447.6 ± 367.9 b | 1603.3 ± 766.1 a |
| tL of day-old larvae (mm) | 8.2 ± 1.2 a | 8.3 ± 1.2 a | 8.0 ± 0.9 a | 8.3 ± 0.6 a |
| iHg absorption by day-old larvae (µg/kg of larvae) | 115.4 ± 4.6 c | 150.7± 10.7 b | 192.35 ± 11.10 a | 214.8 ± 21.2 a |
| Hours (Stage) | 0 µg/L | 25 µg/L | 50 µg/L | 100 µg/L |
|---|---|---|---|---|
| 1 HPF (blastula) | 4.7 ± 1.2 c | 7.4 ± 2.8 b | 8.8 ± 4.0 ab | 17.6 ± 5.0 a |
| 5 HPF (gastrula) | 4.8 ± 3.3 c | 7.0 ± 2. 6 b c | 15.3 ± 6.2 a | 11.2 ± 8.8 ab |
| 8 HPF (segmentation) | 8.2 ± 2.9 b | 15.6 ± 5.2 a | 19.1 ± 5.3 a | 15.0 ± 1.9 a |
| 11 HPF (pharyngula) | 10.0 ± 7.7 b | 45.8 ± 9.1 a | 49.3 ± 16.0 a | 46.6 ± 9.2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madariaga-Mendoza, D.; Marrugo-Negrete, J.; Atencio-García, V. Effect of Inorganic Mercury on Semen Quality, Embryo and Larval Development of Bocachico Prochilodus magdalenae. Fishes 2023, 8, 445. https://doi.org/10.3390/fishes8090445
Madariaga-Mendoza D, Marrugo-Negrete J, Atencio-García V. Effect of Inorganic Mercury on Semen Quality, Embryo and Larval Development of Bocachico Prochilodus magdalenae. Fishes. 2023; 8(9):445. https://doi.org/10.3390/fishes8090445
Chicago/Turabian StyleMadariaga-Mendoza, Diana, José Marrugo-Negrete, and Víctor Atencio-García. 2023. "Effect of Inorganic Mercury on Semen Quality, Embryo and Larval Development of Bocachico Prochilodus magdalenae" Fishes 8, no. 9: 445. https://doi.org/10.3390/fishes8090445
APA StyleMadariaga-Mendoza, D., Marrugo-Negrete, J., & Atencio-García, V. (2023). Effect of Inorganic Mercury on Semen Quality, Embryo and Larval Development of Bocachico Prochilodus magdalenae. Fishes, 8(9), 445. https://doi.org/10.3390/fishes8090445